
Regulacja ekspresji genów w wektorach plazmidowych: …
16
Regulacja ekspresji genów w wektorach plazmidowych: system zale¿ny od doksycykliny i regulowany przez niedotlenienie S³awomir Go³da*, Paulina Kucharzewska*, Jaros³aw Cisowski, Urszula Florczyk, Anna Zagórska, Agnieszka JaŸwa, Agnieszka £oboda, Alicja Józkowicz, Józef Dulak Zak³ad Biotechnologii Medycznej, Wydzia³ Biochemii, Biofizyki i Bio- technologii, Uniwersytet Jagielloñski, Kraków Regulation of gene expression in plasmid vectors: doxocyclin-de- pendent and hypoxia-regulated systems Summary Regulation of gene expression in gene therapy is crucial for obtaining the therapeutic effects, thanks to limitation of transgene activity to the selected cells in a given time. In this paper we have focused on plasmid expression sys- tems regulated by doxycycline or hypoxia. We have described in details the structure, regulatory elements and biological applications of 1) the modified, commercially available Tet-On system, expressing doxycycline-controlled b-galactosidase and 2) hypoxia-activated FGF-4/VEGF expression plasmid con- taining the hypoxia responsive sequence. The presented expression systems can also be used in viral vectors, enabling not only regulated, but also high and long-term expression of transgenes. Key words: Tet-On system, Tet-Off system, HIF-1, hypoxia, regulable gene expression, gene therapy. 1. Wstêp Podstawow¹ cech¹, a zarazem ograniczeniem wiêkszoœci wektorów u¿ywanych w terapii genowej jest stosowanie promo- torów konstytutywnych, nie pozwalaj¹cych na regulacjê poziomu PRACE EKSPERYMENTALNE Adres do korespondencji Józef Dulak, Zak³ad Biotechnologii Medycznej, Wydzia³ Biochemii, Biofizyki i Biotechnologii, Uniwersytet Jagielloñski, ul. Gronostajowa 7, 31-271 Kraków; e-mail: [email protected] 3 (78) 82–97 2007 * Wk³ad tych osób do pracy by³ taki sam.
Transcript of Regulacja ekspresji genów w wektorach plazmidowych: …

Regulacja ekspresji genóww wektorach plazmidowych: systemzale¿ny od doksycykliny i regulowanyprzez niedotlenienie
S³awomir Go³da*, Paulina Kucharzewska*, Jaros³aw Cisowski,
Urszula Florczyk, Anna Zagórska, Agnieszka JaŸwa,
Agnieszka £oboda, Alicja Józkowicz, Józef Dulak
Zak³ad Biotechnologii Medycznej, Wydzia³ Biochemii, Biofizyki i Bio-
technologii, Uniwersytet Jagielloñski, Kraków
Regulation of gene expression in plasmid vectors: doxocyclin-de-
pendent and hypoxia-regulated systems
S u m m a r y
Regulation of gene expression in gene therapy is crucial for obtaining the
therapeutic effects, thanks to limitation of transgene activity to the selected
cells in a given time. In this paper we have focused on plasmid expression sys-
tems regulated by doxycycline or hypoxia. We have described in details the
structure, regulatory elements and biological applications of 1) the modified,
commercially available Tet-On system, expressing doxycycline-controlled
�-galactosidase and 2) hypoxia-activated FGF-4/VEGF expression plasmid con-
taining the hypoxia responsive sequence. The presented expression systems
can also be used in viral vectors, enabling not only regulated, but also high and
long-term expression of transgenes.
Key words:
Tet-On system, Tet-Off system, HIF-1, hypoxia, regulable gene expression,
gene therapy.
1. Wstêp
Podstawow¹ cech¹, a zarazem ograniczeniem wiêkszoœci
wektorów u¿ywanych w terapii genowej jest stosowanie promo-
torów konstytutywnych, nie pozwalaj¹cych na regulacjê poziomu
P R A C E E K S P E R Y M E N T A L N E
Adres do korespondencji
Józef Dulak,Zak³ad BiotechnologiiMedycznej,Wydzia³ Biochemii,Biofizyki i Biotechnologii,Uniwersytet Jagielloñski,ul. Gronostajowa 7,31-271 Kraków;e-mail:[email protected]
3 (78) 82–97 2007 * Wk³ad tych osób do pracy by³ taki sam.

transkrypcji transgenu. Obecnie coraz czêœciej siêga siê jednak po sekwencje pro-
motorowe, które mog¹ byæ aktywowane przez czynniki wewnêtrzne lub przez po-
dawane leki, umo¿liwiaj¹c regulacjê ekspresji transgenu i jego indukcjê tylko
w okreœlonych komórkach i sytuacjach patofizjologicznych.
1.1. Regulacja ekspresji genów w systemie Tet-Off i Tet-On
Regulowan¹ ekspresjê transgenu w wektorze plazmidowym zastosowano po raz
pierwszy u bakterii. Niektóre bia³ka uzyskiwane na skalê przemys³ow¹ w bakteriach
s¹ dla nich w du¿ych stê¿eniach toksyczne, zatem geny koduj¹ce po¿¹dane bia³ka
uruchamia siê dopiero, wówczas gdy hodowla bakteryjna jest ju¿ w fazie zaawanso-
wanego wzrostu. W jednym z efektywniejszych systemów tego typu wykorzystano
naturaln¹ zdolnoœæ niektórych bakterii do opornoœci na tetracyklinê, antybiotyk ha-
muj¹cy translacjê poprzez wi¹zanie siê do rybosomu. Wykazano, ¿e ulegaj¹cy kon-
stytutywnej ekspresji represor promotora dla bia³ka wypompowuj¹cego tetracykli-
nê z komórki jest inaktywowany po zwi¹zaniu z tetracyklin¹, co w konsekwencji
chroni komórki bakteryjne przed antybiotykiem. Tê zale¿noœæ wykorzystano do
konstrukcji bakteryjnego systemu regulowanej ekspresji transgenu, a nastêpnie prze-
niesiono j¹ na modele eukariotyczne.
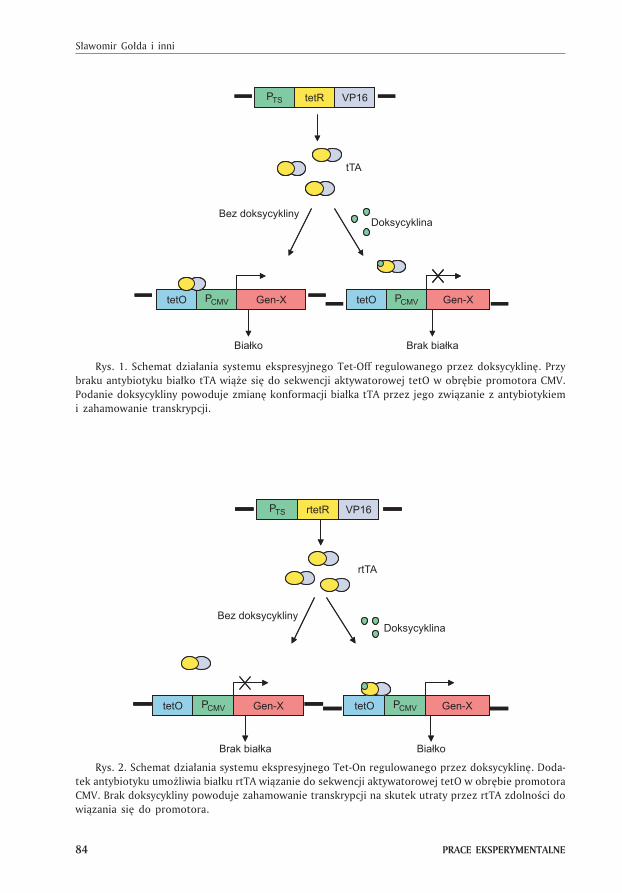
Istniej¹ dwa modele regulacji ekspresji genów przez antybiotyki z grupy tetracy-
klin: Tet-Off (1) i Tet-On (2). W systemie Tet-Off ekspresja genu jest hamowana tak
d³ugo dopóki antybiotyk nie zostanie usuniêty z po¿ywki. W systemie Tet-On do ak-
tywacji transgenu dochodzi po dodaniu doksycykliny (antybiotyku z grupy tetracy-
klin), a maksymalna ekspresja jak¹ udajê siê uzyskaæ w tym systemie jest porówny-
walna do nieregulowanych promotorów CMV (3).
W plazmidowym systemie Tet-Off istotne znaczenie ma bia³ko regulatorowe,
którego sekwencja kodowana jest przez bakteryjny gen TetR oraz fragment genomu
virusa HSV koduj¹cego aktywuj¹c¹ domenê VP16 (4). Fuzja domeny VP16 do TetR
przekszta³ca inhibitor w aktywator transkrypcji, a bia³ko powstaj¹ce na jego matry-
cy i nosz¹ce nazwê tTA (ang. tetracycline-controlled transactivator) indukuje ekspresjê
genu poprzez zwi¹zanie siê do operatora tet-O (rys. 1). Pojawienie siê w komórce
doksycykliny blokuje ekspresjê transgenu, gdy¿ antybiotyk wi¹¿¹c siê z bia³kiem
tTA zmienia jego konformacjê uniemo¿liwiaj¹c przy³¹czanie siê do promotora tetO
(rys. 1).
W systemie Tet-On do sekwencji TetR-VP16 doczepiony zosta³ fragment ko-
duj¹cy cztery aminokwasy (rTetR) (2,5), co w konsekwencji odwróci³o ca³kowicie
funkcjê bia³ka tTA zmieniaj¹c to bia³ko w aktywator rtTA (ang. reverse TA). Przy bra-
ku doksycykliny bia³ko to nie mo¿e przy³¹czyæ siê do promotora tetO i nie dochodzi
wówczas do ekspresji transgenu. Dopiero dodanie antybiotyku zmienia konforma-
cjê rtTA, umo¿liwiaj¹c aktywacjê promotora tetO i w konsekwencji indukcjê ekspre-
sji genu. Typowy system Tet-On przedstawiono na rysunku 2.
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 83
S³awomir Go³da i inni
84 PRACE EKSPERYMENTALNE
Rys. 1. Schemat dzia³ania systemu ekspresyjnego Tet-Off regulowanego przez doksycyklinê. Przy
braku antybiotyku bia³ko tTA wi¹¿e siê do sekwencji aktywatorowej tetO w obrêbie promotora CMV.
Podanie doksycykliny powoduje zmianê konformacji bia³ka tTA przez jego zwi¹zanie z antybiotykiem
i zahamowanie transkrypcji.
Rys. 2. Schemat dzia³ania systemu ekspresyjnego Tet-On regulowanego przez doksycyklinê. Doda-
tek antybiotyku umo¿liwia bia³ku rtTA wi¹zanie do sekwencji aktywatorowej tetO w obrêbie promotora
CMV. Brak doksycykliny powoduje zahamowanie transkrypcji na skutek utraty przez rtTA zdolnoœci do
wi¹zania siê do promotora.

W praktyce, aby ograniczyæ tzw. przeciekanie plazmidu (czyli samoistn¹, niekon-
trolowan¹ ekspresjê transgenu), do transfekcji komórek u¿ywa siê czêsto zmodyfi-
kowanego systemu Tet-On. W uk³adzie takim zmniejszenie niekontrolowanej eks-
presji osi¹gane jest poprzez wprowadzenie dodatkowego represora, tzw. tetS. Sys-
tem ten zosta³ wykorzystany w obecnej pracy, a schemat jego dzia³ania jest omó-
wiony w rozdz. 2.
1.2. Regulacja ekspresji genów przez hipoksjê
Hipoksja (niedotlenienie) jest jednym z istotnych bodŸców reguluj¹cych ekspres-
jê genów zaanga¿owanych w angiogenezê, czyli powstawanie nowych naczyñ krwio-
noœnych. Stê¿enie tlenu w tkankach ulega nieustannym wahaniom w zale¿noœci od
stanu i kondycji organizmu. Przy prawid³owym ciœnieniu parcjalnym tlenu (w nor-
moksji) system hydroksylaz prolinowych hamuje ekspresjê wybranych grup genów
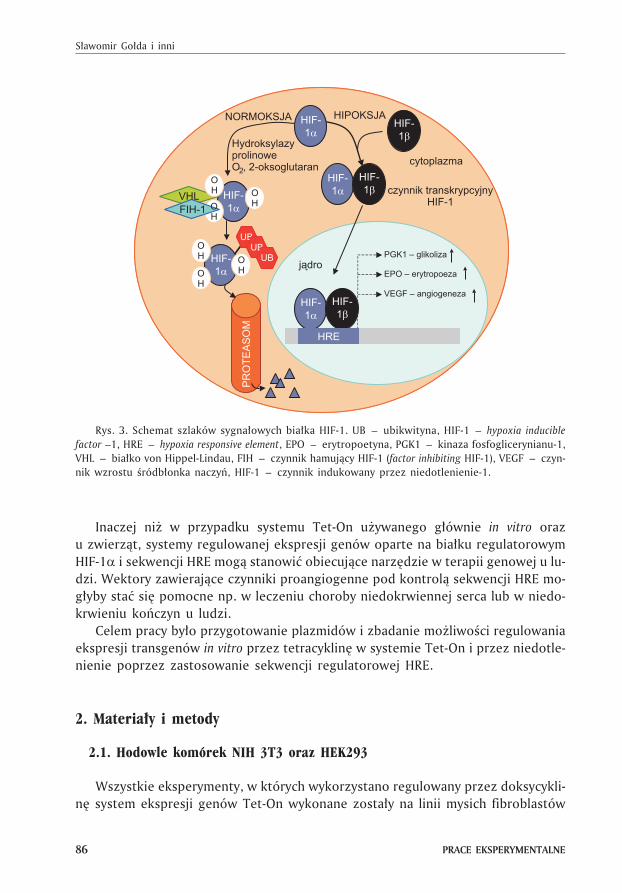
poprzez inaktywacjê bia³ka HIF-1 (zob. (6,7)). HIF-1 (ang. hypoxia inducible factor-1)
jest czynnikiem transkrypcyjnym odpowiedzialnym za utrzymanie równowagi ener-
getycznej zarówno na poziomie komórkowym jak i systemowym. Do procesów pod-
legaj¹cych kontroli bia³ka HIF-1 nale¿¹ m. in. glikoliza, erytropoeza i angiogeneza
(8).
Bia³ko HIF-1 to heterodimer zbudowany z podjednostki HIF-1� i HIF-1� zdol-
nych do wi¹zania DNA (9). Oba bia³ka wytwarzane s¹ konstytutywnie, ale HIF-1� jest
podjednostk¹ stabiln¹, natomiast HIF-1� jest form¹ o czasie pó³trwania krótszym
ni¿ 5 minut w warunkach normoksji. Za rozpad podjednostki HIF-1� odpowiedzial-
na jest ubikwitynacja prowadz¹ca do degradacji bia³ka w proteasomie (10) (rys. 3).
U podstaw tego procesu le¿y posttranslacyjna modyfikacja tej podjednostki po-
przez zale¿n¹ od tlenu hydroksylacjê reszt proliny w pozycjach 402 i 564 przez hy-
droksylazy prolinowe (11). Taka hydroksylacja stanowi sygna³ do ubikwitynacji, kie-
ruj¹c podjednostkê HIF-1� do degradacji z udzia³em bia³ka VHL (von Hippel-Lindau)
bêd¹cego fragmentem rozpoznawczym ligazy wi¹¿¹cej ubikwitynê E3 z bia³kiem
przeznaczonym do degradacji (12,13).
W warunkach hipoksji zarówno hydroksylacja proliny jak i kierowana przez do-
menê VHL degradacja HIF-1� s¹ ma³o wydajne, co w konsekwencji prowadzi do
wzrostu stê¿enia HIF-1� w komórce. Stabilna podjednostka HIF-1� dimeryzuje
z HIF-1� i razem tworz¹ kompleks transkrypcyjny wi¹¿¹cy siê do genomowej se-
kwencji HRE (ang. hypoxia responsive element) w obrêbie promotora genu regulowa-
nego przez hipoksjê (rys. 3).
Poœród wielu genów indukowanych przez HIF-1 mo¿na wyró¿niæ przede
wszystkim geny zaanga¿owane w angiogenezê takie jak czynnik wzrostu
œródb³onka naczyñ (VEGF, ang. vascular endothelial growth factor), receptor VEGFR-1
(flt-1), neuropilinê-1, angiopoetyny oraz indukowaln¹ syntazê tlenku azotu (iNOS)
(zob. (6,14)).
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 85
Inaczej ni¿ w przypadku systemu Tet-On u¿ywanego g³ównie in vitro oraz
u zwierz¹t, systemy regulowanej ekspresji genów oparte na bia³ku regulatorowym
HIF-1� i sekwencji HRE mog¹ stanowiæ obiecuj¹ce narzêdzie w terapii genowej u lu-
dzi. Wektory zawieraj¹ce czynniki proangiogenne pod kontrol¹ sekwencji HRE mo-
g³yby staæ siê pomocne np. w leczeniu choroby niedokrwiennej serca lub w niedo-
krwieniu koñczyn u ludzi.
Celem pracy by³o przygotowanie plazmidów i zbadanie mo¿liwoœci regulowania
ekspresji transgenów in vitro przez tetracyklinê w systemie Tet-On i przez niedotle-
nienie poprzez zastosowanie sekwencji regulatorowej HRE.
2. Materia³y i metody
2.1. Hodowle komórek NIH 3T3 oraz HEK293
Wszystkie eksperymenty, w których wykorzystano regulowany przez doksycykli-
nê system ekspresji genów Tet-On wykonane zosta³y na linii mysich fibroblastów
S³awomir Go³da i inni
86 PRACE EKSPERYMENTALNE
Rys. 3. Schemat szlaków sygna³owych bia³ka HIF-1. UB – ubikwityna, HIF-1 – hypoxia inducible
factor –1, HRE – hypoxia responsive element, EPO – erytropoetyna, PGK1 – kinaza fosfoglicerynianu-1,
VHL – bia³ko von Hippel-Lindau, FIH – czynnik hamuj¹cy HIF-1 (factor inhibiting HIF-1), VEGF – czyn-
nik wzrostu œródb³onka naczyñ, HIF-1 – czynnik indukowany przez niedotlenienie-1.

NIH 3T3 zakupionych w American Type Tissue Culture Collection (Manassas, VA, USA).
Komórki te hodowano w po¿ywce DMEM HG (4,5 g/L glukozy; Gibco) wzbogaconej
w 10% surowicê wo³ow¹ oraz penicylinê (100 U/mL) i streptomycynê (10 �g/mL).
Do badania poziomu ekspresji genów FGF-4 oraz VEGF-A pochodz¹cej z regulo-
wanego przez niedotlenienie plazmidu pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF wy-
korzystano liniê komórkow¹ HEK-293 (ang. human embryonic kidney epithelial cell line,
Stratagene), dostarczon¹ przez Stefana Kochanka (Ulm, Niemcy). Komórki te hodo-
wano podobnie jak NIH 3T3.
2.2. Konstrukcja wektora pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF zawie-raj¹cego geny FGF4 i VEGF indukowane przez hipoksjê
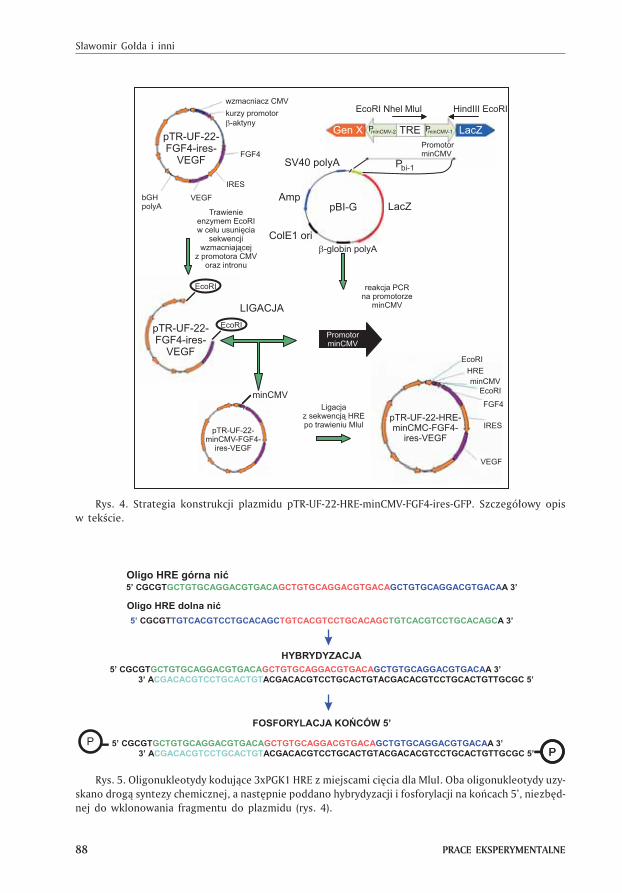
Strategia konstrukcji plazmidu pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF zosta³a
przedstawiona na rysunku 4. Plazmid pTR-UF-22-FGF4-ires-VEGF, którego dok³adn¹
konstrukcjê opisano w pracy (15), zosta³ poddany trawieniu endonukleaz¹ EcoRI
(Fermentas) w celu usuniêcia wstawki (zawieraj¹cej fragment CMV, kurzy promotor
�-aktyny oraz intron), a nastêpnie oczyszczony przy u¿yciu zestawu Gel-Out (A&A
Biotechnology), po rozdziale w 1% ¿elu agarozowym. Przeciêty plazmid zosta³ pod-
dany defosforylacji przy u¿yciu fosfatazy (Antarctic Phosphatase, New England Bio-
labs) w celu unikniêcia recyrkularyzacji. Minimalny promotor CMV (minCMV) zosta³
powielony z plazmidu pBI (Clontech) przy u¿yciu reakcji PCR (temperatura ³¹czenia
starterów 54°C, 30 cykli) z zastosowaniem starterów nCMV1_F oraz CMV1_R (tab.).
T a b e l a
Para starterów u¿ytych do klonowania promotora minCMV
Starter Sekwencja
nCMV1_F 5’ TATGAATTCGCTAGCACGCGTGTCGAGTAGGCGTGTACG 3’
CMV1_R 5’ ACGGAATTCAAGCTTGTACCGAGCTCGGATTCGGG 3’
EcoRI NhI MluI Hind III
Produkt reakcji PCR rozdzielono w 2% ¿elu agarozowym, wyizolowano zesta-
wem Gel-Out (A&A Biotechnology), a nastêpnie poddano ligacji (ligaz¹ T4; New En-
gland Biolabs) z przeciêtym uprzednio plazmidem pTR-UF-22-FGF4-ires-VEGF. Mie-
szanin¹ ligacyjn¹ transformowano kompetentne komórki E. coli (X-Gold; Stratagene)
metod¹ z zastosowaniem szoku termicznego. Po wyizolowaniu plazmidu z transfor-
mantów potwierdzono zarówno obecnoœæ jak i prawid³ow¹ orientacjê promotora
minCMV reakcj¹ PCR. W kolejnym etapie plazmid pTR-UF-22-minCMV-FGF4-ires-VEGF
zosta³ poddany trawieniu enzymem MluI (Promega), a nastêpnie ponownie rozdzie-
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 87
S³awomir Go³da i inni
88 PRACE EKSPERYMENTALNE
Rys. 4. Strategia konstrukcji plazmidu pTR-UF-22-HRE-minCMV-FGF4-ires-GFP. Szczegó³owy opis
w tekœcie.
Rys. 5. Oligonukleotydy koduj¹ce 3xPGK1 HRE z miejscami ciêcia dla MluI. Oba oligonukleotydy uzy-
skano drog¹ syntezy chemicznej, a nastêpnie poddano hybrydyzacji i fosforylacji na koñcach 5’, niezbêd-
nej do wklonowania fragmentu do plazmidu (rys. 4).

lony w 1% ¿elu agarozowym, wyciêty i oczyszczony zestawem Gel-Out (A&A Biotech-
nology) oraz poddany defosforylacji. Do dalszego klonowania u¿yto dwóch oligonu-
kleotydów koduj¹cych sekwencjê 3xPGK1 HRE, sklonowan¹ z promotora genu kina-
zy fosfoglicerynowej – PGK1 (16) i zawieraj¹cych na obydwu koñcach miejsce re-
strykcyjne dla enzymu MluI (rys. 5). Obydwa nukleotydy wymieszano ze sob¹ i pod-
dano denaturacji w 95°C przez 10 minut, a nastêpnie sch³odzono do 50°C w tempie
1°C/min. Po osi¹gniêciu 50°C obni¿ono temperaturê skokowo do 4°C.
Tak przygotowan¹ sekwencjê HRE poddano fosforylacji z u¿yciem kinazy polinu-
kleotydowej T4 (New England Biolabs), a nastêpnie wklonowano do uprzednio stra-
wionego MluI plazmidu pTR-UF-22-minCMV-FGF4-ires-VEGF. ObecnoϾ wstawki
HRE, po wczeœniejszej amplifikacji plazmidu w komórkach bakterii X-Gold, potwier-
dzona zosta³a analiz¹ restrykcyjn¹ oraz sekwencjonowaniem.
2.3. Przejœciowa transfekcja komórek HEK293 wektorami plazmidowymi(system regulowany przez niedotlenienie)
Dzieñ przed transfekcj¹ komórki wysiewano na szalki 24-do³kowe, tak aby na-
stêpnego dnia uzyskaæ konfluencjê 50-70%. Transfekcje wykonywano za pomoc¹ oli-
godendrymeru SuperFect (Qiagen) zgodnie z zaleceniami producenta. W tym celu
w probówce przygotowywano mieszaninê transfekcyjn¹ sk³adaj¹c¹ siê z: 1 �g pla-
zmidowego DNA, 60 �L po¿ywki wolnej od surowicy i antybiotyków oraz 5 �l od-
czynnika SuperFect. Po 10 minutach inkubacji w temperaturze pokojowej do mie-
szaniny dodawano 350 �L pe³nej po¿ywki i podawano na komórki (410 �L/do³ek).
Po up³ywie 6 godzin usuwano mieszaninê transfekcyjn¹ i po przep³ukaniu komórek
PBS dodawano 0,5 mL œwie¿ej po¿ywki. Komórki hodowano przez nastêpne 48 h.
3. Stymulacja HIF-1 w komórkach HEK293 – inkubacja z DMOG
W celu dokonania indukcji czynnika transkrypcyjnego HIF-1 24 h po transfekcji
plazmidami pTR-UF-22-minCMVFGF4-ires-VEGF i pTR-UF-22-HRE-minCMV-FGF4-ires-
-VEGF komórki by³y poddawane dzia³aniu dimetylooksoalliloglicyny, (DMOG 500 �M;
Alexis Biochemicals), bêd¹cej czynnikiem imituj¹cym hipoksjê poprzez hamowanie
aktywnoœci hydroksylaz prolinowych (17). Po 24 h od dodania DMOG zbierano po-
¿ywki, w których oznaczano poziom FGF-4 oraz VEGF-A przy zastosowaniu testu
ELISA (R&D).
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 89
4. Stabilna transfekcja komórek NIH 3T3 – zmodyfikowany systemTet-On regulowany przez doksycyklinê
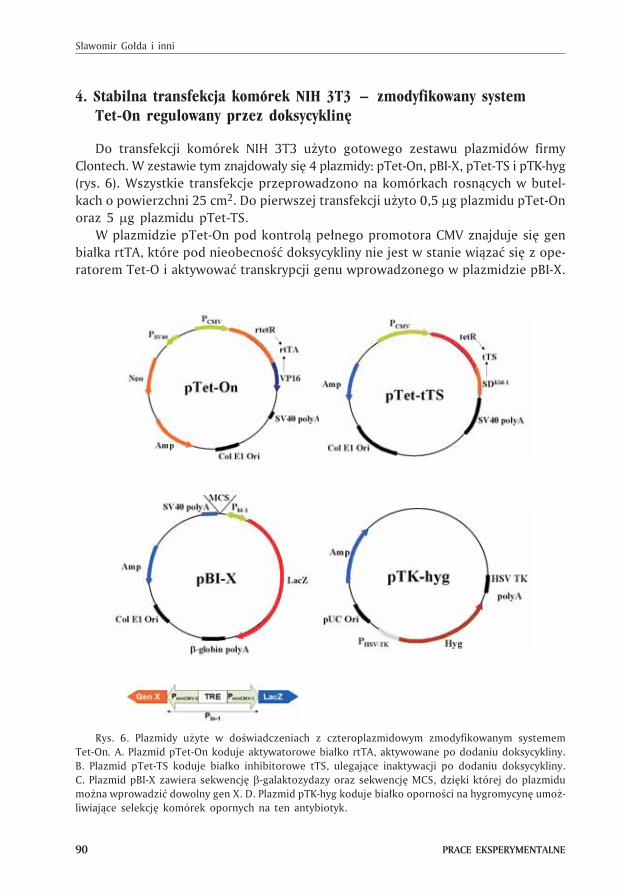
Do transfekcji komórek NIH 3T3 u¿yto gotowego zestawu plazmidów firmy
Clontech. W zestawie tym znajdowa³y siê 4 plazmidy: pTet-On, pBI-X, pTet-TS i pTK-hyg
(rys. 6). Wszystkie transfekcje przeprowadzono na komórkach rosn¹cych w butel-
kach o powierzchni 25 cm2. Do pierwszej transfekcji u¿yto 0,5 �g plazmidu pTet-On
oraz 5 �g plazmidu pTet-TS.
W plazmidzie pTet-On pod kontrol¹ pe³nego promotora CMV znajduje siê gen
bia³ka rtTA, które pod nieobecnoœæ doksycykliny nie jest w stanie wi¹zaæ siê z ope-
ratorem Tet-O i aktywowaæ transkrypcji genu wprowadzonego w plazmidzie pBI-X.
S³awomir Go³da i inni
90 PRACE EKSPERYMENTALNE
Rys. 6. Plazmidy u¿yte w doœwiadczeniach z czteroplazmidowym zmodyfikowanym systemem
Tet-On. A. Plazmid pTet-On koduje aktywatorowe bia³ko rtTA, aktywowane po dodaniu doksycykliny.
B. Plazmid pTet-TS koduje bia³ko inhibitorowe tTS, ulegaj¹ce inaktywacji po dodaniu doksycykliny.
C. Plazmid pBI-X zawiera sekwencjê �-galaktozydazy oraz sekwencjê MCS, dziêki której do plazmidu
mo¿na wprowadziæ dowolny gen X. D. Plazmid pTK-hyg koduje bia³ko opornoœci na hygromycynê umo¿-
liwiaj¹ce selekcjê komórek opornych na ten antybiotyk.

W celu unikniêcia przeciekania z plazmidu pBI-X genu X dodatkowo zastosowano
kotransfekcjê z drugim plazmidem pTet-TS koduj¹cym bia³ko tTS hamuj¹ce tran-
skrypcjê pod nieobecnoœæ doksycykliny. Bia³ko tTS sk³ada siê z aktywatora tetR
po³¹czonego z hamuj¹ca domen¹ SDKid-1. Procedurê transfekcji wykonano stosuj¹c
oligodendrymery SuperFect jak przy transfekcji przejœciowej.
Po oko³o trzech tygodniach selekcji w po¿ywce zawieraj¹cej 450 �g/mL genety-
cyny (G418; Gibco) komórki oporne na antybiotyk, czyli zawieraj¹ce plazmid
pTet-On z opornoœci¹ na neomycynê, poddano kolejnej kotransfekcji, do wykonania
której u¿yto 5 �g plazmidu pBI-X koduj¹cego �-galaktozydazê (gen reporterowy)
i wklonowany w miejsce MCS inny transgen (praca w przygotowaniu) oraz 0,1 �g
plazmidu pTK-hyg umo¿liwiaj¹cego niezale¿n¹ selekcjê pozytywnych klonów w obec-
noœci hygromycyny (rys. 6).
Po nastêpnych 3 tygodniach hodowli w obecnoœci G418 oraz hygromycyny (Invi-
trogen) pozytywne klony zosta³y sprawdzone na aktywnoœæ �-galaktozydazy po
podaniu ró¿nych dawek doksycykliny (Sigma). Ogólny schemat dzia³ania uzyskane-
go w ten sposób systemu Tet-On/Tet-S przedstawiono na rysunku 7.
Analiza statystyczna. Doœwiadczenia wykonywano w duplikatach i powtarzano
co najmniej dwukrotnie. Istotnoœæ statystyczn¹ sprawdzano testem t-studenta.
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 91
Rys. 7. Schemat dzia³ania zmodyfikowanego systemu ekspresyjnego Tet-On/TetS. Dodatek antybio-
tyku umo¿liwia bia³ku rtTA wi¹zanie siê do sekwencji aktywatorowej tetO i jednoczeœnie inaktywuje
bia³ko inhibitora transkrypcji tTS wi¹¿¹ce siê w tym samym miejscu. Usuniêcie doksycykliny powoduje
od³¹czenie siê bia³ka rtTA, aktywacjê inhibitora tTS i zahamowanie transkrypcji genu X.
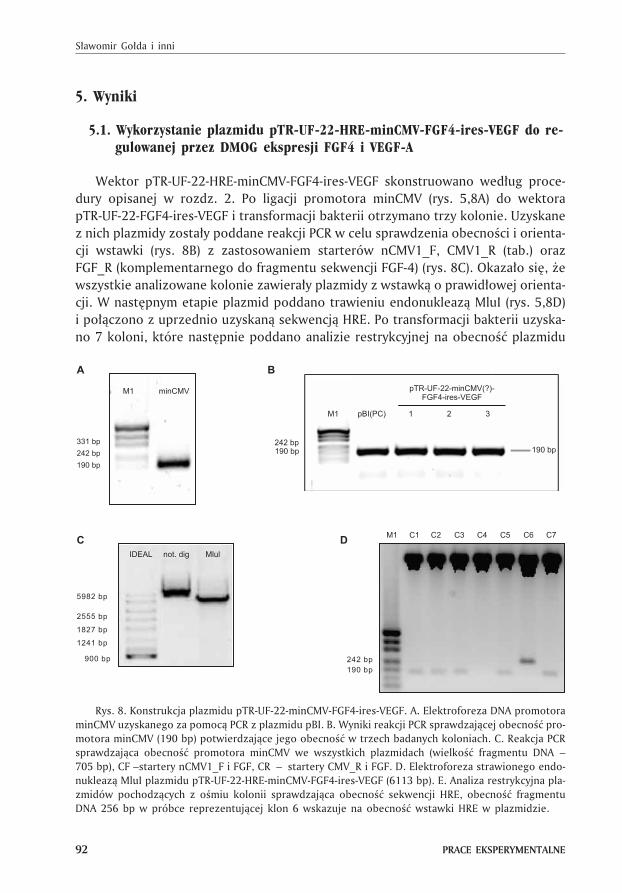
5. Wyniki
5.1. Wykorzystanie plazmidu pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF do re-gulowanej przez DMOG ekspresji FGF4 i VEGF-A
Wektor pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF skonstruowano wed³ug proce-
dury opisanej w rozdz. 2. Po ligacji promotora minCMV (rys. 5,8A) do wektora
pTR-UF-22-FGF4-ires-VEGF i transformacji bakterii otrzymano trzy kolonie. Uzyskane
z nich plazmidy zosta³y poddane reakcji PCR w celu sprawdzenia obecnoœci i orienta-
cji wstawki (rys. 8B) z zastosowaniem starterów nCMV1_F, CMV1_R (tab.) oraz
FGF_R (komplementarnego do fragmentu sekwencji FGF-4) (rys. 8C). Okaza³o siê, ¿e
wszystkie analizowane kolonie zawiera³y plazmidy z wstawk¹ o prawid³owej orienta-
cji. W nastêpnym etapie plazmid poddano trawieniu endonukleaz¹ MluI (rys. 5,8D)
i po³¹czono z uprzednio uzyskan¹ sekwencj¹ HRE. Po transformacji bakterii uzyska-
no 7 koloni, które nastêpnie poddano analizie restrykcyjnej na obecnoœæ plazmidu
S³awomir Go³da i inni
92 PRACE EKSPERYMENTALNE
Rys. 8. Konstrukcja plazmidu pTR-UF-22-minCMV-FGF4-ires-VEGF. A. Elektroforeza DNA promotora
minCMV uzyskanego za pomoc¹ PCR z plazmidu pBI. B. Wyniki reakcji PCR sprawdzaj¹cej obecnoœæ pro-
motora minCMV (190 bp) potwierdzaj¹ce jego obecnoœæ w trzech badanych koloniach. C. Reakcja PCR
sprawdzaj¹ca obecnoœæ promotora minCMV we wszystkich plazmidach (wielkoœæ fragmentu DNA –
705 bp), CF –startery nCMV1_F i FGF, CR – startery CMV_R i FGF. D. Elektroforeza strawionego endo-
nukleaz¹ MluI plazmidu pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF (6113 bp). E. Analiza restrykcyjna pla-
zmidów pochodz¹cych z oœmiu kolonii sprawdzaj¹ca obecnoœæ sekwencji HRE, obecnoœæ fragmentu
DNA 256 bp w próbce reprezentuj¹cej klon 6 wskazuje na obecnoœæ wstawki HRE w plazmidzie.
pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF z zastosowaniem enzymu EcoRI. Tylko jed-
na kolonia (numer 6) zawiera³a po¿¹dany plazmid (rys. 5,8E). Zosta³ on namno¿ony i u¿yty
do doœwiadczeñ sprawdzaj¹cych mo¿liwoœæ indukcji ekspresji FGF-4 oraz VEGF.
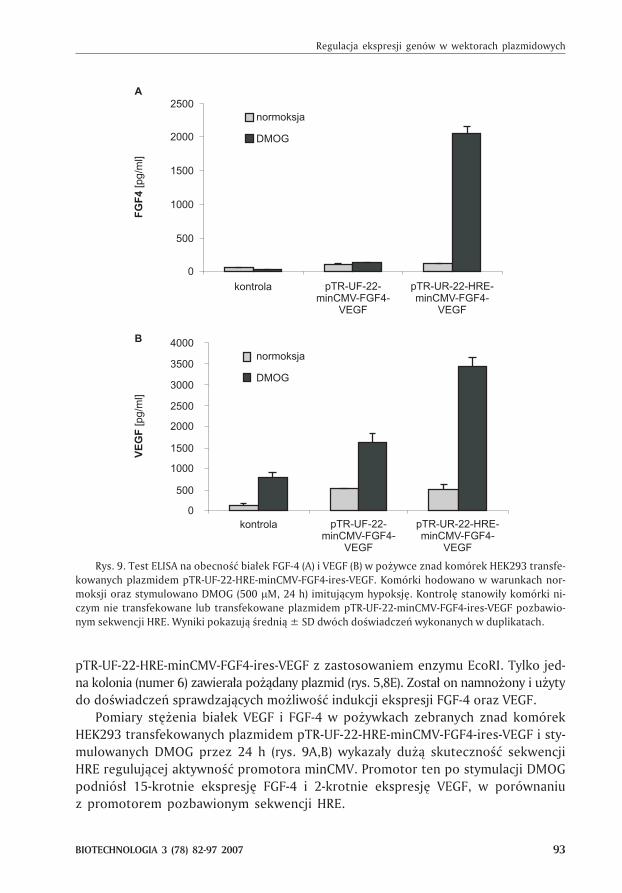
Pomiary stê¿enia bia³ek VEGF i FGF-4 w po¿ywkach zebranych znad komórek
HEK293 transfekowanych plazmidem pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF i sty-
mulowanych DMOG przez 24 h (rys. 9A,B) wykaza³y du¿¹ skutecznoœæ sekwencji
HRE reguluj¹cej aktywnoœæ promotora minCMV. Promotor ten po stymulacji DMOG
podniós³ 15-krotnie ekspresjê FGF-4 i 2-krotnie ekspresjê VEGF, w porównaniu
z promotorem pozbawionym sekwencji HRE.
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 93
Rys. 9. Test ELISA na obecnoœæ bia³ek FGF-4 (A) i VEGF (B) w po¿ywce znad komórek HEK293 transfe-
kowanych plazmidem pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF. Komórki hodowano w warunkach nor-
moksji oraz stymulowano DMOG (500 �M, 24 h) imituj¹cym hypoksjê. Kontrolê stanowi³y komórki ni-
czym nie transfekowane lub transfekowane plazmidem pTR-UF-22-minCMV-FGF4-ires-VEGF pozbawio-
nym sekwencji HRE. Wyniki pokazuj¹ œredni¹ ± SD dwóch doœwiadczeñ wykonanych w duplikatach.
5.2. Wykorzystanie systemu regulowanego przez doksycyklinê ekspresji ge-nów Tet-On
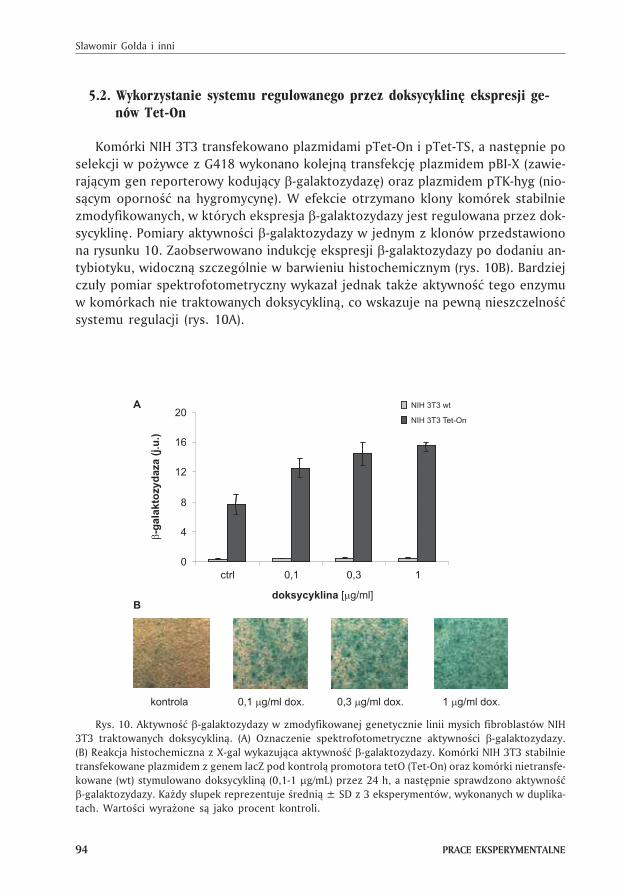
Komórki NIH 3T3 transfekowano plazmidami pTet-On i pTet-TS, a nastêpnie po
selekcji w po¿ywce z G418 wykonano kolejn¹ transfekcjê plazmidem pBI-X (zawie-
raj¹cym gen reporterowy koduj¹cy �-galaktozydazê) oraz plazmidem pTK-hyg (nio-
s¹cym opornoœæ na hygromycynê). W efekcie otrzymano klony komórek stabilnie
zmodyfikowanych, w których ekspresja �-galaktozydazy jest regulowana przez dok-
sycyklinê. Pomiary aktywnoœci �-galaktozydazy w jednym z klonów przedstawiono
na rysunku 10. Zaobserwowano indukcjê ekspresji �-galaktozydazy po dodaniu an-
tybiotyku, widoczn¹ szczególnie w barwieniu histochemicznym (rys. 10B). Bardziej
czu³y pomiar spektrofotometryczny wykaza³ jednak tak¿e aktywnoœæ tego enzymu
w komórkach nie traktowanych doksycyklin¹, co wskazuje na pewn¹ nieszczelnoœæ
systemu regulacji (rys. 10A).
S³awomir Go³da i inni
94 PRACE EKSPERYMENTALNE
Rys. 10. Aktywnoœæ �-galaktozydazy w zmodyfikowanej genetycznie linii mysich fibroblastów NIH
3T3 traktowanych doksycyklin¹. (A) Oznaczenie spektrofotometryczne aktywnoœci �-galaktozydazy.
(B) Reakcja histochemiczna z X-gal wykazuj¹ca aktywnoœæ �-galaktozydazy. Komórki NIH 3T3 stabilnie
transfekowane plazmidem z genem lacZ pod kontrol¹ promotora tetO (Tet-On) oraz komórki nietransfe-
kowane (wt) stymulowano doksycyklin¹ (0,1-1 �g/mL) przez 24 h, a nastêpnie sprawdzono aktywnoœæ
�-galaktozydazy. Ka¿dy s³upek reprezentuje œredni¹ ± SD z 3 eksperymentów, wykonanych w duplika-
tach. Wartoœci wyra¿one s¹ jako procent kontroli.

6. Dyskusja
W prezentowanej pracy opisano dwa systemy regulacji ekspresji genów wyko-
rzystywane w eksperymentalnej terapii genowej. Umo¿liwiaj¹ one uzyskanie eks-
presji wprowadzonego transgenu po podaniu antybiotyku doksycykliny lub poprzez
indukcjê czynnika transkrypcyjnego HIF-1, spowodowan¹ niedotlenieniem lub sty-
mulacj¹ farmakologiczn¹.
System regulacji ekspresji genów przez doksycyklinê zosta³ przedstawiony na
przyk³adzie indukcji �-galaktozydazy w komórkach NIH 3T3. Gen tego enzymu za-
warty jest w plazmidzie pBI posiadaj¹cym dodatkowo sekwencjê MCS umo¿liwia-
j¹c¹ wklonowanie dowolnego genu, którego ekspresjê mo¿na regulowaæ analogicz-
nie do ekspresji �-galaktozydazy. Wykorzystuj¹c ten wektor mo¿na w ³atwy sposób
w³¹czaæ i wy³¹czaæ badany gen, maj¹c jednoczeœnie mo¿liwoœæ wizualnej oceny po-
ziomu jego ekspresji poprzez pomiar aktywnoœci �-galaktozydazy. W naszych bada-
niach system ten wykorzystano do konstrukcji plazmidu zawieraj¹cego obok genu
reporterowego równie¿ sekwencjê koduj¹c¹ reduktazê biliwerdyny (BVR). Umo¿li-
wi³o to znalezienie odpowiedzi na szereg pytañ zwi¹zanych z funkcj¹ i rol¹ BVR in
vitro (praca w przygotowaniu).
Istotne znaczenie dla tego systemu ma wprowadzenie do komórek plazmidu
pTet-tTS koduj¹cego bia³ko tTS hamuj¹ce tzw. przeciekanie genu z plazmidu pBI.
Niestety, w praktyce rzadko udaje siê osi¹gn¹æ ca³kowite wyciszenie genu w plazmi-
dzie pBI, co potwierdzi³y równie¿ nasze obserwacje.
System Tet-On zastosowano m.in. do produkcji szczepionek przeciwko wiruso-
wi HIV-1 (18). W tym celu stworzono wariant wirusa, którego replikacja œciœle zale-
¿a³a od poziomu bia³ka aktywatorowego rtTA regulowanego przez doksycyklinê.
Dziêki temu replikacja wirusa HIV-rtTA mog³a byæ czasowo w³¹czana przez podanie
odpowiednich dawek antybiotyku, co umo¿liwia³o doœæ œcis³¹ kontrolê odpowiedzi
uk³adu odpornoœciowego (18).
Innym przyk³adem zastosowania tego systemu mo¿e byæ wykazanie znaczenia
bia³ka c-Src w rozwoju raka piersi. W tym celu stworzono liniê komórek MCF7 z in-
dukowan¹ przez doksycyklinê negatywn¹, dominuj¹c¹ form¹ bia³ka c-Src (c-SrcDN).
Zbadano zale¿noœci pomiêdzy bia³kiem c-Src, a szeregiem bia³ek sygna³owych oraz
zaobserwowano, ¿e nadekspresja bia³ka c-SrcDN ogranicza migracjê i proliferacjê
komórek. Nastêpnie wykazano, ¿e podanie komórek MCF7 zawieraj¹cych gen SrcDN
do myszy typu nude (z upoœledzonym uk³adem odpornoœciowym) prowadzi³o do
rozwoju nowotworu, a jednoczesne podanie doksycykliny bardzo drastycznie ogra-
nicza³o rozwój guza (19).
W pracach nad plazmidem aktywowanym przez niedotlenienie uzyskano wektor
pTR-UF-22-HRE-minCMV-FGF4-ires-VEGF koduj¹cy czynniki angiogenne FGF-4 i VEGF-A.
Plazmid ten zawiera syntetyczn¹ sekwencjê HRE (3 powtórzenia sekwencji HRE
z promotora genu PGK-1) reguluj¹c¹ aktywnoœæ promotora minCMV. Taki syntetycz-
ny promotor (HREminCMV), o potencjalnie du¿ych mo¿liwoœciach zastosowania
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 95

w terapii genowej, powinien wykazywaæ nisk¹ aktywnoœæ w normoksji, a wysok¹
w tkankach niedotlenionych. Niewielka samoistna ekspresja transgenu jest istotna,
poniewa¿ ogranicza ona dzia³anie bia³ka poza tkank¹ docelow¹, natomiast wysokie
stê¿enie czynnika terapeutycznego w miejscu po¿¹danym zwiêksza prawdopodo-
bieñstwo, ¿e wystêpuje on w dawce wystarczaj¹cej do wywo³ania efektu biologicz-
nego.
Wyniki testu ELISA wykaza³y du¿¹ skutecznoœæ syntetycznego konstruktu
HRE-minCMV, bowiem stymulacja DMOG, zwi¹zkiem aktywuj¹cym HIF-1, prowa-
dzi³a do 15-krotnego wzrostu stê¿enia FGF-4 i 2-krotnego VEGF-A. Na podstawie
stê¿enia FGF-4 i VEGF w po¿ywce stwierdzono równie¿, ¿e podstawowa aktywnoœæ
konstruktu HRE-minCMV w normoksji jest niska. Warto zauwa¿yæ, ¿e w warunkach
symulacji niedotlenienia przez DMOG (zahamowania aktywnoœci hydroksylaz proli-
nowych) poziom VEGF-A by³ wy¿szy ni¿ FGF-4. Mog³o to wynikaæ st¹d, ¿e aktywowa-
ny HIF-1 indukuje ekspresjê tak¿e endogennego VEGF-A, wykrywanego przez ten
sam test ELISA. Endogenny VEGF mo¿e byæ te¿ indukowany w odpowiedzi na sam¹
procedurê transfekcji, dlatego w komórkach kontrolnych transfekowanych plazmi-
dem pozbawionym sekwencji HRE poziom bia³ka VEGF w stosunku do komórek nie
transfekowanych by³ wy¿szy. Ekspresja FGF-4 w komórkach kontrolnych nie prze-
kracza³a natomiast progu detekcji.
Plazmid zawieraj¹cy geny czynników proangiogennych, takich jak FGF-4 i VEGF-A,
pod kontrol¹ promotora HRE-minCMV móg³by znaleŸæ zastosowanie na przyk³ad
w terapii genowej choroby niedokrwiennej serca lub leczeniu niedokrwienia koñ-
czyn. Co wa¿ne, opisany w pracy wektor plazmidowy mo¿e byæ u¿yty do produkcji
wektorów AAV (ang. adeno-associated virus), które mog¹ zapewniæ d³ugotrwa³¹ i wy¿-
sz¹ ekspresjê transgenów w docelowych tkankach.
Ograniczenie ekspresji czynników proangiogennych, a zw³aszcza VEGF, wy³¹cz-
nie do obszaru tkanek niedotlenionych, mo¿e mieæ zasadnicze znaczenie dla zdro-
wia pacjenta. Niekontrolowana nadekspresja VEGF mo¿e bowiem prowadziæ do po-
wstania naczyniaków (20), a tak¿e, poprzez stymulacjê angiogenezy, mo¿e nasilaæ
rozwój ukrytych nowotworów.
Kilka interesuj¹cych przyk³adów zastosowania wektorów regulowanych przez
niedotlenienie opisano w eksperymentalnej terapii chorób uk³adu kr¹¿enia. Uzyska-
no w ten sposób zwiêkszenie ekspresji VEGF w niedotlenionych tkankach transdu-
kowanych wektorem AAV zawieraj¹cym gen VEGF pod kontrol¹ HRE (21). Rozmaite
modyfikacje systemu regulacji ekspresji zale¿nej od HIF-1 wykorzystano tak¿e do
indukcji oksygenazy hemowej-1 w niedotlenionych kardiomiocytach (22-26) i ko-
mórkach progenitorowych œródb³onka (27). Szerszy opis tych strategii mo¿na zna-
leŸæ w pracach przegl¹dowych (28,29).
Skutecznoœæ systemów regulowanej ekspresji genów zosta³a potwierdzona
w pracach badawczych, co stwarza nadziejê na ich wykorzystanie w próbach klinicz-
nych. Mog¹ byæ one szczególnie przydatne tam, gdzie wymagana jest precyzyjna
kontrola zarówno miejsca jak i poziomu ekspresji transgenu.
S³awomir Go³da i inni
96 PRACE EKSPERYMENTALNE

Badania finansowano z projektu PBZ-KBN 096 P05 2004 i PBZ-KBN 107 P04 2004 (Ministerstwo Na-
uki i Szkolnictwa Wy¿szego). Agnieszka JaŸwa jest stypendystk¹ Funduszu im. Stanis³awa Estreichera.
Alicja Józkowicz jest beneficjentem Wellcome Trust (International Senior Research Fellowship).
Agnieszka £oboda jest stypendystk¹ Fundacji na rzecz Nauki Polskiej.
Literatura
1. Gossen M., Bujard H., (1992), Proc. Natl. Acad. Sci. USA, 89, 5547-5551.
2. Gossen M., Freundlieb S., Bender G., Muller G., Hillen W., Bujard H., (1995), Science, 268, 1766-1769.
3. Yin D. X., Zhu L., Schimke R. T., (1996), Anal. Biochem., 235, 195-201.
4. Triezenberg S. J., Kingsbury R. C., McKnight S. L., (1988), Genes Devel., 2, 718-729.
5. Hillen W., Berens C., (1994), Annual. Rev. Microbiol., 48, 345-369.
6. Zagórska A., Dulak J., (2004), Acta Biochim. Pol., 51, 563-585.
7. Lahiri S., Roy A., Baby S. M., Hoshi T., Semena G. L., Prabhakar N. R., (2006), Prog. Biophys. Mol.
Biol., 91, 249-286.
8. Semenza G. L., (2004), Physiology (Bethesda), 19, 176-182.
9. Semenza G. L., Wang G. L., (1992), Mol. Cell Biol., 12, 5447-5454.
10. Huang L. E., Gu J., Schau M., Bunn H. F., (1998), Proc. Natl. Acad. Sci. USA, 7, 7987-7992.
11. Bruick R. K., McKnight S. L., (2001), Science, 9, 1337-1340.
12. Ivan M., Kondo K., Yang H., Kim W., Valiando J., Ohh M., Salic A., Asara J. M., Lane W. S., Kaelin W.
G. Jr., (2001), Science, 20, 464-468.
13. Jaakkola P., Mole D. R., Tian Y. M., Wilson M. I., Gielbert J., Gaskell S. J., Kriegsheim A. V., Hebestreit H.
F., Mukherji M., Schofield C. J., Maxwell P. H., Pugh C. W., Ratcliffe P. J., (2001), Science, 20, 468-472.
14. Semenza G. L., (2002), Bioch. Pharm., 64, 993-998.
15. Kucharzewska P., Zagórska A., Leja J., JaŸwa A., Gozdecka M., Józkowicz A., Dulak J., (2007), Bio-
technologia, 3, 66-81.
16. Wegiel B., (2004), Genetic approach to the role of heme oxygenase 1 in human keratinocytes. Construction
of AAV vectors, praca magisterska, Wydzia³ Biotechnologii UJ, Kraków.
17. £oboda A., (2006), Transkrypton angiogenny komórki œródb³onka mikronaczyñ – wp³yw niedotlenienia
i oksygenazy hemowej, praca doktorska, Wydzia³ Biotechnologii UJ, Kraków.
18. Zhou X., Vink M., Berkhout B., Das A. T., (2006), Retrovirology, 3, 82.
19. Gonzalez L., Agullo-Ortuno M. T., Garcia-Martinez J. M., Calcabrini A., Gamallo C., Palacios J., Aran-
da A., Martin-Perez J,. (2006), J. Biol. Chem., 281, 20851-20864.
20. Lee R. J., Springer M. L., Blanco-Bose W. E., Shaw R., Ursell P. C., Blau H., (2000), Circulation, 102,
898-901.
21. Su H., Joho S., Huang Y., Barcena A., Arakawa-Hoyt J., Grossman W., Kan Y. W., (2004), Proc. Natl.
Acad. Sci. USA, 101, 16280-16285.
22. Tang Y., Jackson M., Qian K., Phillips M. I., (2002), Hypertension, 39, 695-698.
23. Tang Y. L., Tang Y., Zhang Y. C., Qian K., Shen L., Phillips M. I., (2004), Hypertension, 43, 746-751.
24. Tang Y. L., Tang Y., Zhang Y. C., Agarwal A., Kashara H., Qian K., Shen L., Phillips M. I., (2005), Gene
Ther., 12, 1163-1170.
25. Pachori A. S., Melo L. G., Hart M. L., Noiseux N., Zhang L., Morello F., Solomon S. D., Stahl G. L.,
Pratt R. E., Dzau V. J., (2004), Proc. Natl. Acad. Sci. USA, 101, 12282-12287.
26. Pachori A. S., Melo L.G., Zhang L., Solomon S. D., Dzau V. J., (2006), J. Am. Coll. Cardiol., 47,
635-643.
27. Tang Y. L., Tang Y., Zhang Y. C., Qian K., Shen L., Phillips M. I., (2005), J. Am. Coll. Cardiol., 46,
1339-1350.
28. Dulak J., Zagorska A., Wegiel B., £oboda A., Józkowicz A., (2006), Cell Biochem. Biophys., 44, 31-42.
29. JaŸwa A., Józkowicz A., Dulak J., (2007), Current Gene Therapy, 7, 7-23.
Regulacja ekspresji genów w wektorach plazmidowych
BIOTECHNOLOGIA 3 (78) 82-97 2007 97


















