
Udział bakteryjnych dioksygenaz z rodziny AlkB w naprawie ... Krowisz praca... · Gen alkB jest...
83
Szkoła Główna Gospodarstwa Wiejskiego w Warszawie Wydział Rolnictwa i Biologii Beata Krowisz Numer albumu 118999 Udział bakteryjnych dioksygenaz z rodziny AlkB w naprawie egzocyklicznych uszkodzeń DNA The role of bacterial AlkB dioxygenases in repair of exocylic DNA adducts Praca magisterska na kierunku Biologia Praca wykonana pod kierunkiem dr Agnieszki Maciejewskiej Zakład Biologii Molekularnej IBB PAN Warszawa, rok 2010
Transcript of Udział bakteryjnych dioksygenaz z rodziny AlkB w naprawie ... Krowisz praca... · Gen alkB jest...

Szkoła Główna Gospodarstwa Wiejskiego
w Warszawie
Wydział Rolnictwa i Biologii
Beata Krowisz Numer albumu 118999
Udział bakteryjnych dioksygenaz z
rodziny AlkB w naprawie
egzocyklicznych uszkodzeń DNA
The role of bacterial AlkB dioxygenases in repair of exocylic
DNA adducts
Praca magisterska
na kierunku Biologia
Praca wykonana pod kierunkiem
dr Agnieszki Maciejewskiej
Zakład Biologii Molekularnej IBB PAN
Warszawa, rok 2010

2

3
Pragnę serdecznie podziękować
dr Agnieszce Maciejewskiej
za promotorska opiekę, teoretyczne i praktyczne zapoznanie mnie z tematem,
pomoc i wsparcie podczas pisania niniejszej pracy magisterskiej
prof. dr hab. Jarosławowi Kuśmierkowi
za wsparcie naukowe i umożliwienie mi wykonania części doświadczalnej w Zakładzie
Biologii Molekularnej IBB PAN

4
Praca została zrealizowana w ramach polsko-norweskiego grantu nr PNRF-143-AI-
1/07.
The AlkB protein and its eukaryotic homologues – the role in DNA repair and the
possible role in cancer etiology and target in cancer therapy (Białko AlkB i jego
eukariotyczne homologi – rola w naprawie DNA i potencjalny udział w etiologii
nowotworów oraz jako cel w terapii przeciwnowotworowej).

5
Oświadczenie promotora pracy
Oświadczam, że niniejsza praca została przygotowana pod moim kierunkiem i
stwierdzam, że spełnia ona warunki do przedstawienia jej w postępowaniu o nadanie
tytułu zawodowego.
Data ..................................... Podpis promotora pracy . ............................................
Oświadczenie autora pracy
Świadom odpowiedzialności prawnej oświadczam, że niniejsza praca dyplomowa
została napisana przeze mnie samodzielnie i nie zawiera treści uzyskanych w sposób
niezgodny z obowiązującymi przepisami.
Oświadczam również, że przedstawiona praca nie była wcześniej przedmiotem procedur
związanych z uzyskaniem tytułu zawodowego w wyższej uczelni.
Oświadczam ponadto, że niniejsza wersja pracy jest identyczna z załączoną wersją
elektroniczną.
Data ..................................... Podpis autora pracy . ..................................................

6

7
Streszczenie
Udział bakteryjnych dioksygenaz z rodziny AlkB w naprawie egzocyklicznych
uszkodzeń DNA
Gen alkB jest częścią systemu odpowiedzi adaptacyjnej na środki alkilujące bakterii
Escherichia coli. Białko AlkB jest dioksygenazą zależną od α-ketoglutaranu i Fe (II).
Usuwa grupy metylowe z m3C i m
1A oraz mostki etenowe z cytozyny
i adeniny, pozostawiając natywne DNA. Homologi genu alkB znaleziono u wielu
organizmów, także u człowieka.
Badano właściwości naprawcze białka AlkB i jego homologów: SA-1A i SA-2B,
występujących u Streptomyces avermitilis. Ustalono optymalne warunki naprawy
3,N4-α-hydroksyetanocytozyny i 3,N
4-α-hydroksypropanocytozyny przez AlkB
i SA-1A. Stwierdzono, że SA-1A nie naprawia 1,N6etenoadeniny i 3N
4etenocytozyny
natomiast SA-2B nie wykazuje aktywności wobec żadnego z badanych adduktów.
Słowa kluczowe — dioksygenaza, AlkB, modyfikacje DNA, addukty egzocykliczne,
naprawa DNA
Summary
The role of bacterial AlkB dioxygenases in repair of exocylic DNA adducts
The AlkB gene is a part of the system of adaptive response to alkylating agents in
Escherichia coli. AlkB is a member of the superfamily of α-ketoglutarate- and iron-
dependent dioxygenases. AlkB removes methyl groups from m3C and m
1A, and etheno
bridges from εC and εA, restoring native DNA structure. Analogous alkB genes were
found in various organisms including humans.
Repair activity of AlkB and its homologues from Streptomyces avermitilis: SA-1A and
SA-2B was studied. Optimal conditions for AlkB and SA-1A repair of
3,N4-α-hydroxyethanocytosine and 3,N
4-α-hydroxypropanocytosine were established.
SA-1A does not repair 3,N4-ethenocytosine and 1,N
6-ethenoadenine. SA-2B activity
against the studied adducts was not observed.
Keywords — dioxygenase, AlkB, DNA modifications, exocyclic adducts, DNA repair

8

9
Spis treści
I Wstęp ........................................................................................................................ 13
I.1 Czynniki uszkadzające DNA ............................................................................... 13
I.1.1. Chlorek winylu i produkty jego metabolizmu ............................................... 13
I.1.2. Czynniki alkilujące ...................................................................................... 14
I.1.3. Czynniki oksydacyjne .................................................................................. 15
I.1.4. Produkty peroksydacji lipidów ..................................................................... 16
I.1.5. Egzocykliczne addukty zasad DNA.............................................................. 18
I.1.5.1. Etenoaddukty ......................................................................................... 18
I.1.5.2. Propanoaddukty ..................................................................................... 19
I.2. Naprawa uszkodzonego DNA ............................................................................ 20
I.2.1. Rola białka AlkB w naprawie uszkodzeń DNA ............................................ 22
I.2.1.1. Budowa białka AlkB .............................................................................. 22
I.2.1.2. Mechanizm działania ............................................................................. 23
I.2.1.3. Bakteryjne homologi AlkB .................................................................... 24
I.2.1.4. Homologi AlkB występujące u ssaków .................................................. 26
II Cel pracy ................................................................................................................. 28
III.1. Materiały ......................................................................................................... 29
III.1.1. Odczynniki ................................................................................................ 29
III.1.2. Bufory ....................................................................................................... 30
III.1.2.1. Bufory stosowane do oczyszczania białka ........................................... 30
III.1.2.2. Bufory stosowane do reakcji modyfikacji pentametrów i ich naprawy
przez badane białka............................................................................................ 31
III.1.3. Pentamery ................................................................................................. 31
III.1.4. Enzymy ..................................................................................................... 31
III.1.5. Antybiotyki ............................................................................................... 31
III.1.6. Pożywki .................................................................................................... 31
III.1.7. Aparatura .................................................................................................. 32
III.1.7.1. Aparatura do oczyszczania białek ........................................................ 32
III.1.7.2. Aparatura do reakcji modyfikacji pentametrów i ich naprawy przez
badane białka ..................................................................................................... 32
III.1.8. Inne materiały ........................................................................................... 32
III.2. Metody ............................................................................................................ 33
III.2.1. Sporządzanie buforów ............................................................................... 33
III.2.1.1. Bufory stosowane do oczyszczania białka ........................................... 33
III.2.1.2. Bufory stosowane do reakcji modyfikacji pentametrów i ich naprawy
przez badane białka............................................................................................ 33
III.2.1.3. Bufor do chromatografii wysokociśnieniowej ...................................... 33
III.2.2. Transformacja bakterii szczepu E. coli BL21 (DE3) plazmidem
pET28a/pSA ......................................................................................................... 34
III.2.3. Ekspresja białka SA-2B w szczepie E. coli BL21 (DE3) niosącym plazmid
pET28a/pSA ......................................................................................................... 34
III.2.4. Dysrupcja komórek ................................................................................... 35
III.2.5. Oczyszczanie białka SA-2B ....................................................................... 35
III.2.6. Elektroforeza w żelu poliakrylamidowym .................................................. 36
III.2.7. Oznaczanie stężenia białka metodą Bradford ............................................. 36
III.2.8. Przygotowanie substratów do reakcji naprawy przez badane białka ........... 36

10
III.2.8.1. Otrzymywanie pentamerów zawierających etanocytozynę TT(εC)TT
oraz hydroksyetanocytozynę TT(HEC)TT..........................................................36
III.2.8.2. Otrzymywanie pentamerów zawierających hydroksypropanocytozynę
TT(HPC)TT .......................................................................................................37
III.2.8.3. Otrzymywanie substratu zawierającego etenoadeninę TT(εA)TT.........37
III.2.9. Badanie aktywności białka AlkB, SA-1A i SA-2B .....................................38
III.2.9.1. Zależność naprawy od pH....................................................................38
III.2.9.2. Zależność naprawy od stężenia α-KG ..................................................39
III.2.9.3. Zależność naprawy od stężenia jonów Fe (II) ......................................40
III.2.9.4. Zależność naprawy od czasu ................................................................40
IV Wyniki ...................................................................................................................42
IV.1. Oczyszczanie białka SA-2B .............................................................................42
IV.2. Chemiczna modyfikacja pentamerów TTCTT i TTATT ..................................43
IV.2.1. Modyfikacja pentamerów zawierających cytozynę (TTCTT) .....................43
IV.2.2. Modyfikacja pentamerów zawierających adeninę (TTATT).......................45
IV.3. Badanie naprawy modyfikowanych oligomerów przez białko AlkB in vitro ....46
IV.3.1. Naprawa TT(HEC)TT ...............................................................................47
IV.3.1.1. Zależność naprawy od pH ...................................................................47
IV.3.1.2. Zależność naprawy od stężenia Fe (II) .................................................48
IV.3.1.3. Zależność naprawy od stężenia α-KG ..................................................49
IV.3.1.4. Zależność naprawy od czasu................................................................50
IV.3.2. Naprawa TT(HPC)TT ...............................................................................51
IV.3.2.1. Zależność naprawy od pH ...................................................................51
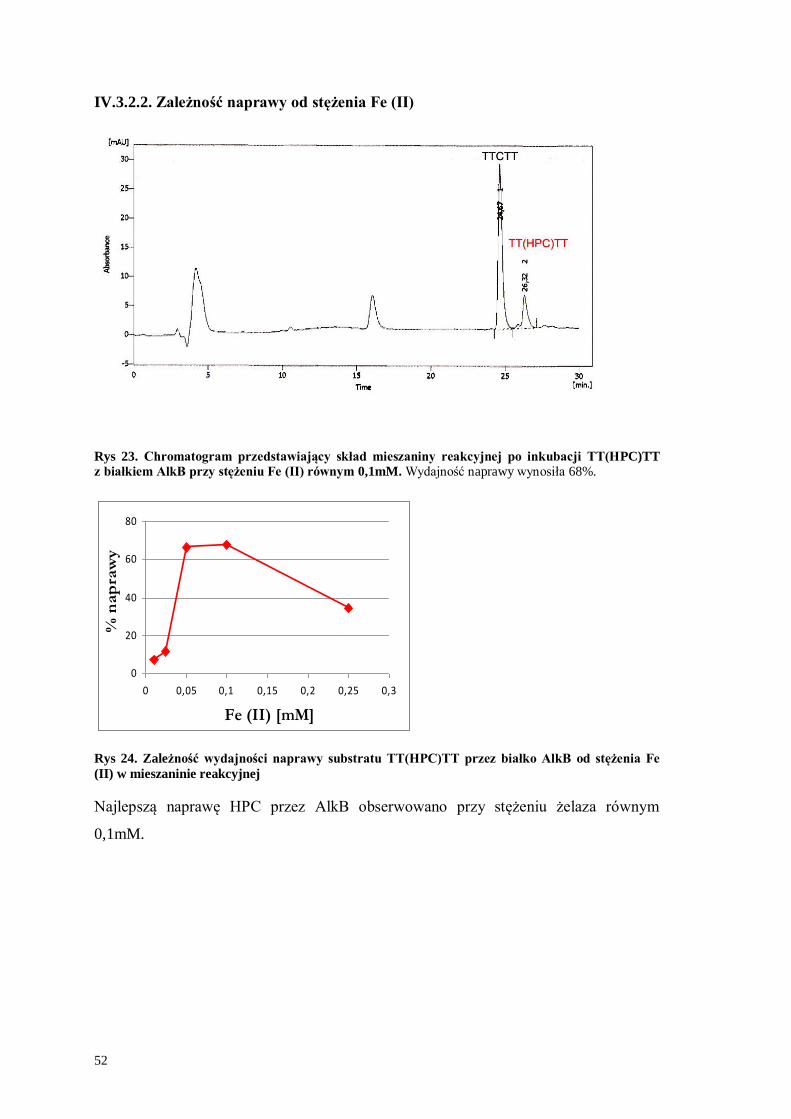
IV.3.2.2. Zależność naprawy od stężenia Fe (II) .................................................52
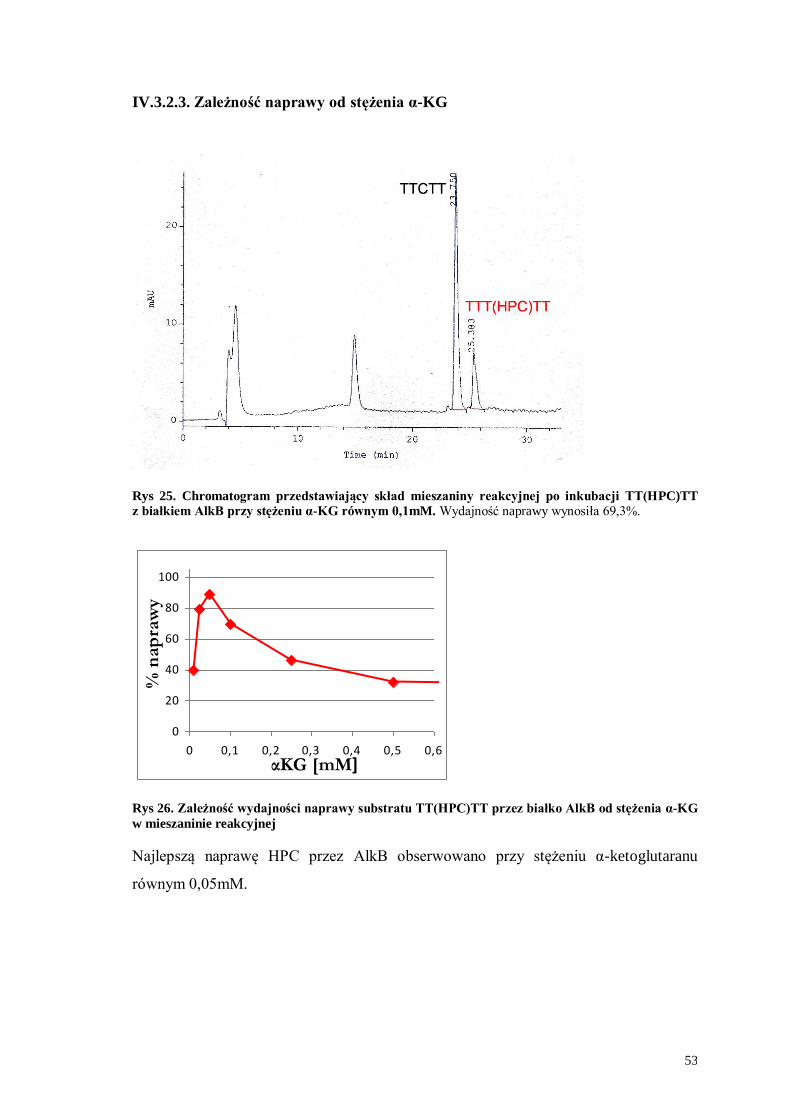
IV.3.2.3. Zależność naprawy od stężenia α-KG ..................................................53
IV.3.2.4. Zależność naprawy od czasu................................................................54
IV.4. Badanie naprawy in vitro modyfikowanych oligomerów przez bakteryjne
homologi białka AlkB: SA-1A i SA-2B ....................................................................55
IV.4.1. Białko SA-1A ............................................................................................55
IV.4.1.1. Zależność naprawy od pH ...................................................................57
IV.4.1.2. Zależność naprawy od stężenia α-KG ..................................................58
IV.4.1.3. Zależność naprawy od stężenia Fe (II) .................................................59
IV.4.2. Białko SA-2B ............................................................................................60
V Dyskusja ..................................................................................................................61
Aneks ..........................................................................................................................69
Spis literatury ..............................................................................................................71
Spis rysunków .............................................................................................................79
Spis tabel .....................................................................................................................81

11
Spis używanych skrótów
VC (ang. vinyl chloride) – chlorek winylu
CEO (ang. chloroethylene oxide) – tlenek chloroetylenu
CAA (ang. chloroacetaldehyde) – aldehyd chlorooctowy
CRA (ang. crotonaldehyde) – aldehyd krotonowy
ACR (ang. acrolein) – akroleina
HNE (ang. trans-4-hydroxy-2-nonenal) – 4-hydroksynonenal
MDA, MA (ang. malondialdehyde) – dialdehyd malonowy
ROS (ang. reactive oxygen species) – reaktywne formy tlenu
εA – 1,N6-etenoadenina
εC – 3,N4-etenocytozyna
HEC – 3,N4-α-hydroksyetanocytozyna
HPC – 3,N4-α-hydroksypropanocytozyna
TT(εA)TT, TεA – pentamer zawierający 4 tyminy i 1,N6-etenoadeninę (εA)
TT(εC)TT, TεC – pentamer zawierający 4 tyminy i 3,N4-etenocytozynę (εC)
TT(HEC)TT, THEC – pentamer zawierający 4 tyminy i 3,N4-α-hydroksyetanocytozynę
(HEC)
TT(HPC)TT, THPC – pentamer zawierający 4 tyminy i 3,N4-α-
hydroksypropanocytozynę (HPC)
EcAlkB – dioksygenaza AlkB pochodząca z E. coli, główny przedstawiciel rodziny
białek AlkB
SA-1A – homolog białka EcAlkB pochodzący z bakterii Streptomyces avermitilis
SA-2B – homolog białka EcAlkB pochodzący z bakterii Streptomyces avermitilis
HPLC (ang. high performance / high pressure liquid chromatography) –
wysokosprawna / wysokociśnieniowa chromatografia cieczowa
MeOH – metanol
TEAA – octan trójetyloaminy
IPTG – izopropylo-β-D-tiogalaktopiranozyd
SDS – dodecylosiarczan sodu
DTT – ditiotreitol

12

13
I Wstęp
Kwas deoksyrybonukleinowy jest podstawowym nośnikiem informacji
genetycznej, determinującym budowę, metabolizm, a nawet zachowanie organizmów
żywych. Uszkodzenia DNA odgrywają bardzo ważną rolę w procesach nowotworzenia
i starzenia się, dlatego też integralność i stabilność DNA jest niezbędna dla
prawidłowego funkcjonowania komórek. Wszystkie organizmy są wystawione na
działanie czynników mutagennych, zarówno pochodzenia egzo- jak i endogennego. Do
egzogennych czynników uszkadzających DNA zalicza się substancje występujące
w środowisku naturalnym (np.: aflatoksyna B1), substancje chemiczne powstałe podczas
procesów przemysłowych (np.: chlorek winylu), promieniowanie UV i jonizujące,
czynniki alkilujące (np.: metanosulfonian metylu - MMS), składniki dymu
papierosowego. Wśród czynnikami endogennych można wyróżnić aktywne formy tlenu
powstałe w procesie oddychania tlenowego w mitochondriach, czy też produkty
peroksydacji lipidów (Śliwiński i Błasiak 2005).
I.1 Czynniki uszkadzające DNA
I.1.1. Chlorek winylu i produkty jego metabolizmu
Chlorek winylu (VC) jest związkiem chemicznym stosowanym do produkcji
polimerów i kopolimerów wykorzystywanych w przemyśle tworzyw sztucznych.
Należy do rodziny halogenowych pochodnych etylenowych. U ludzi narażonych na
długotrwałe działanie VC stwierdzono liczne zmiany naczyniowe i kostne. Już w latach
czterdziestych ubiegłego wieku obserwowano przypadki powstawania nowotworów
u ludzi wystawionych na długotrwałą ekspozycję chlorkiem winylu, dlatego też
prowadzono badania w celu ustalenia mechanizmów kancerogennego i mutagennego
działania VC i jego metabolitów (Mroczkowska-Słupska i Kuśmierek 1994).
Chlorek winylu jest metabolizowany w wątrobie przy udziale cytochromu P450
II E 1. Pierwotnym produktem utleniania chlorku winylu przez ten cytochrom jest
tlenek chloroetylenu (CEO). Jest to związek wysoce nietrwały, szybko ulega
przegrupowaniu do aldehydu chlorooctowego (CAA) oraz hydrolizie do aldehydu
glikolowego, które są związkami trwałymi. CEO i CAA reagują z kwasami
nukleinowymi i białkami. Posiadają dwie aktywne grupy funkcyjne.
W przypadku CEO są to chlor i ugrupowanie epoksydowe, natomiast w przypadku
CAA chlor i grupa aldehydowa. W wyniku reakcji z kwasami nukleinowymi oba te

14
związki tworzą etenopochodne oraz wiązania krzyżowe. Wśród adduktów zasad DNA
powstałych w wyniku działania VC można wyróżnić 7-(2-oksoetylo)guaninę (7oeG),
N2,3-etenoguaninę (N
2,3-εG), 3,N
4-etenocytozynę (3,N
4εC, εC) oraz 1,N
6-etenoadeninę
(1,N6εA, εA) (Mroczkowska-Słupska i Kuśmierek 1994).
C C
H
Cl
H
H
HOCH2
C
O
H
ClCH2
C
O
H
C C
H
Cl
H
H
O
chlorek winylu (VC)
[O]
cytochrom P-450 2E1
aldehyd glikolowy tlenek chloroetylenu
( CEO)
aldehyd chlorooctowy
( CAA)
hydroliza przegrupowanie
Rys 1. Aktywacja metaboliczna chlorku winylu (Mroczkowska-Słupska i Kuśmierek 1994)
I.1.2. Czynniki alkilujące
Do czynników alkilujących należy grupa mutagenów i kancerogenów, które
modyfikują DNA poprzez alkilację. Niektóre z tych czynników są rozpowszechnione
w środowisku, inne natomiast to produkty normalnego metabolizmu. Związki alkilujące
mogą zakłócać replikację, transkrypcję czy też sygnalizować aktywację apoptozy.
U ssaków mogą być zaangażowane w kancerogenezę, choroby neurodegeneracyjne,
procesy starzenia (Nieminuszczy J, Grzesiuk E, 2007). Czynniki alkilujące mogą
dołączać grupy metylowe i etylowe do wszystkich dostępnych atomów tlenu i azotu
w zasadach DNA produkując dużą liczbę uszkodzeń. Dwie spośród 11
zidentyfikowanych modyfikacji DNA: 3-metyloadenina (m3A) oraz O
6-metyloguanina
(O6meG), są głównie odpowiedzialne za biologiczne efekty alkilacji (Singer 1976).
Rodzaj wywoływanych uszkodzeń zależy od czynnika, mechanizmu reakcji i struktury
drugorzędowej substratu (Nieminuszczy i Grzesiuk 2007).
Ze względu na mechanizm reakcji, czynniki alkilujące można podzielić na dwie
podgrupy. Czynniki z podgrupy SN1, takie jak N-metylo-N-nitrozomocznik (MNU)
i N-metylo-N’-nitro-N-nitrozoguanidyna (MNNG), wykorzystują mechanizm

15
jednocząsteczkowy. Natomiast te z podgrupy SN2 np.: metanosulfonian metylu (MMS),
jodek metylu (MeI), bazują na mechanizmie dwucząsteczkowym. Czynniki alkilujące
typu SN1 przyłączają addukty do atomów tlenu i azotu, natomiast czynniki typu SN2
alkilują tylko atom azotu. Głównymi modyfikacjami wywołanymi przez czynniki
metylujące w dwuniciowym DNA są: m7G, m
3A, O
6meG, natomiast m
1A, m
3C, m
7A
i O4meT występują rzadziej. W jednoniciowym DNA m
1A i m
3C występują częściej niż
w dsDNA. Wśród czynników metylujących najbardziej mutagenny jest O6meG.
Modyfikacja ta w dsDNA powoduje tranzycję GC→AT (Falnes i Rognes 2003).
Jak już wspomniano, czynniki alkilujące występują nie tylko w środowisku,
mogą też mieć pochodzenie endogenne. Do takich związków można zaliczyć
S-adenozylometioninę (SAM), która jest głównym biologicznym donorem grup
metylowych oraz prekursorem grup aminopropylowych wykorzystywanych
w biosyntezie poliamin (Lu 2000). Ponieważ metylacja odgrywa niezwykle ważną rolę
w procesach odbywających się w komórce (np.: w ekspresji genów), każda zmiana
stężenia SAM może powodować zaburzenia w funkcjonowaniu komórki.
Nieprawidłowości w metabolizmie SAM mogą być przyczyną chorób wątroby,
niektórych zaburzeń neurologicznych i kancerogenezy (Lutz 1990). SAM może
spontanicznie metylować DNA. Wykorzystuje mechanizm typu SN2 i powoduje
powstawanie głównie m7G i m
3A, rzadziej O
6meG ( Nieminuszczy i Grzesiuk 2007).
Wśród czynników alkilujących występujących w środowisku, bardzo duży
wpływ na zdrowie ludzi mają nitrozoaminy będące składnikami dymu papierosowego.
Do najbardziej kancerogennych należą 4-(metylonitrozoamino)-1-(3-pirydylo)-
1-butanon (NNK), 4-(metylonitrozoamino)-1-(3-pirydylo)-1-butanol (NNAL)
i N’-nitrozonornikotyna (NNN) (Hecht 2002).
I.1.3. Czynniki oksydacyjne
W wyniku wielu procesów metabolicznych zachodzących w komórce powstają
reaktywne formy tlenu takie jak: nadtlenek wodoru, tlen atomowy czy rodnik
hydroksylowy. Związki te mogą reagować z DNA i powodować pęknięcia nici,
uszkodzenia pojedynczej zasady, tworzenie adduktów objętościowych. Termin addukty
objętościowe określa m.in.: wiązania krzyżowe oraz addukty pierścieniowe (Moller
i Tallin 1998). Wśród modyfikacji zasad azotowych można wyróżnić takie, które są
potencjalnie mutagenne oraz takie, które blokują proces replikacji. Jednym

16
z oksydacyjnych uszkodzeń hamujących replikację DNA jest glikol tyminy.
Prawdopodobnie produkty częściowego rozpadu pierścienia purynowego (FapyA
i FapyG) również mogą zablokować proces replikacji (Wallace 1994).
I.1.4. Produkty peroksydacji lipidów
Peroksydacja lipidów to proces fizjologiczny, podczas którego dochodzi do
utlenienia wielonienasyconych kwasów tłuszczowych (PUFA), będących podstawowym
składnikiem błon biologicznych. Powstałe wodoronadtlenki ulegają dalszym
przemianom, w wyniku których powstają stabilne, nierodnikowe związki zawierające
grupy funkcyjne: aldehydowe, ketonowe, hydroksylowe, karboksylowe, peroksylowe
i epoksydowe oraz węglowodory m.in. alkany i alkeny (Marnett 1999). Aktywne
chemicznie związki powstające w wyniku tych procesów, przeważnie o charakterze
aldehydów, są w stanie przemieszczać się do jądra komórkowego, gdzie reagują
z DNA. Mogą też powstawać in situ w wyniku peroksydacji lipidów chromatynowych,
głównie fosfolipidów (Matulewicz i Przybyszewski 2004).
Wśród związków powstałych w wyniku peroksydacji lipidów można wymienić
dwualdehyd malonowy (MDA), akroleinę, aldehyd krotonowy, glioksal, tlenek etylenu,
eten, propen, trans-4-hydroksy-2-nonenal (HNE), 4-hydroksyheksenal (HHE).
Wszystkie działają mutagennie i kancerogennie (Matulewicz i Przybyszewski 2004,
Pastoriza Gallego i Sarasin 2003, Plastaras i wsp. 2000). Wśród nich dużą
toksycznością charakteryzuje się HNE. Najbardziej mutagenny jest MDA (Marnett
1999, Niederhofer i wsp. 2003).
MDA jest czynnikiem mutagennym dla komórek bakterii i ssaków oraz
kancerogennym dla szczurów (Przybyszewski i wsp. 2005). Reakcje MDA z zasadami
DNA, prowadzące do powstawania adduktów, są złożone, m.in.: z powodu procesów
polimeryzacji, którym ulega ten związek. W wyniku tych procesów w komórkach
pojawiają się dimeryczne i trimeryczne postacie MDA, które również reagują
z zasadami azotowymi. Reakcja MDA z deoksyguanozyną prowadzi do powstania
struktury pirymidopurynonu, określanej symbolem M1dG. W wyniku reakcji MDA
z deoksyadenozyną i deoksycytydyną powstają związki oksopropenylowe:
N6-(3-oksopropenyl)deoksyadenozyny oraz śladowe ilości N
6-(3-
oksopropenyl)deoksycytydyny (Marnett 2002, Marnett i wsp. 2003).

17
Rys 2. Powstawanie eteno- i propanoadduktów pod wpływem produktów peroksydacji lipidów
(Przybyszewski i wsp. 2005)
Produkty peroksydacji lipidów: aldehyd krotonowy, akroleina i HNE są
przekształcane do epoksyaldehydów, które modyfikują zasady DNA tworząc
etenoaddukty. W wyniku reakcji epoksyaldehydów z DNA powstają: N2,3
etenoguanina
1,N2etenoguanina, 1,N
6etenoadenina i 3,N
4etenocytozyna. Oddziaływanie akroleiny
z zasadami azotowymi powoduje powstanie 1,N2-etenoguaniny (Golding i wsp. 1996).
Pochodna aldehydu krotonowego, 3-epoksybutanal, prowadzi do powstania 1,N6-
etenoadenozyny, 3,N4-etenocytydyny, 1,N
2-etenoadenozyny (Nair i Offerman 1985).
Natomiast pochodna HNE, 2,3-epoksy-4-hydroksynonenal (EH), w wyniku
podstawienia grup hydroksyheptanowych do guaniny i adeniny tworzy 1,N2- i N
2,3-
etenoguaninę oraz 1,N6-etenoadeninę (Sodum i Chung 1989, 1991).

18
I.1.5. Egzocykliczne addukty zasad DNA
Egzocykliczne addukty zasad DNA charakteryzują się dodatkowym
pierścieniem przy atomie azotu grupy aminowej. Uszkodzenia te, jeżeli nie są
naprawione, mogą wywoływać u ssaków różnego rodzaju choroby chroniczne
i nowotworowe. Wiele z adduktów egzocyklicznych ma pochodzenie endogenne,
prawdopodobnie jako rezultat reakcji produktów peroksydacji lipidów z zasadami DNA
(Borys-Brzywczy i wsp. 2005).
I.1.5.1. Etenoaddukty
Etenoaddukty należą do grupy egzocyklicznych adduktów zasad DNA i są
tworzone przez różne czynniki egzo- i endocykliczne takie jak chlorek winylu, aldehyd
chlorooctowy, produkty peroksydacji lipidów (Maciejewska i wsp. 2010). Powstają
w wyniku utworzenia dodatkowego pierścienia imidazolowego w strukturze cząsteczek
zasad DNA. Możliwe jest utworzenie 4 etenoadduktów: 1,N6-etenoadeniny (εA);
3,N4-etenocytozyny (εC); N
2,3-eteonoguaniny (N
2,3-εG) oraz 1,N
2-etenoguaniny
(1,N2-εG) (Barbin 2000). W reakcjach aldehydu chlorooctowego lub tlenku
chloroetylenu z adeniną i cytozyną zaobserwowano powstawanie uwodnionych form
pośrednich takich jak 3,N4-α-hydroksyetanocytozyna (HEC) (Krzyżosiak i wsp. 1979,
Kuśmierek i Singer 1982, Guengerich i Raney 1992). W DNA formy te są
przekształcane do pochodnych etenowych. W dwuniciowym DNA etenopochodne są
stosunkowo trwałe. εA może być jednak degradowane do 4-amino-5(-imidazol-2-
yl)imidazolu (β) (Yip i Tsou 1973). Natomiast 1,N2-εG może być przekształcona do
N1-2-(oksoetyl)guaniny (Akasaka i Guengerich 1999).
Pierwsze badania nad właściwościami mutagennymi etenoadduktów były
prowadzone w 1981 roku w trzech niezależnych laboratoriach i wykazały, że εA może
powodować błędy w przyłączaniu guaniny lub adeniny. Natomiast εC może wpływać
na niewłaściwe przyłączanie tyminy lub adeniny (Barbin i wsp. 1981, Hall i wsp. 1981,
Spengler i Singer 1981). Kolejne badania wykazały, że u Escherichia coli
i w komórkach nerki małpy εA powoduje tranzycję A→G oraz rzadziej transwersję
A→T i A→C. εC wywołuje w tych komórkach substytucje C→A i C→T, rzadziej
C→G. Dodatkowo εC powoduje jedno- i dwuzasadowe delecje (Basu i wsp. 1993,
Shibutani i wsp. 1996, Palejwala i wsp. 1993). HEC, podobnie jak εC, prowadzi do
tranzycji C→T u E.coli (Jacobsen i wsp. 1989, Borys i wsp. 1994). N2,3-εG in vitro
i w komórkach E.coli może wywoływać tranzycje G→A (Singer i wsp. 1983),
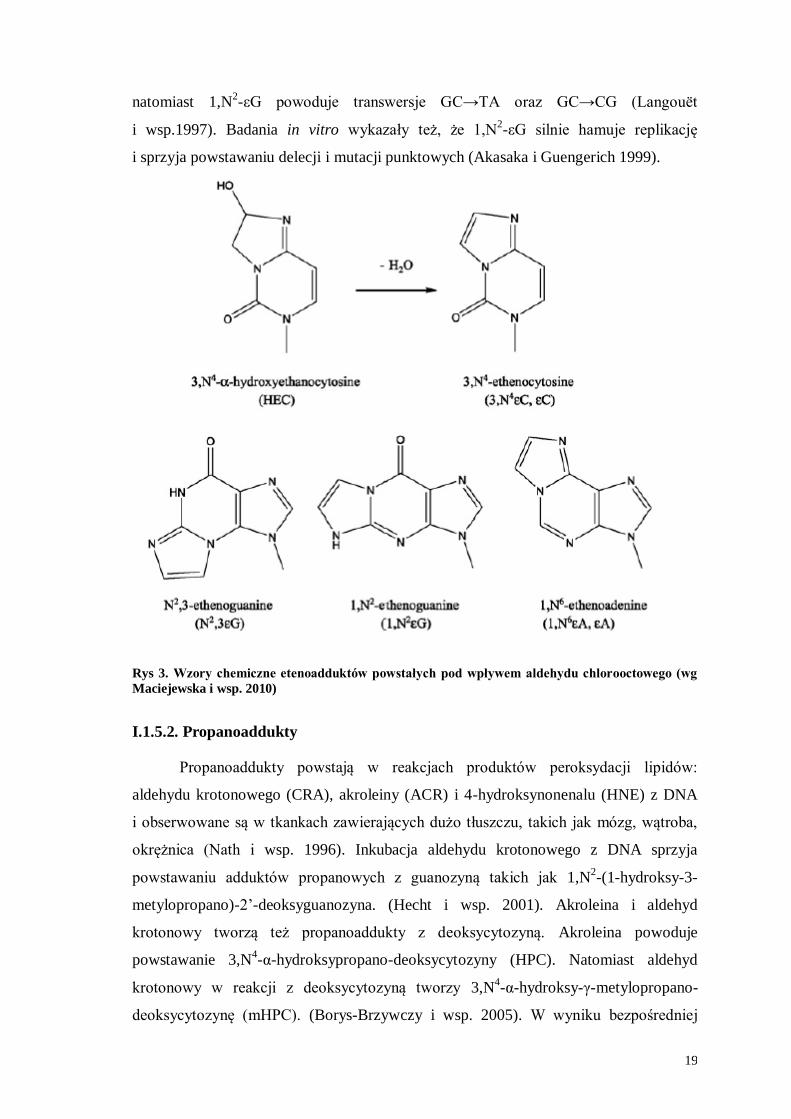
19
natomiast 1,N2-εG powoduje transwersje GC→TA oraz GC→CG (Langouët
i wsp.1997). Badania in vitro wykazały też, że 1,N2-εG silnie hamuje replikację
i sprzyja powstawaniu delecji i mutacji punktowych (Akasaka i Guengerich 1999).
Rys 3. Wzory chemiczne etenoadduktów powstałych pod wpływem aldehydu chlorooctowego (wg
Maciejewska i wsp. 2010)
I.1.5.2. Propanoaddukty
Propanoaddukty powstają w reakcjach produktów peroksydacji lipidów:
aldehydu krotonowego (CRA), akroleiny (ACR) i 4-hydroksynonenalu (HNE) z DNA
i obserwowane są w tkankach zawierających dużo tłuszczu, takich jak mózg, wątroba,
okrężnica (Nath i wsp. 1996). Inkubacja aldehydu krotonowego z DNA sprzyja
powstawaniu adduktów propanowych z guanozyną takich jak 1,N2-(1-hydroksy-3-
metylopropano)-2’-deoksyguanozyna. (Hecht i wsp. 2001). Akroleina i aldehyd
krotonowy tworzą też propanoaddukty z deoksycytozyną. Akroleina powoduje
powstawanie 3,N4-α-hydroksypropano-deoksycytozyny (HPC). Natomiast aldehyd
krotonowy w reakcji z deoksycytozyną tworzy 3,N4-α-hydroksy-γ-metylopropano-
deoksycytozynę (mHPC). (Borys-Brzywczy i wsp. 2005). W wyniku bezpośredniej

20
reakcji HNE z DNA powstaje głównie propanowy addukt guaniny podstawiony
łańcuchem alifatycznym (Douki i wsp. 2004). Badania in vitro wykazały, że HNE
oddziałuje z deoksynukleozydami tworząc propano- i etenocykliczne addukty
posiadające boczny łańcuch hydroksyalkilowy. Powodują one wzrost częstości mutacji
oraz hamują syntezę DNA in vitro u E. coli (Kowalczyk i wsp. 2004).
Rys 4. Wzory chemiczne propanoadduktów akroleiny i aldehydu krotonowego z deoksycytozyną
(na podstawie Borys-Brzywczy i wsp. 2005)
I.2. Naprawa uszkodzonego DNA
DNA wszystkich żywych organizmów jest nieustannie modyfikowane przez
związki pochodzenia endo- i egzogennego. Aby zachować prawidłowe funkcjonowanie
komórek, wykształcone zostały różne mechanizmy naprawy DNA, które zapobiegają
kumulacji szkodliwych mutacji w komórce.
U bakterii Escherichia coli można wyróżnić cztery mechanizmy naprawy
uszkodzeń kwasów nukleinowych spowodowanych przez związki alkilujące. Pierwszy
z nich to BER (ang. base excision repair), czyli naprawa przez wycinanie zasad.
Kolejnym jest NER (ang. nucleotide excision repair), który polega na wycinaniu
uszkodzonych nukleotydów. Trzeci mechanizm, MMR (ang. mismatch repair),

21
naprawia błędnie sparowane zasady. Ostatni sposób to naprawa bezpośrednia przez
odwrócenie np.: przez metylotransferazy (Nieminuszczy i Grzesiuk 2007).
Większość oksydacyjnych modyfikacji DNA np.: zmiany struktury chemicznej
zasady, jest naprawiana z wykorzystaniem mechanizmu naprawy przez wycinanie zasad
(BER) (Gros i wsp. 2003). Wycięcie zmodyfikowanej zasady polega na hydrolizie
wiązania N-glikozydowego pomiędzy zmienioną chemicznie zasadą a cukrem.
N-glikozylaza rozpoznaje uszkodzoną zasadę i przecina wiązanie β-N-glikozydowe
między uszkodzoną zasadą a cząsteczką deoksyrybozy, tworząc miejsce
apurynowe/pirymidynowe. Następnie endonukleaza AP rozpoznaje to miejsce i nacina
DNA w kierunku 5' od miejsca AP tworząc wolny koniec 3'-OH. Polimeraza DNA,
Pol I, wydłuża nić DNA od wolnego końca 3'-OH wykorzystując aktywność
egzonukleazy do zastąpienia nukleotydu z uszkodzoną zasadą oraz kilku następnych.
Na końcu nowa nić jest łączona ze starą za pomocą ligazy DNA (Roszkowski 2002,
Krwawicz i wsp. 2007).
Naprawa przez wycięcie nukleotydu (NER) jest bardziej złożonym procesem niż
BER oraz wymaga ATP jako źródła energii. Jest też mniej specyficznym systemem niż
BER (Wood 1996; Sancar 1996). Fragment DNA, który zawiera uszkodzenie jest
nacinany po obu stronach przez nukleazę, a następnie usuwany. U bakterii jest to
fragment składający się z 12-13 nukleotydów. W komórkach eukariotycznych fragment
ten ma długość 27-29 nukleotydów (Pietrzykowska i Krwawicz 1999; Ma i wsp. 1995).
System naprawy błędnie sparowanych zasad azotowych (MMR) usuwa głównie
błędy, które powstały podczas replikacji DNA i nie zostały naprawione przez
polimerazę DNA oraz niewłaściwe pary zasad tworzące się w wyniku rekombinacji
DNA oraz spontanicznej lub indukowanej deaminacji, utleniania bądź metylacji zasad
azotowych (Modrich i Lahue 1996).
Do naprawy DNA, oprócz skomplikowanych kompleksów naprawczych,
komórka może wykorzystywać także proste mechanizmy naprawy kwasów
nukleinowych, które nie naruszają ich integralności. Należy do nich bezpośrednia
naprawa przez odwrócenie, w której bierze udział wysoce specyficzne białko. Białko to
eliminuje dodatkowe grupy atomów bezpośrednio w miejscu ich przyłączenia. Dzieje
się tak na przykład w przypadku dioksygenazy AlkB naprawiającej 3-metylocytozynę
oraz 1-metyloadeninę (Falnes i wsp. 2002).

22
I.2.1. Rola białka AlkB w naprawie uszkodzeń DNA
Gen alkB został odkryty ponad 20 lat temu podczas izolacji mutantów E. coli ze
zwiększoną wrażliwością na czynniki alkilujące (Hataoka i wsp. 1983). Gen ten
wchodzi w skład systemu odpowiedzi adaptacyjnej na czynniki alkilujące u E. coli wraz
z genami ada, alkA i aidB. Białka Ada, Alka i AlkB naprawiają uszkodzone zasady
DNA przy użyciu różnych mechanizmów. Funkcja AidB nie została jeszcze ostatecznie
ustalona (Nieminuszczy i Grzesiuk 2007). Białko AlkB preferuje jednoniciowe DNA
jako substrat (Falnes i wsp. 2004). Badania biochemiczne wykazały, że najlepszymi
substratami dla AlkB są 3-metylocytozyna (m3C) oraz 1-metyloadenina (m
1A) (Dinglay
i wsp. 2000). Analiza bioinformatyczna wykazała, że białko AlkB należy do rodziny
dioksygenaz zależnych od Fe(II) i α-ketoglutaranu (Aravind i Koonin 2001).
I.2.1.1. Budowa białka AlkB
Badania nad strukturą przestrzenną białka AlkB wykazały, że składa się ono
z 216 aminokwasów i zawiera trzy ważne domeny. Są to:
C-terminalna domena dioksygenazowa, część katalityczna (aa 90-216)
Sekwencja NRL rozpoznająca nukleotyd (aa 46-89),
N-terminalna domena (aa 1-45) (Yang i wsp. 2008; Yu i wsp. 2006).
Rys 5. Struktura kompleksu białka AlkB z Fe(II), α-ketoglutaranem i metylowanym
trójnukleotydem (Yu i wsp. 2006). Niebieskim kolorem oznaczona jest domena rozpoznająca nukleotyd, zielonym rdzeń katalityczny, a żółtym fragment N-końcowy.

23
Wszystkie homologi AlkB posiadają motywy i reszty niezbędne dla
enzymatycznej aktywności (motyw HXD, pojedyncze H i motyw RXXXXR). U AlkB
motywy te wyglądają następująco: H131XD133………H187………R204YNLTFR210. Dwie
histydyny i kwas asparaginowy biorą udział w wiązaniu żelaza, natomiast pierwsza
arginina oddziałuje z α-KG (Wei i wsp. 1996; Aas i wsp. 2003; Kurowski i wsp. 2003).
I.2.1.2. Mechanizm działania
Białko AlkB usuwa alkilacyjne oraz etenowe uszkodzenia zasad DNA. W obu
przypadkach mechanizm naprawy odbywa się w dwóch etapach.
W przypadku naprawy modyfikacji alkilowych pierwszy etap polega na
hydroksylacji wiązania C-H w grupie alkilowej dołączonej do zasady. Wiąże się to
z oksydatywną dekarboksylacją α-ketoglutaranu, w wyniku czego następuje uwolnienie
cząsteczki bursztynianu i dwutlenku węgla. Jeden z atomów cząsteczki tlenu zostaje
wbudowany we fragment alkilowy jako grupa hydroksylowa, drugi natomiast stanowi
cześć grupy karboksylowej bursztynianu (Schofield i Hang 1999; Prescott i Lloyd 2000;
Ryle i Hausinger 2002). W ten sposób białko AlkB bezpośrednio przekształca
3-metylocytozynę i 1-metyloadeninę w cytozynę i adeninę z jednoczesnym
uwolnieniem utlenionej grupy metylowej w postaci formaldehydu. Produktami
przejściowymi w tych reakcjach są niestabilne 3-hydroksymetylocytozyna
i 1-hydroksymetyloadenina (Trewick i wsp. 2002) (Rys. 6).
Rys 6. Mechanizm naprawy uszkodzeń alkilacyjnych przez białko AlkB (Begley i Samson 2003)
W podobny sposób naprawiane są uszkodzenia etenowe. W pierwszej kolejności
mostek eteonowy utleniany jest do epoksydu, który w wyniku addycji cząsteczki wody

24
tworzy glikol. Następnie powstały alkohol uwalnia się spontanicznie w postaci
glioksalu z odtworzeniem natywnej formy zasady (Delaney i wsp. 2005) (Rys. 6).
Rys 6. Mechanizm naprawy uszkodzeń etenowych przez białko AlkB (Delaney i wsp. 2005)
I.2.1.3. Bakteryjne homologi AlkB
Van den Born i współpracownicy na podstawie analizy sekwencji bakteryjnych
białek AlkB opracowali drzewo filogenetyczne i wyodrębnili 4 grupy: 1A, 1B, 2A i 2B.
Dowiedziono, że białka z grup 1A i 2B posiadają kilka tych samych konserwowanych
ewolucyjnie aminokwasów co białka 1B, znajdujących się w regionie odpowiadającym
domenie NRL. Pozwala to przypuszczać, że te 3 grupy białek mogą oddziaływać z tym
samym substratem. (van den Born i wsp. 2009). W białkach grup 1B i 2B w domenie
N-terminalnej występują takie same konserwowane sekwencje. Również w obrębie
domeny NRL występują wspólne dla grup 1B i 2B konserwowane ewolucyjnie
aminokwasy. Nie wykazano natomiast wspólnych konserwowanych sekwencji dla
białek z grup 1A i 2A. W trakcie badań nie ujawniono podobieństwa sekwencji domeny
N-terminalnej białek z grup 1A i 1B. Przypuszcza się jednak, że zawierają one kilka
kluczowych reszt w obrębie domeny NRL, łącznie z resztami Trp69 i Tyr76. (Yu i wsp.
2006) Wykazano też, że białka z grup 1A, 1B oraz 2B uczestniczą głównie w naprawie
DNA, podczas gdy funkcją białek z grupy 2A jest przede wszystkim modyfikacja tRNA
(van den Born i wsp. 2009).
Grupa 1A
Białka z tej grupy są szeroko rozpowszechnione w królestwie bakterii,
zwłaszcza u Proteobacteria. W grupie tej znajduje się białko AlkB E. coli, a także białka
SA-1A i SC-1A występujące u bakterii z rodzaju Streptomyces. Białka te mają w 79%
identyczne sekwencje aminokwasowe i wykazują podobne właściwości. Przypuszcza
się, że grupa 1A zawiera białka o funkcji podobnej do białka modelowego dla rodziny
AlkB – EcAlkB (van den Born i wsp. 2009).

25
Grupa 1B
Białka z grupy 1B występują u β- i γ-proteobacteria, a także u cyanobacteria.
Wykazują podobieństwo do homologów AlkB występujących u kręgowców: ALKBH2
i ALKBH3, które posiadają podobną aktywność naprawczą do EcAlkB. W grupie tej
znajduje się białko XC-1B, występujące u Xanthomonas campestris i wykazujące
wysoką aktywność naprawczą. Analiza bioinformatyczna i badania doświadczalne
wykazały, że białka z grupy 1B posiadają aktywność naprawczą, a ich funkcja jest
zbliżona do EcAlkB (van den Born i wsp. 2009).
Grupa 2B
Białka z grupy 2B występują głównie u Actinobacteria. Wykazano również
występowanie tych białek u kilku patogenów roślinnych z rodzaju Xanthomonas
(γ-proteobacteria) i Burkholderia (β-proteobacteria). Bakterie te mogły nabyć geny
kodujące białka AlkB przez horyzontalny transfer genów od Actinobacteria, będących
w większości bakteriami glebowymi (Madigan i wsp. 2003). Analiza sekwencji
wykazała, że białka z grupy 1B i 2B posiadają kilka takich samych zakonserwowanych
reszt w regionie NRL. Białka z grupy 2B, takie jak: MT-2B, SC-2B, SA-2B i XC-2B
wykazują aktywność naprawczą. Ich specyficzność substratowa jest jednak dość
zróżnicowana. Białko MT-2B, w przeciwieństwie do XC-2B, wykazuje wysoką
aktywność w stosunku do uszkodzeń metylowych i niską w stosunku do etenoadduktów
(van den Born i wsp. 2009).
Grupa 2A
Białka z grupy 2A występują u proteobacteria takich jak: Agrobacterium,
Rickettsia i Rhizobium. Część z tych bakterii to patogeny roślin, które podczas inwazji
na gospodarza przenoszą fragmenty swojego DNA do komórek roślinnych.
U niektórych bakterii np.: Roseobacter denitrificans, Roseobacter etli, Sinorhizobium
meliloti, Sinorhizobium medicae, geny kodujące białka z grupy 2A znajdują się częściej
na plazmidach niż na chromosomach, co sugeruje że być może zostały one uzyskane
z genomu gospodarza (van den Born i wsp. 2009). Białka z grupy 2A wykazują dużą
homologię względem zwierzęcych i roślinnych białek ALKBH8 (Falnes i wsp. 2009).
Po analizach bioinformatycznych zasugerowano, że białka te są raczej zaangażowane
w modyfikację tRNA niż w naprawę DNA (van den Born i wsp. 2009).

26
Tabela 1. Rozmieszczenie bakteryjnych białek AlkB w kompletnie zsekwencjonowanych genomach
Liczba białek
AlkB
Grupa Liczba
gatunków
Przykłady rodzajów/gatunków bakterii
1
1
1
1
1
2
2 2
2
2
2
2
2
2 2
2
3
3
3
1A
1B
2A
2B
Razem
1A
1A+1B 1A+2A
1A+2B
1B
1B+2A
1B+2B
2A
2A+2B 2B
Razem
1A(2)+1B
1B
Razem
65
61
6
12
144
1
2 5
3
1
0
4
0
0 2
18
1
1
2
Escherichia, Salmonella, Bradyrhizobium
Shewanella, Burkholderia, Xylella
Agrobacterium, Rickettsia
Mycobacterium, Nocardia
Ralstonia metallidurans
Pseudomonas fluorescens Sinorhizobium, Rhizobium, Roseobacter
Streptomyces, Rhodococcus
Acaryochloris marina
-
Xanthomonas, Burkholderia enovorans
-
- Frankia
Burkholderia sp. 383
Flavobacterium johnsoniae
Razem 164
I.2.1.4. Homologi AlkB występujące u ssaków
Geny kodujące homologi białka AlkB wykryto nie tylko u bakterii, ale również
w DNA ssaków. U ssaków zidentyfikowano 9 homologów tego białka: ALKBH1-
ALKBH8 (Drablos i wsp. 2004; Aravind i Koonin 2001) oraz białko FTO, które jest
związane z otyłością (Gerken i wsp. 2007; Sanchez-Pulido i Andrade-Navarro 2007).
Ekspresja poszczególnych genów kodujących homologi AlkB nie jest taka sama
we wszystkich tkankach ludzkich i może wynikać z rozbieżności funkcji tych białek
(Lee i wsp. 2005). Zaobserwowano wysoki poziom hABH2 w nieproliferujących
tkankach wątroby, natomiast wysoka ekspresja hABH3 występowała w sercu, wątrobie,
prostacie i jądrach (Sedgwick i wsp. 2007).
Pierwszy ludzki homolog, hABH1, wykazuje 52% podobieństwo i jest w 23%
identyczne z EcAlkB (Mishina i He 2006). Jest funkcjonalnym mitochondrialnym
homologiem AlkB, który naprawia m3C w jednoniciowym DNA i RNA, ale nie
naprawia m1A, ani dsDNA. Aktywność naprawcza hABH1 jest niższa niż AlkB,
hABH2 i hABH3, ale nie można wykluczyć, że białko to ma jeszcze inne substraty
(Westbye 2008). hABH1 wykazuje też aktywność liazy wobec miejsc AP w DNA
(Muller i wsp. 2010).

27
Odkryto, że hABH2 jest odpowiedzialny głównie za naprawę m1A, natomiast ta
sama funkcja u hABH3 pozostaje niejasna (Ringvoll i wsp. 2006). hABH2 jako substrat
bardziej preferuje dwuniciowe DNA (dsDNA) niż jednoniciowe (ssDNA) oraz ma
niskie powinowactwo w stosunku do RNA. Tymczasem hABH3 i AlkB jest bardziej
aktywne w przypadku substratów ssDNA i ssRNA (Aas i wsp. 2003; Falnes i wsp.
2004). hABH2 i hABH3, oprócz tego że preferują różne substraty, mają też inną
lokalizację w komórce. hABH2 występuje tylko w jądrze komórkowym i uczestniczy
w naprawie DNA w pobliżu widełek replikacyjnych. Natomiast hABH3 występuje
w jądrze komórkowym i w cytoplazmie (Duncan i wsp. 2002; Aas i wsp. 2003).
W 2007 roku wielką niespodzianką było odkrycie, że związany z otyłością gen
FTO koduje funkcjonalny homolog białka AlkB (Gerken i wsp. 2007; Sanchez-Pulido
i Andrade-Navarro 2007). Wysoka ekspresja tego genu występuje w podwzgórzu,
a defekt genu FTO powiązano ze zwiększeniem się ilości tkanki tłuszczowej
w organizmie (Dina i wsp. 2007; Frayling i wsp. 2007, Scott i wsp. 2007). Białko FTO
posiada homologiczną sekwencję do białek z rodziny AlkB. Wykazano, że enzym ten
w obecności żelaza, α-KG i O2 ma zdolność do demetylacji m3T w jednoniciowym
DNA (Gerken i wsp. 2007).
Zróżnicowanie właściwości występujących u ssaków homologów białka AlkB
sugeruje, że mogą pełnić one różne funkcje w komórce (Sundheim i wsp. 2006).

28
II Cel pracy
Celem pracy było ustalenie warunków naprawy HEC i HPC przez białko AlkB
oraz określenie specyficzności substratowej homologów AlkB w stosunku do adduktów
egzocyklicznych i wyznaczenie optymalnych warunki naprawy tych uszkodzeń
w zależności od pH buforu, stężenia -ketoglutaranu i jonów żelaza. Wykorzystano tu
modyfikujące własności aldehydu chlorooctowego i akroleiny. Naprawa
powodowanych przez nie uszkodzeń przebiega w sposób bezpośredni - bez naruszania
ciągłości nici DNA. Przedmiotem badań były wyizolowane i oczyszczone białka: AlkB
oraz SA-1A i SA-2B, pochodzące od bakterii Streptomyces avermitilis.

29
III Materiały i metody
III.1. Materiały
III.1.1. Odczynniki
α-ketoglutaran (α-KG) – Fluka
Akroleina (ACR) – Fluka
Aldehyd chlorooctowy (CAA) – Fluka
Bis-Tris – Sigma
Ditiotreitol (DTT) – Sigma
Kwas octowy (CH3COOH, AcOH) – POCh
Kwas solny (HCl) – Chempur
Kwas 4-(2-hydroksyetylo)piperazino-1-etanosulfonowy (HEPES) – Sigma
Metanol (CH3OH, MeOH) – Lab-scan
Octan sodu (NaCOOH) – Chempur
Sześciouwodniony siarczan amonowo-żelazawy (NH4)2Fe(SO4)2 · 6H2O – Fluka
Trietyloamina (TEA, Et3N) – Merck
Wodorotlenek sodu (NaOH) – Stanlab
Odczynniki do elektroforezy
Żel rozdzielający (15%):
30% akrylamid - 2,5ml – Fluka
1,5M Tris (pH 8.8) – 1,3ml
10% SDS – 0,05ml
miliQ H2O – 1,1ml
10% nadsiarczan amonu (APS) - 0,05ml – Bio-Rad
N, N, N’ N’ - tetrametyletylenodiamina (TEMED) – 0,002ml - Fluka
Żel zagęszczający (4,5%):
30% akrylamid – 0,17ml
1M Tris (pH 6,8) – 0,13ml
10% SDS – 0,01ml
miliQ H2O – 0,68ml
10% nadsiarczan amonu (APS) – 0,01ml
N, N, N’ N’ - tetrametyletylenodiamina (TEMED) – 0,001ml

30
Marker
SigmaMarker™ Low Range (M.W. 6,500 - 66,000) – Sigma
Barwnik Coomasie Brillant Blue G – 250
0,2 % Coomasie Brillant Blue G – 250
10 % kwas octowy
45 % metanol
Odbarwiacz
10 % kwas octowy
25 % metanol
III.1.2. Bufory
III.1.2.1. Bufory stosowane do oczyszczania białka
Bufor sodowo-fosforanowy 160mM pH 7,4
Bufor do wiązania (BB) pH 7,4
Bufor do elucji (EB) pH 7,4
Bufor do przechowywania (ST) pH 7,4
Bufor do elektroforezy SDS-PAGE (pH 8,3)
0,125M Tris/HCl pH 8,3
0,96M glicyna
0,5% SDS
Bufor Laemmli’ego (pH 6,8)
0,25M Tris/HCl pH 6,8
50% glicerol
25% β-merkaptoetanol
10% SDS
0,05% błękit bromofenolowy
Bufor do dializy (pH 7,4)
40mM Tris/HCl pH 7,4
200mM NaCl
10% glicerol
5mM DTT
1mM PMSF

31
III.1.2.2. Bufory stosowane do reakcji modyfikacji pentametrów i ich naprawy
przez badane białka
0,5M Bis-Tris w zakresie pH 5,8 – 7,0
0,5M bufor octanowy w zakresie pH 4,2 – 5,6
0,5M HEPES w zakresie pH 6,8 – 7,8
Bufor kakodylanowy pH 6,5 – (CH3)2AsO2Na x 3H2O (BDH)
Bufor TE pH 7,8 (10mM Tris-HCl pH 7,8 + 1mM EDTA pH 8,0)
1,5M TEAB pH ok. 7-8 (węglan trietyloaminy)
Bufor do chromatografii wysokociśnieniowej
1M TEAA (octan trietyloaminy) pH ok. 6,5
30% metanol w H2O
III.1.3. Pentamery
TTATT – Metabion, Martinsried, Germany
TTCTT – Metabion, Martinsried, Germany
III.1.4. Enzymy
Plazmid niosący sklonowany gen alkB z E. coli otrzymano z pracowni prof.
Seeberga (Uniwersytet w Oslo). Plazmidy pET28a niosące sklonowane geny białek
SA-1A i SA-2B otrzymano z pracowni prof. Falnesa (Uniwersytet w Oslo). Plazmidem
transformowano szczep E. coli BL21 (DE3). Preparaty oczyszczonych białek EcAlkB,
SA-1A, SA-2B (His-tag) zostały otrzymane w Zakładzie Biologii Molekularnej IBB
PAN wykorzystując plazmid pET28a. Białka EcAlkB i SA-1A zostały otrzymane przez
dr Jadwigę Nieminuszczy, natomiast białko SA-2B zostało otrzymane przeze mnie.
III.1.5. Antybiotyki
Kanamycyna – Sigma
III.1.6. Pożywki
Pożywka LB płynna
0,5 % NaCl
0,5 % ekstrakt drożdżowy
1 % trypton

32
Pożywka LB stała
Podłoże LB płynne z dodatkiem 1 % agaru
III.1.7. Aparatura
III.1.7.1. Aparatura do oczyszczania białek
Zestaw do chromatografii niskociśnieniowej Äcta prime (Amersham)
Kolumna niklowa HisTrap (Amersham)
Spektrofotometr: Cary 3 UBV – Visible Spectrophotometer Varian
Spektrofotometr ULTROSPEC PLUS 4054 UV/VISIBLE Spectrophotometer LKB
BIOCHROM
Elektroporator: TransPorator Plus - BTX
Wirówka J2-21M/E – Beckman
Wirówka miniSpin – Eppendorf
Sonikator Branson Sonifier 250
pH-metr - inoLab
III.1.7.2. Aparatura do reakcji modyfikacji pentametrów i ich naprawy przez
badane białka
HPLC (High Pressure Liquid Chromatography) firmy Knauer na bazie systemu
podwójnego, zapewniającego gradient dwuskładnikowy, obsługiwany przez program
EuroChrom Basic Edition (wersja 3.05) i wyposażony w detektor UV.
Analityczny i preparatywny rozdział próbek prowadzono na kolumnach:
- Waters Nova-Pak® C18, 60Å, 4μm, 4,6mm x 250mm, z prędkością przepływu
1 ml/min,
- HR C18, 60Å, 6μm, 3,9mm x 300mm, z prędkością przepływu 1,75 ml/min,
z użyciem liniowego gradientu 20mM octanu trietyloaminy o pH 6,5 x 30% MeOH
w wodzie przez 30 minut i detekcją UV przy długości fali 270nm
Spektrofotometr: Cary 3 UBV – Visible Spectrophotometer Varian
Wirówka miniSpin – Eppendorf
pH-metr – MeterLab®
III.1.8. Inne materiały
bibuła 3mm – Whatman

33
folia spożywcza do przykrywania żeli
kuwety do elektroporacji – Bio-Rad
kuwety do spektrofotometrii – Medlab
papierki wskaźnikowe – MERCK
szyby 14cm x 16cm z przekładkami i grzebieniem 0,4mm
III.2. Metody
III.2.1. Sporządzanie buforów
Wszystkie bufory zostały wykonane na bazie wody dejonizowanej mQ.
III.2.1.1. Bufory stosowane do oczyszczania białka
Bufor wiążący (BB)
400ml buforu wiążącego uzyskano poprzez zmieszanie 50ml buforu sodowo-
fosforanowego z 2ml 10mM imidazolu i uzupełnienie wodą miliQ. Mieszaninę
stabilizowano kwasem solnym do pH o wartości 7,4.
Bufor do elucji (EB)
W celu sporządzenia buforu do elucji zmieszano 12,5ml buforu sodowo-
fosforanowego z 25ml 500mM imidazolu i uzupełniono wodą do 100ml.
pH ustabilizowano również do wartości 7,4. Tak przygotowane bufory BB i EB
schłodzono.
Bufor do przechowywania (ST)
Aby przygotować bufor do przechowywania zmieszano 40mM Tris/HCl,
200mM NaCl, 10% glicerolu. pH ustabilizowano do wartości 7,4.
III.2.1.2. Bufory stosowane do reakcji modyfikacji pentametrów i ich naprawy
przez badane białka
Sporządzono 0,2M bufory z przygotowanych wcześniej 0,5M buforów:
octanowego, HEPES i Bis-Tris. Do każdego dodano DTT do końcowego stężenia 5mM.
III.2.1.3. Bufor do chromatografii wysokociśnieniowej
1M octan trójetyloaminy (TEAA) o pH ~ 6,5 przygotowano w dwóch etapach.
I etap: Destylacja TEA
Kilka sit molekularnych wrzucono do kolby (o objętości 500ml) z TEA ~ 300ml
i ogrzano do wrzenia. Następnie obniżono temperaturę do 1800C. Przedgon zbierano do

34
momentu ustabilizowania się temperatury na termometrze (była nieco niższa niż 90ºC).
Następnie zebrano ok. 200ml destylatu TEA. Wyłączono transformator i pozostawiono
wszystko do wystygnięcia.
II etap
Schłodzono 1 mol (139,2 ml) świeżo destylowanej trójetyloaminy (Et3N, TEA),
1 mol (57,5 ml z 17,4M) kwasu octowego (AcOH) i 700 ml H2O mQ. Zlewkę z wodą
wstawiono w łaźni lodowej na mieszadło. Dodano TEA, który pływał po powierzchni
jak olej. Następnie powoli dodawano AcOH aż do uzyskania pH 6,5. Uzupełniono wodą
do 1000 ml i całość przesączono na sączku do HPLC.
Do rozdziałów chromatograficznych używano 20mM TEAA.
III.2.2. Transformacja bakterii szczepu E. coli BL21 (DE3) plazmidem pET28a/pSA
Dwie porcje zawierające po 100μl komórek elektrokompetentnych bakterii
szczepu E. coli BL21 (DE3) rozmrożono na lodzie. Do jednego z nich dodano 1μl
plazmidu pET28a/pSC, niosącego sklonowany gen białka SA-2B z Streptomyces
avermitilis, drugi natomiast stanowił kontrolę negatywną. Przygotowaną mieszaninę
transformacyjną przenoszono następnie do schłodzonej w lodzie kuwety. Kuwetę
wstawiono do elektroporatora i poddawano działaniu impulsu elektrycznego o napięciu
2,5 kV/cm. Do mieszaniny transformacyjnej dodawano następnie 1ml LB i inkubowano
przez 1 godz. w 37ºC w celu wyrażenia genu oporności na kanamycynę. Po
transformacji mieszaninę bakteryjną wysiano na płytki ze stałym podłożem LB
zawierającym kanamycynę o końcowym stężeniu 50 μg/ml. Płytki inkubowano przez
noc w temperaturze 37ºC. Następnie sprawdzano na płytkach obecność transformantów
oraz brak wzrostu na kontroli negatywnej.
III.2.3. Ekspresja białka SA-2B w szczepie E. coli BL21 (DE3) niosącym plazmid
pET28a/pSA
100ml płynnej pożywki LB z dodatkiem kanamycyny (50μg/ml) zaszczepiano
pojedynczą kolonią szczepu E. coli BL21 (DE3), niosącego plazmid kodujący białko
AlkB pET28a-SA-2B. Bakterie hodowano przez noc w 37ºC z wytrząsaniem 200 rpm.
Następnie hodowlę przeszczepiano na świeżą pożywkę (3 x 1 litr). 20 ml hodowli
nocnej dodano do każdej kolby zawierającej 1 litr płynnej pożywki LB z kanamycyną,
zawartość kolb połączono. Całość hodowano w 37ºC z wytrząsaniem do momentu

35
osiągnięcia gęstości OD600=0,6 (ok. 2,5 godziny) Z zawiesiny bakterii pobrano 1 ml
(kontrola nieindukowana). Resztę przeniesiono do 16ºC i hodowano przez 1 godzinę
w celu schłodzenia hodowli. Następnie indukowano ekspresję białka poprzez dodanie
izopropylo-β-tiogalaktopiranozyd (IPTG) do końcowego stężenia 0,1mM. Po indukcji
bakterie hodowano przez 16 godzin w 16ºC z wytrząsaniem 150 rpm. Po tym czasie
hodowle wirowano (6000 rpm w 4ºC przez 15 min). Supernatant odrzucono, a osady
zamrożono w -20ºC.
III.2.4. Dysrupcja komórek
Do osadu komórek bakteryjnych dodano β-merkaptoetanol (70μl/100ml)
i PMSF (1ml/100ml). Całość zawieszono w 60 ml buforu wiążącego pH 7,4 (20mM
bufor fosforanowy, 0,5M NaCl, 10mM imidazol). Zawiesinę przeniesiono do falkonów
i umieszczono w ciekłym azocie w celu szybkiego zamrożenia, następnie rozmrożono
pod zimną, bieżącą wodą. Po przeniesieniu zawartości falkonów do dwóch zlewek
i umieszczeniu w łaźni lodowej komórki sonikowano 2 razy po 3 minuty za pomocą
ultradźwięków o energii 250W, używając sonikatora Branson Sonifier 250. Kolejnym
etapem było zwirowanie próbek z prędkością 12000 x g/10 min w 4ºC (ultrawirówka
Beckman J2-21M/E). Osad odrzucono, a supernatant zebrano i przefiltrowano przez filtr
celulozowo-nitrowy 0,2μm (Milipore).
III.2.5. Oczyszczanie białka SA-2B
Plazmid bakteryjny pSA kodujący białko SA-2B (homologi AlkB) otrzymano od
prof. Falnesa z Uniwersytetu w Oslo. Plazmid pET28a/pSA zawierał wklonowaną
sekwencję cDNA kodującą białko SA-2B.
Wszystkie etapy oczyszczania białka odbywały się w temperaturze 4ºC.
Supernatant nakładano na kolumny ze złożem niklowym HisTrap (Amersham)
z wykorzystaniem urządzenia Äcta Prime. Białko eluowano w buforze elucyjnym
gradientem stężenia imidazolu od 10mM do 0,5M. Zebrane frakcje zachowano do
dalszej analizy, natomiast frakcje, które zawierały białko SA-2B zostały poddane
dializie do buforu do przechowywania 2xST (storage buffer) o pH 7,4 (40mM
Tris-HCl, 200mM NaCl, 10% glicerol). Na koniec dodano glicerol do końcowego
stężenia 50%.
Oczyszczone białka przechowywano w temperaturze -80ºC.

36
III.2.6. Elektroforeza w żelu poliakrylamidowym
Białka obecne w zebranych frakcjach (osad bakterii nieidukowanych IPTG,
supernatant przed nałożeniem na złoże, frakcje zebrane po dializie) zostały rozdzielone
elektroforetycznie w żelu poliakrylamidowym w obecności SDS-PAGE (warunki
denaturujące). Zastosowano metodę według modyfikacji Laemmli’ego, czyli w tzw.
systemie nieciągłym. W metodzie tej żel składa się z dwóch części. Część górna
zawiera 4,5% akrylamid o pH 6,8 i służy do zagęszczenia białek. Natomiast część dolna
zawiera 15% akrylamid o pH 8,8. Dochodzi w niej do rozdziału białek w zależności od
ich wielkości. Zanim próbki o objętości 5µl nałożono na żel, dodano do nich 1,5µl
5x stężonego buforu Laemmli’ego. Elektroforezę prowadzono w buforze do
elektroforezy SDS-PAGE (1x) pod napięciem 200 V do momentu wyjścia barwnika
z żelu. Rozdziałowi poddano też marker wielkości białek SigmaMarker™ Low Range
w zakresie 6,5 - 66 kDa, służący do oceny wielkości białka. Po rozdziale białka
barwiono Coomassie Brillant Blue G-250 przez 30 minut. Następnie barwnik zlewano,
a żel umieszczano w odbarwiaczu, który zmieniano kilkakrotnie, aż do całkowitego
odbarwienia się tła.
III.2.7. Oznaczanie stężenia białka metodą Bradford
Każdy pomiar wykonano w trzech powtórzeniach, ostateczny wynik stanowił
średnią arytmetyczną wszystkich wyników dla danej frakcji. 5μl każdej
z analizowanych frakcji rozcieńczano w kuwetach zawierających po 795μl sterylnej
wody miliQ, dodawano 200μl odczynnika Bradford, mieszano i odstawiano na 10 minut
w ciemności. Po tym czasie mierzono absorbancję roztworów przy długości fali 595 nm
względem kontroli (mieszanina 800μl wody i 200μl odczynnika Bradford). Stężenie
białka w analizowanych frakcjach określano na podstawie krzywej wzorcowej.
III.2.8. Przygotowanie substratów do reakcji naprawy przez badane białka
III.2.8.1. Otrzymywanie pentamerów zawierających etanocytozynę TT(εC)TT oraz
hydroksyetanocytozynę TT(HEC)TT
Do 276,5μl 1,5M buforu TEAB pH ok. 7-8 dodano 12,5μl pentameru TTCTT
o stężeniu 2 OD/μl (555nmoli) oraz 29,75μl (180μmol) 45% roztworu aldehydu

37
chlorooctowego (CAA). Całość inkubowano przez 2,5h w 37°C. Następnie mieszaninę
reakcyjną (nieprzereagowany TTCTT oraz TT(HEC)TT i TT(εC)TT) oczyszczono
i rozfrakcjonowano za pomocą HPLC na kolumnie preparatywnej, nakładając całkowitą
objętość reakcji. W celu określenia stężenia oligomeru zebrane frakcje poddano
pomiarowi absorbancji, po czym rozdzielono je za pomocą rozdziału analitycznego na
HPLC w celu sprawdzenia ich składu (nakładano po ok. 1nmol substancji). Następnie
frakcje zawierające modyfikowane pentamery liofilizowano do sucha i zawieszono
w buforze TE pH 8 w końcowym stężeniu 500pm/μl. Frakcje zawierające czysty THEC
umieszczono w temp. - 20ºC. Do frakcji zawierających mieszaninę TεC i THEC dodano
1M bufor kakodylanowy pH 6,5 i poddano procesowi dehydratacji przez ogrzewanie
w 37ºC w pH 6,5 przez 72h.
III.2.8.2. Otrzymywanie pentamerów zawierających hydroksypropanocytozynę
TT(HPC)TT
Do 214μl 1M buforu octanowego o pH 4,5 dodano 5μl pentameru TTCTT
o stężeniu 2 OD/μl (222nmoli) oraz 39μl (1nmol) akroleiny (ACR). Całość inkubowano
przez 4h w 37°C. Następnie mieszaninę reakcyjną oczyszczono za pomocą HPLC na
kolumnie preparatywnej, nakładając całkowitą objętość reakcji. W celu określenia
stężenia oligomeru zebrane frakcje zawierające TT(HPC)TT poddano pomiarowi
absorbancji, po czym rozdzielono je za pomocą HPLC na kolumnie analitycznej
(nakładano po ok. 1nmol substancji). Frakcje zawierające pożądany produkt połączono,
zliofilizowano, a następnie rozpuszczono w buforze TE do stężenia 500pmoli/μl.
III.2.8.3. Otrzymywanie substratu zawierającego etenoadeninę TT(εA)TT
Do 442,4μl 0,5 M buforu octanowego o pH 4,6 dodano 5μl pentameru TTATT
o stężeniu 2 OD/μl (222 nmole) oraz 47,6 μl (288 μmol) 45 % roztworu aldehydu
chlorooctowego (CAA). Całość inkubowano przez 24 h w 37°C. Następnie mieszaninę
reakcyjną oczyszczano za pomocą HPLC na kolumnie, nakładając całkowitą objętość
reakcji. Zebrane frakcje zawierające TTεATT poddano pomiarowi absorbancji w celu
określenia stężenia oligomeru, a następnie rozdziałowi za pomocą HPLC na kolumnie
analitycznej (nakładano po ok. 1 nmol substancji). Frakcje zawierające pożądany
produkt połączono, zliofilizowano, a następnie rozpuszczono w buforze TE do stężenia
500 pmoli/μl.

38
III.2.9. Badanie aktywności białka AlkB, SA-1A i SA-2B
W celu badania aktywności badanych białek przygotowano 0,2M bufory:
Bis-Tris, HEPES i octanowy w różnych zakresach pH wraz z dodatkiem 1M DTT
(5μl/ml). Reakcje prowadzono w zależności od pH buforu, czasu, stężenia jonów Fe (II)
i stężenia α-ketoglutaranu. Objętości wszystkich mieszanin reakcyjnych wynosiły 20μl,
a ich inkubacja w temp. 37ºC trwała 30 minut (w przypadku badania zależności od
czasu inkubacja trwała do 30 minut). Po upływie tego czasu reakcje zatrzymywano
przez dodanie 230μl wody i zamrożenie. Analizę składu mieszaniny reakcyjnej
prowadzono za pomocą HPLC na kolumnie analitycznej.
Wydajność naprawy obliczano porównując procentowy udział modyfikowanego
i niemodyfikowanego pentameru w rozdziale chromatograficznym HPLC przed i po
reakcji z badanym białkiem.
III.2.9.1. Zależność naprawy od pH
Reakcje startowano buforem o odpowiednim pH. α-ketoglutaran i jony Fe (II)
w postaci (NH4)2Fe(SO4)2 x 6H2O dodawano tuż przed reakcją ze względu na ich
nietrwałość.
TT(HEC)TT
W przypadku białka AlkB reakcje naprawy prowadzono w buforze octanowym
o pH 5,4 oraz w buforze Bis-Tris o pH: 5,8; 6,0; 6,2 lub 6,6. Reakcje z białkami SA-1A
i SA-2B prowadzono w buforze Bis-Tris o pH: 5,8; 6,0; 6,2; 6,6 i 7,0. We wszystkich
mieszaninach z białkiem SA-1A i SA-2B końcowe stężenia α-ketoglutaranu i Fe (II)
wynosiły po 0,5mM. Natomiast w reakcjach z białkiem AlkB stężenie Fe (II) wynosiło
1mM, a stężenie α-KG 0,5mM. Pozostałe składniki reakcji stanowiły: woda miliQ,
substrat TT(HEC)TT (500pm) oraz białko (AlkB 25pm, SA-1A 20pm i SA-2B 50pm).
TT(HPC)TT
Reakcje z udziałem białka AlkB i SA1A prowadzono w buforze HEPES
o pH: 6,8; 7,2; 7,4; 7,6 oraz 7,8. W reakcji z białkiem SA-1A stężenie α-KG było równe
0,5mM, natomiast stężenie Fe (II) wynosiło 0,1mM. W reakcjach z białkiem AlkB
stężenie Fe (II)i α-KG wynosiło 0,05mM. Pozostałe składniki stanowiła woda, substrat
TT(HPC)TT (500pm) i białko (AlkB 25pm, SA-1A 20pm).

39
TT(εC)TT i TT(εA)TT
Reakcje dla białek SA-1A i SA-2B prowadzono w buforze octanowym o pH:
4,2; 4,6; 5,0; 5,4 i 5,6. Stężenie α-ketoglutaranu wynosiło 0,5mM, natomiast stężenie Fe
(II) 3mM. Pozostałe składniki stanowiła woda miliQ, substrat (500pm) oraz białko
(50pm).
III.2.9.2. Zależność naprawy od stężenia α-KG
W celu porównania uzyskanych wyników i zminimalizowania błędu
pipetowania z wyjściowego roztworu o stężeniu 100mM przygotowano szereg
rozcieńczeń α-KG.
TT(HEC)TT
Reakcje z białkiem AlkB i SA-1A prowadzono w buforze Bis-Tris o pH 5,8.
Stężenia α-KG w reakcjach z AlkB wynosiły 0,01mM; 0,025mM; 0,05mM; 0,1mM;
0,25mM; 0,5mM, a w reakcjach z SA-1A wynosiły 0mM; 0,01mM; 0,025mM;
0,05mM; 0,1mM i 0,5mM. Pozostałe składniki reakcji to: Fe (II) (1mM w reakcji
z AlkB, 500μM w reakcji z SA-1A), woda miliQ, substrat TT(HEC)TT (500pm)
i białko (AlkB 25pm, SA-1A 20pm).
TT(HPC)TT
Reakcje z białkiem AlkB prowadzono w buforze HEPES o pH 7,4, natomiast
reakcje z białkiem SA-1A prowadzono w buforze HEPES o pH 7,4. Stężenia α-KG
w reakcjach z AlkB wynosiły 0,025mM; 0,05mM; 0,1mM; 0,25mM; 0,5mM i 1mM.
Natomiast w reakcjach z SA-1A stężenia α-KG wynosiły 0mM; 0,01mM; 0,025mM;
0,05mM; 0,1mM i 0,5mM. Pozostałe składniki reakcji to: 100μM Fe (II), woda miliQ,
substrat TT(HPC)TT (500pm) i białko (50pm).
TT(εA)TT i TT(εC)TT
Reakcje naprawy przez białko SA-1A przeprowadzono w buforze octanowym
o pH 5,0. Stężenia α-KG w reakcjach wynosiły 0mM; 0,01mM; 0,025mM; 0,05mM;
0,1mM i 0,5mM. Pozostałe składniki stanowiły: Fe (II) o stężeniu 3mM, woda miliQ,
substrat (500pm) i białko (50pm).

40
III.2.9.3. Zależność naprawy od stężenia jonów Fe (II)
W celu porównania uzyskanych wyników i zminimalizowania błędu
pipetowania z wyjściowego roztworu o stężeniu 100mM przygotowano szereg
rozcieńczeń jonów Fe (II) w postaci (NH4)2Fe(SO4)2 x 6H2O.
TT(HEC)TT
Reakcje z białkiem AlkB, SA-1A i SA-2B prowadzono w buforze Bis-Tris o pH
5,8. Stężenia Fe (II) w reakcjach z AlkB wynosiły 0,25mM; 0,5mM; 1mM; 1,5mM;
2mM; 2,5mM i 3mM, a w reakcjach z SA-1A i SA-2B: 0mM; 0,1mM; 0,5mM; 1mM;
2mM i 3mM. Pozostałe składniki reakcji to: α-ketoglutaran (500 μM w reakcji z AlkB,
25μM w reakcji z SA-1A i 250μM w reakcji z SA-2B), woda miliQ, substrat
TT(HEC)TT (500pm) i białko (AlkB 25pm, SA-1A i SA-2B 50pm).
TT(HPC)TT
Reakcje z białkiem AlkB i SA-1A prowadzono w buforze HEPES o pH 7,4.
Stężenia Fe (II) w reakcjach z AlkB wynosiły 0,01mM; 0,025mM; 0,05mM; 0,1mM;
0,25mM, a w reakcjach z SA-1A: 0mM; 0,01mM; 0,05mM; 0,1mM; 0,25mM i 0,5mM.
Pozostałe składniki reakcji to: α-ketoglutaran (50μM), woda miliQ, substrat
TT(HPC)TT (500pm) i białko (AlkB 25pm, SA-1A 50pm).
TT(εA)TT i TT(εC)TT
Reakcje naprawy przez białko SA-2B przeprowadzono w buforze octanowym
o pH 5,0. Użyto α-KG o stężeniu 250μM. Stężenia Fe (II) w reakcjach wynosiły
0,5mM, 1,5mM, 3mM i 5mM. Pozostałe składniki stanowiła woda, substrat (500pm)
i białko (50pm).
III.2.9.4. Zależność naprawy od czasu
W reakcjach z białkiem AlkB inkubacja mieszaniny reakcyjnej w temp. 37ºC
trwała odpowiednio: 1; 2,5; 5; 10; 20 lub 30 minut.
TT(HEC)TT
Reakcje z białkiem AlkB prowadzono w buforze Bis-Tris o pH 5,8; przy
stężeniu Fe (II) równym 1mM i stężeniu α-ketoglutaranu wynoszącym 500 μM.

41
Pozostałe składniki mieszaniny reakcyjnej to: woda miliQ, substrat TT(HEC)TT
(500pm) i białko (25pm).
TT(HPC)TT
Reakcje z białkiem AlkB prowadzono w buforze HEPES o pH 7,5; przy stężeniu
Fe (II) równym 100 μM i stężeniu α-ketoglutaranu 500 μM. Pozostałe składniki
mieszaniny reakcyjnej stanowiły: woda miliQ, substrat TT(HPC)TT (500pm) i białko
(12,5pm).
42
IV Wyniki
IV.1. Oczyszczanie białka SA-2B
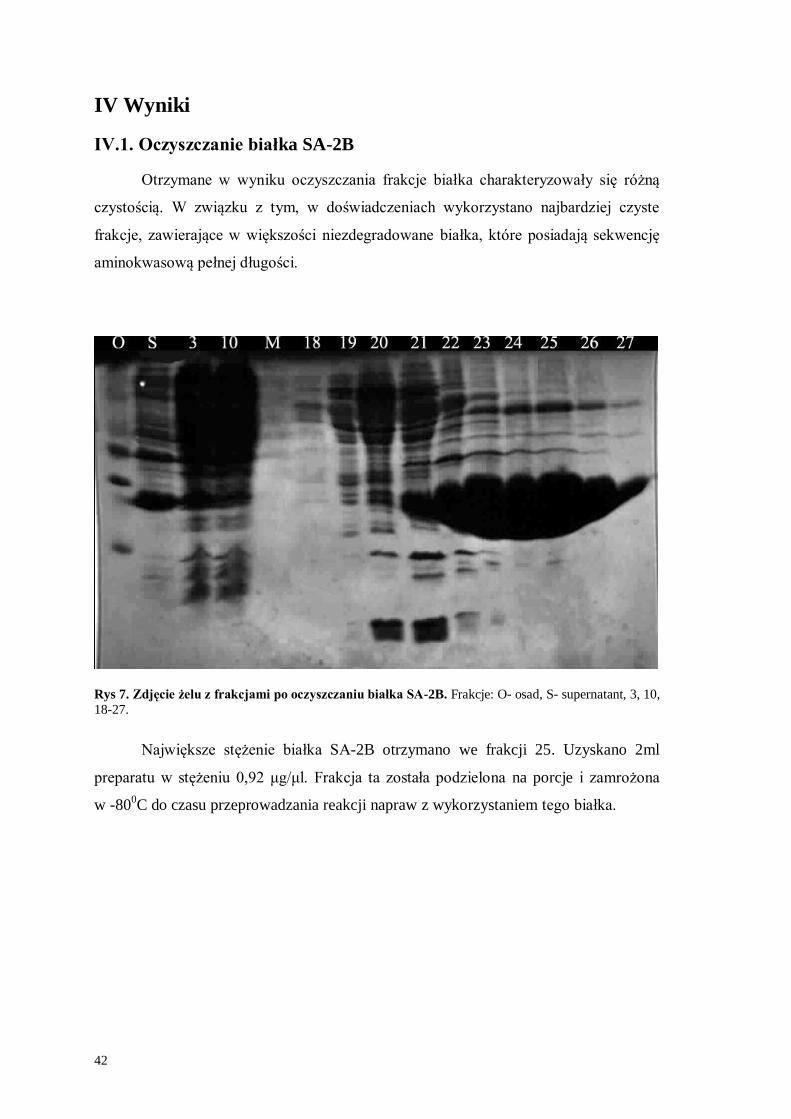
Otrzymane w wyniku oczyszczania frakcje białka charakteryzowały się różną
czystością. W związku z tym, w doświadczeniach wykorzystano najbardziej czyste
frakcje, zawierające w większości niezdegradowane białka, które posiadają sekwencję
aminokwasową pełnej długości.
Rys 7. Zdjęcie żelu z frakcjami po oczyszczaniu białka SA-2B. Frakcje: O- osad, S- supernatant, 3, 10,
18-27.
Największe stężenie białka SA-2B otrzymano we frakcji 25. Uzyskano 2ml
preparatu w stężeniu 0,92 μg/μl. Frakcja ta została podzielona na porcje i zamrożona
w -800C do czasu przeprowadzania reakcji napraw z wykorzystaniem tego białka.
43
IV.2. Chemiczna modyfikacja pentamerów TTCTT i TTATT
IV.2.1. Modyfikacja pentamerów zawierających cytozynę (TTCTT)
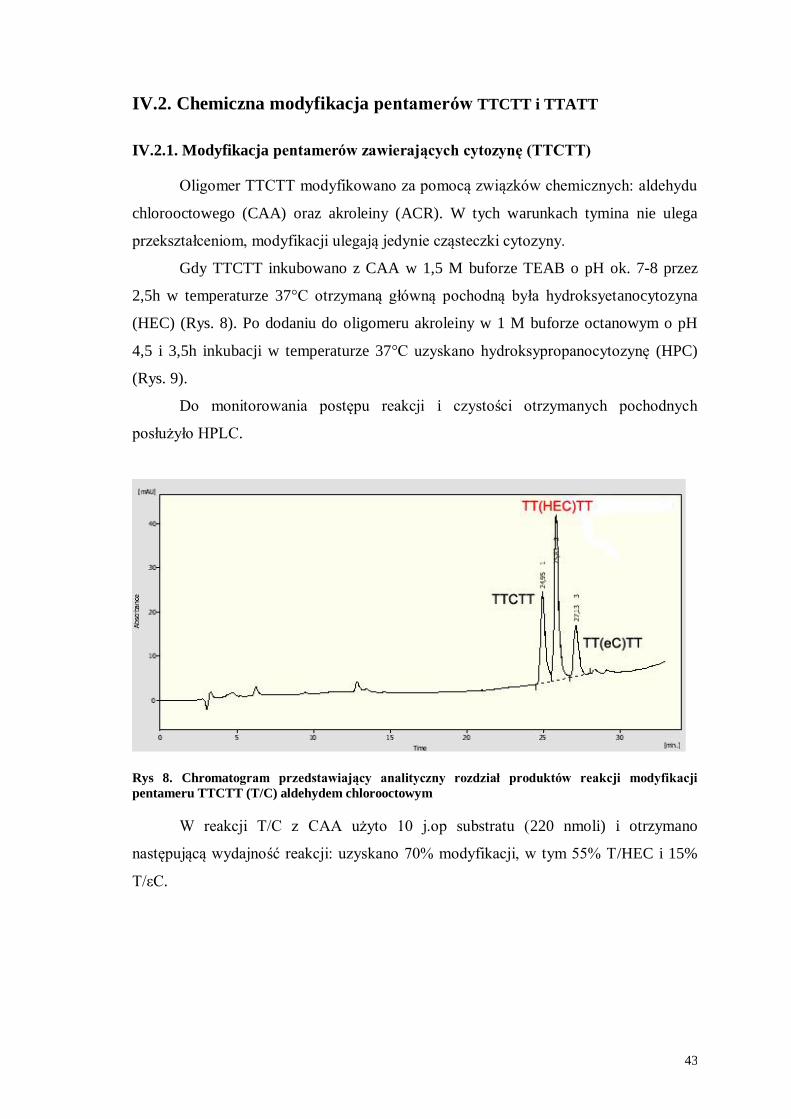
Oligomer TTCTT modyfikowano za pomocą związków chemicznych: aldehydu
chlorooctowego (CAA) oraz akroleiny (ACR). W tych warunkach tymina nie ulega
przekształceniom, modyfikacji ulegają jedynie cząsteczki cytozyny.
Gdy TTCTT inkubowano z CAA w 1,5 M buforze TEAB o pH ok. 7-8 przez
2,5h w temperaturze 37°C otrzymaną główną pochodną była hydroksyetanocytozyna
(HEC) (Rys. 8). Po dodaniu do oligomeru akroleiny w 1 M buforze octanowym o pH
4,5 i 3,5h inkubacji w temperaturze 37°C uzyskano hydroksypropanocytozynę (HPC)
(Rys. 9).
Do monitorowania postępu reakcji i czystości otrzymanych pochodnych
posłużyło HPLC.
Rys 8. Chromatogram przedstawiający analityczny rozdział produktów reakcji modyfikacji
pentameru TTCTT (T/C) aldehydem chlorooctowym
W reakcji T/C z CAA użyto 10 j.op substratu (220 nmoli) i otrzymano
następującą wydajność reakcji: uzyskano 70% modyfikacji, w tym 55% T/HEC i 15%
T/εC.
44
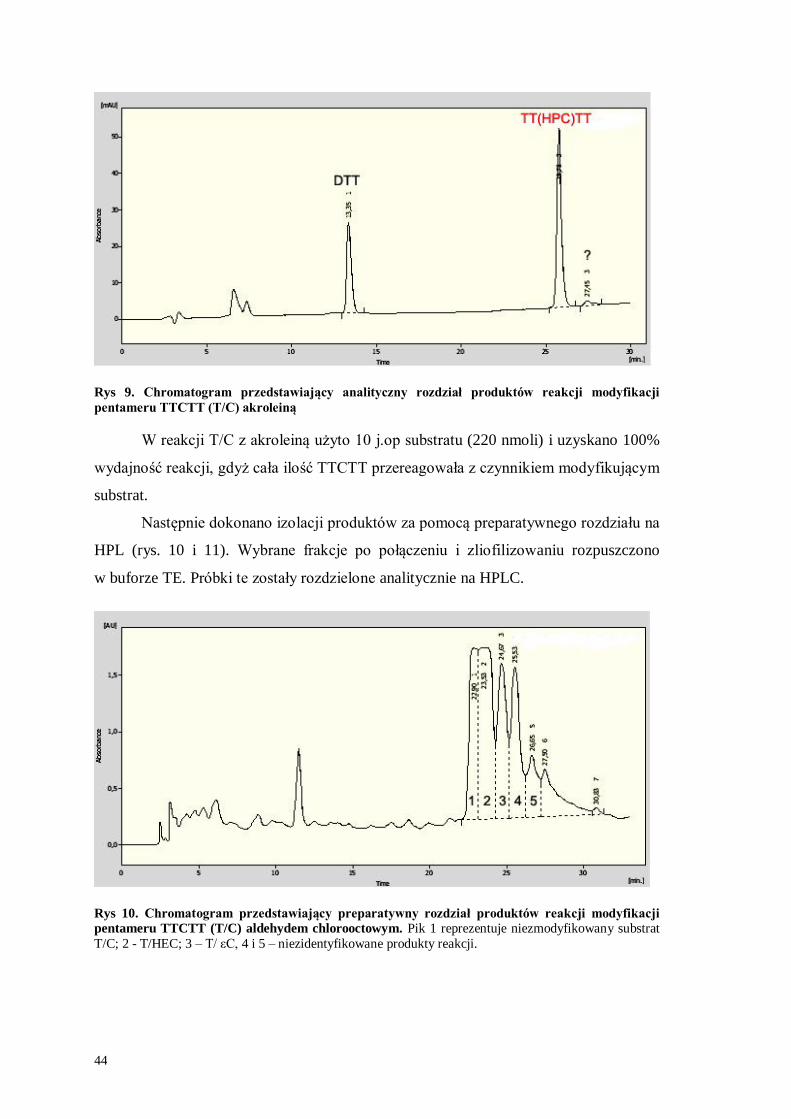
Rys 9. Chromatogram przedstawiający analityczny rozdział produktów reakcji modyfikacji
pentameru TTCTT (T/C) akroleiną
W reakcji T/C z akroleiną użyto 10 j.op substratu (220 nmoli) i uzyskano 100%
wydajność reakcji, gdyż cała ilość TTCTT przereagowała z czynnikiem modyfikującym
substrat.
Następnie dokonano izolacji produktów za pomocą preparatywnego rozdziału na
HPL (rys. 10 i 11). Wybrane frakcje po połączeniu i zliofilizowaniu rozpuszczono
w buforze TE. Próbki te zostały rozdzielone analitycznie na HPLC.
Rys 10. Chromatogram przedstawiający preparatywny rozdział produktów reakcji modyfikacji
pentameru TTCTT (T/C) aldehydem chlorooctowym. Pik 1 reprezentuje niezmodyfikowany substrat
T/C; 2 - T/HEC; 3 – T/ εC, 4 i 5 – niezidentyfikowane produkty reakcji.
45
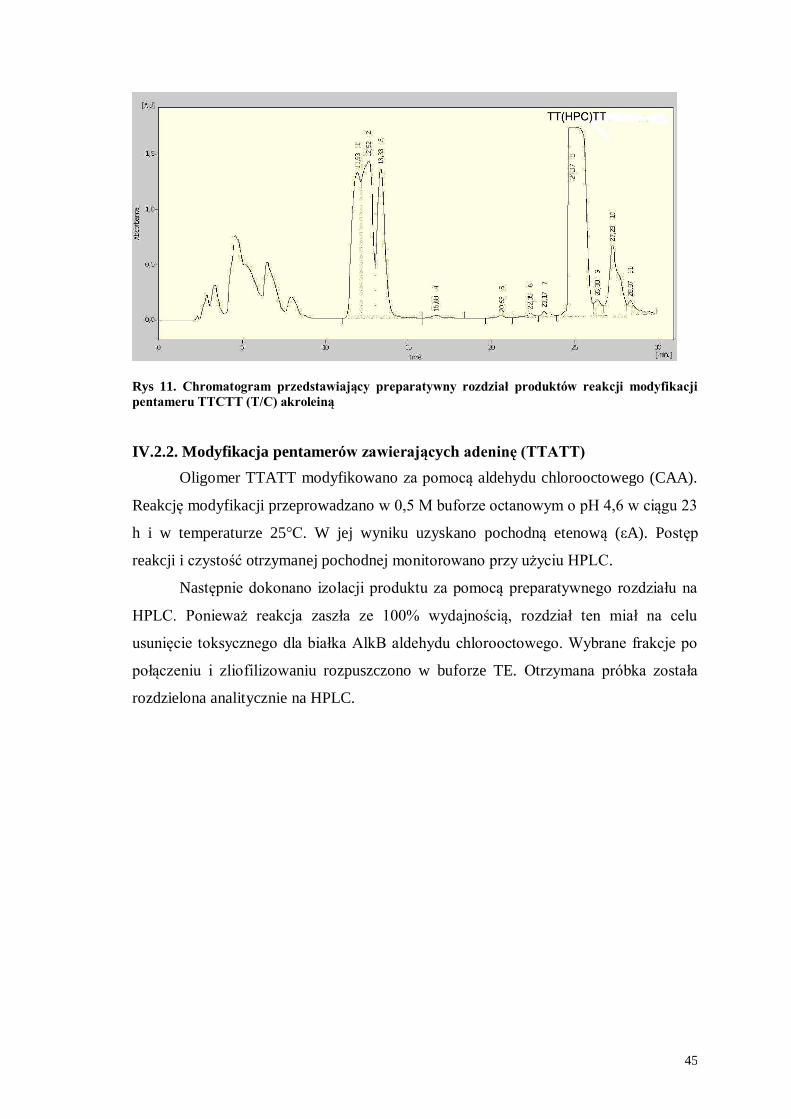
Rys 11. Chromatogram przedstawiający preparatywny rozdział produktów reakcji modyfikacji
pentameru TTCTT (T/C) akroleiną
IV.2.2. Modyfikacja pentamerów zawierających adeninę (TTATT)
Oligomer TTATT modyfikowano za pomocą aldehydu chlorooctowego (CAA).
Reakcję modyfikacji przeprowadzano w 0,5 M buforze octanowym o pH 4,6 w ciągu 23
h i w temperaturze 25°C. W jej wyniku uzyskano pochodną etenową (εA). Postęp
reakcji i czystość otrzymanej pochodnej monitorowano przy użyciu HPLC.
Następnie dokonano izolacji produktu za pomocą preparatywnego rozdziału na
HPLC. Ponieważ reakcja zaszła ze 100% wydajnością, rozdział ten miał na celu
usunięcie toksycznego dla białka AlkB aldehydu chlorooctowego. Wybrane frakcje po
połączeniu i zliofilizowaniu rozpuszczono w buforze TE. Otrzymana próbka została
rozdzielona analitycznie na HPLC.

46
Rys 12. Chromatogram przedstawiający analityczny rozdział produktów reakcji
modyfikacji pentameru TTATT (T/A) aldehydem chlorooctowym
IV.3. Badanie naprawy modyfikowanych oligomerów przez białko
AlkB in vitro
W celu analizy aktywności naprawczej białka wobec otrzymanych substratów
przeprowadzono inkubację oczyszczonych pentametrów z białkiem AlkB w obecności
α-ketoglutaranu (α-KG), jonów żelaza (II), buforu o odpowiednim pH, ditiotreitolu
(DTT) oraz wody. Badano wpływ różnych czynników takich jak: pH, stężenie Fe (II),
stężenie α-KG, czas; na wydajność naprawy poszczególnych substratów przez białko
AlkB. Stopień naprawy oligomeru przez białko AlkB był określany na podstawie
analizy uzyskanych chromatogramów.
47
IV.3.1. Naprawa TT(HEC)TT
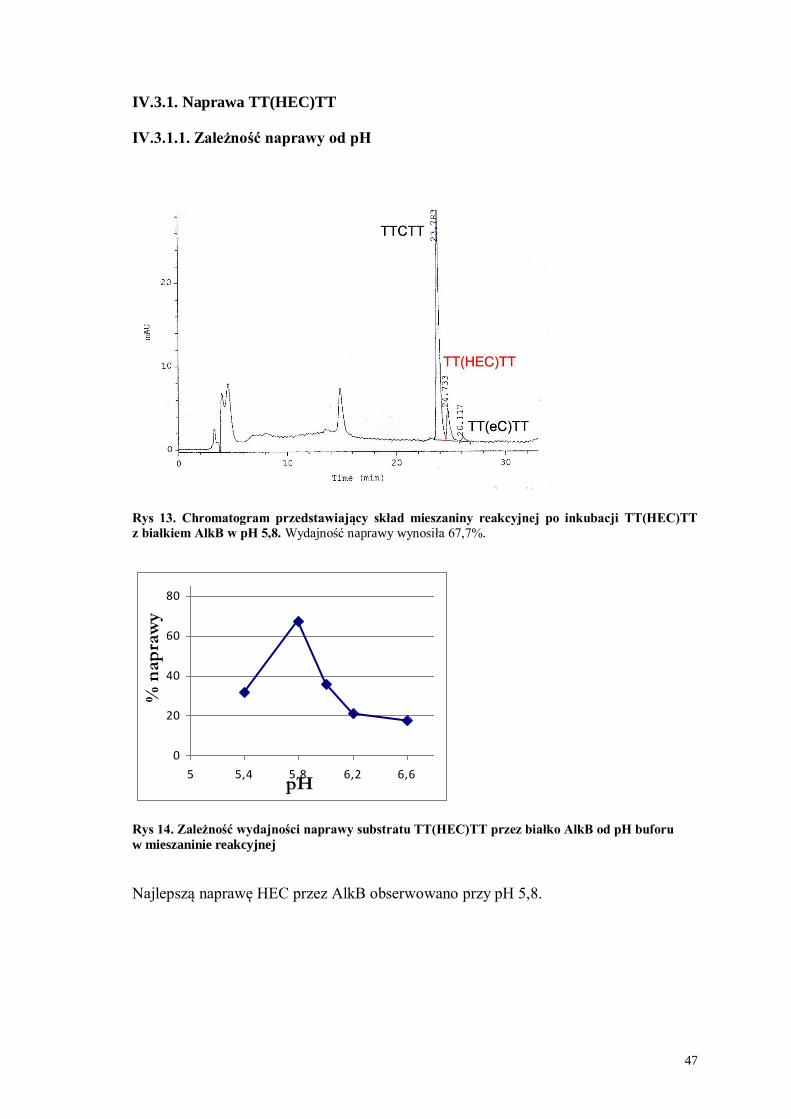
IV.3.1.1. Zależność naprawy od pH
Rys 13. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HEC)TT
z białkiem AlkB w pH 5,8. Wydajność naprawy wynosiła 67,7%.
0
20
40
60
80
5 5,4 5,8 6,2 6,6pH
% n
ap
raw
y
Rys 14. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od pH buforu
w mieszaninie reakcyjnej
Najlepszą naprawę HEC przez AlkB obserwowano przy pH 5,8.
48
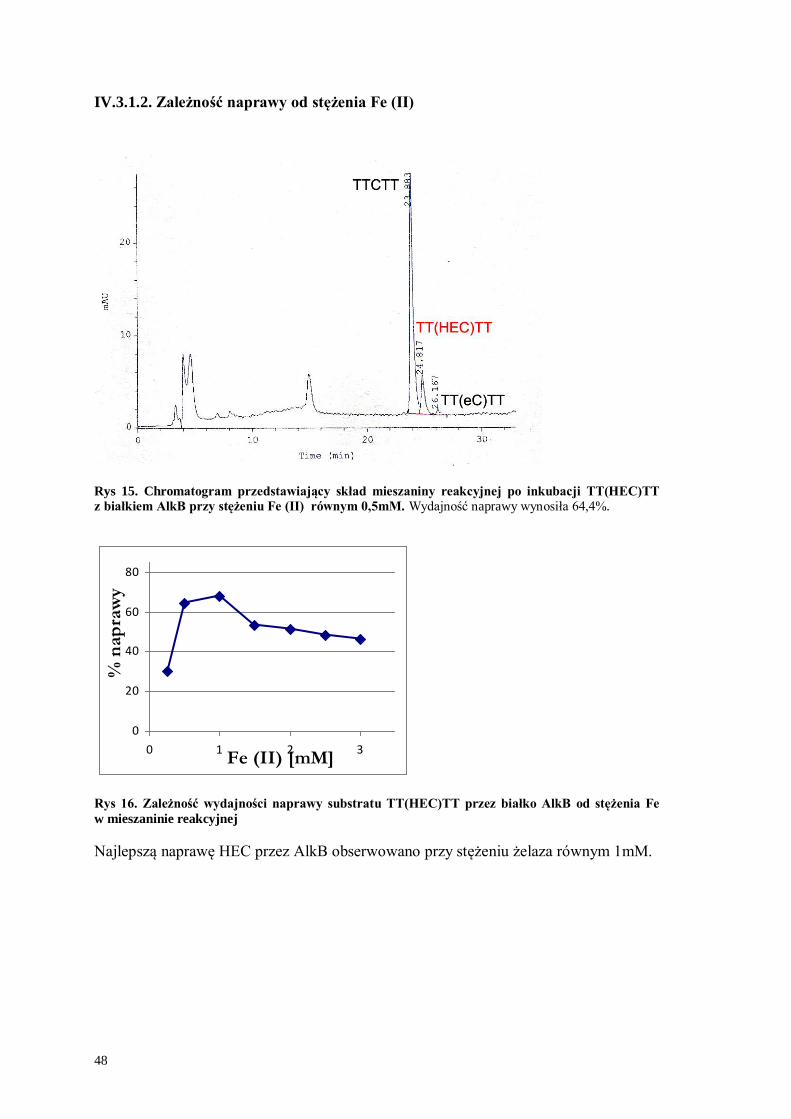
IV.3.1.2. Zależność naprawy od stężenia Fe (II)
Rys 15. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HEC)TT
z białkiem AlkB przy stężeniu Fe (II) równym 0,5mM. Wydajność naprawy wynosiła 64,4%.
0
20
40
60
80
0 1 2 3Fe (II) [mM]
% n
ap
raw
y
Rys 16. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od stężenia Fe
w mieszaninie reakcyjnej
Najlepszą naprawę HEC przez AlkB obserwowano przy stężeniu żelaza równym 1mM.

49
IV.3.1.3. Zależność naprawy od stężenia α-KG
Rys 17. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HEC)TT
z białkiem AlkB przy stężeniu α-KG równym 0,05mM. Wydajność naprawy wynosiła 71,8%.
0
20
40
60
80
0 0,1 0,2 0,3 0,4 0,5 0,6αKG [mM]
% n
ap
raw
y
Rys 18. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od stężenia α-KG
w mieszaninie reakcyjnej
Najlepszą naprawę HEC przez AlkB obserwowano przy stężeniu α-ketoglutaranu
równym 0,025mM.

50
IV.3.1.4. Zależność naprawy od czasu
Rys 19. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HEC)TT
z białkiem AlkB w czasie 30min. Wydajność naprawy wynosiła 42%.
0
10
20
30
40
50
0 10 20 30czas [min.]
% n
ap
raw
y
Rys 20. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od czasu
Najlepszą naprawę HEC przez AlkB obserwowano przy reakcji prowadzonej przez
30min.
51
IV.3.2. Naprawa TT(HPC)TT
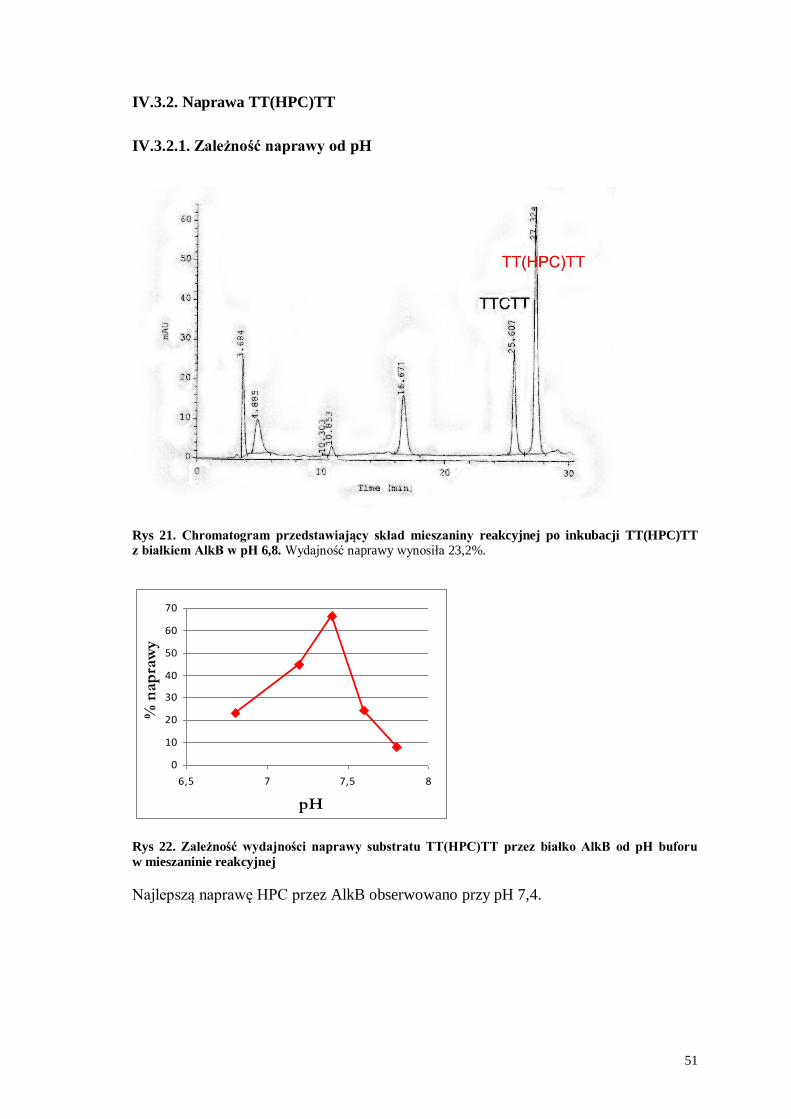
IV.3.2.1. Zależność naprawy od pH
Rys 21. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HPC)TT
z białkiem AlkB w pH 6,8. Wydajność naprawy wynosiła 23,2%.
0
10
20
30
40
50
60
70
6,5 7 7,5 8
pH
% n
ap
raw
y
Rys 22. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od pH buforu
w mieszaninie reakcyjnej
Najlepszą naprawę HPC przez AlkB obserwowano przy pH 7,4.
52
IV.3.2.2. Zależność naprawy od stężenia Fe (II)
Rys 23. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HPC)TT
z białkiem AlkB przy stężeniu Fe (II) równym 0,1mM. Wydajność naprawy wynosiła 68%.
0
20
40
60
80
0 0,05 0,1 0,15 0,2 0,25 0,3
Fe (II) [mM]
% n
ap
raw
y
Rys 24. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od stężenia Fe
(II) w mieszaninie reakcyjnej
Najlepszą naprawę HPC przez AlkB obserwowano przy stężeniu żelaza równym
0,1mM.
53
IV.3.2.3. Zależność naprawy od stężenia α-KG
Rys 25. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HPC)TT
z białkiem AlkB przy stężeniu α-KG równym 0,1mM. Wydajność naprawy wynosiła 69,3%.
0
20
40
60
80
100
0 0,1 0,2 0,3 0,4 0,5 0,6αKG [mM]
% n
ap
raw
y
Rys 26. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od stężenia α-KG
w mieszaninie reakcyjnej
Najlepszą naprawę HPC przez AlkB obserwowano przy stężeniu α-ketoglutaranu
równym 0,05mM.
54
IV.3.2.4. Zależność naprawy od czasu
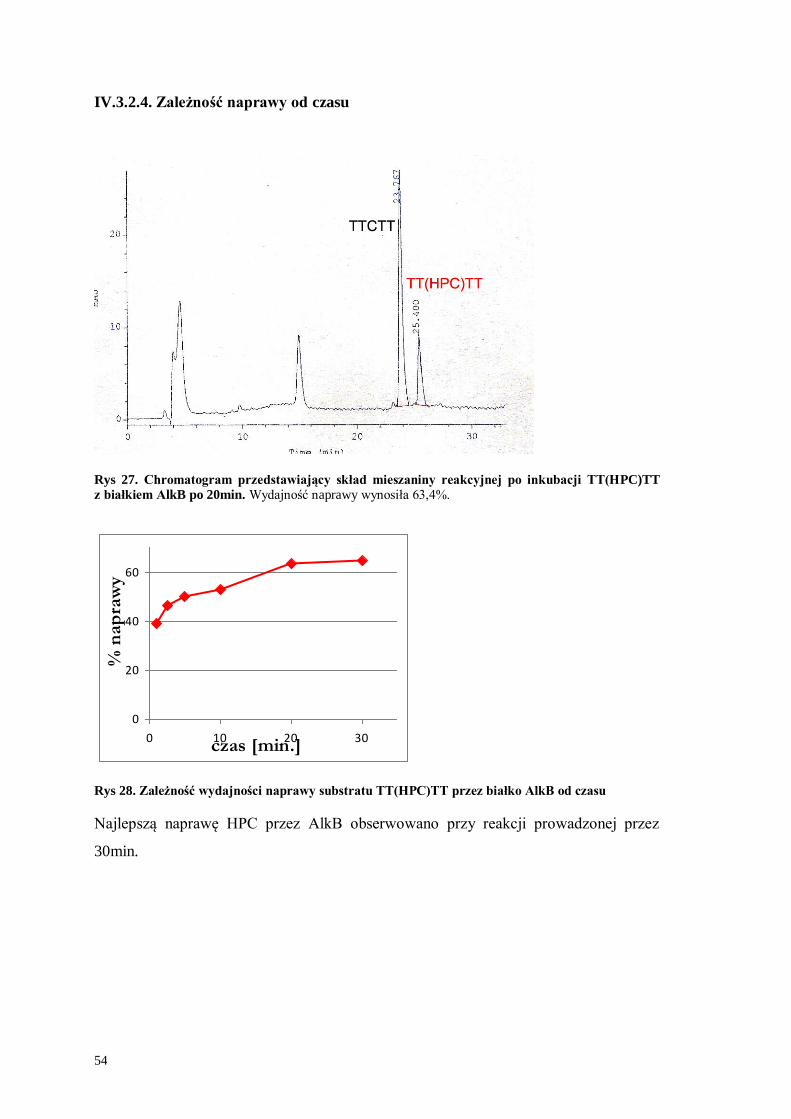
Rys 27. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HPC)TT
z białkiem AlkB po 20min. Wydajność naprawy wynosiła 63,4%.
0
20
40
60
0 10 20 30czas [min.]
% n
ap
raw
y
Rys 28. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od czasu
Najlepszą naprawę HPC przez AlkB obserwowano przy reakcji prowadzonej przez
30min.

55
IV.4. Badanie naprawy in vitro modyfikowanych oligomerów przez
bakteryjne homologi białka AlkB: SA-1A i SA-2B
Przeprowadzono analizę aktywności naprawczej białka SA-1A i SA-2B wobec
zsyntetyzowanych substratów. W tym celu inkubowano w 37°C otrzymane oczyszczone
pentamery z każdym z tych białek w obecności α-ketoglutaranu (α-KG), jonów Fe (II),
buforu o odpowiednim pH, ditiotreitolu (DTT) oraz wody. W celu zatrzymania reakcji
mieszaniny reakcyjne rozcieńczano wodą i zamrażano. Badano wpływ różnych
czynników takich jak: pH, stężenie Fe, stężenie α-KG, czas; na wydajność naprawy
poszczególnych substratów przez białko SA-1 i SA-2. Analizując wyniki reakcji po
rozdziale chromatograficznym określano stopień naprawy oligomeru przez homologi
białka AlkB.
IV.4.1. Białko SA-1A
Na rys. 29, 30, 31 i 32 przedstawiono wzorce substratów użytych do badania
specyficzności substratowej białka SA-1:
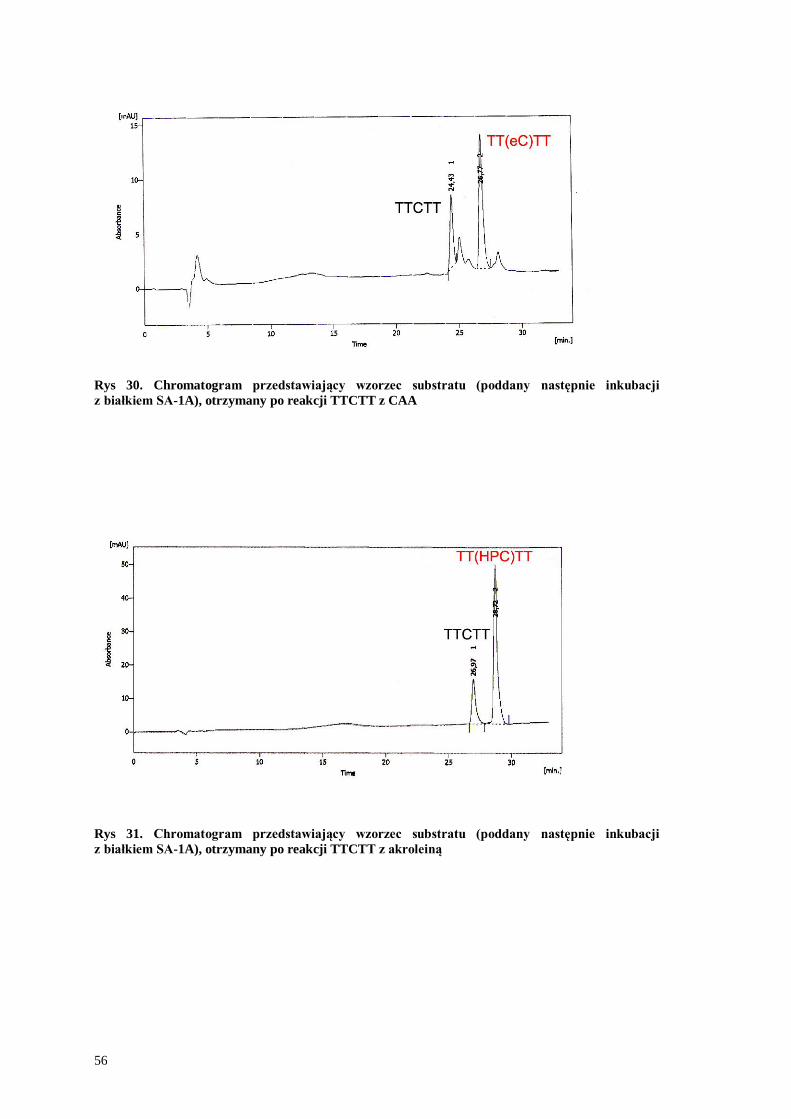
Rys 29. Chromatogram przedstawiający wzorzec substratu (poddany następnie inkubacji
z białkiem SA-1A), otrzymany po reakcji TTCTT z CAA
56
Rys 30. Chromatogram przedstawiający wzorzec substratu (poddany następnie inkubacji
z białkiem SA-1A), otrzymany po reakcji TTCTT z CAA
Rys 31. Chromatogram przedstawiający wzorzec substratu (poddany następnie inkubacji
z białkiem SA-1A), otrzymany po reakcji TTCTT z akroleiną
57
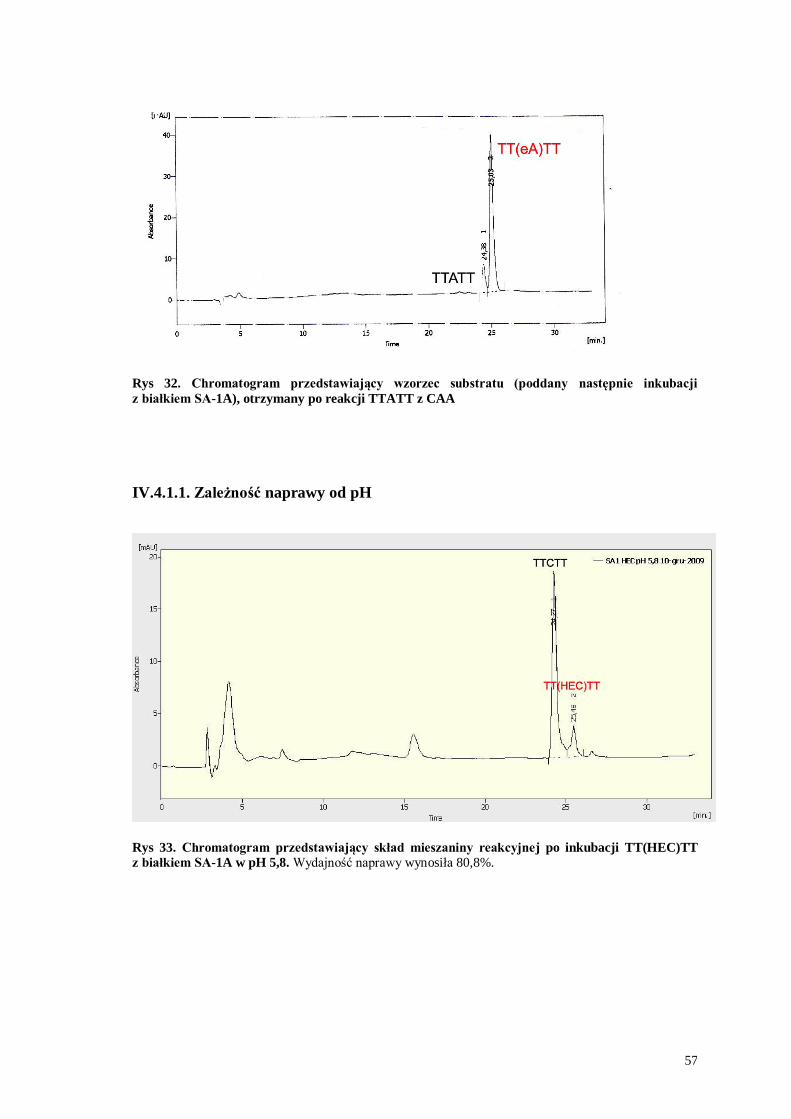
Rys 32. Chromatogram przedstawiający wzorzec substratu (poddany następnie inkubacji
z białkiem SA-1A), otrzymany po reakcji TTATT z CAA
IV.4.1.1. Zależność naprawy od pH
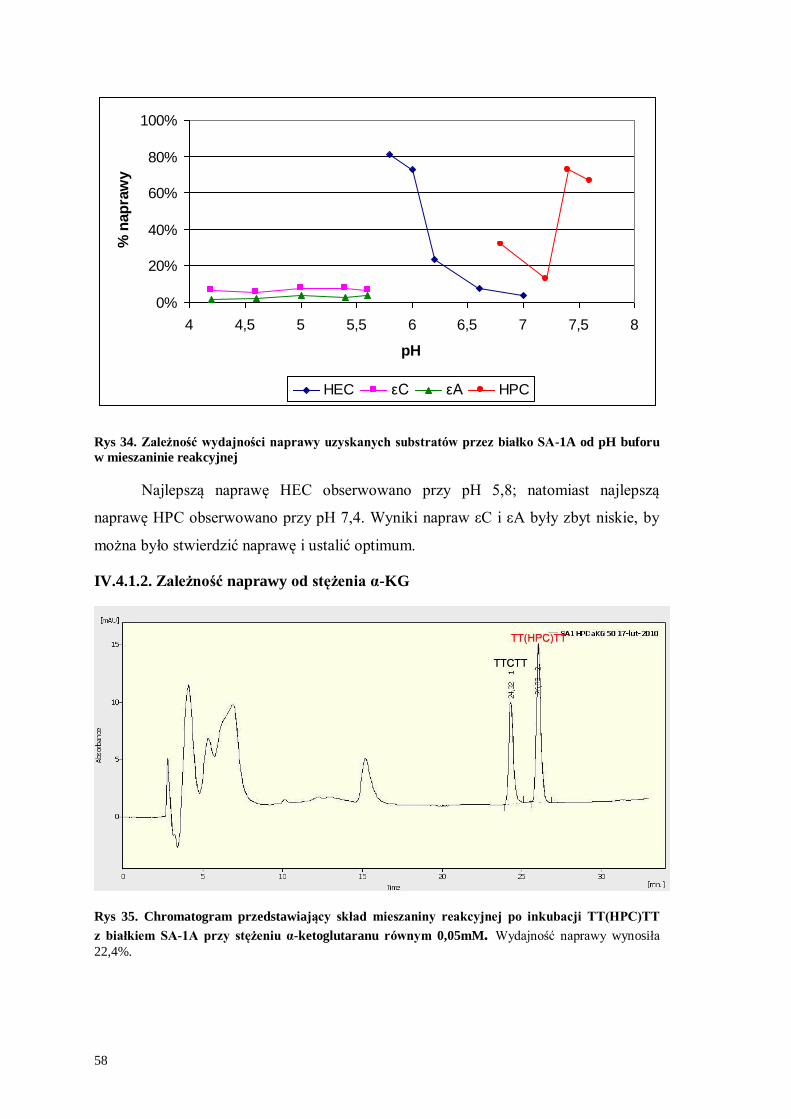
Rys 33. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HEC)TT
z białkiem SA-1A w pH 5,8. Wydajność naprawy wynosiła 80,8%.
58
0%
20%
40%
60%
80%
100%
4 4,5 5 5,5 6 6,5 7 7,5 8
pH
% n
ap
raw
y
HEC εC εA HPC
Rys 34. Zależność wydajności naprawy uzyskanych substratów przez białko SA-1A od pH buforu
w mieszaninie reakcyjnej
Najlepszą naprawę HEC obserwowano przy pH 5,8; natomiast najlepszą
naprawę HPC obserwowano przy pH 7,4. Wyniki napraw εC i εA były zbyt niskie, by
można było stwierdzić naprawę i ustalić optimum.
IV.4.1.2. Zależność naprawy od stężenia α-KG
Rys 35. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HPC)TT
z białkiem SA-1A przy stężeniu α-ketoglutaranu równym 0,05mM. Wydajność naprawy wynosiła
22,4%.
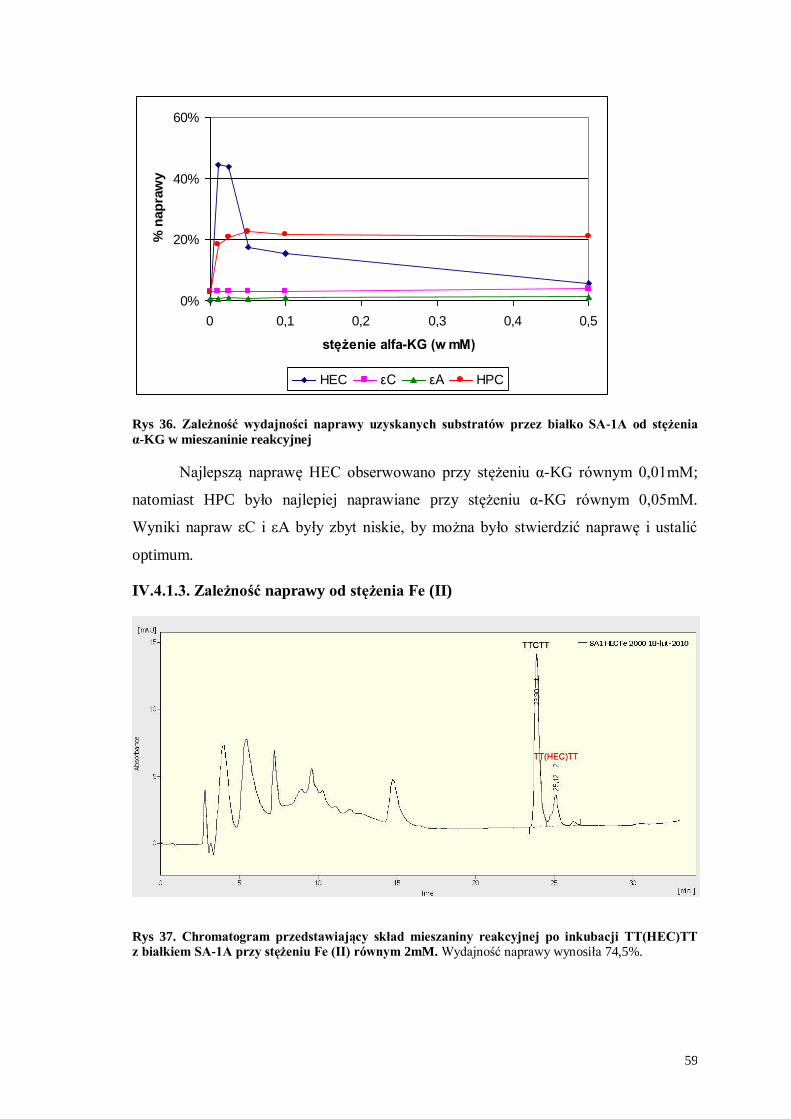
59
0%
20%
40%
60%
0 0,1 0,2 0,3 0,4 0,5
stężenie alfa-KG (w mM)
% n
ap
raw
y
HEC εC εA HPC
Rys 36. Zależność wydajności naprawy uzyskanych substratów przez białko SA-1A od stężenia
α-KG w mieszaninie reakcyjnej
Najlepszą naprawę HEC obserwowano przy stężeniu α-KG równym 0,01mM;
natomiast HPC było najlepiej naprawiane przy stężeniu α-KG równym 0,05mM.
Wyniki napraw εC i εA były zbyt niskie, by można było stwierdzić naprawę i ustalić
optimum.
IV.4.1.3. Zależność naprawy od stężenia Fe (II)
Rys 37. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji TT(HEC)TT
z białkiem SA-1A przy stężeniu Fe (II) równym 2mM. Wydajność naprawy wynosiła 74,5%.
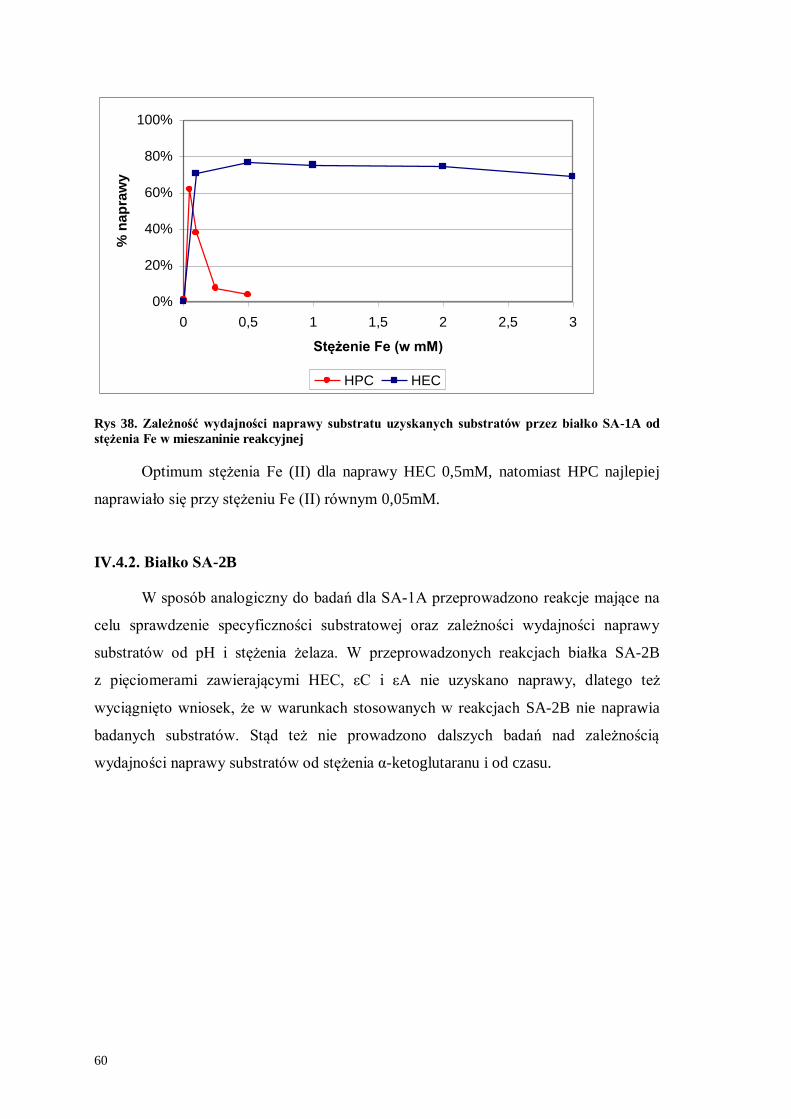
60
0%
20%
40%
60%
80%
100%
0 0,5 1 1,5 2 2,5 3
Stężenie Fe (w mM)
% n
ap
raw
y
HPC HEC
Rys 38. Zależność wydajności naprawy substratu uzyskanych substratów przez białko SA-1A od
stężenia Fe w mieszaninie reakcyjnej
Optimum stężenia Fe (II) dla naprawy HEC 0,5mM, natomiast HPC najlepiej
naprawiało się przy stężeniu Fe (II) równym 0,05mM.
IV.4.2. Białko SA-2B
W sposób analogiczny do badań dla SA-1A przeprowadzono reakcje mające na
celu sprawdzenie specyficzności substratowej oraz zależności wydajności naprawy
substratów od pH i stężenia żelaza. W przeprowadzonych reakcjach białka SA-2B
z pięciomerami zawierającymi HEC, εC i εA nie uzyskano naprawy, dlatego też
wyciągnięto wniosek, że w warunkach stosowanych w reakcjach SA-2B nie naprawia
badanych substratów. Stąd też nie prowadzono dalszych badań nad zależnością
wydajności naprawy substratów od stężenia α-ketoglutaranu i od czasu.

61
V Dyskusja
Białko AlkB jest enzymem, który naprawia DNA według unikalnego
mechanizmu, polegającego na usuwaniu uszkodzeń z zasady bez naruszania
integralności nici DNA, podczas gdy większość znanych mechanizmów naprawy polega
na usuwaniu i wymianie uszkodzonych fragmentów podwójnej helisy DNA. AlkB
utlenia grupy metylowe 1-metyloadeniny i 3-metylocytozyny w ssDNA (Trewick i wsp.
2002, Falnes i wsp. 2002), dsDNA oraz ssRNA (Aas i wsp. 2003). Efektywniej
naprawia uszkodzenia w ssDNA niż w dsDNA (Dinglay i wsp. 2000). Naprawia
również etenoadeninę, etenocytozynę oraz hydroksyetanocytozynę (Delaney i wsp.
2005, Mishina i wsp. 2005, Maciejewska i wsp. 2010). Badania prowadzone w
Zakładzie Biologii Molekularnej IBB przez prof. Jarosława Kuśmierka i dr Agnieszkę
Maciejewską wykazały, że białko AlkB naprawia również z dużą wydajnością
hydroksypropanocytozynę (Maciejewska i wsp., publikacja w przygotowaniu).
Niniejsza praca miała na celu ustalenie optymalnych warunkach naprawy
3,N4-α-hydroksyetanocytozyny i 3,N
4-α-hydroksypropanocytozyny przez AlkB oraz
określenie specyficzności substratowej homologów AlkB: SA-1A i SA-2B,
pochodzących z Streptomyces avermitilis. Te występujące w glebie bakterie, należące
do rzędu promieniowców, produkują wiele metabolitów wtórnych, w tym awermektyny,
które są wykorzystywane w medycynie jako czynniki antypasożytnicze (Ōmura
i współpracownicy 2001). Efektywność naprawy przez SA-1 i SA-2B badano dla
3,N4-α-hydroksyetanocytozyny (HEC), 3,N
4-α-hydroksypropanocytozyny (HPC),
1,N6-etenoadeniny oraz 3,N
4-etenocytozyny. Dla naprawianych przez te homologi
substratów ustalono optymalne warunki naprawy.
Pierwszym etapem badań było uzyskanie oczyszczonych białek
wykorzystywanych następnie do reakcji naprawy. Jak pokazują wyniki metoda,
polegająca na transformacji bakterii szczepu E. coli BL21 (DE3) plazmidem
pET28a/alkB, następnie ekspresji białka w szczepie E. coli BL21 (DE3) i oczyszczaniu
go na kolumnie niklowej HisTrap (Amersham) z wykorzystaniem urządzenia Akta
Prime, jest skuteczna i pozwala uzyskać aktywne enzymy, które można przechowywać
w temperaturze -800C i wykorzystywać do badań nad ich specyficznością substratową.
Drugim etapem badań była modyfikacja komercyjnie dostępnych pentamerów
DNA, które służyły jako substraty do reakcji naprawy przez białko AlkB i jego
homologi. Używane do modyfikacji pentamery składają się z 4 tymin i A lub C,

62
ponieważ tymina jako jedyna zasada nie ma egzocyklicznej grupy NH2 i nie tworzy
adduktów egzocyklicznych, ponieważ nie reaguje z aldehydem chlorooctowym
i akroleiną. Za pomocą ACR i CAA modyfikowano pentamery zawierające cytozynę.
W reakcji z akroleiną otrzymywano HPC, natomiast w reakcji
z aldehydem chloroocrowym otrzymywano HEC i εC. Pentamery zawierające adeninę
modyfikowano aldehydem chlorooctowym, w wyniku czego otrzymywano εA.
Najszybciej przebiega modyfikacja cytozyny aldehydem chlorooctowym. HEC i εC
otrzymuje się już po 2,5h inkubacji. Nieco wolniej (4h) przebiega modyfikacja
cytozyny akroleiną. Najdłużej natomiast trzeba inkubować mieszaninę reakcyjną
zawierającą pentamer z adeniną oraz aldehyd chlorooctowy. Jak wykazały badania mgr
Zuzanny Kaczmarskiej (praca mgr UW 2009), zależność stopnia przereagowania
TTATT do TTεATT od czasu jest liniowa przez niecałe 8 godzin. Po tym czasie,
reakcja zwalnia i powoli osiąga 100 % przereagowania w ciągu 23 godzin. Po inkubacji
interesujące nas produkty oczyszczano za pomocą preparatywnego rozdziału na HPLC.
W wyniku rozdziału na HPLC otrzymano czyste pochodne: TT(HEC)TT, TT(HPC)TT
oraz TT(εA)TT. Kolejnym etapem badań było ustalenie optimum naprawy HEC i HPC
przez AlkB. Badano następujące parametry: pH, stężenie Fe(II), stężenie α-KG oraz
czas reakcji. HEC okazał się być wydajnie naprawiany przez białko AlkB. Optymalne
pH mieszaniny reakcyjnej wyniosło 5,8. Najlepszą naprawę HEC przez AlkB
obserwowano przy stężeniu żelaza równym 1mM. Optymalnym stężeniem α-KG było
0,025mM. HEC najlepiej naprawiany był po czasie 30 minut (wydajność naprawy
wyniosła ponad 40%). HPC okazał się lepszym substratem dla AlkB niż HEC.
Najwyższa naprawa, jaką zaobserwowano, wynosiła 96,9%. Optymalne pH naprawy
HPC było wyższe niż dla HEC i wynosiło 7,4. Podobnie w przypadku α-KG, optymalne
stężenie było wyższe (0,05mM). Natomiast optymalne stężenie żelaza było niższe niż
dla HEC, wynosiło 0,1mM.
Uzyskane wyniki zostały porównane z wynikami badań nad specyficznością
substratową AlkB prowadzonymi przez dr Agnieszkę Maciejewską (Zakład Biologii
Molekularnej PAN), które uwzględniały również takie związki jak: m3C, εA, εC i εG.
Z danych przedstawionych w tabeli 2 wynika, że najlepszym substratem dla AlkB jest
m3C, natomiast najsłabiej naprawiane jest εC. Nie zaobserwowano naprawy εG.
Naprawa adduktów egzocyklicznych przez AlkB wymaga wyższego stężenia Fe(II)
i niższego stężenia α-KG, niż dla naprawy m3C.
63
Tabela 2. Zależność optymalnych warunków naprawy substratów AlkB od ich pKa (Maciejewska
·i wsp., plakat 2009).
Substrat
AlkB
pKa Optymalne pH Optymalne
stęż. Fe(II)
[µM]
Optymalne
stęż. α-KG
[µM]
Relatywna
wydajność
naprawy
m3C 9,0 8,0 50 500 +++++
HPC 8,6 7,5 100 50 +++
HEC 6,9 5,8 1000 25 ++
εA 3,9 4,6 3000 25 ++++
εC 3,7 4,6 5000 25 +
εG 2,2 – 2,5 Nie zaobserwowano naprawy
Z danych, które zaprezentowano w zeszłym roku w postaci plakatu na
konferencji Europejskiego Towarzystwa Mutagenów Środowiskowych we Florencji
wynika, że optimum pH dla naprawy substratów AlkB jest zależne od ich pKa
(Maciejewska i wsp., publikacja w przygotowaniu). pKa jest to wartość pH, przy której
połowa cząstek danego związku występuje w formie uprotonowanej, zaś druga połowa
w formie obojętnej. Dalsza protonacja odbywa się w niższym pH. Przyjmuje się, że
wszystkie cząsteczki danego związku są uprotonowane w pH niższym o 1 od pKa. Jak
wynika z danych przedstawionych w tabeli: m3C, HEC i HPC są naprawiane najlepiej
w pH niższym o ok. 1 od pKa, czyli wtedy gdy wszystkie ich cząsteczki są
uprotonowane. Sugeruje to, że centrum aktywne białka AlkB wiąże łatwiej
uprotonowane (dodatnio naładowane) formy substratów. W przypadku εA i εC
najlepszą naprawę obserwuje się przy pH 4,6, natomiast pKa wynosi odpowiednio 3,9
i 3,7. Zatem oba te substraty są całkowicie uprotonowane w pH<3. Prawdopodobnie
optimum pH wynosi 4,6, ponieważ w niższym pH enzym ulega dezaktywacji. Jednakże
w pH 4,6 ok. 15% cząsteczek εA i ok. 5% cząsteczek εC jest w formie uprotonowanej,
a ściślej każda z cząsteczek εA i εC jest przez odpowiednio 15% i 5% czasu ich trwania
w formie uprotonowanej. Lepsza naprawa εA może ponadto wynikać z jej dobrego
dopasowania do centrum aktywnego AlkB. Jak wynika z analizy badań
krystalograficznych przedstawionych w pracy Yu i współpracowników, w centrum
aktywnym AlkB, zawierającym substrat m3C, jest wolne miejsce, w którym dobrze
zmieściłaby cząsteczka εA (Yu i wsp. 2006).
Zebrane dane pokazują również, że im wyższe pH, tym wymagane stężenie
jonów żelaza (II) jest niższe. Być może wynika to z faktu, iż w niższym pH znajduje się
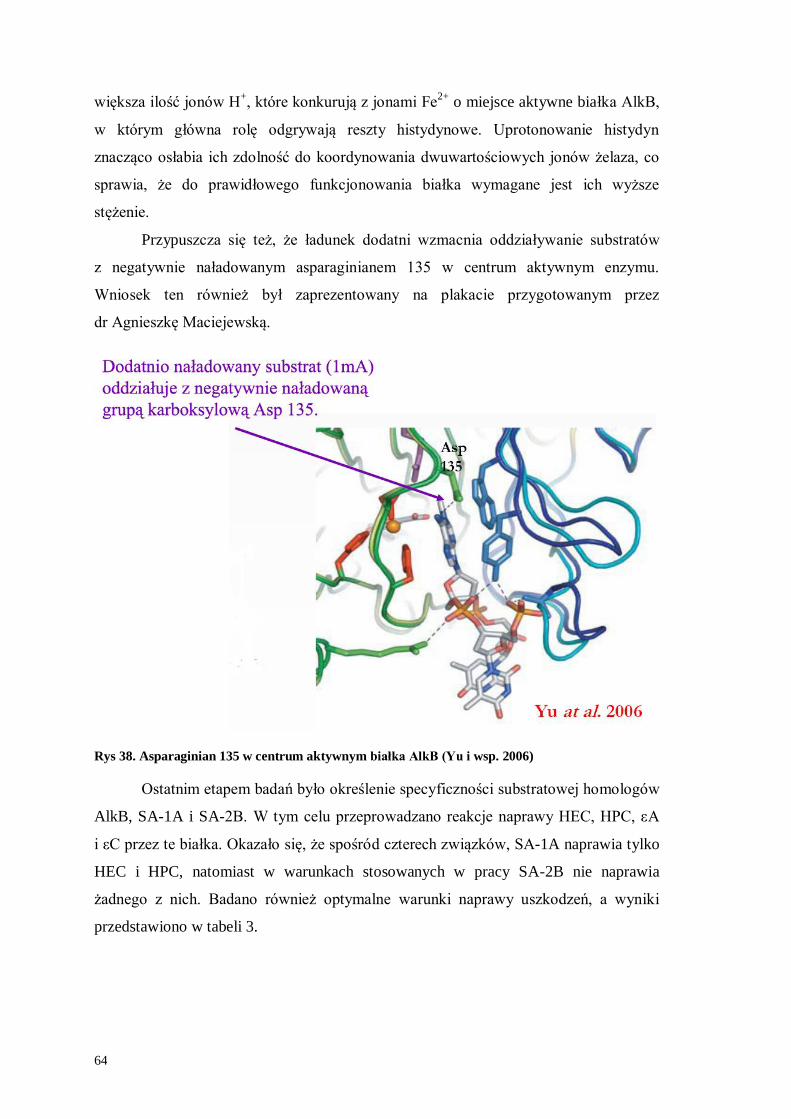
64
większa ilość jonów H+, które konkurują z jonami Fe
2+ o miejsce aktywne białka AlkB,
w którym główna rolę odgrywają reszty histydynowe. Uprotonowanie histydyn
znacząco osłabia ich zdolność do koordynowania dwuwartościowych jonów żelaza, co
sprawia, że do prawidłowego funkcjonowania białka wymagane jest ich wyższe
stężenie.
Przypuszcza się też, że ładunek dodatni wzmacnia oddziaływanie substratów
z negatywnie naładowanym asparaginianem 135 w centrum aktywnym enzymu.
Wniosek ten również był zaprezentowany na plakacie przygotowanym przez
dr Agnieszkę Maciejewską.
Rys 38. Asparaginian 135 w centrum aktywnym białka AlkB (Yu i wsp. 2006)
Ostatnim etapem badań było określenie specyficzności substratowej homologów
AlkB, SA-1A i SA-2B. W tym celu przeprowadzano reakcje naprawy HEC, HPC, εA
i εC przez te białka. Okazało się, że spośród czterech związków, SA-1A naprawia tylko
HEC i HPC, natomiast w warunkach stosowanych w pracy SA-2B nie naprawia
żadnego z nich. Badano również optymalne warunki naprawy uszkodzeń, a wyniki
przedstawiono w tabeli 3.
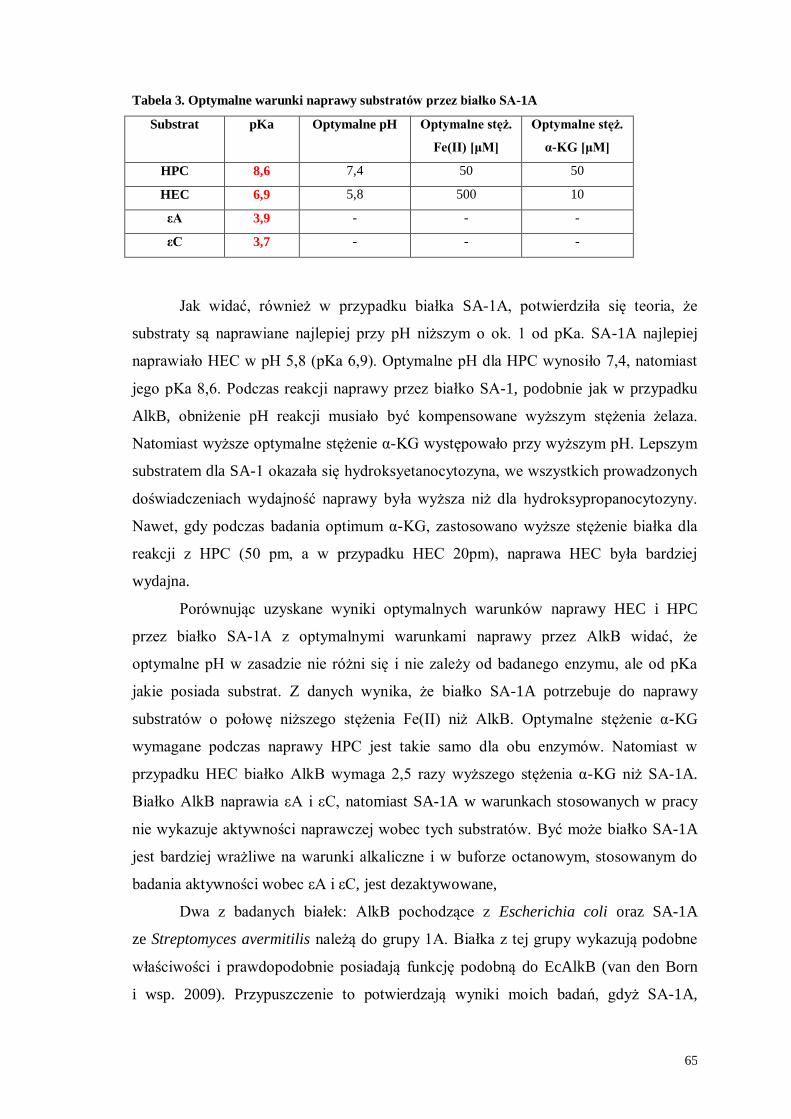
65
Tabela 3. Optymalne warunki naprawy substratów przez białko SA-1A
Substrat pKa Optymalne pH Optymalne stęż.
Fe(II) [μM]
Optymalne stęż.
α-KG [μM]
HPC 8,6 7,4 50 50
HEC 6,9 5,8 500 10
εA 3,9 - - -
εC 3,7 - - -
Jak widać, również w przypadku białka SA-1A, potwierdziła się teoria, że
substraty są naprawiane najlepiej przy pH niższym o ok. 1 od pKa. SA-1A najlepiej
naprawiało HEC w pH 5,8 (pKa 6,9). Optymalne pH dla HPC wynosiło 7,4, natomiast
jego pKa 8,6. Podczas reakcji naprawy przez białko SA-1, podobnie jak w przypadku
AlkB, obniżenie pH reakcji musiało być kompensowane wyższym stężenia żelaza.
Natomiast wyższe optymalne stężenie α-KG występowało przy wyższym pH. Lepszym
substratem dla SA-1 okazała się hydroksyetanocytozyna, we wszystkich prowadzonych
doświadczeniach wydajność naprawy była wyższa niż dla hydroksypropanocytozyny.
Nawet, gdy podczas badania optimum α-KG, zastosowano wyższe stężenie białka dla
reakcji z HPC (50 pm, a w przypadku HEC 20pm), naprawa HEC była bardziej
wydajna.
Porównując uzyskane wyniki optymalnych warunków naprawy HEC i HPC
przez białko SA-1A z optymalnymi warunkami naprawy przez AlkB widać, że
optymalne pH w zasadzie nie różni się i nie zależy od badanego enzymu, ale od pKa
jakie posiada substrat. Z danych wynika, że białko SA-1A potrzebuje do naprawy
substratów o połowę niższego stężenia Fe(II) niż AlkB. Optymalne stężenie α-KG
wymagane podczas naprawy HPC jest takie samo dla obu enzymów. Natomiast w
przypadku HEC białko AlkB wymaga 2,5 razy wyższego stężenia α-KG niż SA-1A.
Białko AlkB naprawia εA i εC, natomiast SA-1A w warunkach stosowanych w pracy
nie wykazuje aktywności naprawczej wobec tych substratów. Być może białko SA-1A
jest bardziej wrażliwe na warunki alkaliczne i w buforze octanowym, stosowanym do
badania aktywności wobec εA i εC, jest dezaktywowane,
Dwa z badanych białek: AlkB pochodzące z Escherichia coli oraz SA-1A
ze Streptomyces avermitilis należą do grupy 1A. Białka z tej grupy wykazują podobne
właściwości i prawdopodobnie posiadają funkcję podobną do EcAlkB (van den Born
i wsp. 2009). Przypuszczenie to potwierdzają wyniki moich badań, gdyż SA-1A,

66
podobnie jak AlkB, naprawia egzocykliczne addukty DNA: HEC i HPC. Białko SA-2B
należy natomiast do grupy 2B. Białka z tej grupy, takie jak: MT-2B, SC-2B, SA-2B
i XC-2B wykazują aktywność naprawczą, ale ich specyficzność substratowa jest dość
zróżnicowana. Niektóre z nich, tak jak XC-2B, wykazują wysoką aktywność naprawczą
dla uszkodzeń etenowych, inne natomiast dla metylowych. (van den Born i wsp. 2009).
Prawdopodobnie dlatego SA-2B nie wykazywało aktywności naprawczej w stosunku do
etenoadduktów, a być może, podobnie jak MT-2B, naprawia tylko uszkodzenia
metylowe.
W badaniach in vitro prowadzonych przez van den Borna i współpracowników
białko SA-1A wykazywało wysoką aktywność naprawczą wobec m3C występującego
w dsDNA, porównywalną do aktywności naprawczej AlkB. Naprawiało również εA
w ssDNA, jednak z dużo niższą aktywnością niż AlkB. Nie naprawiało natomiast m1A
w dsDNA oraz εA w dsDNA. Białko SA-2B wykazywało aktywność naprawczą
porównywalną do AlkB wobec m3C występującego w dsDNA, dużo niższą dla εA
w ssDNA oraz εA w dsDNA, nie naprawiało natomiast m1A w dsDNA (van der Born
2009). Jak widać wyniki te różnią się od tych uzyskanych przeze mnie. W przypadku
moich badań SA-1A i SA-2B nie naprawiały εA. Być może rozbieżność w wynikach
jest spowodowana różnicami w stosowanej metodzie. W moich badaniach
wykorzystywane były pentamery, natomiast w badaniach zespołu van der Borna
wykorzystywano dużo dłuższe oligonukleotydy. W badaniach norweskich naukowców
stosowano następujące składniki reakcji: 100pm białka, 16,6 nM substratu, 50mM Tris
HCl o pH 8,0, 2mM kwas askorbinowy, 100μM α-KG i 40μM FeSO4. Reakcja po 30
min inkubacji w 370C była przerywana poprzez dodanie 18mM EDTA, 0,4% SDS
i 0,4mg/ml proteinazy K. Następnie DNA było trawione przez enzym restrykcyjny
wrażliwy na modyfikację DNA, a produkcji reakcji były rozdzielane na 20%
denaturującym PAGE.
Badania nad białkami SA-1A i SA-2B były prowadzone w Zakładzie Biologii
Molekularnej również przez mgr Annę Domańską (praca mgr UW 2010).
W doświadczeniach przeprowadzonych przez nią SA-1A wykazywało aktywność
naprawczą wobec etenoadeniny i etenoguaniny, jednak o połowę niższą niż
w przypadku AlkB (ok. 35% naprawionych oligodeoksynukleotydów w dsDNA, przy
najwyższym stężeniu białka). Natomiast aktywność enzymatyczna SA-1A względem
etenocytozyny była zbyt niska, aby uznać ją za białko naprawcze dla tego uszkodzenia
(około 7,5%). SA-2B z dużą wydajnością naprawiało etenoguaninę (ok. 70%). Mniej

67
efektywnie naprawiana była etenoadenina (ok. 20%), natomiast naprawa etenocytozyny
była znikoma (poniżej 15%). Wyniki te nie są zbieżne z tymi uzyskanymi przeze mnie,
ponieważ w moich doświadczeniach SA-1A i SA-2B nie naprawiały etenocytozyny
i etenoadeniny. Należy jednak wziąć pod uwagę różnice w stosowanej metodzie. Mgr
Domańska po przeprowadzeniu reakcji badanego oligomeru z homologiem AlkB,
używała glikozylazy ze szlaku BER (MUG, HAP-1 lub ANPG), która usuwała
nienaprawione wcześniej uszkodzenia i pozostawiała miejsce
apurunowe/apirymidynowe (AP). Po rozdziale w żelu poliakrylamidowym obserwowała
wystąpienie lub brak fragmentacji nici DNA. Fragmentacja była spowodowana
pęknięciami w miejscu AP i świadczyła o braku naprawy. Korzystała również z dużo
dłuższych substratów niż te zastosowane przeze mnie. Reakcje prowadziła
w odmiennych warunkach pH i temperatury. Używała także dużo większych stężeń
badanego białka. Zdarzało się, że nadmiar enzymu doprowadzał do zahamowania
procesu naprawy.
Liczne badania wykazały, że wiele z dioksygenaz z rodziny AlkB bierze udział
w naprawie uszkodzeń DNA i RNA. Różnią się one specyficznością substratową oraz
wydajnością, z jaką naprawiają uszkodzone zasady. Prawdopodobnie dla dioksygenaz
naprawiających DNA z dużą efektywnością, usuwanie uszkodzeń zasad DNA jest
główną funkcją. Natomiast dla białek działających z dużo niższą efektywnością, być
może jest to funkcja drugorzędna i pełnią one również inne ważne funkcje w komórce,
takie jak modyfikacja RNA. Wydaje się, że za naprawę DNA i modyfikacja RNA
w komórkach eukariotycznych odpowiadają różne dioksygenazy, natomiast
w komórkach bakteryjnych jeden enzym może pełnić obie te funkcje. W przyrodzie
występują bakterie, które posiadają tylko jeden homolog AlkB np.: E. coli. Inne
natomiast (np.: Streptomyces avermitilis, Streptomyces coelicolor, Xanthomonas
campestris) posiadają dwa lub więcej homologów AlkB. Może to wskazywać na fakt, iż
u niektórych bakterii nastąpiło częściowe rozdzielenie funkcji naprawy DNA
i modyfikacji RNA przez białka z rodziny AlkB.
W niniejszej pracy wykazano aktywność naprawczą dwóch, z trzech badanych
dioksygenaz z rodziny AlkB wobec egzocyklicznych adduktów zasad DNA. Najwyższą
efektywność naprawy uszkodzonych zasad DNA obserwuje się w przypadku EcAlkB,
niższą efektywnością charakteryzowało się białko SA-1A, natomiast SA-2B nie
naprawiała uszkodzeń etenowych w warunkach stosowanych w pracy.

68
VI Wnioski
Na podstawie uzyskanych wyników sformułowano następujące wnioski:
1. Stosowana w badaniach metoda uzyskiwania oczyszczonych białek jest skuteczna
i pozwala na otrzymanie enzymatycznie aktywnych białek.
2. Białko AlkB naprawia takie uszkodzenia jak HEC i HPC. HPC jest nowym,
dotychczas niepublikowanym substratem białek typu AlkB.
3. Optymalne warunki naprawy przez AlkB:
- dla TT(HEC)TT – pH 5,8, Fe 1mM, α-KG 0,025mM, czas 30min;
- dla TT(HPC)TT – pH 7,5, Fe 0,1mM, α-KG 0,05mM, czas 30min.
4. Białko SA-1A jest homologiem AlkB naprawiającym takie uszkodzenia jak HEC
i HPC.
5. Optymalne warunki naprawy przez SA-1A:
- dla TT(HEC)TT – pH 5,8, Fe 0,01mM, α-KG 0,5mM, czas 10min;
- dla TT(HPC)TT – pH 7,4, Fe 0,05mM (50μM), α-KG 0,05mM, czas 10min;
- dla TT(εA)TT – brak naprawy w stosowanych warunkach reakcji;
- dla TT(εC)TT – brak naprawy w stosowanych warunkach reakcji.
5. Białko SA-2B nie naprawia egzocyklicznych uszkodzeń zasad DNA w warunkach
stosowanych w pracy.
6. Badane białka różnią się specyficznością substratową względem badanych
egzocyklicznych adduktów oraz efektywnością naprawy uszkodzeń w DNA.

69
Aneks
Wyniki uzyskane w tej pracy były prezentowane w postaci plakatu na konferencji
Europejskiego Towarzystwa Mutagenów Środowiskowych
40th annual meeting of The European Environmental Mutagen Society (EEMS),
September 15-18 2010, Oslo
NEW SUBSTRATES FOR ALKB DIOXYGENASE FROM ESCHERICHIA COLI
AND OTHER BACTERIA
Maciejewska1, Agnieszka M; Nieminuszczy
1, Jadwiga; van den Born
2, Edwin;
Krowisz1, Beata; Wrona
1, Joanna; Grzesiuk
1, Elżbieta; Falnes
2,3, Pål Ø and Kuśmierek
1,
Jarosław T
1Institute of Biochemistry and Biophysics, Polish Academy of Sciences, 02-106 Warsaw,
5A Pawińskiego Str, Poland
2Department of Molecular Biosciences, University of Oslo, PO Box 1041 Blindern,
0316 Oslo, Norway,
3Centre for Molecular Biology and Neuroscience, Institute of Medical Microbiology,
Oslo University Hospital and University of Oslo, N-0027 Oslo, Norway,
Name and E-mail of presenting author: Agnieszka M. Maciejewska;
Keywords: DNA repair; AlkB dioxygenase homologues; exocyclic adducts to DNA
bases;
Conference: EEMS
The AlkB-type proteins are repair enzymes which remove alkyl lesions from bases via
an oxidative mechanism restoring native DNA structure. Recently, it was found that
they also repair DNA etheno()-adducts. Exocyclic adducts (-adducts among them)
can originate endogenously from reaction of lipid peroxidation products: α, β-
unsaturated- and epoxy- aldehydes with DNA bases. They are also formed upon
exposure to various industrial carcinogens: e.g., vinyl chloride metabolites,

70
chloroethylene epoxide and chloroacetaldehyde (CAA), or dietary and
environmental toxin acrolein (ACR). We reacted 5-mer oligodeoxynucleotides,
TTXTT, where X is C or A, with CAA, ACR, croton aldehyde or MMS, and we
obtained 5-mers containing modified cytosines: 3,N4-α-hydroxyethano- (HEC),
3,N4-etheno- (εC), 3,N
4-α-hydroxypropano- (HPC), 3,N
4-α-hydroxy-γ-methyl-
propano- (mHPC) or3-methyl- (3mC), respectively, or 1,N6-ethenoadenine (εA).
The hydrolytic deamination of TT(3mC)TT at pH 9.2 gave TT(3mU)TT. All
prepared modified 5-mers were studied as substrates for E. coli AlkB protein. Using
HPLC technique, which allows to separate the modified from unmodified (repaired)
oligomer, we have found that with exception of mHPC, all the modifications are
repaired by E. coli AlkB dioxygenase. Appearance of repaired oligomer was also
confirmed by mass spectrometry. We observed that the repair efficiency is pH
dependent and correlated with pKa value of repaired adduct what suggests that
enzyme recognizes protonated form of substrate. The efficiency of repair decreases
in the following order 3mC>εA>HPC>HEC>εC>3mU. The repair of 3mU can be
observed only at a high enzyme/substrate ratio. We have also checked the repair of
HEC, εC, HPC and εA by following bacterial AlkB homologues: Streptomyces
avermitilis SA-1A and SA-2A, Streptomyces coelicolor SC-1A and SC-2A, and
Mycobacterium tuberculosis MT-2B. All of them were known to repair 3mC. The
SA-1A and SC-1A proteins repair HEC and HPC efficiently, but εC weakly; εA is
repaired by SC-1A but not by SA-1A protein. We did not observe any repair activity
of SA-2B, SC-2B and MT-2B against the studied exocyclic adducts.

71
Spis literatury
Aas P.A., Otterlei M., Falnes P.Ø., Vågbø C.B., Skorpen F., Akbari M., Sundheim
O., Bjørås M., Slupphaug G., Seeberg E., Krokan H.E. (2003) Human and bacterial
oxidative demethylases repair alkylation damage in both RNA and DNA. Nature 421:
859–863
Akasaka S., Guengerich F.P. (1999) Mutagenicity of site-specifically located 1,N2-
ethenoguanine in Chinese hamster ovary cell chromosomal DNA. Chem. Res. Toxicol.
12: 501–507
Aravind L., Koonin E.V. (2001) The DNA-repair protein AlkB, EGL-9, and leprecan
define new families of 2-oxoglutarate- and iron-dependent dioxygenases. Genome
Biology 2(3): 0007.1-0007.8
Aravind L., Koonin E.V. (2001) The DNA-repair protein AlkB, EGL-9, and leprecan
define new families of 2-oxoglutarate- and iron-dependent dioxygenases. Genome Biol.
2 RESEARCH0007
Barbin A. (2000) Etheno-adduct-forming chemicals: from mutagenicity testing to
tumor mutation spectra. Mutation Research 462: 55-69
Barbin A., Bartsch H., Leconte P., Radman M. (1981) Studies on the miscoding
properties of 1,N6-ethenoadenine and 3,N
4-ethenocytosine, DNA reaction products of
vinyl chloride metabolites, during in vitro DNA synthesis. Nucleic Acids. Res. 9: 375–
387
Basu A.K., Wood M.L., Niedernhofer L.J., Ramos L.A., Essigmann J.M. (1993)
Mutagenic and genotoxic effects of three vinyl chloride-induced DNA lesions: 1,N6-
ethenoadenine, 3,N4-ethenocytosine, and 4-amino-5-(imidazol-2-yl) imidazole
Biochemistry 32: 12793–12801
Borys E., Mroczkowska-Słupska M.M., J.T. Kusmierek (1994) The induction of
adaptive response to alkylating agents in Escherichia coli reduces the frequency of
specific C→T mutations in chloroacetaldehyde-treated M13 glyU phage. Mutagenesis
9: 407–410
Borys-Brzywczy E., Arczewska K.D., Saparbaev M., Hardeland U., Schar P.,
Kusmierek J.T. (2005) Mismatch dependent uracil/thymine-DNA glycosylases excise
exocyclic hydroxyethano and hydroxypropano cytosine adducts. Acta Biochim. Pol. 52:
149-65

72
Delaney J.C., Smeester L., Wong C., Frick L.E., Taghizadeh K., Wishnok J.S.,
Drennan C.L., Samson L.D., Essigmann J.M. (2005) AlkB reverses etheno DNA
lesions caused by lipid oxidation in vitro and in vivo. Nat Struct Mol Biol 12: 855-60
Dina C., Meyre D., Gallina S., Durand E., Körner A., Jacobson P., Carlsson
L.M.S., Kiess W., Vatin V., Lecoeur C., Delplanque J., Vaillant E., Pattou F., Ruiz
J., Weill J., Levy-Marchal C., Horber F., Potoczna N., Hercberg S., Stunff C.L.,
Bougnères P., Kovacs P., Marre M., Balkau B., Cauchi S., Chèvre, J., Froguel, P.
(2007) Variation in FTO contributes to childhood obesity and severe adult obesity. Nat.
Genet. 39: 724–726
Dinglay S., Trewick S.C., Lindahl T., Sedgwick B. (2000) Defective processing of
methylated single-stranded DNA by E. coli AlkB mutants. Genes Dev. 14: 2097–2105
Domańska A. (2010) Naprawa etenoadduktów DNA przez bakteryjne dioksygenazy
AlkB. Praca magisterska UW
Douki T., Odin F., Caillat S., Favier A., Cader J. (2004) Predominance of the 1,N2-
propano-2’-deoksyguanosine adduct among 4-hydroxy-2-nonenal induced DNA-lesions.
Free Radic. Biol. Med. 37: 62-70
Drablos F., Feyzi E., Aas P.A., Vaagbo C.B., Kavli B., Bratlie M.S., Pena-Diaz J.,
Otterlei M., Slupphaug G., Krokan H.E. (2004) Alkylation damage in DNA and RNA
– repair mechanisms and medical significance. DNA Repair (Amst) 3: 1389-407
Duncan T., Trewick S.C., Koivisto P., Bates P.A., Lindahl T., Sedgwick B. (2002)
Reversal of DNA alkylation damage by two human dioxygenases. Proc. Natl. Acad. Sci.
USA 99: 16660–16665
Falnes P.Ø., Bjørås M., Aas P.A., Sundheim O., Seeberg E. (2004) Substrate
specificities of bacterial and human AlkB proteins. Nucleic Acids Res. 32: 3456–3461
Falnes P.Ø., Johansen R.F., Seeberg E. (2002) AlkB-mediated oxidative
demethylation reverses DNA damage in Escherichia coli. Nature 419: 178-182
Falnes P.Ø., Rognes T. (2003) DNA repair by bacterial AlkB proteins. Res Microbiol.
Oct;154(8):531-8
Falnes P.Ø., van den Born E., Meza T.J. (2009) Demethylation of DNA and RNA by
AlkB Proteins. In Grosjean H (ed.), DNA and RNA Modification Enzymes: Structure,
Mechanism, Function and Evolutio, Landes Bioscience, Austin, USA
Frayling T.M., Timpson N.J., Weedon M.N., Zeggini E., Freathy R.M., Lindgren
C.M., Perry J.R.B., Elliott K.S., Lango H., Rayner N.W., Shields B., Harries L.W.,
Barrett J.C., Ellard S., Groves C.J., Knight B., Patch A., Ness A.R., Ebrahim S.,

73
Lawlor D.A., Ring S.M., Ben-Shlomo Y., Jarvelin, M., Sovio U., Bennett A.J.,
Melzer D., Ferrucci L., Loos R.J.F., Barroso I., Wareham N.J., Karpe F., Owen
K.R., Cardon L.R., Walker M., Hitman G.A., Palmer C.N.A., Doney A.S.F.,
Morris A.D., Smith G.D., Hattersley A.T., McCarthy, M.I. (2007) A common variant
in the FTO gene is associated with body mass index and predisposes to childhood and
adult obesity. Science 316: 889–894
Gerken T., Girard C.A., Tung Y.L., Webby C.J., Saudek V., Hewitson K.S., Yeo
G.S.H., McDonough M.A., Cunliffe S., McNeill L.A., Galvanovskis J., Rorsman P.,
Robins P., Prieur X., Coll A.P., Ma M., Jovanovic Z., Farooqi I.S., Sedgwick B.,
Barroso I., Lindahl T., Ponting C.P., Ashcroft F.M., O_Rahilly S., Schofield C.J.
(2007) The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic
acid demethylase. Science 318: 1469–1472
Golding B.T., Slaich P.K., Kennedy G., Bleasdale C., Waston W.P. (1996)
Mechanisms of formation of adducts from reactions of glycidaldehyde with 2’-
deoxyguanosise and/or guanosine. Chem Res Toxicol 9: 147-57
Gros L., Ishchenko A.A., Saparbaev M. (2003) Enzymology of repair of etheno-
adducts. Mutat Res 531: 219-29
Guengerich F.P., Raney V.M. (1992) Formation of etheno adducts of adenosine and
cytidine from 1-halooxiranes. Evidence for a mechanism involving initial reaction with
the endocyclic nitrogen atoms. J. Am. Chem. Soc. 3: 1074–1080
Hall J.A., Saffhill R., Green T., Hathway D.E. (1981) The induction of errors during
in vitro DNA synthesis following chloroacetaldehyde treatment of poly (dA–dT) and
poly (dC–dG) templates. Carcinogenesis 2: 141–146
Hecht S.S. (2002) Human urinary carcinogen metabolites: biomarkers for investigating
tobacco and cancer. Carcinogenesis. Jun;23(6):907-22
Hecht, S.S., McIntee, E.J., Wang, M. (2001) New DNA adducts of crotonaldehyde
and acetaldehyde. Toxicology 166: 31-36
Jacobsen J.S., Perkins C.P., Callahan J.T., Sambamurti K., Humayun M.Z. (1989)
Mechanisms of mutagenesis by chloroacetaldehyde. Genetics 121: 213–222
Kaczmarska Z. (2009) Nowe substraty dioksygenazy AlkB z Escherichia coli. Praca
magisterska UW
Kataoka H., Yamamoto Y., Sekiguchi M. (1983) A new gene (alkB) of Escherichia
coli that controls sensitivity to methyl methane sulfonate. J. Bacteriol. 153: 1301–1307

74
Kowalczyk, P., Cieśla, J.M., Komisarski, M., Kuśmierek, J.T., Tudek, B. (2004)
Long chain adducts of trans-4-hydroxy-2-nonenal to DNA bases cause recombination,
base substitution and frameshift mutations in M23 phage. Mutat. Res. 550: 33-48
Krwawicz J. Arczewska K.D., Speina E., Maciejewska A., Grzesiuk E. (2007)
Bacterial DNA repair genes and their eukaryotic homologues: 1. Mutations in genes
involved in base excision repair (BER) and DNA-end processors and their implication
in mutagenesis and human disease. Acta Biochim Pol 54(3): 414-434
Krzyżosiak W.J., Biernat J., Ciesiołka J., Górnicki P., Wiewiórowski M. (1979)
Further studies on adenosine and cytidine reaction with chloroacetaldehyde, a new
support for the cyclic carbinolamine structure of the stable reaction intermediate and
its relevance to the reaction mechanism and tRNA modification. Polish J Chem 53:
243–52
Kurowski M.A., Bhagwat A.S., Papaj G., Bujnicki J.M. (2003) Phylogenomic
identification of five new human homologs of the DNA repair enzyme AlkB. BMC
Genomics 4: 48
Kusmierek J.T., Singer B. (1982) Chloroacetaldehyde-treated riboand
deoxyribopolynucleotides: 1. Reaction products. Biochemistry 21: 5717–5722
Langouet S., Muller M., Guengerich F.P. (1997) Misincorporation of dNTPs opposite
1,N2-ethenoguanine and 5,6,7,9-tetrahydro-7-hydroxy-9-oxoimidazo[1,2-α]purine in
oligonucleotides by Escherichia coli polymerases I exo- and II exo-, T7 polymerase exo-
, human immunodeficiency virus-1 reverse transcriptase, and rat polymerase β.
Biochemistry 36: 6069–6079.
Lee D.-H. , Jin S.-G., S. Cai, Chen Y., Pfeifer G.P., O’Connor T.R. (2005) Repair of
methylation damage in DNA and RNA by mammalian AlkB homologues. J. Biol. Chem.
280: 39448–39459
Lu S.C. (2000) S-Adenosylmethionine. Int J Biochem Cell Biol 32: 391–395
Lutz W.K. (1990) Endogenous genotoxic agents and processes as a basis of
spontaneous carcinogenesis. Mutat Res 238: 287–295
Ma L., Hoeijmakers J.H.J., van der Eb A.J. (1995) Mammalian nucleotide excision
repair. Biochem Bioph Acta 1242: 137-64
Maciejewska A.M., Ruszel K.P., Nieminuszczy J., Lewicka J., Sokołowska B.,
Grzesiuk E., Kuśmierek J.T. (2010) Chloroacetaldehyde-induced mutagenesis in
Escherichia coli: The role of AlkB protein in repair of 3,N4-ethenocytosine and 3,N
4-α
hydroxyethanocytosine. Mutation Research 684: 24-34

75
Madigan M.T., Martinko J.M., Parker J. (2003) Brock Biology of Microorganisms.
Marnett L.J. (1999) Lipid peroxidation – DNA damage by malondialdehyde. Mutat.
Res. 424: 83–95
Marnett L.J. (2002) Oxy radicals, lipid peroxidation and DNA damage. Toxicology
181/182: 219–222
Marnett L.J., Riggins J.N., West J.D. (2003) Endogenous generation of reactive
oxidants and electrophiles and their reactions with DNA. J. Clin. Invest. 111: 583–593
Matulewicz, Ł., Przybyszewski, W.M. (2004) Wpływ lipidów na aktywność polimeraz
DNA. Post. Biol. Kom. 31: 277-284
Mishina Y., He C. (2006) Oxidative dealkylation DNA repair mediated by the
mononuclear non-heme iron AlkB proteins. Journal of Inorganic Biochemistry 100:
670–678
Mishina Y., Yang C.G, He C. (2005) Direct repair of the exocyclic DNA adduct 1,N6-
ethenoadenine by the DNA repair AlkB proteins. J Am Chem Soc 127(42): 14594–5
Modrich P., Lahue R. (1996) Mismatch repair in replication fidelity, genetic
recombination, and cancer biology. Annu Rev Biochem 65: 101-33
Moller P., Wallin H. (1998) Adduct formation, mutagenesis and nucleotide excision
repair of DNA damage produced by reactive oxygen species and lipid peroxidation
product. Mutat. Res. 410: 271–290
Mroczkowska-Słupska M. Kuśmierek J.T. (1994) Molekularne podstawy
mutagennego działania chlorku winylu. Postępy Biochemii 40(1):31-39
Muller T.A., Meek K. and Hausinger R.P. (2010) Human AlkB homologue 1 (ABH1)
exhibits DNA lyase activity at abasic sites. DNA Repair (Amst), 9, 58-65.
Nair V., Offerman R.J. (1985) Ring-extended products from the reaction of epoxy
carbonyl compounds and nucleic acid bases. J. Org. Chem. 50: 5627-5631
Nath, R.G., Ocando, J.E., Chung, F.L. (1996) Detection of
1,N2propanodeoxyguanosine adducts as potential endogenous DNA lesions in rodent
and human tissues. Cancer Research 56: 452-456
Niederhofer L.J., Daniels J.S., Rauzer C.A., Greene R.E., Marnett L.J. (2003)
Malonaldehyde, a product of lipid peroxidation is mutagenic in human cells. J. Biol.
Chem. 278: 31426–31433
Nieminuszczy J., Grzesiuk E. (2007) Bacterial DNA repair genes and their eukaryotic
homologues: 3. AlkB dioxygenase and Ada methyltransferase in the direct repair of
alkylated DNA. Acta Biochim Pol 54(3): 459-68

76
Ōmura S., Ikeda H., Ishikawa J., Hanamoto A., Takahashi C., Shinose M.,
Takahashi Y., Hiroshi H., Nakazawa H., Osonoe T., Kikuchi H., Shibai T., Sakaki
Y., Hattori M. Genome sequence of an industrial microorganism Streptomyces
avermitilis: Deducing the ability of producing secondary metabolites. PNAS October 9,
2001 vol. 98 no. 21 12215–12220
Palejwala V.A. Rzepka , R.W., Humayun M.Z. (1993) UV irradiation of Escherichia
coli modulates mutagenesis at a sitespecific ethenocytosine residue on M13 DNA.
Evidence for an inducible recA-independent effect. Biochemistry 32: 4112–4120.
Palejwala V.A. Rzepka , R.W., Simha D., Humayun M.Z. (1993) Quantitative
multiplex sequence analysis of mutational hot spots. Frequency and specificity of
mutations induced by a site-specific ethenocytosine in M13 viral DNA. Biochemistry 32:
4105–4111
Pastoriza Gallego M., Sarasin A. (2003) Transcription-coupled repair of 8-
oxoguanine in human cells and its defi ciency in some DNA repair diseases. Biochimie
85: 1073–1082
Pietrzykowska I., Krwawicz J. (1999) Mechanizmy naprawy DNA u bakterii i
człowieka. Kosmos 245: 315-28
Plastaras J.P., Riggins J.N., Otteneder M., Marnett L.J. (2000) Reactivity and
mutagenicity of endogenous DNA oxopropenylating agents base propenals,
malonaldehyde,and N (epsilon)-oxopropenallysine. Chem. Res. Toxicol. 13: 1235–1242
Prentice Hall, Upper Saddle River, NJ
Prescott A.G., Lloyd M.D. (2000) The iron (II) and 2-oxoacid-dependent dioxygenases
and their role in metabolism. Nat Prod Rep 17: 367-83
Przybyszewski W.M., Kasperczyk J., Stokłosa K., Bkhiyan A. (2005) Uszkodzenia
DNA powodowane przez produkty peroksydacji lipidów. Postepy Hig. Med. Dosw. 59:
75-81
Ringvoll J., Nordstrand L.M., Vågbø, C.B., Talstad V., Reite K., Aas P.A.,
Lauritzen K.H., Liabakk N.B., Bjørk A., Doughty A., Falnes P.Ø., Krokan H.E.,
Klungland, A. (2006) Repair deficient mice reveal mABH2 as the primary oxidative
demethylase for repairing 1meA and 3meC lesions in DNA. EMBO J. 25: 2189–2198
Roszkowski K. (2002) Mechanizmy naprawy oksydacyjnych uszkodzeń DNA.
Współczesna Onkologia 6:6, 360-365
Ryle M.J., Hausinger R.P. (2002) Non-heme iron oxygenases. Curr Opin Chem Biol 6:
193-201

77
Sancar A. (1996) DNA excision repai.r Ann Rev Biochem 65: 43-81
Sanchez-Pulido L., Andrade-Navarro M.A. (2007) The FTO (fat mass and obesity
associated) gene codes for a novel member of the non-heme dioxygenase superfamily.
BMC Biochem. 8: 23
Schofield C.J., Zhang Z. (1999) Structural and mechanistic studies on 2-oxoglutarate-
dependent oxygenases and related enzymes. Curr Opin Struct Biol 9: 722-31.
Scott L.J., Mohlke K.L., Bonnycastle L.L., Willer C.J., Li Y., Duren W.L., Erdos
M.R., Stringham H.M., Chines P.S., Jackson A.U., Prokunina-Olsson L., Ding C.J.,
Swift A.J., Narisu N., Hu T., Pruim R., Xiao R., Li X.Y., Conneely K.N., Riebow
N.L., Sprau A.G., Tong M., White P.P., Hetrick K.N., Barnhart M.W., Bark C.W.,
Goldstein J.L., Watkins L., Xiang F., Saramies J., Buchanan T.A., Watanabe
R.M., Valle T.T., Kinnunen L., Abecasis G.R., Pugh E.W., Doheny K.F., Bergman
R.N., Tuomilehto J., Collins F.S., Boehnke M. (2007) A genome-wide association
study of type 2 diabetes in finns detects multiple susceptibility variants. Science 316:
1341–1345
Sedgwick B., Bates P.A., Paik J., Jacobs S.C., Lindahl T. (2007) Repair of alkylated
DNA: recent advances. DNA Repair (Amst) 6: 429–442
Shibutani S., Suzuki N., Matsumoto Y., Grollman A.P. (1996) Miscoding properties
of 3,N4-etheno-2’-deoxycytidine in reactions catalyzed by mammalian DNA
polymerases. Biochemistry 35: 14992–14998
Singer B. (1976) All oxygens in nucleic acids react with carcinogenic ethylating agents.
Nature 264: 333–339
Singer B., Kusmierek J.T., Fraenkel-Conrat H. (1983) In vitro discrimination of
replicases acting on carcinogen-modified polynucleotide templates. Proc. Natl. Acad.
Sci. U.S.A. 80: 969–972.
Śliwiński T., Błasiak J. (2005) Naprawa DNA przez wycinanie zasad. Postępy
Biochemii tom 51, nr 2, str. 120-128
Sodum R.S., Chung F.L. (1989) Structural characterization of adducts formed in the
reaction of 2,3-epoxy-4-hydroxynonanal with deoxyguanosine. Chem Res Toxicol 2:
23-8
Sodum R.S., Chung F.L. (1991) Stereoselective formation of in vitro nucleic acid
adducts by 2,3-epoxy-4-hydroxynonanal. Cancer Res 51(1): 137-43

78
Spengler S., Singer B. (1981) Transcriptional errors and ambiguity resulting from the
presence of 1,N6-ethenoadenosine or 3,N
4-ethenocytidine in polyribonucleotides.
Nucleic Acids Res. 9: 365–373
Sundheim O., Vågbø,C.B., Bjørås M., Sousa M.M.L., Talstad V., Aas P.A., Drabløs
F., Krokan H.E., Tainer J.A., Slupphaug G. (2006) Human ABH3 structure and key
residues for oxidative demethylation to reverse DNA/RNA damage. EMBO J. 25: 3389–
3397
Trewick S.C., Henshaw T.F., Hausinger R.P., Lindahl T., Sedgwick B. (2002)
Oxidative demethylation by Escherichia coli AlkB directly reverts DNA base damage.
Nature 419: 174-7
van den Born E., Bekkelund A., Moen M.N., Omelchenko M.V., Klungland A.,
Falnes P.Ø. (2009) Bioinformatics and functional analysis define four distinct groups
of AlkB DNA-dioxygenases in bacteria. Nuc Acid Res 37: 7124-36
Wei Y.F., Carter K.C., Wang R.P., Shell B.K. (1996) Molecular cloning and
functional analysis of a human cDNA encoding an Escherichia coli AlkB homolog, a
protein involved in DNA alkylation damage repair. Nucleic Acids Res 24: 931–937
Westbye M.P., Feyzi E., Aas P.A., Vagbo C.B., Talstad V.A., Kavli B., Hagen L.,
Sundheim O., Akbari M., Liabakk N.B., Slupphaug G., Otterlei M. and Krokan,
H.E. (2008) Human AlkB homolog 1 is a mitochondrial protein that demethylates 3-
methylcytosine in DNA and RNA. J. Biol. Chem., 283, 25046-25056
Wood R.D. (1996) DNA repair in Eukaryotes. Ann Rev Biochem 65: 135-67
Yang C.G., Yi C., Duguid E.M., Sullivan C.T., Jian X., Rice P.A., He C. (2008)
Crystal structures of DNA/RNA repair enzymes AlkB and ABH2 bound to dsDNA.
Nature 452: 961–65
Yip K.F., Tsou K.C. (1973) Synthesis of fluorescent adenosine derivatives.
Tetrahedron Lett. 33: 3087–3090
Yu B., Edstrom W.C., Benach J., Hamuro Y., Weber P.C., Gibney B.R., Hunt J.F.
(2006) Crystal structures of catalytic complexes of the oxidative DNA/RNA repair
enzyme AlkB. Nature 439: 879-84

79
Spis rysunków
Rys 1. Aktywacja metaboliczna chlorku winylu .......................................................... 14
Rys 2. Powstawanie eteno- i propanoadduktów pod wpływem produktów peroksydacji
lipidów ........................................................................................................................ 17
Rys 3. Wzory chemiczne etenoadduktów powstałych pod wpływem aldehydu
chlorooctowego ........................................................................................................... 19
Rys 4. Wzory chemiczne propanoadduktów akroleiny i aldehydu krotonowego z
deoksycytozyną ........................................................................................................... 20
Rys 5. Struktura kompleksu białka AlkB z Fe(II), α-ketoglutaranem i metylowanym
trójnukleotydem .......................................................................................................... 22
Rys 6. Mechanizm naprawy uszkodzeń alkilacyjnych przez białko AlkB..................... 23
Rys 7. Zdjęcie żelu z frakcjami po oczyszczaniu białka SA-2B. .................................. 42
Rys 8. Chromatogram przedstawiający analityczny rozdział produktów reakcji
modyfikacji pentameru TTCTT (T/C) aldehydem chlorooctowym............................... 43
Rys 9. Chromatogram przedstawiający analityczny rozdział produktów reakcji
modyfikacji pentameru TTCTT (T/C) akroleiną .......................................................... 44
Rys 10. Chromatogram przedstawiający preparatywny rozdział produktów reakcji
modyfikacji pentameru TTCTT (T/C) aldehydem chlorooctowym. .............................. 44
Rys 11. Chromatogram przedstawiający preparatywny rozdział produktów reakcji
modyfikacji pentameru TTCTT (T/C) akroleiną .......................................................... 45
Rys 12. Chromatogram przedstawiający analityczny rozdział produktów reakcji
modyfikacji pentameru TTATT (T/A) aldehydem chlorooctowym .............................. 46
Rys 13. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HEC)TT z białkiem AlkB w pH 5,8.. .................................................................... 47
Rys 14. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od pH
buforu w mieszaninie reakcyjnej ................................................................................ 47
Rys 15. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HEC)TT z białkiem AlkB przy stężeniu Fe (II) równym 0,5mM.. ........................ 48
Rys 16. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od
stężenia Fe w mieszaninie reakcyjnej.......................................................................... 48
Rys 17. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HEC)TT z białkiem AlkB przy stężeniu α-KG równym 0,05mM .......................... 49
Rys 18. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od
stężenia α-KG w mieszaninie reakcyjnej .................................................................... 49
Rys 19. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HEC)TT z białkiem AlkB w czasie 30min ............................................................ 50
Rys 20. Zależność wydajności naprawy substratu TT(HEC)TT przez białko AlkB od
czasu ........................................................................................................................... 50
Rys 21. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HPC)TT z białkiem AlkB w pH 6,8 ...................................................................... 51
Rys 22. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od pH
buforu w mieszaninie reakcyjnej ................................................................................ 51
Rys 23. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HPC)TT z białkiem AlkB przy stężeniu Fe (II) równym 0,1mM ........................... 52
Rys 24. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od
stężenia Fe (II) w mieszaninie reakcyjnej .................................................................... 52
Rys 25. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HPC)TT z białkiem AlkB przy stężeniu α-KG równym 0,1mM ............................ 53

80
Rys 26. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od
stężenia α-KG w mieszaninie reakcyjnej .....................................................................53
Rys 27. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HPC)TT z białkiem AlkB po 20min .....................................................................54
Rys 28. Zależność wydajności naprawy substratu TT(HPC)TT przez białko AlkB od
czasu ...........................................................................................................................54
Rys 29. Chromatogram przedstawiający wzorzec substratu (poddany następnie
inkubacji z białkiem SA-1A), otrzymany po reakcji TTCTT z CAA ............................55
Rys 30. Chromatogram przedstawiający wzorzec substratu (poddany następnie
inkubacji z białkiem SA-1A), otrzymany po reakcji TTCTT z CAA ............................56
Rys 31. Chromatogram przedstawiający wzorzec substratu (poddany następnie
inkubacji z białkiem SA-1A), otrzymany po reakcji TTCTT z akroleiną. .....................56
Rys 32. Chromatogram przedstawiający wzorzec substratu (poddany następnie
inkubacji z białkiem SA-1A), otrzymany po reakcji TTATT z CAA ............................57
Rys 33. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HEC)TT z białkiem SA-1A w pH 5,8 ...................................................................57
Rys 34. Zależność wydajności naprawy uzyskanych substratów przez białko SA-1A od
pH buforu w mieszaninie reakcyjnej ...........................................................................58
Rys 35. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HPC)TT z białkiem SA-1A przy stężeniu α-ketoglutaranu równym 0,05mM.. ......58
Rys 36. Zależność wydajności naprawy uzyskanych substratów przez białko SA-1A od
stężenia α-KG w mieszaninie reakcyjnej .....................................................................59
Rys 37. Chromatogram przedstawiający skład mieszaniny reakcyjnej po inkubacji
TT(HEC)TT z białkiem SA-1A przy stężeniu Fe (II) równym 2mM ...........................59
Rys 38. Zależność wydajności naprawy substratu uzyskanych substratów przez białko
SA-1A od stężenia Fe w mieszaninie reakcyjnej ..........................................................60
Rys 38. Asparaginian 135 w centrum aktywnym białka AlkB ......................................64

81
Spis tabel
Tabela 1. Rozmieszczenie bakteryjnych białek AlkB w kompletnie
zsekwencjonowanych genomach ................................................................................. 26
Tabela 2. Zależność optymalnych warunków naprawy substratów AlkB od ich pKa. ... 63
Tabela 3. Optymalne warunki naprawy substratów przez białko SA-1A ...................... 65

82

83
Wyrażam zgodę na udostępnienie mojej pracy w czytelniach Biblioteki SGGW
...................................................... (czytelny podpis autora)


















