
Syntazy tlenku azotu (NOS) -...
56
Biochemia stresu oksydacyjnego Literatura: Grzegorz Bartosz „Druga twarz tlenu” Barry Halliwell & John Gutteridge „Free radicals in biology and medicine” Syntazy tlenku azotu (NOS) Wykład 3
Transcript of Syntazy tlenku azotu (NOS) -...

Biochemia stresu oksydacyjnego
Literatura:Grzegorz Bartosz „Druga twarz tlenu”
Barry Halliwell & John Gutteridge „Free radicals in biology and medicine”
Syntazy tlenku azotu (NOS)
Wykład 3

Tlenek azotu- Tlenek azotu jest wolnym rodnikiem (ma niesparowany elektron).
- Jest dobrze rozpuszczalny w rozpuszczalnikach organicznych – łatwo dyfunduje przez błonę komórkową.
- Odgrywa kluczową rolę w prawidłowym funkcjonowaniu układu krążenia, układu nerwowego i układuodpornościowego:
* jest wazodylatorem (nadciśnienie – nitrogliceryna, azotany organiczne)* jest neurotransmiterem* jest czynnikiem toksycznym dla patogenów* odgrywa zasadniczą rolę w przebiegu szoku septycznego: nadprodukcja NO przez
iNOS prowadzi do spadku ciśnienia krwi
- Produkowany jest:* przez syntazy tlenku azotu (NOS) z argininy* z azotanów i azotynów
- Tlenek azotu reaguje z białkami, zwłaszcza zawierającymi:* centra żelazowo-siarkowe* jony metali przejściowych* grupy hemowe
- W mitochondriach tlenek azotu wiąże się z oksydazą cytochromu = kompetycja z tlenem
- Tlenek azotu może wykazywać właściwości antyoksydacyjne: jest wolnym rodnikiem, więc łatworeaguje z innymi wolnymi rodnikami (np. biorącymi udział w peroksydacji lipidów) i terminuje reakcję.

Wazodylatacja zależna od NO
- activation of a calcium-sensitive potassium channel through the cGMP-dependent protein kinase, which results in membrane hyperpolarization?- More likely, rise in cGMP probably initiates Ca extrusion from the smooth muscle cell which causes relaxation
NO i erekcja:- mediator komórek nerwowych
autonomicznego układu nerwowego
- wazodylatacja naczyń prącia
Viagra (Sildenafil) = PDE5 inhibitor
(NO pełni również funkcję w utrzymaniu cyklu estrogenowego i w implantacji zapłodnionego jaja)
NO kowalencyjnie wiąże grupę hemową sGC

Tlenek azotu
- Tlenek azotu jest nietrwały w obecności tlenu. W natlenionych roztworachwodnych jego okres półtrwania wynosi kilka sekund, a tlenek azotu reaguje z tlenemdając dwutlenek azotu:
2NO˙ + O2 2NO2˙
- Dwutlenek azotu, jest bardziej reaktywny niż tlenek azotu, reaguje ze związkaminienasyconymi tworząc wolne rodniki, w których niesparowany elektron jestzlokalizowany na atomie węgla.
- W roztworach wodnych o pH obojętnym dwutlenek azotu ulegadysproporcjonowaniu, tworząc anion azotynowy i azotanowy:
NO˙ + H2O NO2- + NO3- + 2H+
- Tlenek azotu bardzo szybko reaguje z anionorodnikiem ponadtlenkowym tworzącnadtlenoazotyn:
NO˙ + O2-˙ ONOO-

Aktywność syntaz tlenku azotu (NOS)
Alderton et al. Biochem J. 2001
- Elektrony są przekazywane z NADPH dodomeny reduktazowej NOS, a następnieprzez FAD i FMN przenoszone są dodomeny oksygenazowej NOS. Tu reagują
z żelazem hemowym i BH4 w centrumaktywnym, katalizując reakcję tlenu z L-argininą, prowadzącą do powstaniacytruliny i NO.
- Transport elektronów przez domenęreduktazową wymaga obecnościCa2+/CaM.- Dimeryzacja NOS jest kluczowa dla jej
aktywności
BH4 - tetrahydrobiopterynaCaM - kalmodulina
Syntaza tlenku azotu (NOS) jest enzymem zawierającym FAD (dinukleotydflawinoadeninowy) i FMN (mononukleotyd flawinowy). Jej substratem jest arginina, któraulega redukcji przy udziale NADPH, a z której po odłączeniu NO powstaje cytrulina.Kofaktorem tej reakcji jest tetrahydrobiopteryna (BH4). Analogi argininyblokują działaniesyntazy.

Reakcje katalizowane przez NOS
Alderton et al. Biochem J. 2001
- Anion nitroksylowy jest utleniany doNO w reakcji katalizowanej przezSOD.- SOD może nasilać biodostępność NOpoprzez dysmutację anionorodonikaponadtlenkowego, który wprzeciwnym razie reagowałby z NOtworząc nadtlenoazotyn.
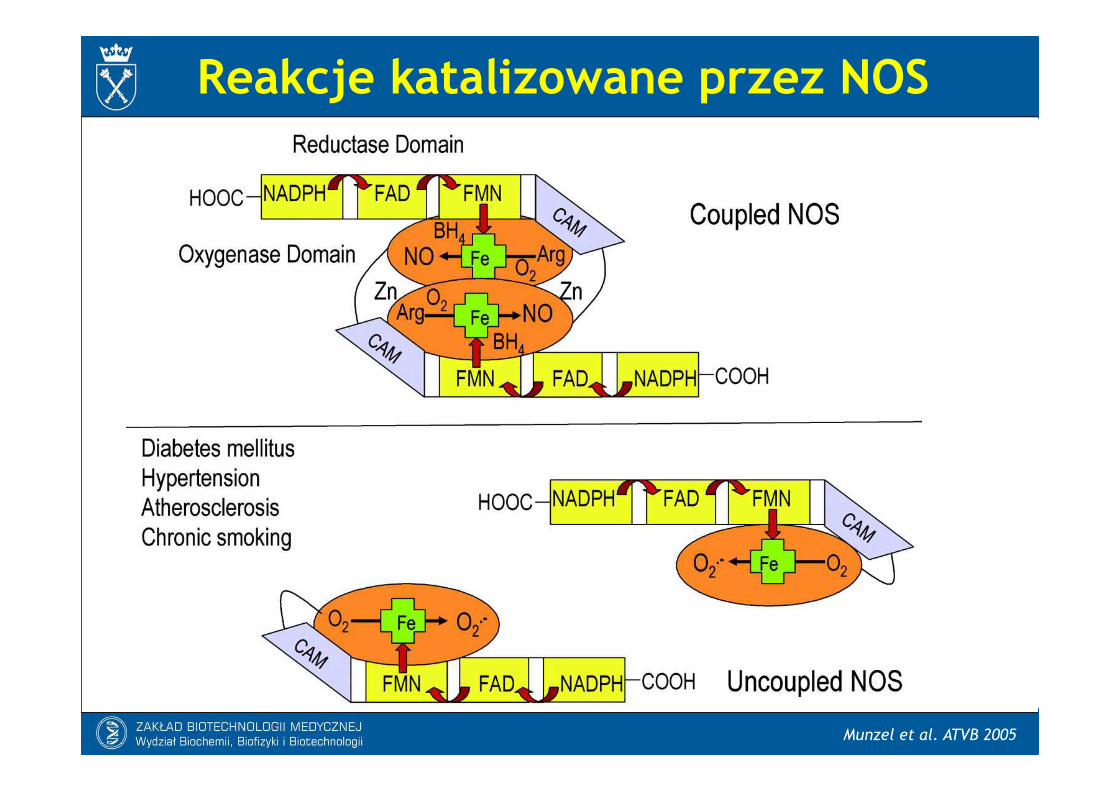
Munzel et al. ATVB 2005
Reakcje katalizowane przez NOS

Liu & Velazquez. Antioxid Redox Signal 2008
Regulacja aktywności eNOS
XO
XO: xathine oxidase
XO
XO: xathine oxidase- NO może wiązać się do eNOS-Fe2+ przed opuszczeniem miejsca katalitycznego,tworząc nieaktywny kompleks.
- Ponowna aktywacja eNOS wymaga uwolnienia NO lub reakcji z O2, tak byodtworzyć formę eNOS-Fe3+.
- NOS jest białkiem hemowym zawierającym Fe3+. Obecność NADPH pozwala naredukcję reszty flawinowej w eNOS, transfer elektronu na hem i redukcję hemu doformy eNOS-Fe2+.
- eNOS-Fe2+ katalizuje utlenianie argininy do NO i cytruliny lub (jeśli nie ma argininy)produkcję anionorodnika ponadtlenkowego.

Inhibitory NOS
Alderton et al. Biochem J. 2001;
- Mechanizmy inhibicji:* blokowanie miejsca wiązania L-argininy (np. tiocytrulina)* blokowanie miejsca wiązania BH4 (np.4-amino-BH4)* jednoczesne blokowanie miejsc wiązania L-argininy i BH4 (np. 7-NI)* blokowanie miejsca wiązania hemu (np. imidazol)* blokowanie miejsca wiązania CaM (np. trifluoperazine)* blokowanie miejsca wiązania flawin* blokowanie dimeryzacji NOS (np. diphenyleneiodonium)

Inhibitory NOS
Alderton et al. Biochem J. 2001
L-NAME

Formy NOS
Alderton et al. Biochem J. 2001
- Znane są 3 formy NOS, każda kodowana jest przez inny gen. Każdy enzym jestodmiennie regulowany, ma inną lokalizację i może mieć odmienną wrażliwość nainhibitory.
- Struktura genów wszystkich form jest podobna, co sugeruje ich wspólnepochodzenie. Homologia strukturalna form NOS na poziomie DNA u człowieka to ok.55%.
- Enzymy zbudowane są z dwóch domen połączonych miejscem wiążącymkalmodulinę:
* N-końcowej domeny oksygenazowej (wiążącej hem, BH4 i L-argininę)* C-końcowej domeny reduktazowej (wiążącej FAD, FMN i NADPH)

Formy NOS
Alderton et al. Biochem J. 2001
- Nazwy i synonimy izoform NOS:* nNOS (neuronalna NOS, NOS typu I, NOS-I, NOS-1)* iNOS (indukowalna NOS, NOS typu II, NOS-II, NOS-2)* eNOS (śródbłonkowa NOS, NOS typu III, NOS-III, NOS-3)
- Dawna (nieprecyzyjna) klasyfikacja to:* NOS indukowalna (iNOS) i konstytutywna (nNOS i eNOS)* NOS zależna od wapnia (nNOS i eNOS) i niezależna od wapnia (iNOS)
- Mimo odmienności, formy NOS wykazują znaczne podobieństwo strukturalne,podobną orientację i lokalizację kofaktorów i podobne właściwości stereochemicznecentrum aktywnego.
- Miejsce wiązania BH4 jest bardzo podobne we wszystkich izoformach.

Inhibitory NOS
-
Alderton et al. Biochem J. 2001
- Aby inhibitor mógł być wykorzystany w praktyce jako selektywny dla danej formyNOS in vivo, jego powinowactwo dla danej formy musi być 50-100 razy większe niżdla pozostałych.
- Pozorna selektywność in vivo może wynikać z tkankowo-specyficznego wchłanianiainhibitora (np. 7-nitroindazol hamuje głównie nNOS), lub różnej dostępności wtkankach L-argininy i BH4.
- Inhibitory NOS mogą również wpływać na aktywność innych enzymów.

Reakcje katalizowane przez NOS
- Inhibitory NOS mogą mieć bardzo odmienne efekty w zależności od tegoczy hamują syntezę zarówno NO jak i innych pochodnych (np. L-NNA i nNOS)czy nie (np. L-NMMA i nNOS).
- NOS mogą katalizować rozsprzężoną oksydację (bez tworzenia NO) tworzącO2'
-. W efekcie powstaje nadtlenoazotyn. Najwięcej O2'- tworzy nNOS,
zwłaszcza przy niskich stężeniach argininy lub BH4. Izoforma eNOSsyntetyzuje znacznie mniej O2'
-, choć ilości te wciąż są mierzalne. Najniższaprodukcja O2'
- charakteryzuje iNOS.
- Nadtlenoazotyn może być produkowany również jeśli pierwszymproduktem NOS jest nitroksyl zamiast NO, choć reakcja jest wolniejsza.
Alderton et al. Biochem J. 2001

Andrew & Mayer. Cardiovasc Res 1999; Alderton et al. Biochem J. 2001
Porównanie form NOS
- Struktura domen NOS została określona na podstawie badań krystalograficznych wlatach 1998-1999.
- Aktywne NOS są zwykle określane jako dimery, ale działają w rzeczywistości jakotetramery (2 cząsteczki NOS i 2 cząsteczki kalmoduliny). Zawierają też kofaktory:
* BH4 (tetrahydrobiopteryna)* FAD (dinukleotyd flawinoadeninowy)* FMN (mononukleotyd flawinowy)* protoporfiryna-IX żelaza (hem)

Porównanie form NOS
- Dimeryzacja iNOS zależy wyłącznieod domeny oksygenazowej, natomiastnNOS i eNOS od domeny oksygena-zowej i reduktazowej. DimeryzacjaiNOS jest bardziej niż dimeryzacjanNOS i eNOS zależna od kalmoduliny.
- Dimeryzacja iNOS jest w komórkachw warunkach fizjologicznychpraktycznie nieodwracalna. Nie jestpewne czy to samo odnosi się do nNOSi eNOS.
- Dla indukcji dimeryzacji kluczowajest obecność hemu, natomiaststabilizacja dimerów zależy także odBH4 i L-argininy.
Andrew & Mayer. Cardiovasc Res 1999; Alderton et al. Biochem J. 2001

Struktura białek NOS
Alderton et al. Biochem J. 2001

Tkankowo-specyficzna ekspresja NOS
Liu et al. Cardiovasc Res 2007
- Ekspresja nNOS (z reguły konstytutywna):* neurony* mięśnie szkieletowe
- Ekspresja eNOS (z reguły konstytutywna):* śródbłonek
- Ekspresja iNOS (z reguły indukowalna):* komórki linii monocytarno-makrofagowej
wątroba, eNOS (zielony)
podwzgórze, nNOS (zielony)
mikroglej, iNOS (czerwony)

Porównanie form NOS
Liu et al. Cardiovasc Res 2007

Nietypowe formy NOSmitochondrialny NOS (mtNOS)
- Barwienia immunohistochemiczne wskazują na istnienie formy mitochondrialnejNOS, która jest aktywna metabolicznie.
- Prawdopodobnie jest to forma iNOS, ale związana z błoną, a nie cytozolowa.Wiązanie z błoną może zależeć np. od kaweoliny.
reumatoidalny NOS (OA-NOS)
- Barwienia immunohistochemiczne ipomiar aktywności enzymatycznejsugerowały istnienie nowej formynNOS, ulegającej ekspresji wchondrocytach stymulowanych cytoki-nami prozapalnymi (w reumatoidal-nym zapaleniu stawów). Wydaje sięjednak, że w komórkach tych akty-wacji ulega iNOS.
jądra, oksydaza cytochromowa, mtNOS

Izoformy splicingowe nNOS
Alderton et al. Biochem J. 2001
- Formy splicingowe nNOS:* nNOS-α (aminokwasy 1-1433)* nNOS-β (aminokwasy 236-1433)* nNOS-γ (Tn-nNOS, aminokwasy 336-1433)* nNOS-µ (aminokwasy 1-1433 + 34 aminokwasowy insert w domenie wiążącej FMN)* nNOS-2 (aminokwasy 1-1433, z delecją aminokwasów 504-608)

nNOS-2
- Delecja fragmentu genu 504-608 bp prowadzi do powstania białka nNOS-2pozbawionego 105 aminokwasów, z zachowaniem ramki odczytu. Brakujący regionjest istotny dla wiązania L-argininy, prawdopodobnie więc nNOS-2 jest katalitycznienieaktywna i może działać jako inhibitor nNOS (dominant negative).
Izoformy splicingowe nNOS
nNOS-ββββ- Zachowuje ok. 80% aktywności formy nNOS-α
nNOS-γγγγ- Zachowuje jedynie ok. 3% aktywności formy nNOS-α.
- U ludzi podobna forma TnNOS ulega ekspresji główniew jądrach (komórki Leydiga) i ma dodatkowy fragment5'-UTR regulujący ekspresję w sposób tkankowospecyficzny. Jej aktywność enzymatyczna w jądrachjest podobna do nNOS-α.
Liu et al. Cardiovasc Res 2007; R. Nelson, ohio State University

Izoformy splicingowe nNOS
nNOS-µµµµ- Ulega specyficznej ekspresji w sercu, mięśniach szkieletowych, penisie i cewcemoczowej.
- Ma dodatkowy fragment 34 aminokwasów pomiędzy domenami wiążącymi CaM iflawiny.
- Kinetyka katalizowanej reakcji jestbardzo podobna jak w formie natywnej,choć być może produkcja anionorodnikaponadtlenkowego jest nieco niższa.
- Jest wrażliwa na działanie kalpainy(główna proteaza degradująca nNOS,działająca w sposób zależny od wapnia),ale rozkładana jest znacznie wolniej niżforma natywna.
nNOSµµµµ
nNOSαααα
WT
Liu et al. Cardiovasc Res 2007; Precival et al. J Clin Invest 2010

nNOS KO
- Szczepy myszy KO stworzono unieczynniając egzon 2 (pozostaje wówczas około 5%aktywności enzymatyczej cytozolowych form splicingowych) lub egzon 6 (pozostajeok. 0.3% aktywności).
* Znaczne powiększenie żołądka* Zaburzenia relaksacji mięśni gładkich* Zwiększona oporność mózgu na niedotlenienie (mniejsze uszkodzenia w wyniku
reperfuzji)* Myszy są płodne i żywotne, ale samce są bardzo agresywne
Liu et al. Cardiovasc Res 2007; R. Nelson, ohio State University

- Istnieją trzy linie myszy iNOS KO, pozbawione całkowicie ekspresji iNOS i wszystkie(mimo różnych mechanizmów inaktywacji genu) wykazują podobny fenotyp:
* brak zaburzeń wzrostu, rozwoju czy płodności* zwiększona wrażliwość na zakażenia wewnątrzkomórkowe bakteriami (np.
Listeria monocytogenes) lub pierwotniakami (np. Leishmania major).* Zwiększona oporność na skutki sepsy - znacznie mniejsza odpowiedź
hipotensyjna.
iNOS KO
Hierholzer et al. J Exp Med. 1998; Liu et al. Cardiovasc Res 2007
Hepatic injury in WT and iNOS knockout mice subjected to hemorrhagic shock.

- Istnieje kilka szczepów myszy eNOS KO,pozbawionych całkowicie ekspresji eNOS iwszystkie (mimo inaktywacji różnych fragmentówgenu) mają podobny fenotyp:
* brak anomalii rozwojowych i anatomicznych
* normalna żywotność i płodność
* nadciśnienie (wzrost o ok. 30%)
* zaburzona relaksacja naczyń
* nasilone tworzenie blaszek miażdżycowych pozranieniu naczynia
* nasilona miażdżyca indukowana dietą
Liu et al. Cardiovasc Res 2007
eNOS KO

Porównanie form NOS
Liu et al. Cardiovasc Res 2007
Fenotyp myszy pozbawionych genów NOS (nNOS KO, eNOS KO i iNOS KO)

Niedobór NOS
Liu et al. Cardiovasc Res 2007

- Aktywność eNOS jest regulowana przez Hsp90, który działa jak modulatorallosteryczny (trzykrotny wzrost aktywności eNOS). Interakcja ta nasilana jest przez:
* VEGF* histaminę* siły ścinania
- Kompleks z Hsp90 tworzy również nNOS. Tu Hsp90 ułatwia wbudowywanie hemu donNOS, a jego funkcja jest szczególnie istotna przy niskich stężeniach hemu.
- iNOS jest regulowany głównie na poziomie transkrypcji, ale w układzie nerwowymmoże oddziaływać z kaliriną. Białko to hamuje tworzenie dimerów iNOS i możedziałać neuroprotekcyjnie w trakcie zapalenia.
Aktywność NOS
Alderton et al. Biochem J. 2001

Rola BH4 w aktywności NOS
Alderton et al. Biochem J. 2001

NOS jako źródło RFT
Gdy NOS nie jest w dostatecznym stopniu wysycona BH4, enzym aktywowany przezkompleks Ca-kalmodulina nie działa na substrat (argininę), lecz jego domena reduktazowa redukuje tlencząsteczkowy, poprzez przyłączenie jednego elektronu (powstaje anionorodnik ponadtlenkowy), czylidziała jak oksydaza NADPH.
Małe stężenie substratu również sprzyja tej reakcji.
Gdy stężenie BH4 jest w przedzialepośrednim, jedna jednostka NOS działanormalnie, produkując NO, a druga uwalniaanionorodnik ponadtlenkowy. NO+ O2
-˙ →ONOO- (nadtlenoazotyn)Podobna sytuacja, gdy jest mało SOD.
Wytwarzanie nadtlenozaotynu przez eNOS –rozwój miażdżycy

Rola CaM w aktywności NOS
Alderton et al. Biochem J. 2001
- CaM jest niezbędna do aktywności enzymatycznej wszystkich izoform NOS,choć eNOS i nNOS są ściślej regulowane niż iNOS. CaM nasila transferelektronów z NADPH do domen flawinowych.
- nNOS i eNOS różnią się od iNOS, obecnością 40-50 aminokwasów w obrębiedomeny wiążącej FMN, stanowiących tzw. pętlę autoinhibicyjną. Działa onapoprzez destabilizację wiązania CaM przy niskim poziomie Ca2+ i poprzezhamowanie przekazywania elektronów z FMN do hemu przy braku Ca2+/CaM.
kalmodulina

- Aktywność nNOS i eNOS jest regulowana przez fosforylację.* fosforylacja Ser1177 eNOS (przez Akt kinazę) zwiększa
produkcję NO* fosforylacja Ser486 nNOS (przez kinazę zależną od CaM)
zmniejsza produkcję NO.
- Siły ścinania nasilają fosforylację eNOS, zwiększającaktywność enzymatyczną, niezależnie od jonów wapnia.
- N-końcowy fragment nNOS zawiera miejsce wiązania białkaPIN (protein inhibitor of NOS).
- Rola PIN nie jest jasna:?jedynie nNOS może reagować z PIN, który hamuje
aktywność enzymatyczną poprzez destabilizację dimerównNOS
?PIN nie ma wpływu na dimeryzację nNOS, natomiasthamuje aktywność domeny oksygenazowej nNOS, eNOS i iNOSoraz oksygenazy NADPH
?PIN jest inhibitorem lekkiego łańcucha miozyny i dyneiny -może regulować transport aksonalny nNOS, ale nie wpływa naaktywność enzymatyczną nNOS
Regulacja aktywności NOS
Alderton et al. Biochem J. 2001; Magee et al. Biol Reprod 2003.
unerwienie penisa

- eNOS jest mirystylowany i palmitynowany:* przyłączenie mirystynianu do N-końcowej glicyny jest kotranslacyjne i
nieodwracalne* przyłączenie palmitynianu do cysteiny jest po-translacyjne i odwracalnie,
regulowane przez zmiany wewnątrzkomórkowego stężenia Ca2+ (zależnego np. odbradykininy).
- Jednoczesna mirystylacja i pamitynacja jest niezbędna do lokalizacji eNOS wkaweolach plazmalemmy komórek śródbłonka.
Regulacja lokalizacji NOS
Alderton et al. Biochem J. 2001

- Białko NOS jest zlokalizowane w kaweolach. W śródbłonku łączy się z kaweoliną-1,a w kardiomiocytach z kaweoliną-3.
- Kaweolina-1 hamuje aktywność eNOS w sposób zależny od Ca2+/CaM.
- Kaweolina-3 w kardiomiocytach hamuje także aktywność nNOS w sposób zależny odCa2+/CaM.
Regulacja aktywności NOS
Alderton et al. Biochem J. 2001

Liu et al. Cardiovasc Res 2007
Regulacja aktywności eNOS
poziom ekspresjimRNA i białka
dostępność substratu
dostępnośćkofaktorów
lokalizacja komórkowa
fosforylacja
reakcja NO z O2'-

- Wszystkie formy NOS wpływają na regulację ciśnienia krwi:* eNOS - bezpośrednia regulacja wazorelaksacji i funkcji mięśni gładkich
(obniżanie cisnienia krwi)* nNOS - brak wpływu na ciśnienie krwi w warunkach kontrolnych, ale być może
kontrola baroreceptorów (nNOS jest produkowana w centrach wazomotorycznych inerwach periwaskularnych). Być może nNOS może podnosić ciśnienie krwi, chroniączmniejszając ryzyko hipotencji np. podczas anestezji)
* iNOS - wywołuje hipotensję podczas szoku septycznego
- Jedną z głównych przyczyn dysfunkcji śródbłonka (niewłaściwa regulacjawazorelaksacji) jest niedobór NO produkowanego przez eNOS. Jest to elementpatogenezy:
* miażdżycy* nadciśnienia* cukrzycy* hipercholesterolemii* przedwczesnego starzenia
- Dysfunkcja pojawia się przed zmianami anatomicznymi (przed hiperplazjąmięśniówki lub tworzeniem złogów lipidowych w naczyniach).
NOS - regulacja ciśnienia krwi
Liu et al. Cardiovasc Res 2007
blaszka miażdżycowa
Wpływ starzenia na dysfunkcję naczyń
Herrera et al. Aging Res Rev 2010
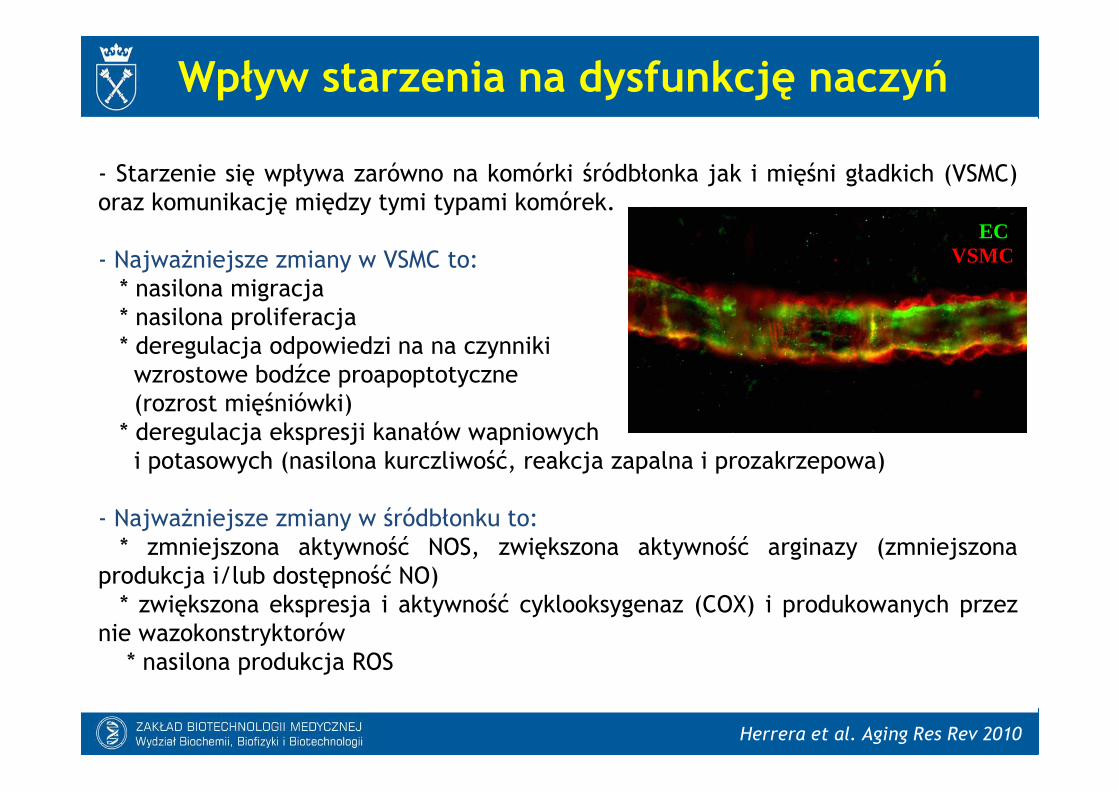
- Starzenie się wpływa zarówno na komórki śródbłonka jak i mięśni gładkich (VSMC)oraz komunikację między tymi typami komórek.
- Najważniejsze zmiany w VSMC to:* nasilona migracja* nasilona proliferacja* deregulacja odpowiedzi na na czynnikiwzrostowe bodźce proapoptotyczne(rozrost mięśniówki)
* deregulacja ekspresji kanałów wapniowychi potasowych (nasilona kurczliwość, reakcja zapalna i prozakrzepowa)
- Najważniejsze zmiany w śródbłonku to:* zmniejszona aktywność NOS, zwiększona aktywność arginazy (zmniejszona
produkcja i/lub dostępność NO)* zwiększona ekspresja i aktywność cyklooksygenaz (COX) i produkowanych przez
nie wazokonstryktorów* nasilona produkcja ROS
ECVSMC

Herrera et al. Aging Res Rev 2010
Wpływ starzenia na dysfunkcję naczyń
DAMA: NG-NG0-dimethyl-arginine asymmetric

- Większość badań wskazuje na zwiększoną ekspresję eNOS u starzejących sięzwierząt. Niektóre badania wskazują na obniżenie ekspresji eNOS w aortachstarzejących się szczurów.
Herrera et al. Aging Res Rev 2010
- Suplementacja arginina może poprawiać funkcje śródbłonka u starzejących sięludzi.
- Poziom BH4 może obniżać się z wiekiem, zwiększając prawdopodobieństwoprodukcji anionorodnika ponadtlenkowego zamiast NO przez eNOS.
Wpływ starzenia na dysfunkcję naczyń
eNOS

Herrera et al. Aging Res Rev 2010
Wpływ starzenia na dysfunkcję naczyń
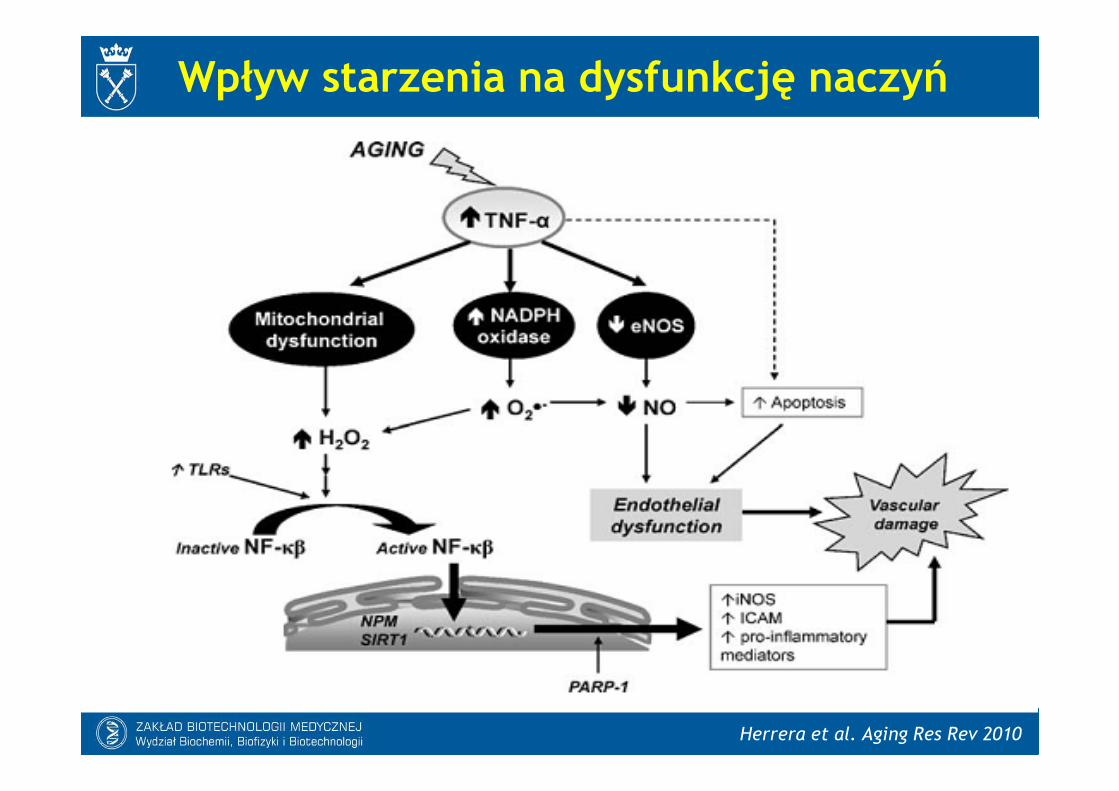
Herrera et al. Aging Res Rev 2010
Wpływ starzenia na dysfunkcję naczyń

Herrera et al. Aging Res Rev 2010
Wpływ starzenia na dysfunkcję naczyń
TERT: telomerase reverse transcriptase.

Gojenie ran u myszy z cukrzycą
Days after wounding
0
20
40
60
80
100
0 1 2 3 4 5 6 7 8 10 11 13 15
Surf
ace o
f th
e w
ound [
%]
WT
db/db**
*****
***
*** *** *** ****** ***
***
Grochot-Przeczek et al. PLoS ONE, 2009
db/db WT
Day 1
Day 3
Day 8
Day 17
Day 0
Liu & Velazquez. Antioxid Redox Signal 2008
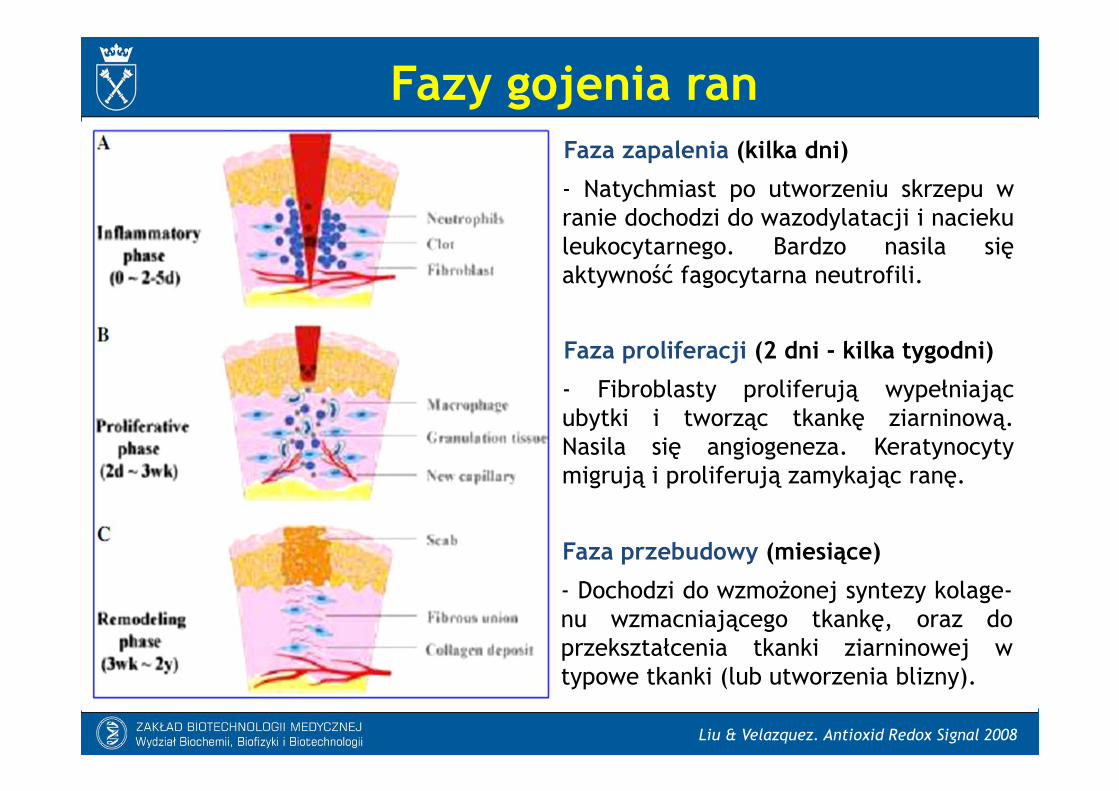
Fazy gojenia ran
Faza zapalenia (kilka dni)
- Natychmiast po utworzeniu skrzepu wranie dochodzi do wazodylatacji i naciekuleukocytarnego. Bardzo nasila sięaktywność fagocytarna neutrofili.
Faza proliferacji (2 dni - kilka tygodni)
- Fibroblasty proliferują wypełniającubytki i tworząc tkankę ziarninową.Nasila się angiogeneza. Keratynocytymigrują i proliferują zamykając ranę.
Faza przebudowy (miesiące)
- Dochodzi do wzmożonej syntezy kolage-nu wzmacniającego tkankę, oraz doprzekształcenia tkanki ziarninowej wtypowe tkanki (lub utworzenia blizny).

Jarajapu & Grant. Circ Res 2010.
Upośledzenie funkcji EPC w cukrzycy
EPC: komórki progenitorowe śródbłonka
- U zdrowych osobników czynniki uwalnianeprzez ischemiczne lub zranione tkankimobilizują komórki progenitorowe (w tymEPC) ze szpiku, a te po dotarciu do miejsczranienia lub niedotlenienia uwalniajączynniki proangenne (w tym niewielkie ilościNO i ROS) oraz biorą udział wneowaskularyzacji i naprawie naczyń.
- W cukrzycy sygnały wysyłane przezniedotlenione lub zranione tkanki są słabsze,przez co mobilizacja komórek progenitoro-wych jest mniejsza. Komórki progenitorowektóre docierają do zranionych tkanekuwalniają niewiele czynników proangiogen-nych, natomiast dużo prozapalnych iantyangiogennych (w tym duże ilości NO iROS).

Brem & Tomic-Canic. J Clin Invest 2007.
Upośledzenie gojenia ran w cukrzycy- EPC: komórki progenitorowe śródbłonka
- VEGF: vascular endothelial growth factor- SDF-1: stromal cell derived growth factor
- eNOS: endothelial nitric oxide synthase

Liu & Velazquez. Antioxid Redox Signal 2008
Mobilizacja EPC
- Zranione tkanki uwalniają między innymi VEGF, który wpływa na komórkipodścieliska szpiku. Prowadzi to do:
* aktywacji enzymatycznej eNOS* nasilenia produkcji NO.
- To zwiększa aktywność MMP-9, która uwalnia sKitL z błonowego białka mKitL.
- Związanie sKitL do receptora c-Kit prowadzi do uwolnienia EPC z niszy szpikowychdo krążenia.
EPC w szpiku

Mechanisms of blood vessels formation
Formation of blood vessels de novo
Incorporation to pre-existing vessels
paracrine stimulationof endothelial cells (VEGF, bFGF, IL-8…)
Carmeliet P. Nat Med, 2000

Liu & Velazquez. Antioxid Redox Signal 2008
Regulacja mobilizacji EPC w hiperoksji i hipoksji
- W cukrzycy zmniejszony jest poziom fosforylacji eNOS przez kinazy PI3K/Akt,przez co zmniejsza się produkcja NO.
- Nasilony stres oksydacyjny i zwiększona produkcja anionorodnikaponadtlenkowego prowadzi do nasilonej syntezy nadtlenoazotynu (ONOO-) izmniejszenia dostępności NO.
- W ranach cukrzycowych osłabiona jest synteza SDF-1, co zmniejsza napływkomórek progenitorowych do uszkodzonych tkanek. To przyczynia się doupośledzenia angiogenezy i opóźnia gojenie.
- Cukrzyca powoduje również zmniejszenie liczby EPC w szpiku.
WT db/db

Wpływ cukrzycy na EPC
Kotlinowski et al., in preparation
Percentage of EPC in bone marrow
0.0000
0.0002
0.0004
0.0006
0.0008
0.0010
0.0012
healthy diabetic
% C
D45-/
KD
R+/S
ca-1
+/l
ecti
n+
*
0
200
400
600
800
0
10
20
30
40
50
0
2
4
6
8
10
num
ber
of
cells
*
Migration to SDF-1
healthy diabetic
num
ber
of
connecti
ons
*
healthy diabetic
Morphogenesis
cum
ula
tive k
ength
of
spro
uts
*
healthy diabetic
Capillary sprouting
(one of) Primary mechanism: oxidative stress leading to increased ROS production,
reduced NO availability, diturbed PI3K/Akt signaling and augmented inflammation

Kotlinowski et al., in preparation
Wpływ cukrzycy na komórki śródbłonka
Krążki aorty zatopione w matriżelu - 5 dni inkubacji. Widoczne tworzące się kapilary.
Myszy WT
Myszy db/db

Odtwarzanie krążenia w mięśniu myszy z cukrzycą
Ebrahimian et al. Am J Pathol 2006
diabetes diabetes + NAC
diabetes + NACdiabetes
NAC - N-acetylocyteina

Liu & Velazquez. Antioxid Redox Signal 2008
Mobilizacja EPC ze szpiku
- EPC: komórki progenitorowe śródbłonka- VEGF: vascular endothelial growth factor- MMP-9: matrix metalloproteinase-9- eNOS: endothelial nitric oxide synthase

Liu & Velazquez. Antioxid Redox Signal 2008
Regulacja mobilizacji EPC w hiperoksji i hipoksji

Slajdy dost ępne na stronie Zakładu Biotechnologii Medycznej
Zapraszam na wykład 4


















