
Ekspresja antygenu Fas i FasL na limfocytach T i B w przerosłych migdałkach gardłowych u dzieci...
6
WSTĘP Podczas odpowiedzi immunologicznej anty- genowo-specyficzne limfocyty T i B ulegają intensywnej proliferacji rozwijając swoje funkcje efektorowi, jak wydzielanie przeciwciał, cytotok- syczność lub wytwarzanie cytokin. Aby ochro- nić własne tkanki przed uszkodzeniem przez te mechanizmy, po zakończeniu odpowiedzi immu- nologicznej efektorowe aktywowane limfocyty są 59 Otolaryngologia Polska 2008, LXII, 1 Autorzy nie zgłaszają konfliktu interesów. Ekspresja antygenu Fas i FasL na limfocytach T i B w przerosłych migdałkach gardłowych u dzieci chorych na wysiękowe zapalenie ucha Expression Fas and FasL antigen protein of lymphocytes T and B in hypertrophied adenoid in children with otitis media with effusion Beata Żelazowska-Rutkowska 1 , Jolanta Wysocka 1 , Karol Ratomski 1 , Bożena Skotnicka 2 , Janusz Żak 1 , Małgorzata Mrugacz 3 1 Zakład Laboratoryjnej Diagnostyki Pediatrycznej AM w Białymstoku Kierownik: prof. dr hab. J. Wysocka 2 Klinika Otolaryngologii Dziecięcej AM w Białymstoku Kierownik: prof. dr hab. E. Hassmann-Poznańska 3 Klinika Okulistyki Dziecięcej AM w Białymstoku Kierownik: prof. dr hab. A. Bakunowicz-Łazarczyk Summary Introduction. Apoptosis of lymphocytes can be induced by different factors mainly however the activation of antigen Fas (CD95) and his ligand FasL (CD95L) route. Apoptosis induced by Fas-FasL has large significance in elimination of lymphocytes T in final phase immune response designate AICD (activation induced cell death). Lymphocytes B undergo apoptosis induced by Fas-FasL in germinal center, which has special meaning in elimination of cells about low specificity and weak affinity of receptor BCR ( B-cell antigen receptor). Aim of study. The aim of this study was evaluation of per- centage of apoptotic lymphocytes and expression of Fas and FasL in CD4 + , CD8 + , CD19 + lymphocytes in hypertrophied adenoid and hypertrophied adenoid and otitis media with effusion. Methods. CD4 + Fas + , CD8 + Fas + , CD19 + Fas + CD4 + FasL + , CD8 + FasL + , CD19 + FasL + cells subpopulation were identified using monoclonal antibodies and flow cytometry method. Results. The percentage of lymphocytes CD4 + Fas + , CD8 + Fas + , CD19 + Fas + was higher in hypertrophied adenoid and otitis media with effusion compored to the control group. Their was no significant difference the percentage CD4 + Fas + , CD8 + Fas + , CD19 + Fas + CD4 + FasL + , CD8 + FasL + , CD19 + FasL + in hypertrophied adenoid and otitis media with effusion then the control group. The percentage of apoptotic lymphocytes was higher in hypertrophied adenoid and otitis media with effusion compored to the control group. Conclusions. The susceptibility the individual subpopulation of lymphocytes in hypertrophied adenoid seems to influence on change of relations of quantitative lymphocytes and upset the immunological function of tonsils which can influence on course otitis media with effusion at children. Hasła indeksowe: apoptoza, wysiękowe zapalenie ucha, cytometria przepływowa Key words: apoptosis, otitis media with effusion, flow cytometry Otolaryngol Pol 2008; LXII (1): 59–64 © 2008 by Polskie Towarzystwo Otorynolaryngologów – Chirurgów Głowy i Szyi
-
Upload
malgorzata -
Category
Documents
-
view
214 -
download
2
Transcript of Ekspresja antygenu Fas i FasL na limfocytach T i B w przerosłych migdałkach gardłowych u dzieci...

WSTĘP
Podczas odpowiedzi immunologicznej anty-genowo-specyficzne limfocyty T i B ulegają intensywnej proliferacji rozwijając swoje funkcje
efektorowi, jak wydzielanie przeciwciał, cytotok-syczność lub wytwarzanie cytokin. Aby ochro-nić własne tkanki przed uszkodzeniem przez te mechanizmy, po zakończeniu odpowiedzi immu-nologicznej efektorowe aktywowane limfocyty są
59Otolaryngologia Polska 2008, LXII, 1
Autorzy nie zgłaszają konfliktu interesów.
Ekspresja antygenu Fas i FasL na limfocytach T i B w przerosłych migdałkach gardłowych u dzieci
chorych na wysiękowe zapalenie uchaExpression Fas and FasL antigen protein of lymphocytes T and B
in hypertrophied adenoid in children with otitis media with effusion
Beata Żelazowska-Rutkowska1, Jolanta Wysocka1, Karol Ratomski1, Bożena Skotnicka2, Janusz Żak1, Małgorzata Mrugacz3
1Zakład Laboratoryjnej Diagnostyki Pediatrycznej AM w Białymstoku Kierownik: prof. dr hab. J. Wysocka
2Klinika Otolaryngologii Dziecięcej AM w BiałymstokuKierownik: prof. dr hab. E. Hassmann-Poznańska3Klinika Okulistyki Dziecięcej AM w BiałymstokuKierownik: prof. dr hab. A. Bakunowicz-Łazarczyk
SummaryIntroduction. Apoptosis of lymphocytes can be induced by different factors mainly however the activation of antigen Fas (CD95) and his ligand FasL (CD95L) route. Apoptosis induced by Fas-FasL has large significance in elimination of lymphocytes T in final phase immune response designate AICD (activation induced cell death). Lymphocytes B undergo apoptosis induced by Fas-FasL in germinal center, which has special meaning in elimination of cells about low specificity and weak affinity of receptor BCR ( B-cell antigen receptor). Aim of study. The aim of this study was evaluation of per-centage of apoptotic lymphocytes and expression of Fas and FasL in CD4+, CD8+, CD19+ lymphocytes in hypertrophied adenoid and hypertrophied adenoid and otitis media with effusion. Methods. CD4+ Fas+, CD8+Fas+, CD19+Fas+ CD4+ FasL+, CD8+FasL+, CD19+FasL+ cells subpopulation were identified using monoclonal antibodies and flow cytometry method. Results. The percentage of lymphocytes CD4+ Fas+, CD8+Fas+, CD19+Fas+ was higher in hypertrophied adenoid and otitis media with effusion compored to the control group. Their was no significant difference the percentage CD4+ Fas+, CD8+Fas+, CD19+Fas+ CD4+ FasL+, CD8+FasL+, CD19+FasL+ in hypertrophied adenoid and otitis media with effusion then the control group. The percentage of apoptotic lymphocytes was higher in hypertrophied adenoid and otitis media with effusion compored to the control group. Conclusions. The susceptibility the individual subpopulation of lymphocytes in hypertrophied adenoid seems to influence on change of relations of quantitative lymphocytes and upset the immunological function of tonsils which can influence on course otitis media with effusion at children.
Hasła indeksowe: apoptoza, wysiękowe zapalenie ucha, cytometria przepływowa
Key words: apoptosis, otitis media with effusion, flow cytometry
Otolaryngol Pol 2008; LXII (1): 59–64 © 2008 by Polskie Towarzystwo Otorynolaryngologów – Chirurgów Głowy i Szyi
�
�

B. Żelazowska-Rutkowska i inni
60 Otolaryngologia Polska 2008, LXII, 1
usuwane między innymi w procesie apoptozy [21, 27].
Apoptoza limfocytów może być indukowana przez wiele czynników, jednak główną rolę w tej indukcji pełni aktywacja receptora śmierci Fas (CD95) przez jego ligand (CD95L). Proces apoptozy zależny od CD95 zachodzi przy udziale dwóch róż-nych mechanizmów. W typie I droga aktywacji omija mitochondria, a w typie II kontrolowanym przez antyapoptotyczne białka należące do rodziny Bcl-2 dochodzi do aktywacji mitochondriów [1, 19].
Fas jest transmembranową glikoproteiną typu I (50kDa) występującą konstytutywnie na powierzch-ni limfocytów, należącą do rodziny receptorów TNF (tumor necrosis factor)/NGT (nerve growth factor). FasL typ II transmembranowego białka (40kDa) nie występuje na limfocytach spoczynkowych, ulega on ekspresji podczas aktywacji limfocytów T i B i komórek NK [6, 10].
Aktywowanie receptora Fas przez komplemen-tarną cząsteczkę Fas ligand rozpoczyna wewnątrz-komórkowy szlak przemian. Po związaniu z ligan-dem dochodzi do oligomeryzacji cząsteczek recep-tora, a następnie sygnał przekazywany jest przez ich wewnątrzcytoplazmatyczną część zwaną domeną śmierci DD (death domain). Powoduje to aktywację wielu białek adapterowych i powstania wielkoczą-steczkowego kompleksu określanego mianem DISC (death-inducing signaling complex). Służy on jako platforma inicjacji enzymatycznego mechanizmu apoptozy. Następnie dochodzi do wzajemnej prote-olitycznej aktywacji cząsteczkowej kaspazy 8, która uruchamia kolejne cząsteczki innych kaspaz inicju-jąc całą kaskadę kaspaz aż do aktywacji efektorowej kaspazy 3 [4, 11].
Apoptoza indukowana przez wiązanie Fas-FasL ma największe znaczenie w eliminacji limfocytów T w końcowej fazie odpowiedzi immunologicznej nazywanej „śmiercią komórki indukowaną aktywa-cją” (AICD – activation induced cell death [1, 10, 12].
Limfocyty B ulegają apoptozie indukowanej przez wiązanie Fas-FasL w ośrodkach rozmnażania, co ma szczególne znaczenie w eliminacji komórek o niskiej swoistości i słabym powinowactwie receptora BCR [3, 8, 18, 25].
Celem pracy była ocena nasilenia apoptozy limfo-cytów w przerosłych migdałkach gardłowych u dzie-ci chorych na wysiękowe zapalenie ucha trwające ponad trzy miesiące przez ocenę odsetka limfocytów apoptotycznych i limfocytów T (CD4+ i CD8+) i B(CD19+) z ekspresją antygenu Fas i FasL.
MATERIAŁ I METODY
Grupę badaną stanowiło 42 dzieci w wieku od 1 do 17 r.ż. z przerostem migdałka gardłowego cho-rych na wysiękowe zapalenie ucha trwające ponad 3 miesiące. Z powodu braku postępów w leczeniu dzieci te zakwalifikowano do zabiegu adenoidekto-mii. Grupę porównawczą stanowiło 40 dzieci obojga płci w wieku od 1 do 17 r.ż. z przerostem migdałka gardłowego, u których nie stwierdzono stanu zapal-nego uszu.
Kwalifikację chorych oraz adenoidektomię prze-prowadzono w Klinice Otolaryngologii Dziecięcej Akademii Medycznej w Białymstoku.
Migdałki gardłowe natychmiast po adenoidekto-mii poddano mechanicznemu rozdrobnieniu w celu uzyskania zawiesiny komórek, której gęstość oce-niano w analizatorze hematologicznym Sysmex XT 2000i. Do analizy użyto zawiesiny komórek o gęsto-ści 4-10 x103/ μl. Wyizolowane komórki jednojądrza-ste w formie zawiesiny w PBS poddano znakowaniu przeciwciałami monoklonalnymi. Przeprowadzono znakowanie antygenów na powierzchni komórki, dodając do 100 μl zawiesiny 10 μl przeciwciał monoklonalnych (anty-CD4-FITC, anty-CD8-FITC, anty-CD19-FITC, anty-CD95-PE, anty-CD95L-PE). Następnie przeprowadzono 15 min. inkubację w temp. pokojowej. Po inkubacji dodawano odczynni-ki lizujące erytrocyty i ponownie przeprowadzono inkubacje. Komórki płukano PBS zawierającym 0,1% azydku sodu i 1 % albuminy wołowej wirując przez 5 min. przy 2000 obr./min., a następnie poddawano analizie cytometrycznej.
Po dokładnym wymieszaniu, próbki oceniano na cytometrze przepływowym EPICS XL Coulter, wypo-sażonym w laser argonowy emitujący promieniowa-nie o długości fali 488 nm.
Oceniano również odsetek komórek apoptotycz-nych zestawem Apoptest.
Badane komórki płukano w PBS i zawieszano w buforze w stężeniu 105–106 komórek/ml. Następnie do 96 μl zawiesiny komórek dodawano 1 μl Anneksyny-V sprzężonej z fluoresceiną oraz 2,5 μl jodku propi-dyny (PI) i inkubowano przez 10 min w ciemności. Po inkubacji do próby dodawano 250 μl buforu i bezpośrednio mierzono fluorescencję komórek wyko-rzystując metodę cytometrii przepływowej.
We wczesnej fazie apoptozy na powierzchni komórki dochodzi do utraty asymetrii fosfolipidów błony komórkowej spowodowanej translokacją fosfa-tydyloseryny z wewnętrznej na zewnętrzną stronę błony komórkowej. Zjawisko to jest charakterystyczne

Ekspresja antygenu Fas i FasL
61Otolaryngologia Polska 2008, LXII, 1
dla procesu apoptozy, ale zachodzi również podczas nekrozy komórki. Różnicując dwie formy śmierci komórki można wykorzystać właściwości Anneksyny V, białka, które w obecności jonów Ca2+i Mg2+ wiąże się z fofadyloseryną i jednocześnie stosując jodek propidyny, który wnika do wnętrza komórki, weryfi-kując integralność błony komórkowej [5, 15].
Jednoczesne dodanie Anneksyny V oraz jodku propidyny pozwala różnicować populację komórek żywych (nie barwiącym się żadnym flurochromem) od komórek w początkowym stadium apoptozy (barwiących się jedynie Anneksyny V) oraz komó-rek chłonących jodek propidyny i wiążących się z Anneksyny V, czyli komórek martwiczych [2, 15].
Wyniki przeprowadzonych badań podano jako procent pozytywnych komórek w poszczególnej pró-bie badanej. Różnice dla p<0,05 uznano za znamien-ne statystycznie.
WYNIKI
Odsetek limfocytów apoptotycznych w grupie dzieci z przerostem migdałka gardłowego i wysię-kowym zapaleniem ucha (WZU) wynosił średnio 3,60 ± 1,13% i był istotnie statystycznie wyższy, p < 0,001 niż w grupie dzieci z przerostem migdał-ka gardłowego (PMG), który wynosił średnio 2,55 ± 0,72%. W podgrupie dzieci młodszych grupy badanej (WZU1) odsetek limfocytów apoptotycz-nych stanowił 3,35 ± 1,20% i był znamiennie statystycznie wyższy niż w analogicznej podgru-pie dzieci grupy porównawczej (PMG1 – 2,48 ± 0,71%), p<0,04. Wykazano różnice znamiennie statystyczną, p < 0,004 między grupą WZU2 (3,85 ± 0,73%) a podgrupą PMG2 (2,63 ± 0,74%).
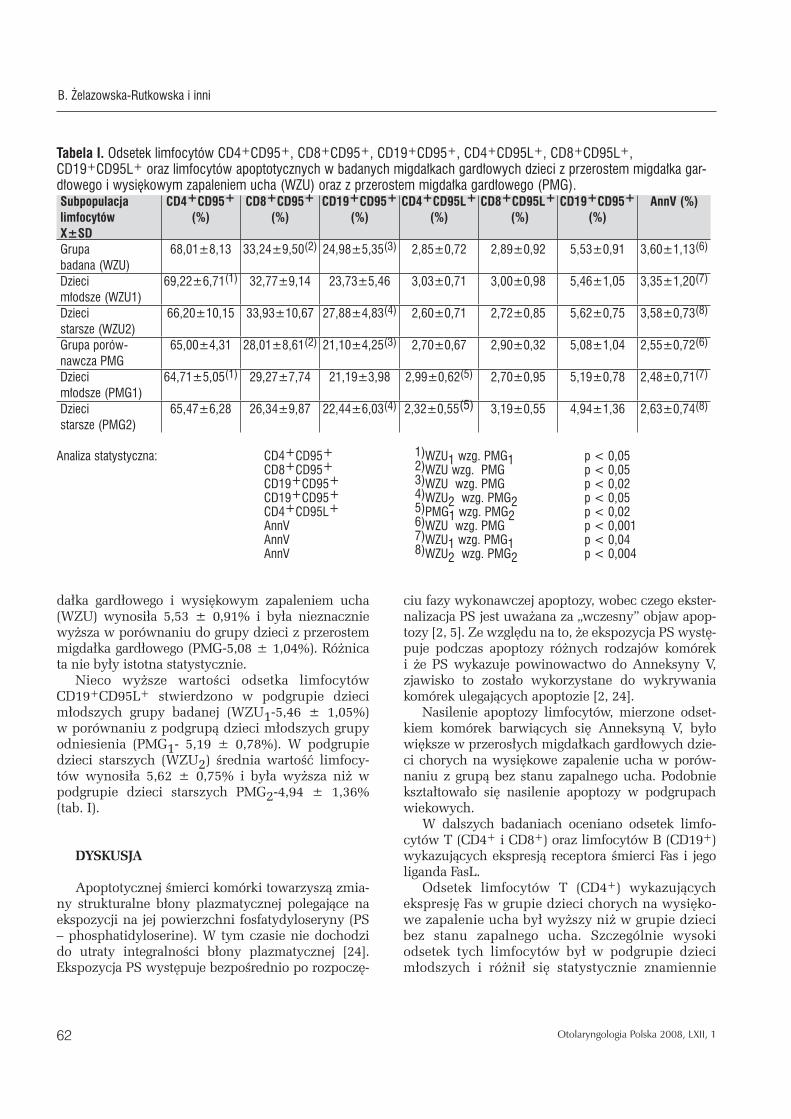
Odsetek limfocytów CD4+CD95+ u grupie dzieci z przerostem migdałka gardłowego i wysiękowym zapaleniem ucha (WZU) wynosił średnio 68,01 ± 8,13% i był nieznacznie wyższy w porównaniu z grupą dzieci z przerostem migdałka gardłowego (PMG), gdzie wynosił 65,00 ± 4,31%. W grupie badanej dzieci młodszych WZU1 stwierdzono sta-tystycznie znamiennie wyższy, p < 0,05 odsetek limfocytów CD4+CD95+, którego wartość wynosiła 69,22 ± 6,71% w porównaniu z podgrupą dzieci młodszych grupy porównawczej (PMG1-64,71 ± 5,05%). Nieznacznie wyższe wartości odsetka limfo-cytów CD4+CD95+ obserwowano w podgrupie dzie-ci starszych grupy badanej (WZU2-66,20 ± 10,15%), w porównaniu z podgrupą dzieci starszych grupy porównawczej (PMG2-65,47 ± 6,28%).
Odsetek limfocytów CD8+CD95+ w grupie dzieci z przerostem migdałka gardłowego i wysiękowym zapaleniem ucha (WZU) wynosił średnio 33,24 ± 9,50%. Był on znamiennie statystycznie wyższy, p < 0,05 względem grupy porównawczej (PMG-28,01 ± 8,61%). W podgrupach dzieci młodszych i dzieci starszych grupy badanej odsetek limfocytów CD8+CD95+ był nieznacznie wyższy (WZU1-32,77 ± 9,14%, WZU2-33,93 ± 10,67%) niż w analogicz-nych podgrupach grypy odniesienia (PMG1-29,27 ± 7,74%, PMG2-26,34 ± 9,87%), chociaż różnice te nie były znamienne statystyczne.
Analiza statystyczna wykazała statystycznie zna-miennie wyższy odsetek limfocytów CD19+CD95+, p<0,02 w grupie badanej (WZU-24,98 ± 5,35%) w porównaniu z grupą odniesienia (PMG-21,10 ± 4,25%). W podgrupie dzieci młodszych grupy bada-nej (WZU1) wartość ta wynosiła 23,73 ± 5,46% i była wyższa niż w podgrupie dzieci młodszych grupy porównawczej PMG1-21,19 ± 3,98%. W podgrupie dzieci starszych grupy badanej WZU2 odsetek limfo-cytów CD19+CD95+ wynosił średnio 27,88 ± 4,83% i był wyższy niż w podgrupie dzieci starszych grupy porównawczej PMG2-22,44 ± 6,03%. Wykazano róż-nicę istotną statystycznie, p < 0,05 pomiędzy grupą WZU2 a grupą PMG2.
Odsetek limfocytów CD4+CD95L+ w grupie dzie-ci z przerostem migdałka gardłowego i wysiękowym zapaleniem ucha (WZU) wynosił średnio 2,85 ± 0,72% i był nieznacznie wyższy niż w grupie dzie-ci z przerostem migdałka gardłowego (PMG), który wynosił średnio 2,70 ± 0,67%.W podgrupie dzieci młodszych i dzieci starszych grupy badanej odsetek limfocytów CD4+CD95L+ wynosił średnio (WZU1-3,03 ± 0,71%, WZU2-2,60 ± 0,71%) i był wyższy niż w analogicznych podgrupach wiekowych dzieci grupy odniesienia (PMG1-2,99 ± 0,62%, PMG2-2,32 ± 0,55%). Wykazano różnice znamiennie statystycz-ną, p < 0,02 między grupą PMG1 a PMG2.
Odsetek limfocytów CD8+CD95L+ w grupie bada-nej wynosił średnio (WZU 2,89 ± 0,92%) i był zbli-żony do grupy porównawczej (PMG-2,90 ± 0,32%). W podgrupie dzieci młodszych grupy badanej WZU1 odsetek limfocytów CD8+CD95L+wynosił 3,00 ± 0,98% i był nieco wyższy niż w grupie porównawczej dzieci młodszych PMG1-2,70 ± 0,95%. W podgrupie dzieci starszych grupy badanej WZU2 odsetek lim-focytów CD8+CD95L+ wynosił 2,72 ± 0,85% i był nieco niższy niż w podgrupie dzieci starszych grupy porównawczej PMG2 (3,19 ± 0,55%).
Wartość odsetka subpopulacji limfocytów CD19+CD95L+ w grupie dzieci z przerostem mig-
B. Żelazowska-Rutkowska i inni
62 Otolaryngologia Polska 2008, LXII, 1
dałka gardłowego i wysiękowym zapaleniem ucha (WZU) wynosiła 5,53 ± 0,91% i była nieznacznie wyższa w porównaniu do grupy dzieci z przerostem migdałka gardłowego (PMG-5,08 ± 1,04%). Różnica ta nie były istotna statystycznie.
Nieco wyższe wartości odsetka limfocytów CD19+CD95L+ stwierdzono w podgrupie dzieci młodszych grupy badanej (WZU1-5,46 ± 1,05%) w porównaniu z podgrupą dzieci młodszych grupy odniesienia (PMG1- 5,19 ± 0,78%). W podgrupie dzieci starszych (WZU2) średnia wartość limfocy-tów wynosiła 5,62 ± 0,75% i była wyższa niż w podgrupie dzieci starszych PMG2-4,94 ± 1,36% (tab. I).
DYSKUSJA
Apoptotycznej śmierci komórki towarzyszą zmia-ny strukturalne błony plazmatycznej polegające na ekspozycji na jej powierzchni fosfatydyloseryny (PS – phosphatidyloserine). W tym czasie nie dochodzi do utraty integralności błony plazmatycznej [24]. Ekspozycja PS występuje bezpośrednio po rozpoczę-
ciu fazy wykonawczej apoptozy, wobec czego ekster-nalizacja PS jest uważana za „wczesny” objaw apop-tozy [2, 5]. Ze względu na to, że ekspozycja PS wystę-puje podczas apoptozy różnych rodzajów komórek i że PS wykazuje powinowactwo do Anneksyny V, zjawisko to zostało wykorzystane do wykrywania komórek ulegających apoptozie [2, 24].
Nasilenie apoptozy limfocytów, mierzone odset-kiem komórek barwiących się Anneksyną V, było większe w przerosłych migdałkach gardłowych dzie-ci chorych na wysiękowe zapalenie ucha w porów-naniu z grupą bez stanu zapalnego ucha. Podobnie kształtowało się nasilenie apoptozy w podgrupach wiekowych.
W dalszych badaniach oceniano odsetek limfo-cytów T (CD4+ i CD8+) oraz limfocytów B (CD19+) wykazujących ekspresją receptora śmierci Fas i jego liganda FasL.
Odsetek limfocytów T (CD4+) wykazujących ekspresję Fas w grupie dzieci chorych na wysięko-we zapalenie ucha był wyższy niż w grupie dzieci bez stanu zapalnego ucha. Szczególnie wysoki odsetek tych limfocytów był w podgrupie dzieci młodszych i różnił się statystycznie znamiennie
Tabela I. Odsetek limfocytów CD4+CD95+, CD8+CD95+, CD19+CD95+, CD4+CD95L+, CD8+CD95L+, CD19+CD95L+ oraz limfocytów apoptotycznych w badanych migdałkach gardłowych dzieci z przerostem migdałka gar-dłowego i wysiękowym zapaleniem ucha (WZU) oraz z przerostem migdałka gardłowego (PMG).Subpopulacjalimfocytów X±SD
CD4+CD95+
(%)CD8+CD95+
(%)CD19+CD95+
(%)CD4+CD95L+
(%) CD8+CD95L+
(%)CD19+CD95+
(%)AnnV (%)
Grupa badana (WZU)
68,01±8,13 33,24±9,50(2) 24,98±5,35(3) 2,85±0,72 2,89±0,92 5,53±0,91 3,60±1,13(6)
Dzieci młodsze (WZU1)
69,22±6,71(1) 32,77±9,14 23,73±5,46 3,03±0,71 3,00±0,98 5,46±1,05 3,35±1,20(7)
Dzieci starsze (WZU2)
66,20±10,15 33,93±10,67 27,88±4,83(4) 2,60±0,71 2,72±0,85 5,62±0,75 3,58±0,73(8)
Grupa porów-nawcza PMG
65,00±4,31 28,01±8,61(2) 21,10±4,25(3) 2,70±0,67 2,90±0,32 5,08±1,04 2,55±0,72(6)
Dzieci młodsze (PMG1)
64,71±5,05(1) 29,27±7,74 21,19±3,98 2,99±0,62(5) 2,70±0,95 5,19±0,78 2,48±0,71(7)
Dzieci starsze (PMG2)
65,47±6,28 26,34±9,87 22,44±6,03(4) 2,32±0,55(5) 3,19±0,55 4,94±1,36 2,63±0,74(8)
Analiza statystyczna: CD4+CD95+ 1)WZU1 wzg. PMG1 p < 0,05 CD8+CD95+ 2)WZU wzg. PMG p < 0,05 CD19+CD95+ 3)WZU wzg. PMG p < 0,02 CD19+CD95+ 4)WZU 2 wzg. PMG2 p < 0,05 CD4+CD95L+ 5)PMG1 wzg. PMG2 p < 0,02 AnnV 6)WZU wzg. PMG p < 0,001 AnnV 7)WZU 1 wzg. PMG1 p < 0,04 AnnV 8)WZU 2 wzg. PMG2 p < 0,004

Ekspresja antygenu Fas i FasL
63Otolaryngologia Polska 2008, LXII, 1
w porównaniu z podgrupą dzieci młodszych bez stanu zapalnego ucha.
Również odsetek limfocytów TCD8+ z ekspresją Fas był statystycznie znamiennie wyższy w podgru-pie dzieci chorych na wysiękowe zapalenie ucha w porównaniu z grupą odniesienia.
Natomiast odsetek limfocytów T (CD4+ i CD8+) z ekspresją FasL był zbliżony w obu grupach badanych i podgrupach wiekowych dzieci.
Czynniki wpływające na żywotność limfocy-tów T zmieniają się w zależności od typu i stanu, w jakim znajdują się te komórki. Naiwne limfocyty T nie wykazują ekspresji receptora Fas i umierają zależnie od mechanizmu zależnego od Bcl-2/Bim [12]. Głównym mechanizmem, od którego zależy śmierć aktywowanych limfocytów T, są sygnały docierające do nich przez powierzchniowy recep-tor Fas [1, 10, 12].
Limfocyty T, wśród których przeważają lim-focyty CD4+, umiejscowione są głównie w prze-strzeni międzygrudkowej migdałków. Po kontakcie z komórkami prezentującymi antygen i interak-cji różnych cząstek kostymulujących dochodzi do pobudzenia i proliferacji limfocytów T [14]. Pobudzone limfocyty T wykazują ekspresję Fas [1], który po połączeniu z FasL inicjuje proces apopto-zy [1, 10, 12]. Proces ten zachodzi w końcowej fazie odpowiedzi immunologicznej.
Z badań immunohistochemicznych Stratera i wsp. [20] oraz Zakrzewskiej i wsp. [26] wynika, że w migdałkach większość komórek wykazują-cych ekspresję mRNA CD95L była zlokalizowana pod nabłonkiem krypt, a w nieco mniejszej ilości komórki takie występowały w przestrzeni mię-dzykomórkowej. Tateyama i wsp. [22] uważają, że ważnym czynnikiem regulującym są limfocyty T CD4+FasL+, gdyż limfocyty CD4+i CD8+ po kon-takcie z nimi ulegały apoptozie.
Wyższy odsetek limfocytów T CD4+ i CD8+ wykazujących ekspresję receptora śmierci Fas w przerosłych migdałkach gardłowych dzieci cho-rych na wysiękowe zapalenie ucha może świad-czyć o większym nasileniu apoptozy aktywowa-nych limfocytów, co z kolei może być przyczyną zaburzonej lokalnej odpowiedzi immunologicznej przerosłych migdałków gardłowych obejmującej ucho środkowe.
Odsetek limfocytów B z ekspresją Fas w gru-pie badanej był statystycznie znamiennie wyższy niż w grupie odniesienia. W podgrupach dzieci młodszych odsetek ten był zbliżony, natomiast w podgrupie dzieci starszych chorych na wysięko-
we zapalenie ucha był statystycznie znamiennie wyższy niż w podgrupie dzieci starszych bez stanu zapalnego ucha. Odsetek limfocytów B z ekspresją FasL był zbliżony w obu grupach badanych i pod-grupach wiekowych.
Nasilenie apoptozy limfocytów B ma znacze-nie w odpowiedzi humoralnej [21] migdałków. Decyzja pomiędzy śmiercią komórki a przeżyciem jest podejmowana już we wczesnym okresie roz-woju limfocytów B [7]. W ośrodkach rozmnażania apoptoza limfocytów B dotyczy komórek o niskiej swoistości i brakiem powinowactwa receptora BCR [18]. Limfocyty B w ośrodkach rozmnażania, szcze-gólnie proliferujące centroblasty, wykazują fenotyp charakterystyczny dla komórek wrażliwych na apoptozę, mianowicie niską zawartość antyapopto-tycznego Bcl-2 [13] i wysoką ekspresję cząsteczek indukujących śmierć takich jak Fas, Bax, c-Myc i p53 [7, 18]. Uważa się, że ponad 90% limfocytów B w ośrodkach rozmnażania umiera w wyniku apoptozy. Przeżywają jedynie te komórki, które syntetyzują przeciwciała o wysokim powinowa-ctwie i mogą się różnicować do komórek pamięci lub komórek plazmatycznych [18].
Znacznie wyższy odsetek limfocytów B z eks-presją Fas w przerosłych migdałkach u dzieci cho-rych na wysiękowe zapalenie ucha, zwłaszcza u dzieci po 5 r.ż., wydaje się świadczyć o większym nasileniu apoptozy tych limfocytów.
W migdałkach receptor Fas limfocytów B wiąże się z FasL występującym na limfocytach T i B [9] oraz na grudkowych komórkach dendrytycznych [8, 20], głównie w strefie jasnej, gdzie zachodzi selekcja centrocytów [25]. Chociaż komórki te peł-nią raczej funkcję antyapoptotyczną wzmagają pro-liferację i różnicowanie limfocytów B w ośrodkach rozmnażania [16]. Tsunoda i wsp. [23] uważają, że w migdałkowych ośrodkach rozmnażania grudko-we komórki dendrytyczne są zdolne do regulacji podatności limfocytów B na apoptozę. Wydaje się, że odsetek limfocytów B z ekspresją FasL nie wpły-wa na nasilenie apoptozy limfocytów B u dzieci chorych na wysiękowe zapalenie ucha, ponieważ jest on zbliżony w obu grupach badanych i podgru-pach wiekowych.
Prawidłowa funkcja immunologiczna migdał-ków gardłowych, które są źródłem komórek immu-nologicznie kompetentnych dla błon śluzowych górnych dróg oddechowych, w tym ucha środko-wego [17], zależy od wzajemnych stosunków iloś-ciowych i jakościowych komórek biorących udział w odpowiedzi immunologicznej.

B. Żelazowska-Rutkowska i inni
64 Otolaryngologia Polska 2008, LXII, 1
WNIOSKI
Jednym z ważnych procesów regulujących homeo-stazę migdałka gardłowego jest apoptoza. Podatność poszczególnych subpopulacji limfocytów w przero-słych migdałkach gardłowych wydaje się wpływać na zmianę stosunków ilościowych limfocytów, a tym samym zaburzać funkcję immunologiczną migdał-ków, co może wpływać na przebieg wysiękowego zapalenia ucha u dzieci.
PIŚMIENNICTWO
01. Baumann S, Krueger A, Kirchhoff S, Krammer PH. Regulation
of T cell apoptosis during the immune response. Curr Mol Med
2002; 2: 257–272.
02. Cornelissen M, Philippe J, De Sitter S, De Ridder L. Annexin
V expression in apoptotic peripheral blood lymphocytes: an
electron microscopic evaluation. Apoptosis 2002; 7: 41–47.
03. Daniel PT, Oettinger U, Mapara YM, Bommert K, Bargou R,
Dorken B. Activation and activation-induced death of human
tonsillar B cells and Burkitt lymphoma cells: lack of CD95 (Fas/
APO-1) ligand expression and function. Eur J Immunol 1997;
27: 1029–1034.
04. Fadeel B, Orrenius S. Apoptosis: a basic biological phenome-
non wide-ranging implications in human disease. J Intern Med
2005; 258: 479–517.
05. Gerbaulet A, Hartmann K, Mekori YA. Mast cell apoptosis.
Methods Mol Biol 2006; 315: 407–423.
06. Gupta S, Su H, Bi R, Agrawal S, Gollapudi S. Life and death
of lymphocytes: a role in immunesenescence. Immun Ageing
2005; 2: 12–26.
07. Guzman-Rojas L, Sims-Mourtada J, Rangel R, Martinez-Valdez
H. Life and death within germinal centres: a double – edged
sword. Immunology 2002; 107: 167–175.
08. Hur DY, Kim DJ, Kim S, Kim YY, Cho D, Lee DS, i wsp. Role of
follicular dendritic cells in the apoptosis in germinal center B
cells. Immunol Lett 2000; 72: 107–111.
09. Koopman G, Keehnen R M, Lindhout E, Zhou DF, de Groot
C, Pals ST. Germinal center B cells rescued from apoptosis by
CD40 ligation or attachment to follicular dendritic cells, but
not by engagement of surface immunoglobulin or adhesion
receptors, become resistant to CD95-induced apoptosis. Eur J
Immunol 1997; 27: 1–7.
10. Krueger A, Fas SC, Baumann S, Krammer PH. The role of CD95
in the regulation of peripheral T-cell apoptosis. Immunol Rev
2003; 193: 58–69.
11. Lavrik IN, Golks A, Krammer PH. Caspases: pharmacological
manipulation of cell death. J Clin Invest 2005; 115: 2665–
2672.
12. Marrack P, Kappler J. Control of T cell viability. Ann Rev
Immunol 2004, 22: 765–787.
13. Musiatowicz M, Hassmann-Poznańska E, Musiatowicz B,
Baltaziak M. The Bcl-2 protein expression in germinal cen-
ters of hypertrophied adenoids in children. Int J Pediatr
Otorhinolaryngol 2003; 67: 1369–1373.
14. Nave H, Gebert A, Pabst R. Morphology and immunology of
human paltine tonsil. Anat Embryol 2001; 204: 367–373.
15. Oancea M, Mazumder S, Crosby ME, Almasan A. Apoptosis
assays. Methods Mol Med 2006; 129: 279–290.
16. Park CS, Choi YS. How do follicular dendritic cells interact
intimately with B cells in germinal centre? Immunology 2005;
114: 2–10.
17. Rynnel-Dag B, gren K. The nasopharynx and the middle ear.
Inflammatory reactions in middle ear disease. Vaccine 2001;
19: S26–S31.
18. Sagaert X, De Wolf-Peeters C. Classification of B-cells according
to their differentiation status, their micro-anatomical localisa-
tion and their develpmental lineage. Immunol Lett 2003; 90:
179–186.
19. Scaffidi C, Schmitz I, Zha J, Korsmeyer SJ, Krammer PH, Peter
ME. Differential modulation of apoptosis sensitivity in CD95
type I and type II cells. J Biol Chem 1999; 374: 22532–22538.
20. Sräter J, Mariani SM, Walczak H, Rücker FG, Leithäuser F,
Krammer PH, i wsp. CD95 Ligand (CD95L) in normal human
lymphoid tissues. Am J Pathol 1999; 154: 193–201.
21. Strasser A, Bouilelet P. The control of apoptosis in lymphocyte
selection. Immunol Rev 2003; 193: 82–92.
22. Tateyama M, Oyaizu N, McCloskey TW, Than S, Pahwa S. CD4
lymphocytes are primed to express Fas ligand by CD4 cross-lin-
king and contribute to CD8 T-cell apoptosis via Fas/FasL death
pathway. Blood 2000; 96: 195–202.
23. Tsunoda R, Heinen E, Sugai N, Follicular dendritic cells in vitro
modulate the expression of Fas and Bcl-2 on germinal center B
cells. Cell Tissue Res 2000, 299: 395–402.
24. Van Engeland M, Nieland LJW, Ramaekers FCS, Schutte B,
Reutelingsperger CPM. AnnexinV-affinity assay: a reviewe on
an apoptosis detection system based on phosphatidylserine
exposure. Cytometry 1998; 31: 1–9.
25. Verbeke CS, Wenthe U, Zengraf H. Fas ligand expression in the
germinal centre. J Pathol 1999, 189: 155–160.
26. Zakrzewska A, Kobos J, Gryczyńska D. Evaluation of CD25,
CD152, Fas-ligand expression in the adenoids of aller-
gic and non-allergic children: a pilot study. Int J Pediatr
Otorhinolaryngol 2003; 67(suppl 1), S205–208.
27. Zhang N, Hartig H, Dzhagalov I, Draper D, He YW. The role of
apoptosis in the development and function of T lymphocytes.
Cell Res 2005; 15: 749–169.
Adres autora:
mgr Beata Żelazowska-Rutkowska
Zakład Laboratoryjnej Diagnostyki Pediatrycznej
AM w Białymstoku
ul. Waszyngtona 17
15-274 Białystok
tel: (085) 745 07 43


















