Zaburzenia funkcji endogennych systemów - Strona startowa · Klasyczne opioidy (t.j.morfina,...
59
Barbara Przewłocka Zakład Farmakologii Bólu, Instytut Farmakologii, PAN Kraków Zaburzenia funkcji endogennych systemów opioidowych w przewlekłym bólu
Transcript of Zaburzenia funkcji endogennych systemów - Strona startowa · Klasyczne opioidy (t.j.morfina,...

Barbara PrzewłockaZakład Farmakologii Bólu, Instytut Farmakologii, PANKraków
Zaburzenia funkcji endogennych systemów
opioidowych w przewlekłym bólu

Dwa oblicza bólu
działanie ostrzegająco-ochronne
przewlekłe zespoły bólowe – choroba sama
w sobie

Ból jest to nieprzyjemne doznanie zmysłowe i
emocjonalne związane z aktualnie
występującym lub potencjalnym uszkodzeniem
tkanek, lub opisywane w kategoriach takiego
uszkodzenia
Międzynarodowe Stowarzyszenie Badania Bólu International Association for the Study of Pain
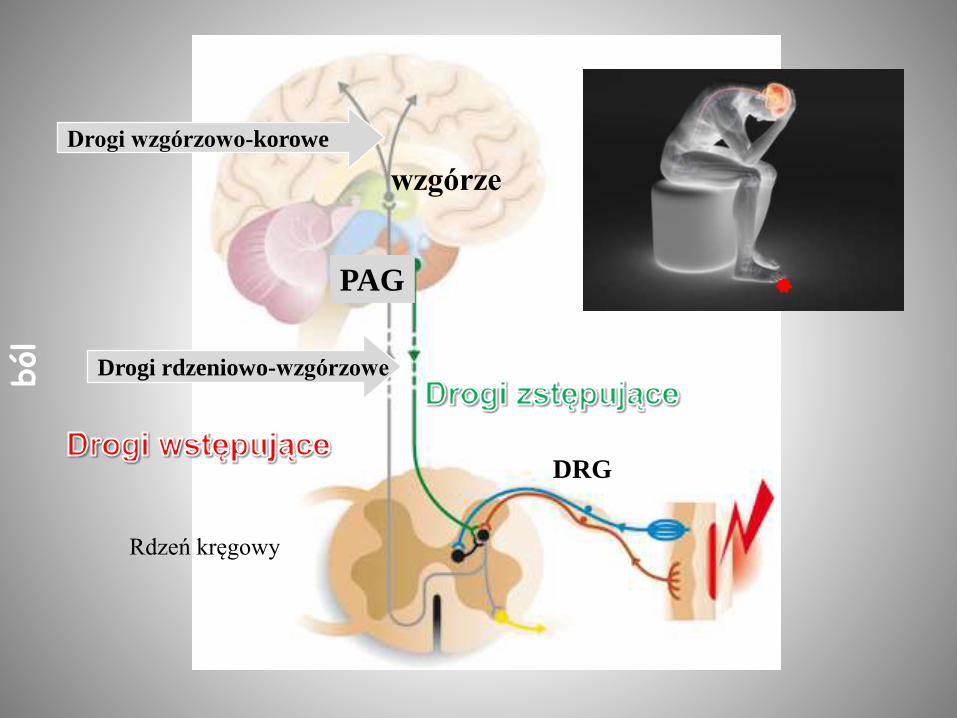
Drogi wzgórzowo-korowe
wzgórze
DRG
Drogi rdzeniowo-wzgórzowe
PAG
Rdzeń kręgowy
ból

Syrop na kaszel ?mor
fina

Radość dla świata. Perry Davis roślinny lek przeciwbólowymor
fina

Syrop na ząbkowaniemor
fina

Rzym, ok. 300 AD
mor
fina

1803 – Serturner wyizolował z opium alkaloid –morfinę
7,8-didehydro-4,5-epoksy-17-metylomorfinan-3,6-diol
Białą, gorzką substancję
nazwał „morfeusz” od
imienia greckiego boga marzeń
sennych
•pierwsze doniesienie dotyczące przeciwbólowego działania morfiny pojawiło się 150 lat temu (Wood, 1855)
mor
fina

Klasyczne opioidy (t.j.morfina, fentanyl, buprenorfina) działają
głównie przez ośrodkowy układ nerwowy
Efekty działania morfiny:
•hamuje perystaltykę żołądka i jelit, wywołuje zaparcia
•hamuje uwalnianie hormonów i neurotransmiterów
•nasila pobieranie pokarmu
•wpływa na temperaturę ciała (hipotermia/hipertermia)
•hamuje czynności oddechowe
•działa przeciwkaszlowo
•wywołuje bradykardię
•pobudza chemoreceptory ośrodka wymiotnego
•silne działa uspokajająco, wywołuje senność
•obniża nastrój – dysforia
•wywoluje tolerancję
MORFINA (μ μ1/μ2) mor
fina

Odkrycie specyficznego wiązania opiatów do tkanki mózgowej
1971 A. Goldstein
1973 C. Pert, S. Snyder
E. Simon, L.Terenius
ERIC J. SIMON PNAS 1973
opioidy

Odkrycie enkefalin
1975 - H. Kosterlitz i J. Hughes
Ekstrakt z mózgu hamuje wydzielanie acetylocholiny w preparacie jelita świnki morskiej.
opioidy

Le Bars et al., 2001
Liczba artykułów opublikowanych w latach 1970-2000, w których stosowano różne bodźce w badaniach
progu bólowego
opioidy

1979 - proopiomelanokortyna (POMC)
1982 - proenkefalina
1982 - prodynorfina
1995 - Nocyceptyna/orfanina (Nociceptin/orphaninFQ) Meunier; Reinscheid
Czy w OUN występuje substancja, która działa na ten sam receptor co morfina, na receptor m?
1997 Zadina - odkrycie dwóch nowych peptydów: endomorfiny-1 i endomorfiny-2
opioidy
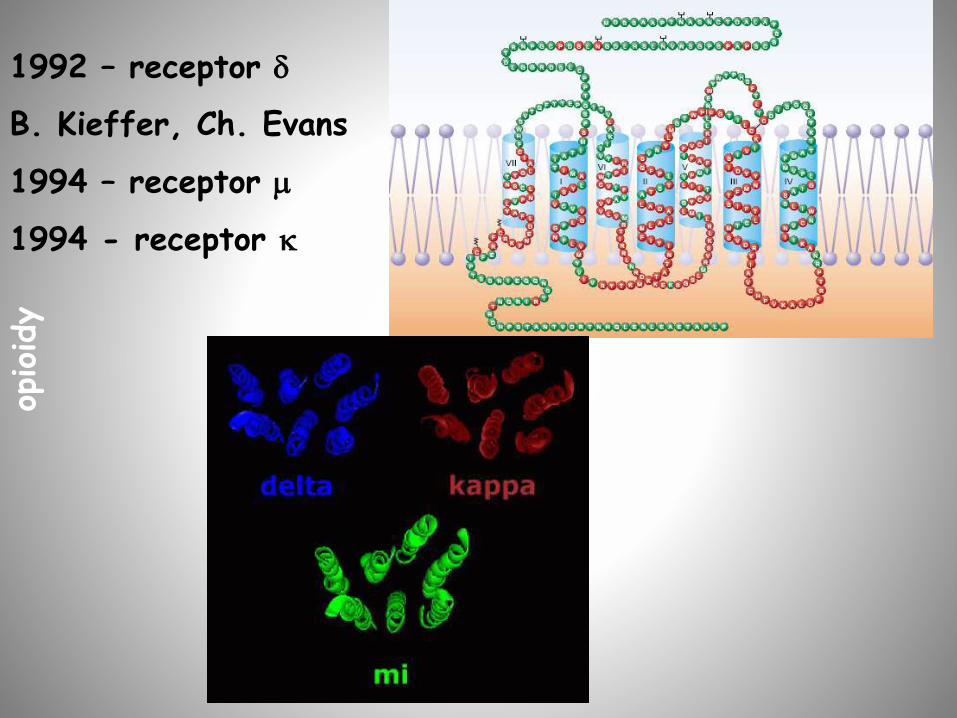
1992 – receptor d
B. Kieffer, Ch. Evans
1994 – receptor m
1994 - receptor k
opioidy

Klasyfikacja ligandów receptorów opioidowych:
• struktura alkaloidowa - opiaty– naturalnie występujące związki - morfina– związki syntetyczne – sufentanyl
• struktura peptydowa - opioidy- typowe peptydy opioidowesekwencja aminokwasowa na N-końcuH-Tyr-Gly-Gly-Phe- (Met-enkefalina, -endorfina, dynorfina)i ich syntetyczne pochodne ([D-Pen2,D-Pen5] enkefalina, DPDPE)(prohormon - proopiomelanokortyna, proenkefalina, prodynorfina)
- nietypowe peptydy opioidowesekwencja aminokwasowa na N-końcuH-Tyr-X-X-X- (kazomorfina, Tyr-W-MIF, endomorfina-1)i ich syntetyczne pochodne (PL017)
opioidy

Opiaty
Peptydyopioidowe
Opioidy
Terminologiaop
ioidy
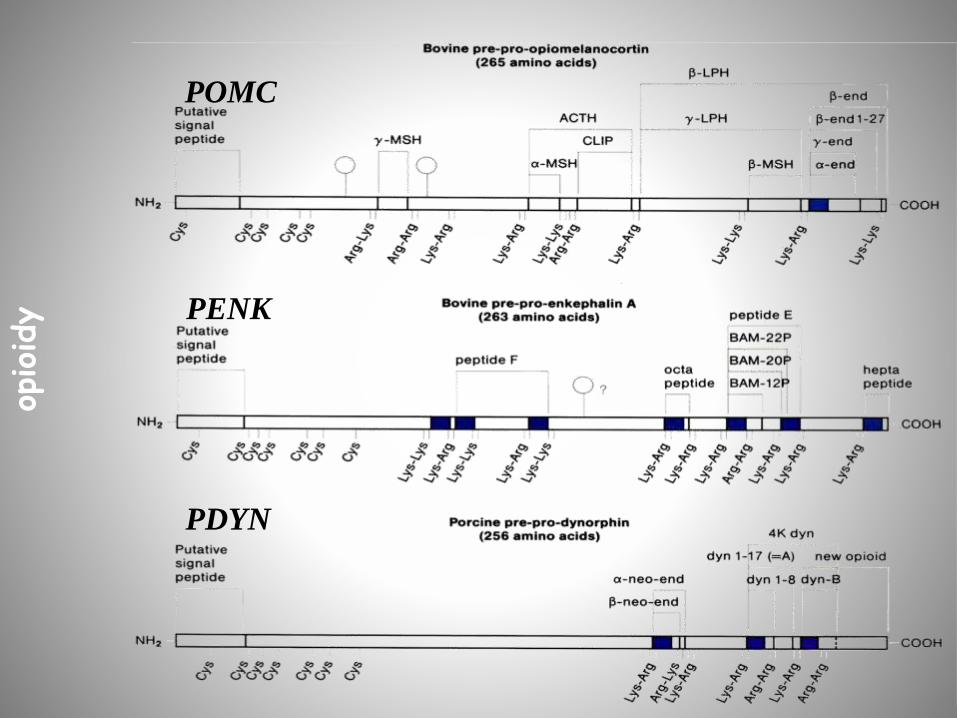
POMC
PENK
PDYN
opioidy
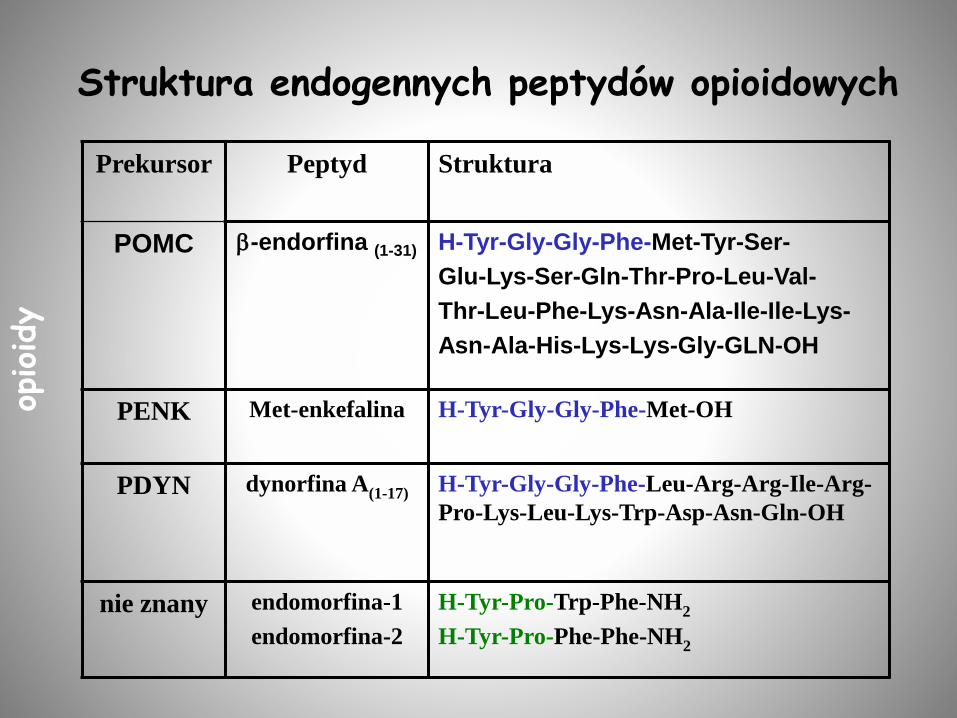
Struktura endogennych peptydów opioidowych
Prekursor Peptyd Struktura
POMC -endorfina (1-31) H-Tyr-Gly-Gly-Phe-Met-Tyr-Ser-
Glu-Lys-Ser-Gln-Thr-Pro-Leu-Val-
Thr-Leu-Phe-Lys-Asn-Ala-Ile-Ile-Lys-
Asn-Ala-His-Lys-Lys-Gly-GLN-OH
PENK Met-enkefalina H-Tyr-Gly-Gly-Phe-Met-OH
PDYN dynorfina A(1-17) H-Tyr-Gly-Gly-Phe-Leu-Arg-Arg-Ile-Arg-
Pro-Lys-Leu-Lys-Trp-Asp-Asn-Gln-OH
nie znany endomorfina-1
endomorfina-2
H-Tyr-Pro-Trp-Phe-NH2
H-Tyr-Pro-Phe-Phe-NH2
opioidy
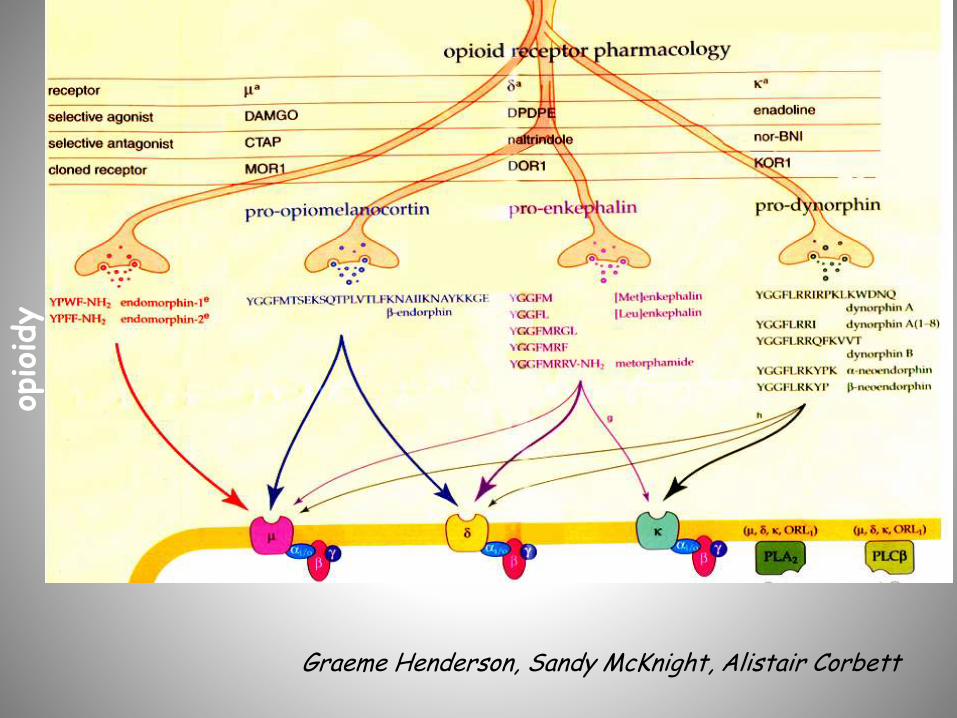
Graeme Henderson, Sandy McKnight, Alistair Corbett
opioidy

Endogenne peptydy opioidowe –selektywność receptorowa
Proopiomelanokortyna -Endorfina m , d
Proenkefalina [Met]enkefalina[Leu]enkefalina
BAM 1-22d , m
Prodynorfina Dynorfina ADynorfina A(1-8) Dynorfina B
k
m , d
Pronocyceptyna / OFQ Nocyceptyna ORL-1
Proendomorfina (?) Endomorfina-1Endomorfina-2
m
opioidy
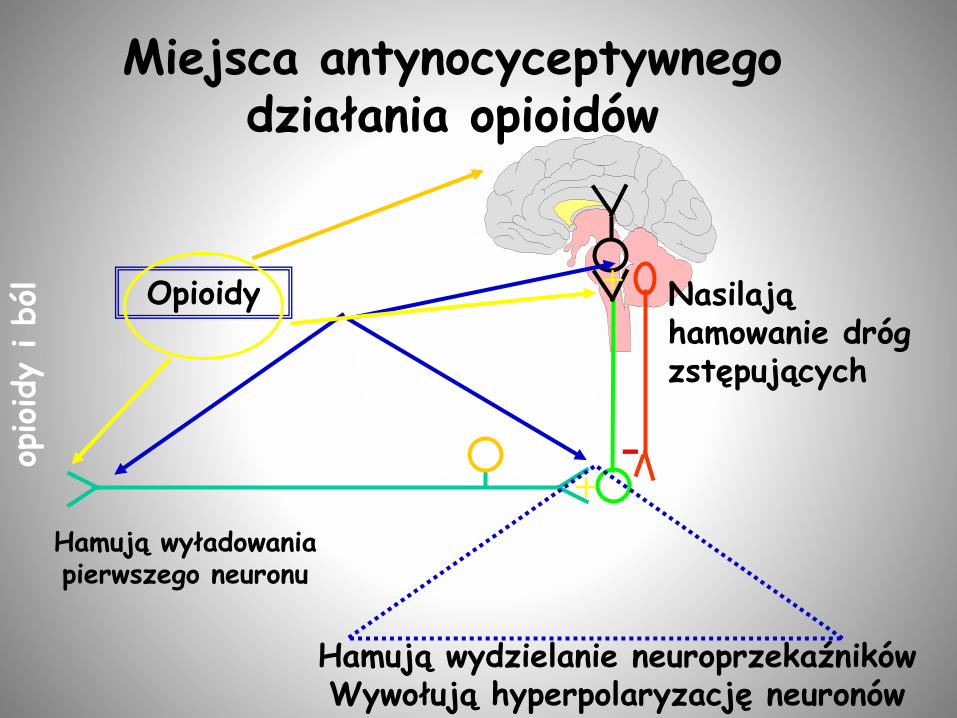
Miejsca antynocyceptywnego działania opioidów
Opioidy
Hamują wyładowania pierwszego neuronu
-+
+
Hamują wydzielanie neuroprzekaźnikówWywołują hyperpolaryzację neuronów
Nasilają hamowanie dróg zstępujących
opioidy i b
ól

Presynaptycznie - na zakończenia neuronów w rdzeniu kręgowym i na obwodzie - hamowanie wydzielania neuroprzekaźników
Postsynaptycznie - na receptory opioidowe –hyperpolaryzacja i hamowanie wywołanej przez bodziec bólowy aktywacji we wstępujących drogach nerwowych
Postsynaptycznie - na neurony hamującenp. GABA i w ten sposób dochodzi do odhamowanie neuronów (np. 5-HT) ograniczających transmisję nocyceptywną
MECHANIZM DZIAŁANIA OPIOIDÓW NA
TRANSMISJĘ NOCYCEPTYWNĄ
Neuron
hamujący
opioidy i b
ól
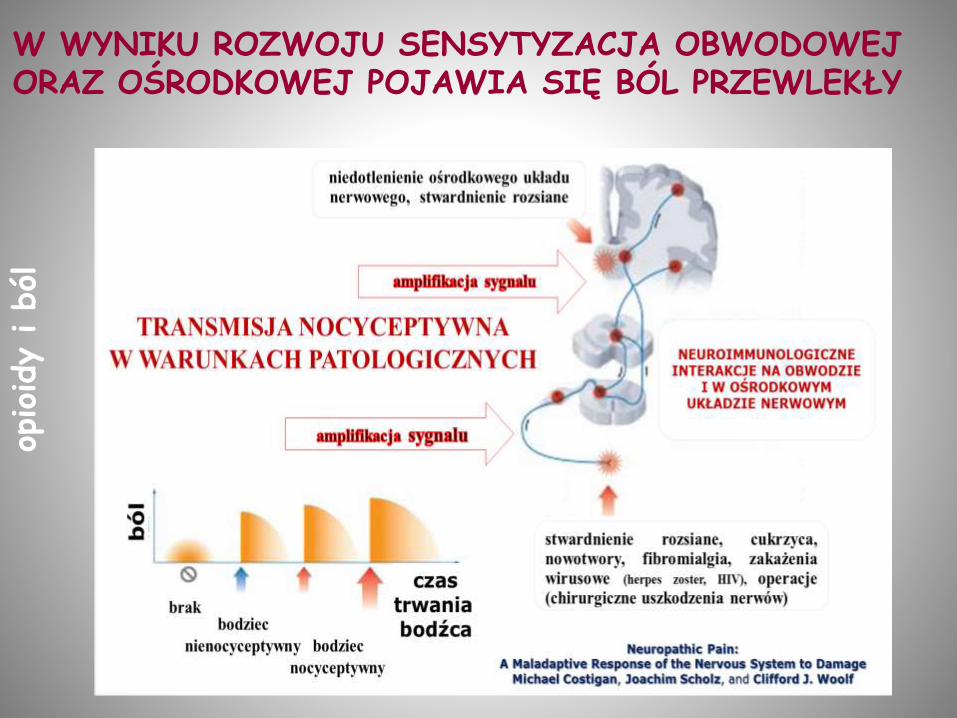
W WYNIKU ROZWOJU SENSYTYZACJA OBWODOWEJ ORAZ OŚRODKOWEJ POJAWIA SIĘ BÓL PRZEWLEKŁY
opioidy i b
ól
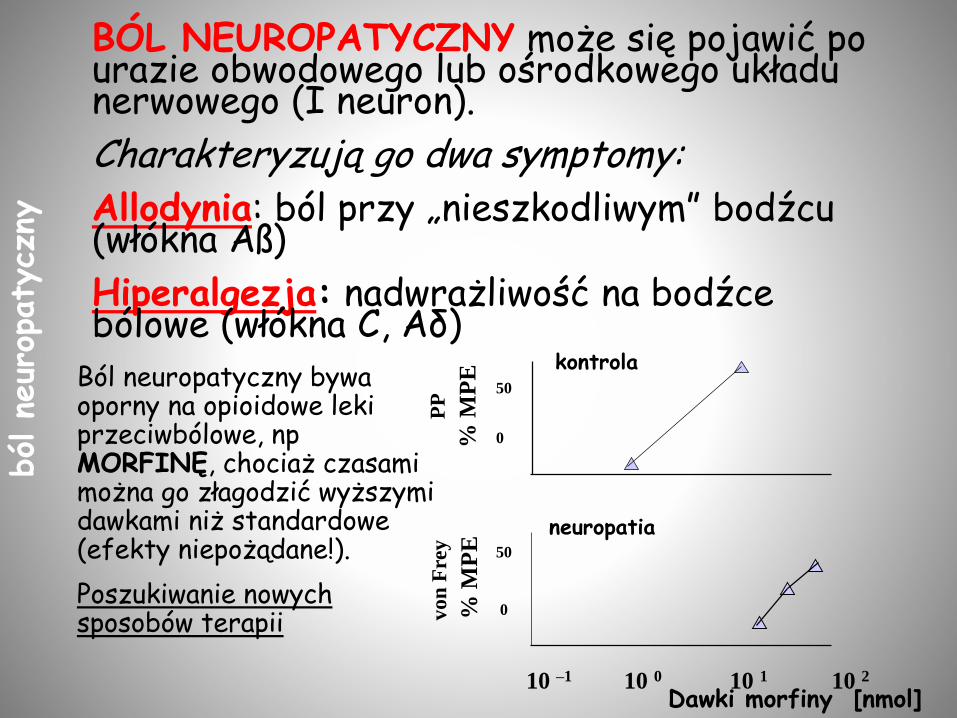
BÓL NEUROPATYCZNY może się pojawić po urazie obwodowego lub ośrodkowego układu nerwowego (I neuron).
Charakteryzują go dwa symptomy:
Allodynia: ból przy „nieszkodliwym” bodźcu (włókna Aß)
Hiperalgezja: nadwrażliwość na bodźce bólowe (włókna C, Aδ)
Ból neuropatyczny bywa oporny na opioidowe leki przeciwbólowe, np MORFINĘ, chociaż czasami można go złagodzić wyższymi dawkami niż standardowe (efekty niepożądane!).
Poszukiwanie nowych sposobów terapii
50
0
50
0
PP
% M
PE
vo
n F
rey
% M
PE
10 –1 10 0 10 1 10 2
neuropatia
kontrola
Dawki morfiny [nmol]
ból n
eur
opaty
czny

Mechanizm bólu neuropatycznego•Degeneracja zakończeń pierwotnych
Nuytten et al. 1992
•Rozrastanie się włókien A i tworzenie nowych synaps Woolf et al. 1992
•Tworzenie się miejsc o spontanicznej aktywności, które nasilają wrażliwość neuronów rdzeniowych
Wolf and Doubell 1994
•Nasilenie przekaźnictwa sympatycznego w zwojach rogów tylnych rdzenia
Xie et al. 1995
•Wzrost poziomu neuropeptydów w rogach tylnych rdzenia
Dubner 1991, Stanfa et al 1994
ból n
eur
opaty
czny
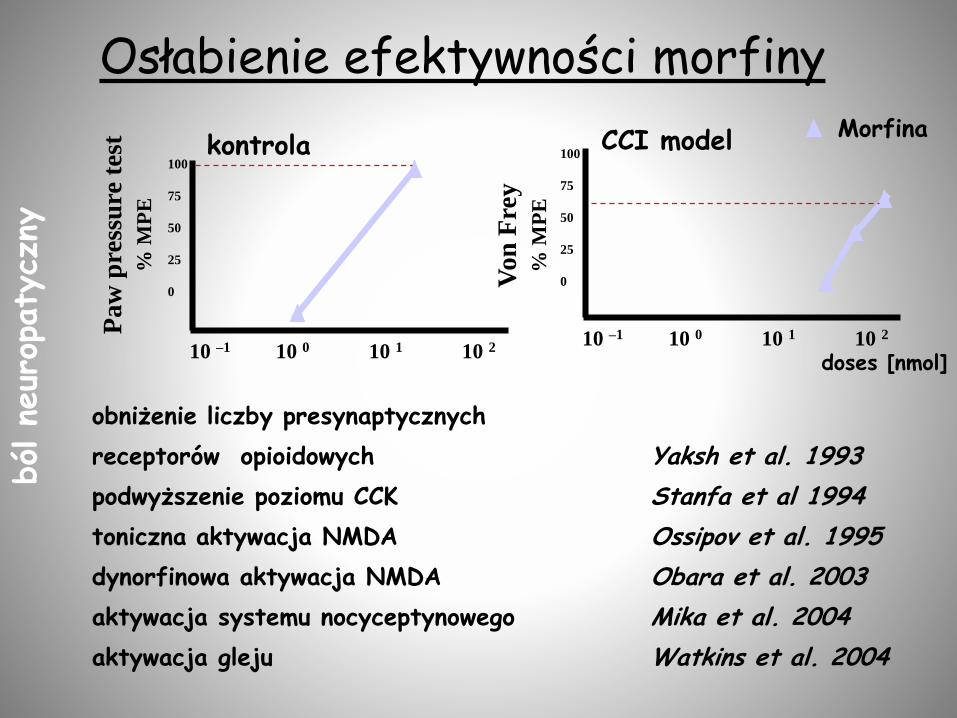
Osłabienie efektywności morfinyP
aw
pre
ssu
re t
est
% M
PE
Von
Fre
y
% M
PE
100
75
50
25
0
100
75
50
25
0
10 –1 10 0 10 1 10 2 10 –1 10 0 10 1 10 2
doses [nmol]
CCI modelkontrolaMorfina
obniżenie liczby presynaptycznych
receptorów opioidowych Yaksh et al. 1993
podwyższenie poziomu CCK Stanfa et al 1994
toniczna aktywacja NMDA Ossipov et al. 1995
dynorfinowa aktywacja NMDA Obara et al. 2003
aktywacja systemu nocyceptynowego Mika et al. 2004
aktywacja gleju Watkins et al. 2004
ból n
eur
opaty
czny
POMC
PENK
PDYN
opioidy
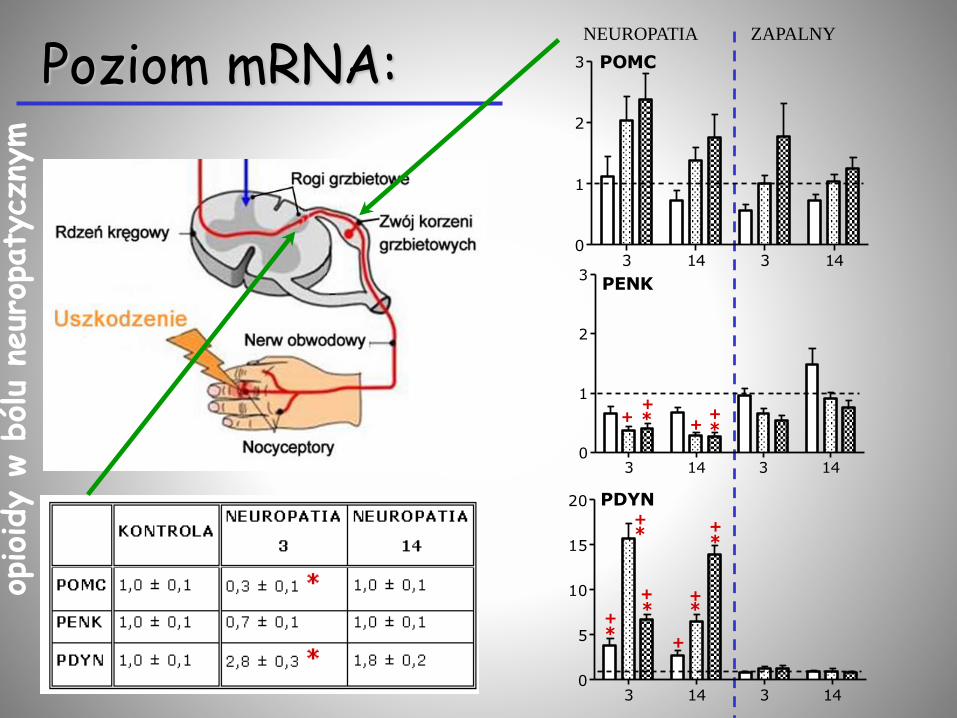
Poziom mRNA:NEUROPATIA ZAPALNY
3 14 3 140
1
2
3 POMC
3 14 3 140
1
2
3PENK
+ *+
+ *+
3 14 3 140
5
10
15
20 PDYN
+*
*
* *
*
+
+
+
+
+
opioidy w
bólu
neur
opaty
czny
m
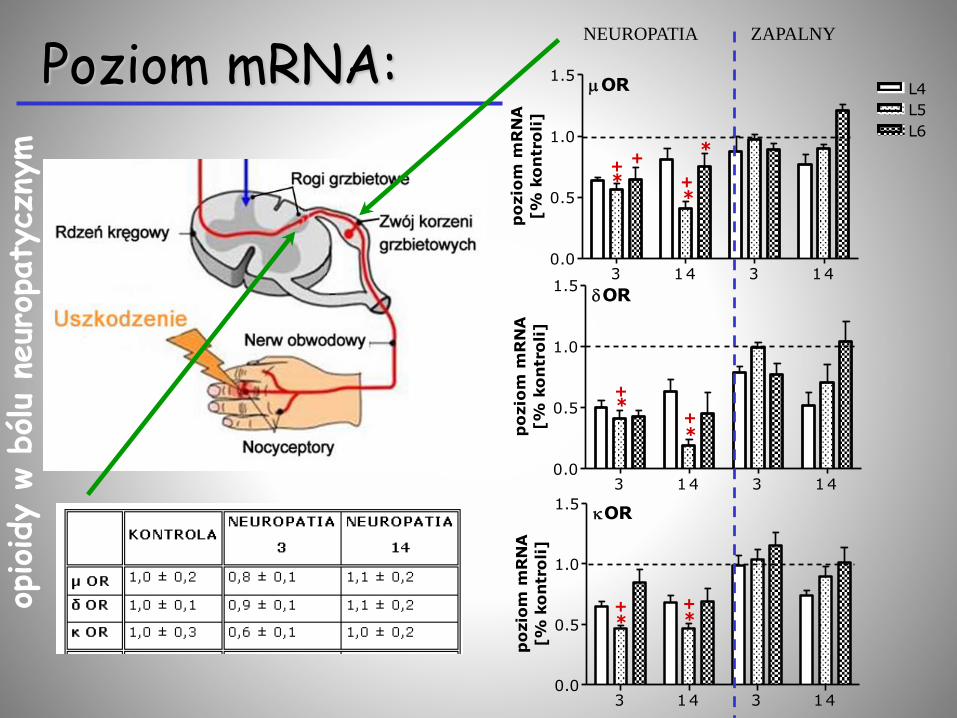
3 14 3 140.0
0.5
1.0
1.5L4
L5
L6
mOR
+*
*
*
+
+
po
zio
m m
RN
A
[%
ko
ntr
oli
]
3 14 3 140.0
0.5
1.0
1.5dOR
+*
*+
po
zio
m m
RN
A
[%
ko
ntr
oli
]
3 14 3 140.0
0.5
1.0
1.5kOR
* *+ +
po
zio
m m
RN
A
[%
ko
ntr
oli
]
Poziom mRNA:NEUROPATIA ZAPALNY
opioidy w
bólu
neur
opaty
czny
m

POMC - wspólny prekursor melanokortyn i opioidów
PROOPIOMELANOKORTYNA (POMC)
Pro--MSH/N-POMC JP -LPHACTH
-LPH -END
N-LPH -MSH
-MSH CLIP3-MSHN-POMC(1-49)
2-MSH
1-MSH
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
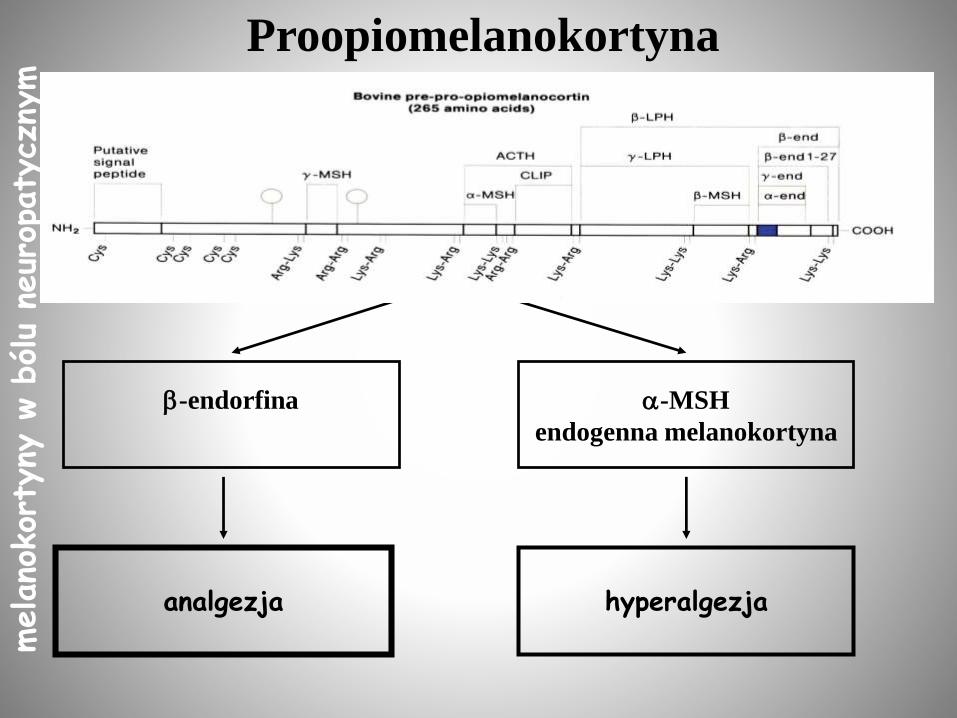
analgezja
-endorfina -MSH
endogenna melanokortyna
hyperalgezja
Proopiomelanokortynamelano
kor
tyny
w b
ólu
neur
opaty
czny
m
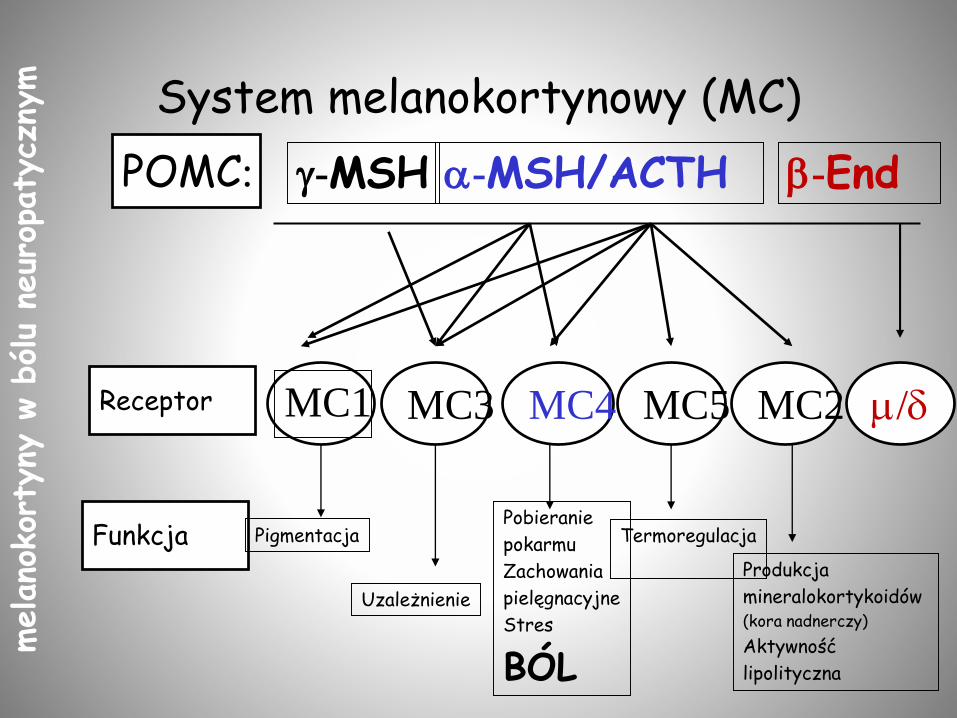
System melanokortynowy (MC)
Funkcja
Receptor MC1
POMC: -MSH -MSH/ACTH -End
Pigmentacja
Uzależnienie
Pobieranie
pokarmu
Zachowania
pielęgnacyjne
Stres
BÓL
Termoregulacja
Produkcja
mineralokortykoidów (kora nadnerczy)
Aktywność
lipolityczna
MC4 MC5 MC2 m/dMC3
melano
kor
tyny
w b
ólu
neur
opaty
czny
m

mRNARdzeń kręgowy
rogi grzbietowe warstwa I +++
warstwa II +++
warstwa X +++
rogi brzuszne +
Podwzgórze +
Wzgórze +++
Jądro migdałowate +++
------------------------------------------------------------------------------------------
Ekspresja receptora MC4 w rejonach OUNzaangażowanych w nocycepcję
Kishi i wsp., Expression of melanocortin 4 receptor mRNA in the central nervous system of the rat.
J Comp Neurol. 2003;457(3):213-35.
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
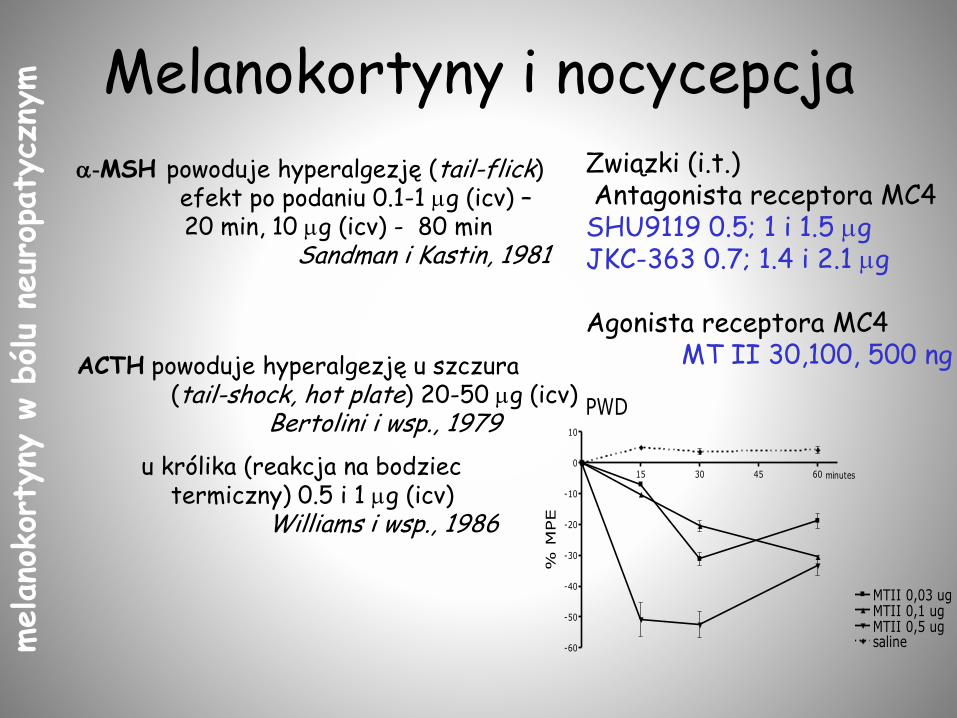
Melanokortyny i nocycepcja
-MSH powoduje hyperalgezję (tail-flick)efekt po podaniu 0.1-1 mg (icv) –20 min, 10 mg (icv) - 80 min
Sandman i Kastin, 1981
ACTH powoduje hyperalgezję u szczura (tail-shock, hot plate) 20-50 mg (icv)
Bertolini i wsp., 1979
u królika (reakcja na bodziec termiczny) 0.5 i 1 mg (icv)
Williams i wsp., 1986
15 30 45 60
-60
-50
-40
-30
-20
-10
0
10
MTII 0,03 ugMTII 0,1 ugMTII 0,5 ug
PWD
minutes
saline
% M
PE
Związki (i.t.) Antagonista receptora MC4
SHU9119 0.5; 1 i 1.5 mgJKC-363 0.7; 1.4 i 2.1 mg
Agonista receptora MC4 MT II 30,100, 500 ng
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
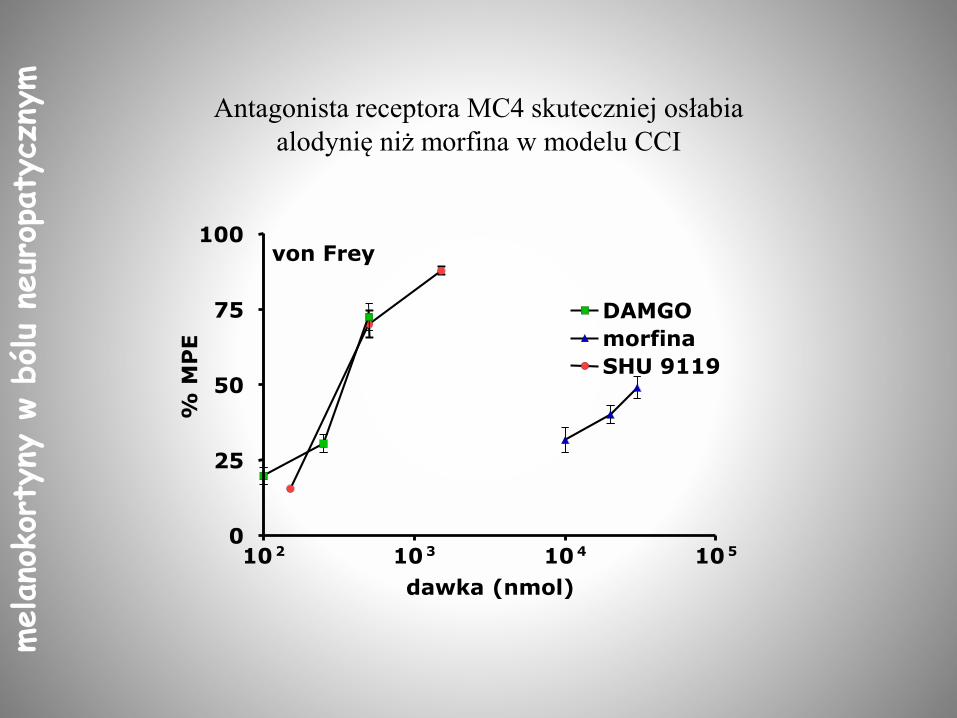
Antagonista receptora MC4 skuteczniej osłabia
alodynię niż morfina w modelu CCI
10 2 10 3 10 4 10 50
25
50
75
100
DAMGO
morfina
SHU 9119
von Frey
dawka (nmol)
% M
PE
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
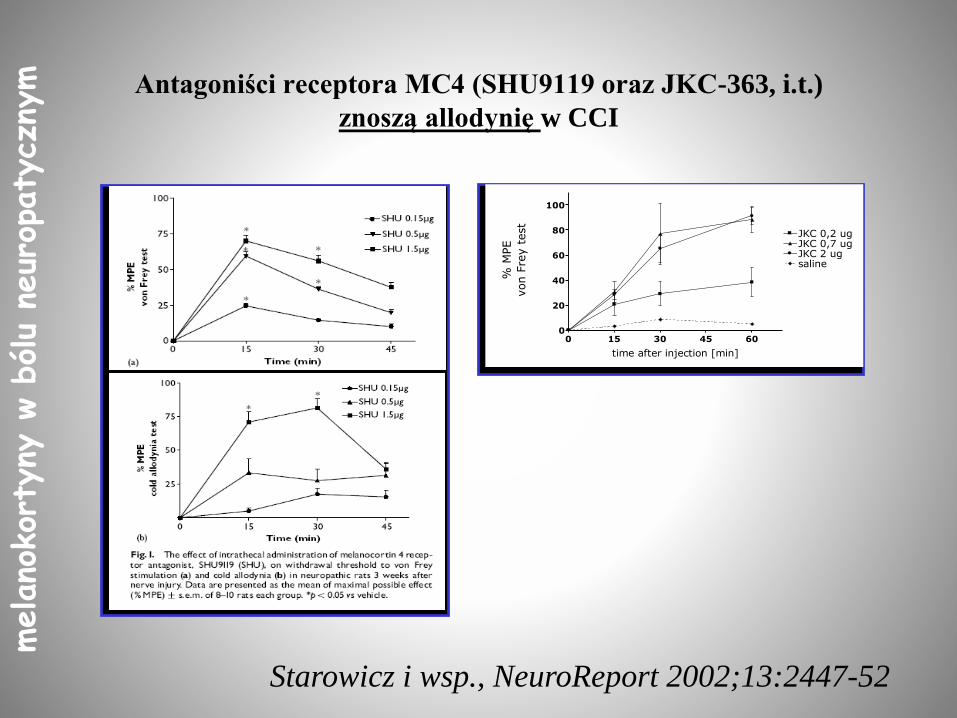
Antagoniści receptora MC4 (SHU9119 oraz JKC-363, i.t.)
znoszą allodynię w CCI
Starowicz i wsp., NeuroReport 2002;13:2447-52
0 15 30 45 600
20
40
60
80
100
JKC 0,2 ugJKC 0,7 ugJKC 2 ugsaline
time after injection [min]
% M
PE
von F
rey t
est
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
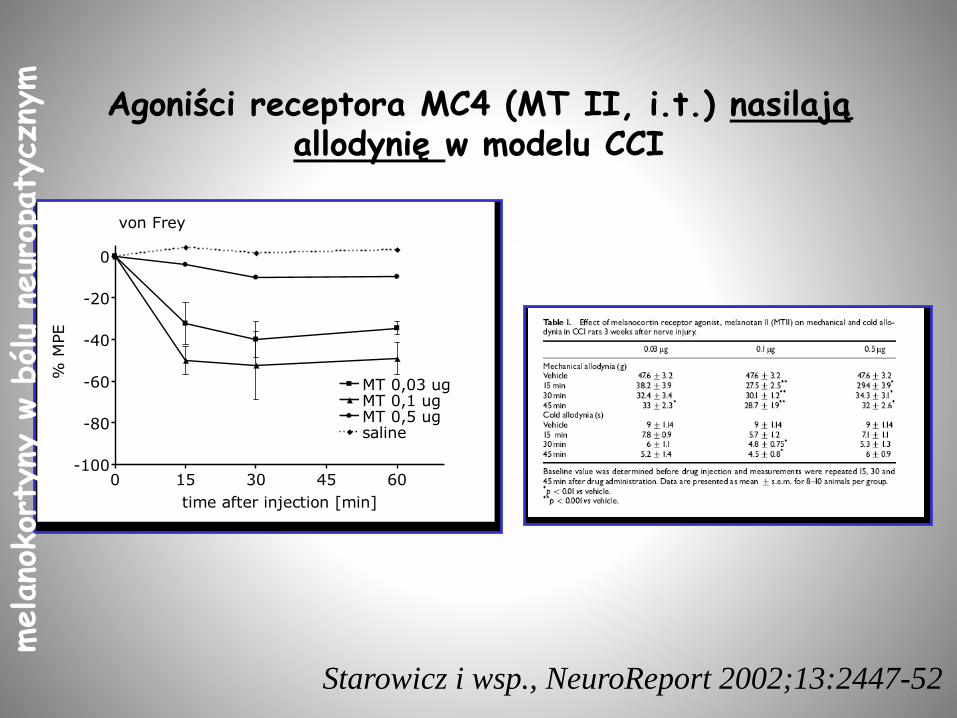
Agoniści receptora MC4 (MT II, i.t.) nasilają allodynię w modelu CCI
0 15 30 45 60-100
-80
-60
-40
-20
0
MT 0,03 ugMT 0,1 ugMT 0,5 ugsaline
von Frey
time after injection [min]
% M
PE
Starowicz i wsp., NeuroReport 2002;13:2447-52
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
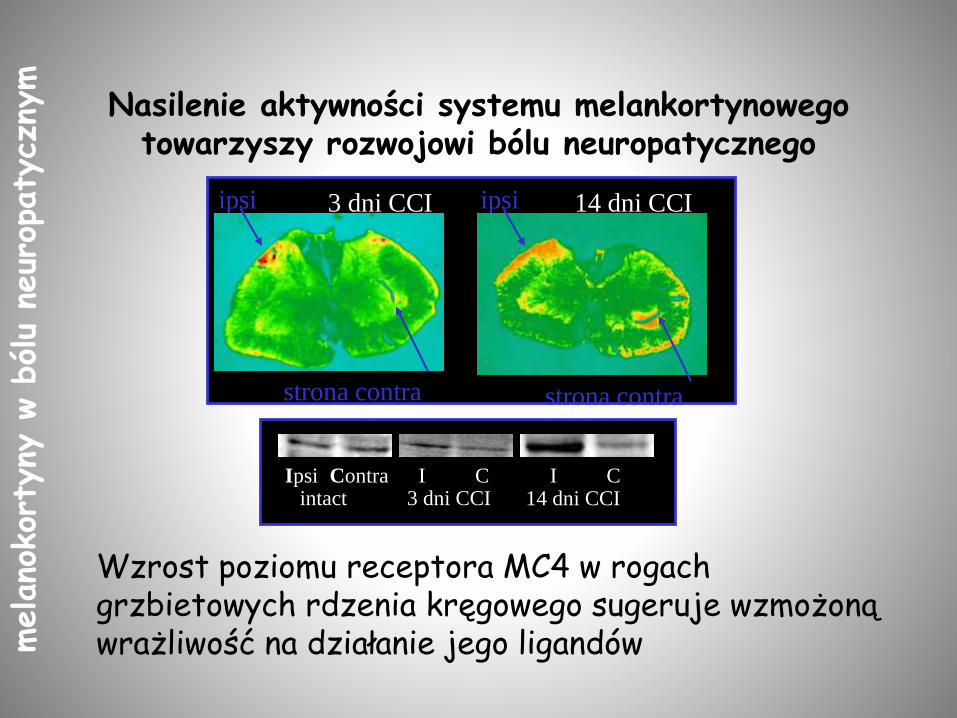
Nasilenie aktywności systemu melankortynowegotowarzyszy rozwojowi bólu neuropatycznego
strona contra
ipsi ipsi
strona contra
3 dni CCI 14 dni CCI
Ipsi Contra I C I Cintact 3 dni CCI 14 dni CCI
Wzrost poziomu receptora MC4 w rogach grzbietowych rdzenia kręgowego sugeruje wzmożoną wrażliwość na działanie jego ligandów melano
kor
tyny
w b
ólu
neur
opaty
czny
m
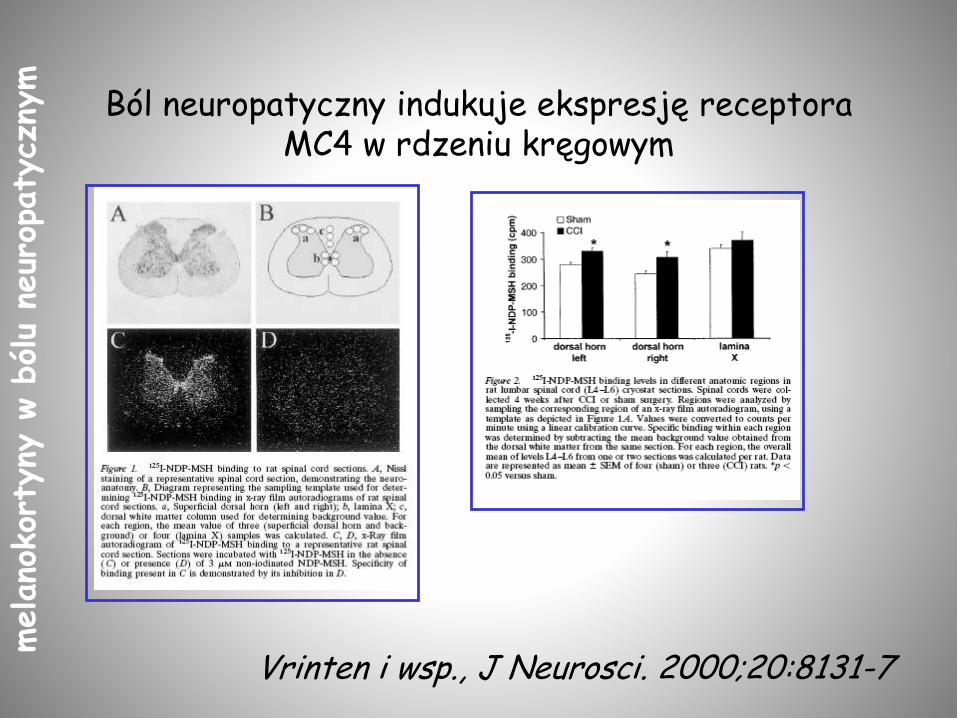
Ból neuropatyczny indukuje ekspresję receptora MC4 w rdzeniu kręgowym
Vrinten i wsp., J Neurosci. 2000;20:8131-7
melano
kor
tyny
w b
ólu
neur
opaty
czny
m

HIPOTEZA
• Równowaga między stałą impulsacją probólową (rdzeniowy system MC) a przeciwbólową (rdzeniowy system opioidowy)
?Czy efektywność działania opioidów może być
nasilona przez modulacje aktywności systemu MC
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
0 15 30 45 600
10
20
30
40
50
60
70
80
90
SHU 0,5MF 30SHU 0,5+MF 30
min
% M
PE (
von
Frey)
0 10 20 300
20
40
60
80
100
MFSHU+MFSHU
**
***
*
####
SHU+MF vs. MFalone
SHU+MF vs. SHU alone
MF dose (ug)%
MP
E
(von
Frey)
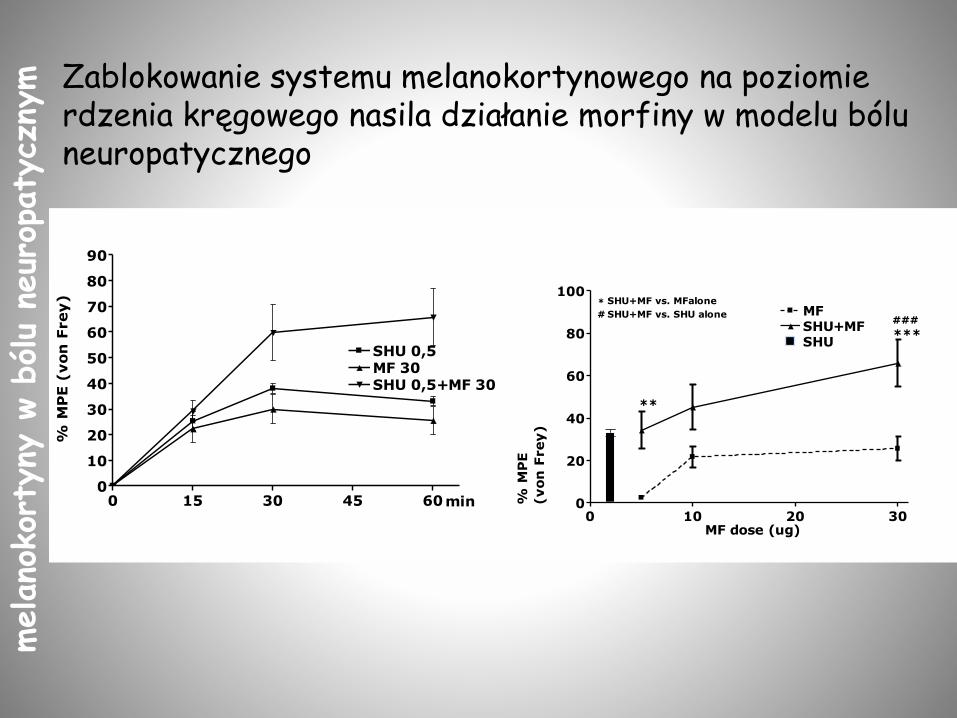
Zablokowanie systemu melanokortynowego na poziomie rdzenia kręgowego nasila działanie morfiny w modelu bólu neuropatycznego
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
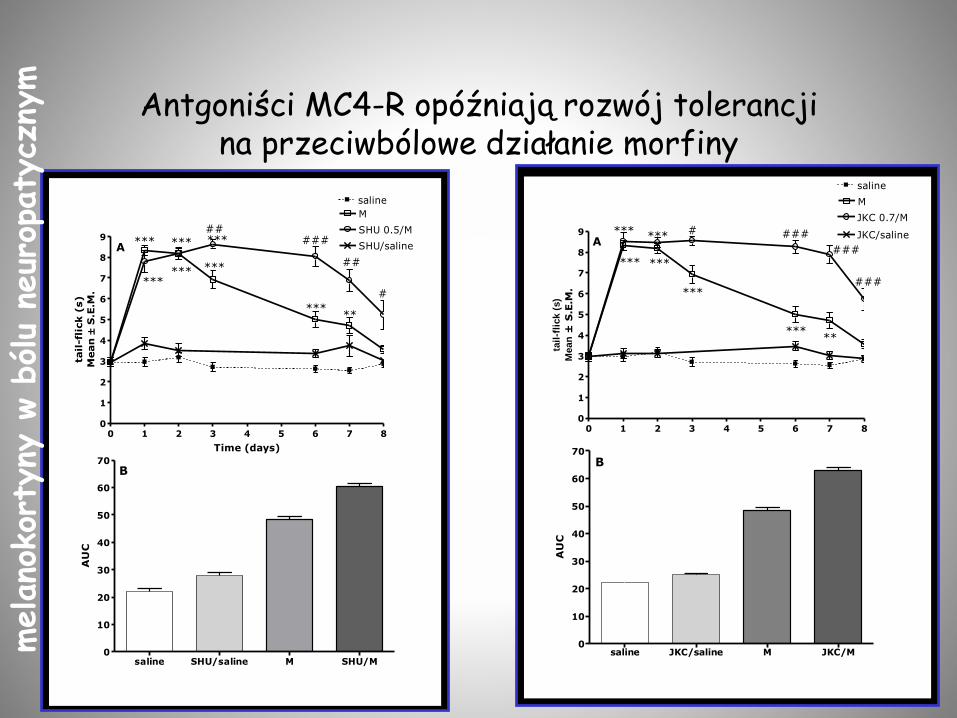
Antgoniści MC4-R opóźniają rozwój tolerancji na przeciwbólowe działanie morfiny
0 1 2 3 4 5 6 7 80
1
2
3
4
5
6
7
8
9
saline
M
SHU 0.5/M***
***
***
*** ***
***
*****
#####
##
#
SHU/salineA
Time (days)
tail
-fl
ick (
s)
Mean
S
.E.M
.
saline SHU/saline M SHU/M0
10
20
30
40
50
60
70B
AU
C
0 1 2 3 4 5 6 7 80
1
2
3
4
5
6
7
8
9
saline
M
JKC 0.7/M
***
***
***
***
***
*****
# ###
###
###
JKC/salineA
tail
-fli
ck (
s)
Mean
± S
.E.M
.
saline JKC/saline M JKC/M0
10
20
30
40
50
60
70
B
AU
C
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
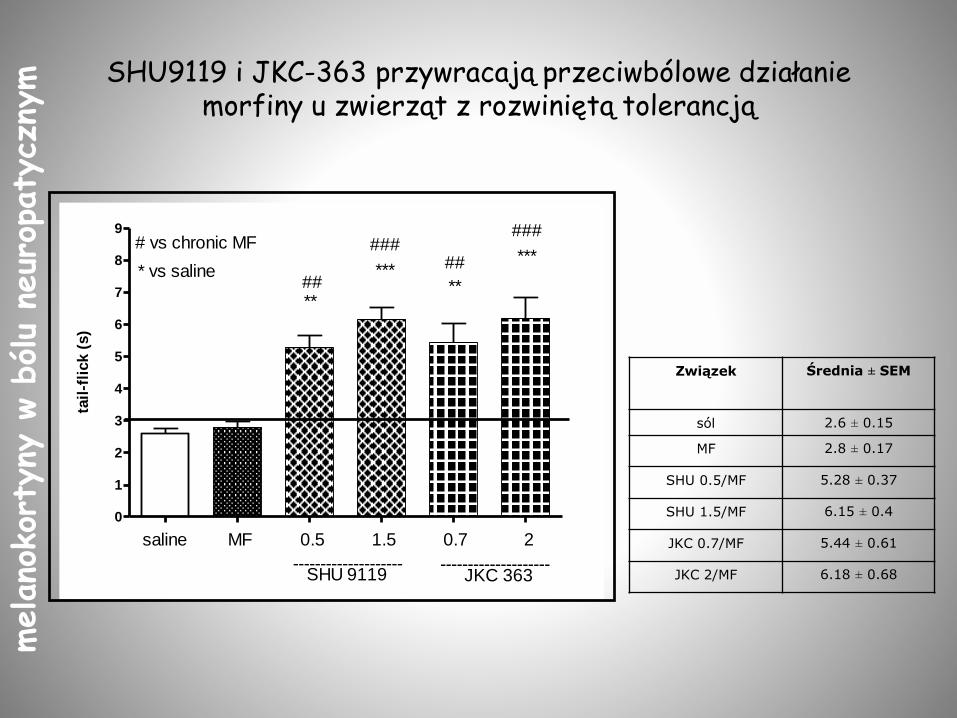
SHU9119 i JKC-363 przywracają przeciwbólowe działanie morfiny u zwierząt z rozwiniętą tolerancją
0
1
2
3
4
5
6
7
8
9
****
******
####
######
# vs chronic MF
saline MF
-------------------- --------------------
0.5 1.5 0.7 2
SHU 9119 JKC 363
* vs saline
tail
-fli
ck (
s)
Związek Średnia ± SEM
sól 2.6 ± 0.15
MF 2.8 ± 0.17
SHU 0.5/MF 5.28 ± 0.37
SHU 1.5/MF 6.15 ± 0.4
JKC 0.7/MF 5.44 ± 0.61
JKC 2/MF 6.18 ± 0.68
melano
kor
tyny
w b
ólu
neur
opaty
czny
m

PodsumowanieRdzeniowe receptory MC4 są zaangażowane w transmisjęnocyceptywną
Antagoniści MC4-R (SHU9119, JKC-363) znoszą allodynię i hiperalgezję, natomiast agoniści (MT II) nasilają te objawy
Zablokowanie rdzeniowych receptorów MC4 nasila działanie morfiny w modelu bólu neuropatycznego
Antagoniści receptora MC4: SHU9119 i JKC 363 (i.t.) opóźniają rozwój tolerancji rozwijającej się po wielokrotnych podaniach morfiny
U zwierząt z rozwiniętą tolerancją na przeciwbólowe działanie morfiny, SHU9119 i JKC-363 blokując rdzeniowe receptory MC4 przywracają jej analgetyczne działanie
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
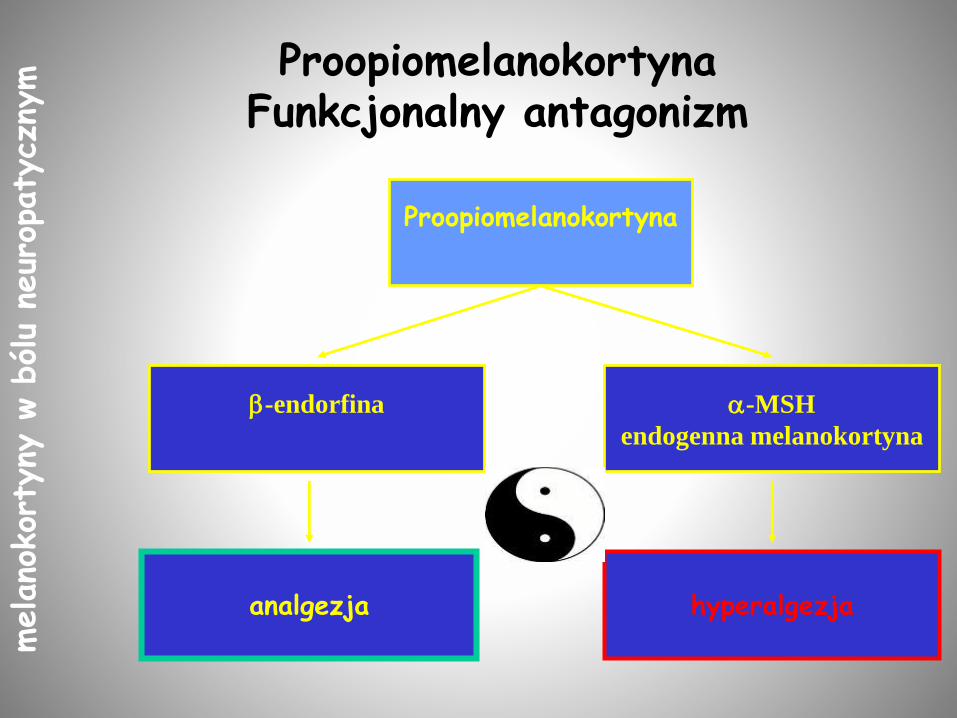
ProopiomelanokortynaFunkcjonalny antagonizm
Proopiomelanokortyna
analgezja
-endorfina -MSH
endogenna melanokortyna
hyperalgezja
melano
kor
tyny
w b
ólu
neur
opaty
czny
m
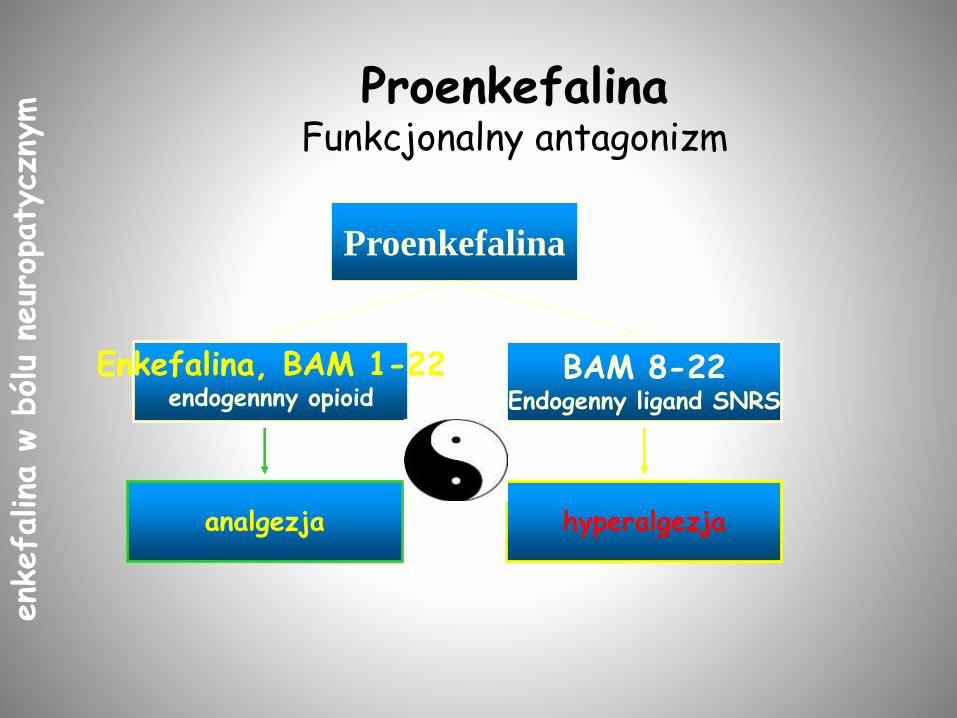
Proenkefalina
analgezja
Enkefalina, BAM 1-22endogennny opioid
BAM 8-22Endogenny ligand SNRS
hyperalgezja
ProenkefalinaFunkcjonalny antagonizm
enk
efa
lina
w b
ólu
neur
opaty
czny
m
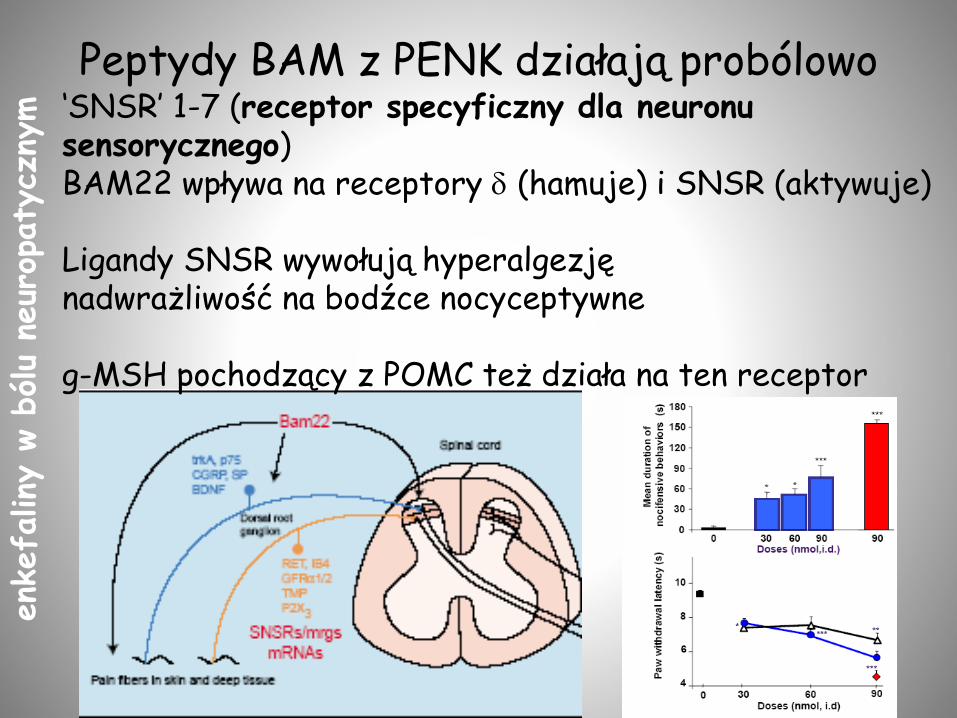
Peptydy BAM z PENK działają probólowo‘SNSR’ 1-7 (receptor specyficzny dla neuronu sensorycznego)BAM22 wpływa na receptory d (hamuje) i SNSR (aktywuje)
Ligandy SNSR wywołują hyperalgezjęnadwrażliwość na bodźce nocyceptywne
g-MSH pochodzący z POMC też działa na ten receptor
enk
efa
liny
w b
ólu
neur
opaty
czny
m
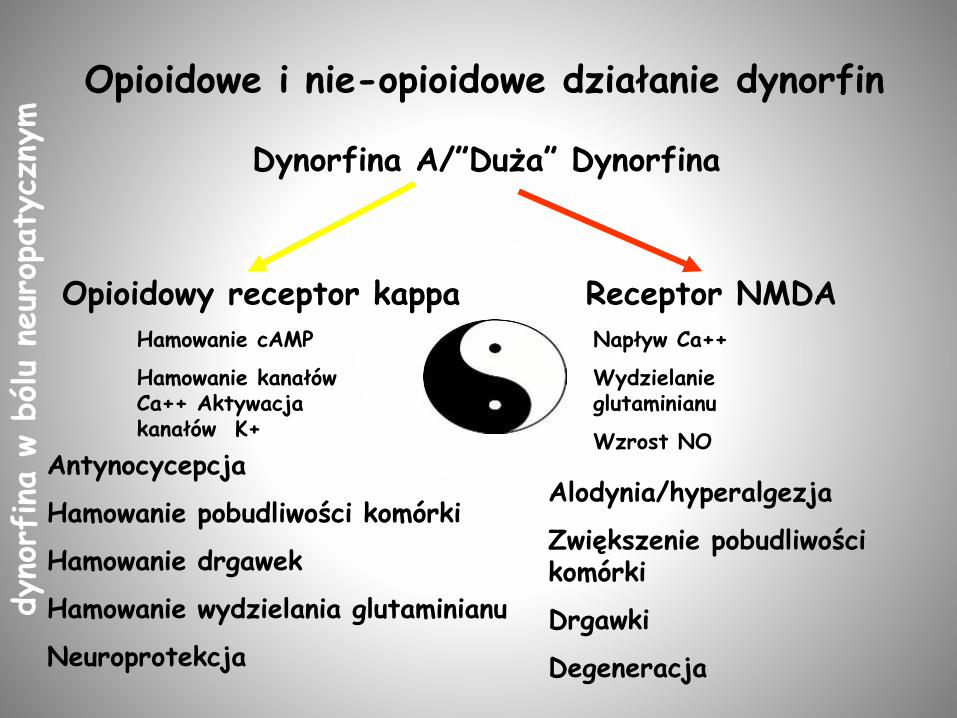
Dynorfina A/”Duża” Dynorfina
Opioidowy receptor kappa Receptor NMDAHamowanie cAMP
Hamowanie kanałów Ca++ Aktywacja kanałów K+
Napływ Ca++
Wydzielanie glutaminianu
Wzrost NO
Alodynia/hyperalgezja
Zwiększenie pobudliwości komórki
Drgawki
Degeneracja
Opioidowe i nie-opioidowe działanie dynorfin
Antynocycepcja
Hamowanie pobudliwości komórki
Hamowanie drgawek
Hamowanie wydzielania glutaminianu
Neuroprotekcja
dyno
rfina w
bólu
neur
opaty
czny
m
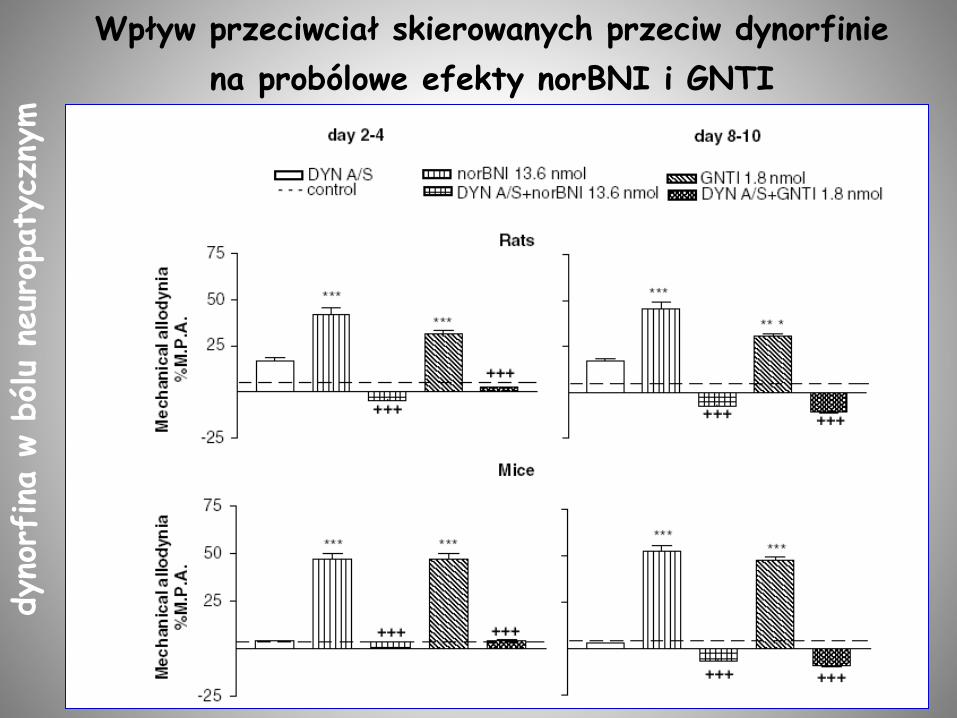
Wpływ przeciwciał skierowanych przeciw dynorfinie
na probólowe efekty norBNI i GNTI
dyno
rfina w
bólu
neur
opaty
czny
m
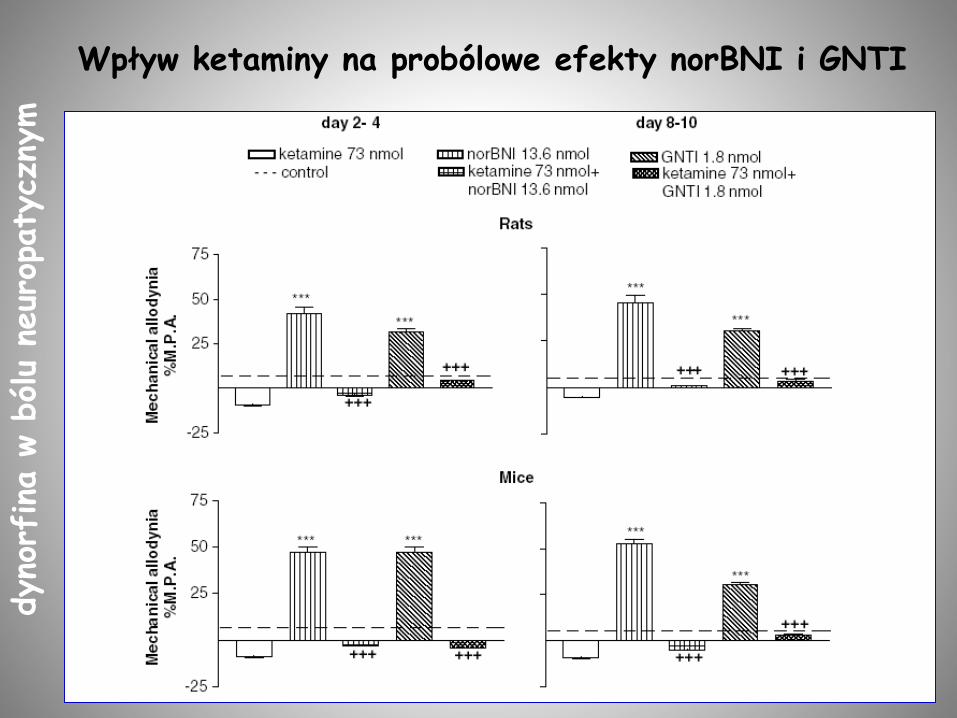
Wpływ ketaminy na probólowe efekty norBNI i GNTIdyno
rfina w
bólu
neur
opaty
czny
m
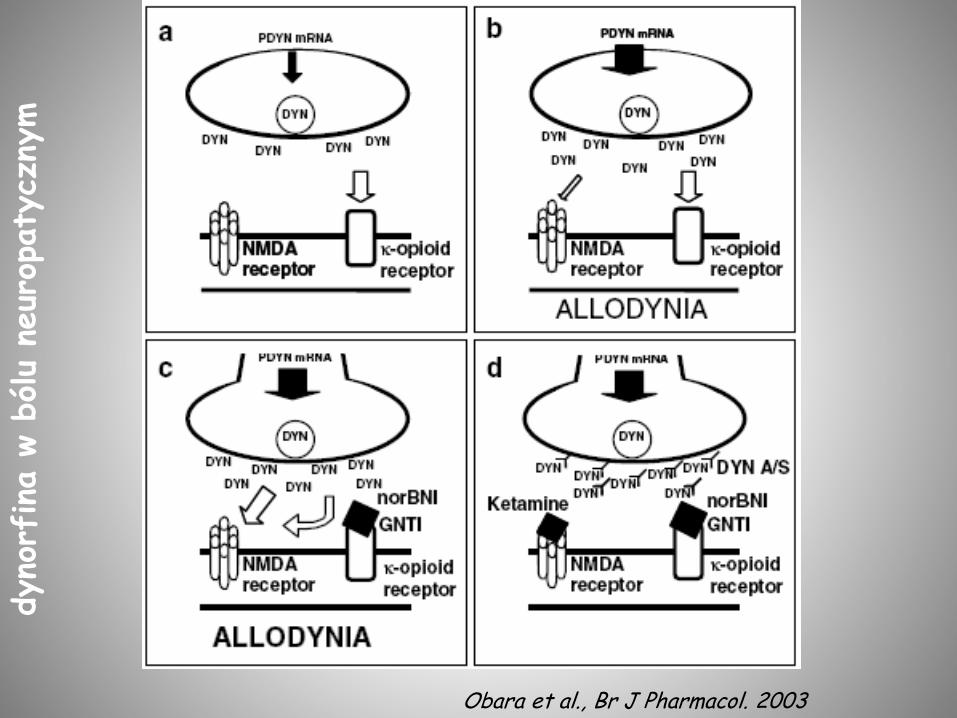
Obara et al., Br J Pharmacol. 2003
dyno
rfina w
bólu
neur
opaty
czny
m
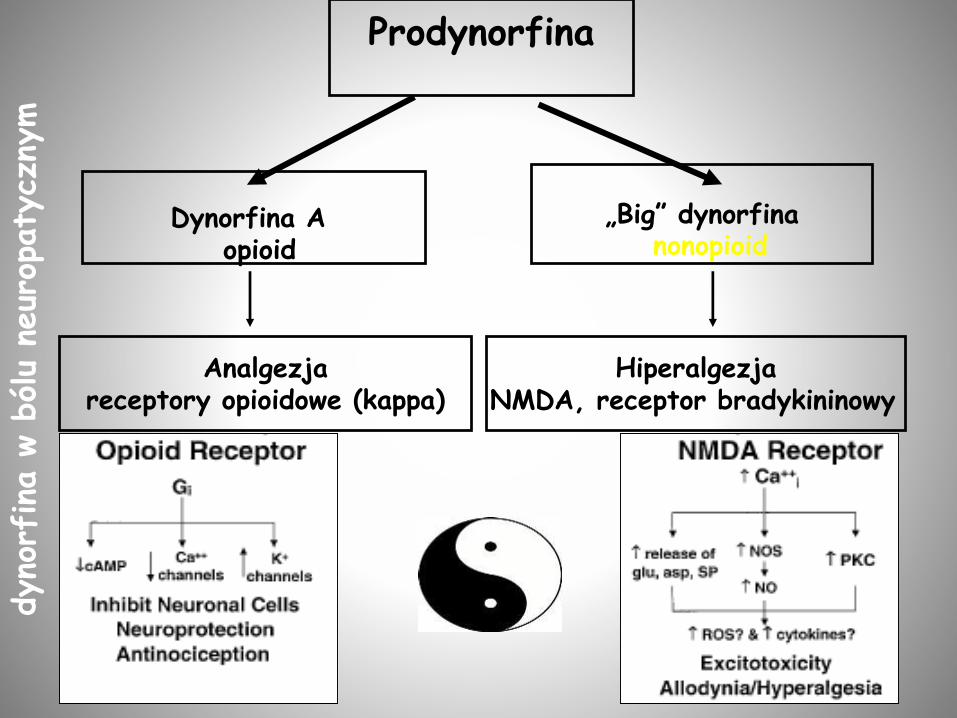
Prodynorfina
Analgezjareceptory opioidowe (kappa)
Dynorfina A opioid
„Big” dynorfina
HiperalgezjaNMDA, receptor bradykininowy
nonopioid
dyno
rfina w
bólu
neur
opaty
czny
m
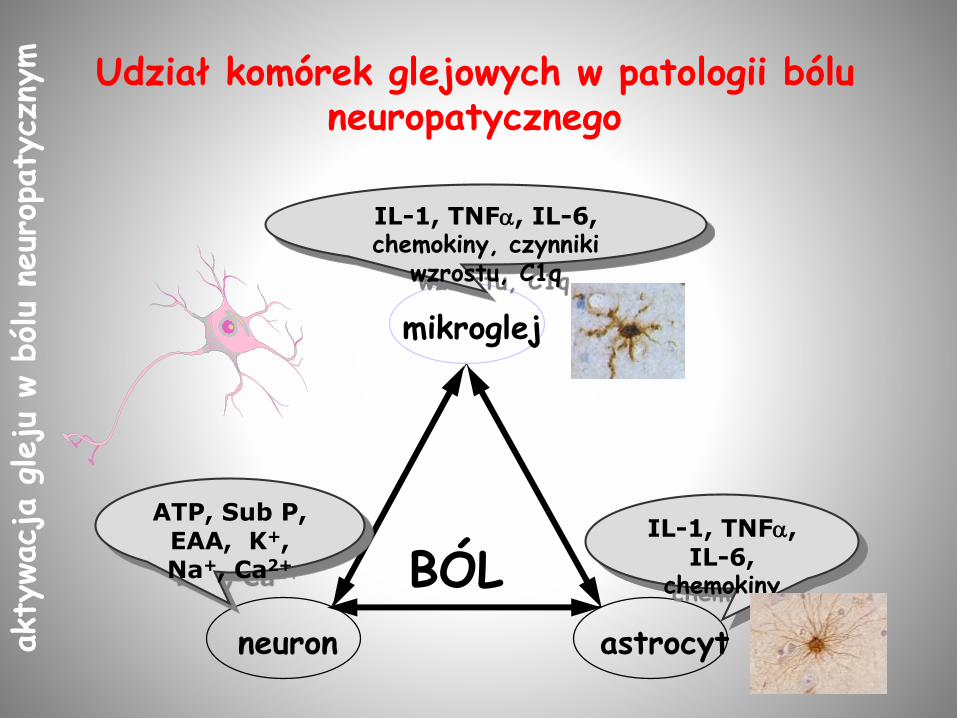
BÓL
astrocytneuron
mikroglej
ATP, Sub P, EAA, K+, Na+, Ca2+
IL-1, TNF, IL-6, chemokiny, czynniki
wzrostu, C1q
IL-1, TNF, IL-6,
chemokiny
Udział komórek glejowych w patologii bólu neuropatycznego
akty
wacj
a g
leju
w b
ólu
neur
opaty
czny
m
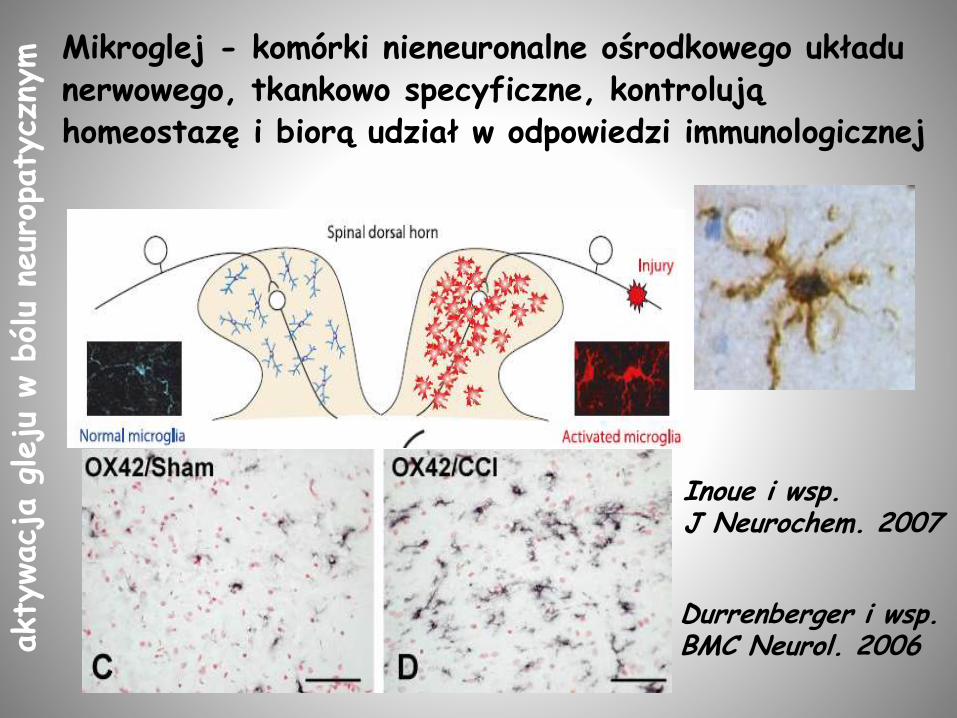
Mikroglej - komórki nieneuronalne ośrodkowego układu nerwowego, tkankowo specyficzne, kontrolują homeostazę i biorą udział w odpowiedzi immunologicznej
Durrenberger i wsp. BMC Neurol. 2006
Inoue i wsp.J Neurochem. 2007
akty
wacj
a g
leju
w b
ólu
neur
opaty
czny
m
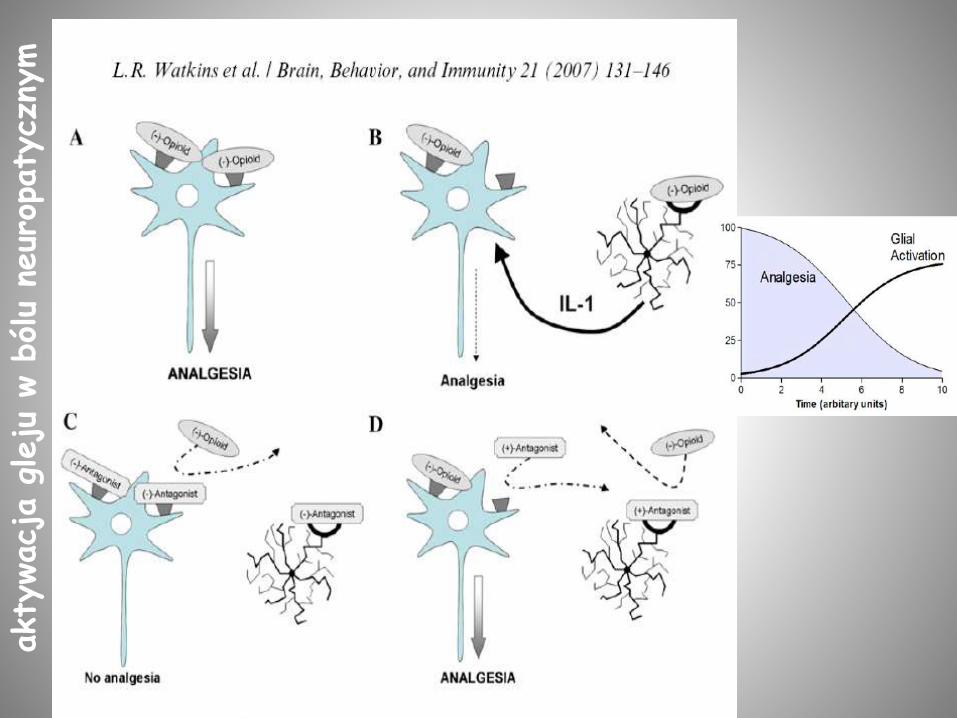
akty
wacj
a g
leju
w b
ólu
neur
opaty
czny
m

http://www.anzca.edu.au/fpm/events/fpm-events/activated-glia-as-culprits-in-opposing-opioid-analgesia-driving-tolerance.html
akty
wacj
a g
leju
w b
ólu
neur
opaty
czny
m

http://www.anzca.edu.au/fpm/events/fpm-events/activated-glia-as-culprits-in-opposing-opioid-analgesia-driving-tolerance.html
akty
wacj
a g
leju
w b
ólu
neur
opaty
czny
m

Zakład Farmakologii Bólu, Instytut Farmakologii, PANKraków
Pracownia Patofizjologii Bólu







![PDF WLA CUN 14 15 [tryb zgodności] - pum.edu.pl · Ośrodkowy układ nerwowy otoczony jest włóknistymi błonami – oponami mózgowo-rdzeniowymi. Opona miękka ... Obwodowy układ](https://static.fdocuments.pl/doc/165x107/5c78725d09d3f23a068ccccf/pdf-wla-cun-14-15-tryb-zgodnosci-pumedupl-osrodkowy-uklad-nerwowy-otoczony.jpg)