
OPTYMALNE KSZTAŁTOWANIE PRZEKROJU PODŁUŻNEGO DŹWIGARÓW AŻUROWYCH KŁADEK DLA PIESZYCH

Strategie życiowe roślin – czy będziemy ciągle szukać pod latarnią?
Jan Kozłowski
Instytut Nauk o Środowisku
Uniwersytetu Jagiellońskiego
Bofizmat 2, Warszawa, 15/16 maja 2015

Jeden rodzaj, wiele stylów życia
Jednoroczne: Veronica agrestis
Wikipedia Wikipedia
Dwuletnie: Veronica polita
Wikimedia
Krzewinka: Veronica fruticans
Wikimedia
Krzew: Veronica (Hebe) speciosa

Jeden rodzaj, wiele stylów życia
Jednoroczne: Veronica agrestis
Wikipedia Wikipedia
Dwuletnie: Veronica polita
Wikimedia
Krzewinka: Veronica fruticans
Wikimedia
Krzew: Veronica (Hebe) speciosa
Rozumiemy nieźle te różnice, ale teoria
dotyczy roślin nie rozmnażających się
wegetatywnie!
Większość roślin rozmnaża się
wegetatywnie!

Rozmnażanie poprzez kłącza
www.shweeashbamboo.com/bamboo%20care%20and%20maintenance.htm
Bambus. Jeden klon może zajmować wiele kilometrów kwadratowych.

Modele optymalnej allokacji zasobów przez rośliny jednoroczne
( )P w
( )tu P w
(1 ) ( )tu P w
Optymalne przełączenie jest prawie
zawsze zero-jedynkowe
ut=0 or u t =1
w= masa
wegetatywna
( )
(1 ) ( )
t
t
w u P w
y u P w
wzrost
reprodukcja
Maksymalizowane jest y(T)

Modele optymalnej allokacji zasobów przez rośliny jednoroczne
( )P w
( )tu P w
(1 ) ( )tu P w
Optymalne przełączenie jest
prawie zawsze zero-jedynkowe
ut=0 or u t =1
w= body mass
( )
(1 ) ( )
t
t
w u P w
y u P w
wzrost
reprodukcja
Jest to najprostszy model.
Można dodać odkładanie
zasobów, inwestowanie w
obronę chemiczną,
mechaniczną, itd.

Metody
• Zasada Maksimum Pontryagina (PMP)– Metoda analityczna
– Konstruowany jest Hamiltonian; zwykle Hamiltonian jest liniowy względem kontroli, co implikuje rozwiązanie typu bang-bang;
– Tylko „biologicznie dziwna” nieliniowość prowadziłaby do przełączenia stopniowego;
– System musi być stosunkowo prosty
• Programowanie dynamiczne (DP)– Metoda czysto numeryczna
– System może być dowolnie skomplikowany
• Inne metody (inne)– Zwykle stosowane do specjalnych przypadków.

Ogólna reguła
• At each time resources should be allocated to the compartment that has maximum marginal value for fitness.
– In the Pontryagin Maximum Principle method it is equivalent to allocatation into compartment with the highest co-state (adjoined) variable.
– F = fitness (dostosowanie; to co maksymalizujemy; tutaj całożyciowa alokacja zasobów w reprodukcję)
– xi = zmienna stanu i
– marginal value ( ( )) ( )( )
i i
i
dFV x t V t
dx t

Zasada Maksimum Pontryagina
( ) ( )
(1 ( )) ( )
w u t P w
y u t P w
1 2
1 2
( ) ( ) ( ( )) ( )(1 ( )) ( ( )
( ) ( ) 0
H p t u t P w t p t u t P w t
p T p T
p1 i p2 są to co-state variables (adjoined variables)
Maksymalizowane jest y(T)
1 2
( ) ( )( ) ( )
dy T dy Tp t p t
dw dy
Aby zmaksymalizować y(T), zmienna kontrolna u(t)
musi maksymalizować H dla każdego t.
marginal value of y
1 2( ) ( )( ) ( )
H Hp t p t
w t y t
marginal value of x
Optymalnym jest lokowanie zasobów w
każdej chwili t w zmienną, która ma
największą marginal value.
Alokacja mieszana (rozwiązanie osobliwe =
singular solution) jeśli obie pochodne są
jednakowe dla przedziału t.

Rozmiar i marginal values
czas czas0 0T T
marg
inal valu
e
masa wegetatywna
masa generatywna
ts ts
rozm
iar

Rośliny jednorocznetempo produkcji proporcjonalne do masy
P(w)=0.2wWikipedia
Masa w
egeta
tyw
na
w
200
400
00150300
Czas do przejścia
Krzywa przełączeń
Koniec sezonu
Cohen (1971); metoda: inne
Wynik potwierdzony przy
pomocy PMP przez
Denholm 1975.

Rośliny jednoroczneprodukcja nieliniowa
P(w)=2w.67- w.75
Wikipedia
400
Masa w
egeta
tyw
na w
800
0
0100200Czas do przejścia
Koniec sezonu
Krzywa przelączeń
Vincent & Pulliam 1980; Ziółko & Kozłowski 1983; metoda: PMP

Układ dynamiczny
• Podstawowy układ równań może być bardziej skomplikowany:
– Jeśli części wegetatywne są częściowo tracone lub starzeją się, a części generatywne są tracone;
– Jeśli dojrzewające owoce mogą fotosyntetyzować.
( )
(1 ) ( )
t w
t y
w u P w w
y u P w y
( )
(1 ) ( )
t w
t
w u P w w
y u P w vy

Marginal valuesWażne jest, co się dzieje z owocami!
czas
mar
gin
al v
alu
e
1
Tts
Jeśli owoce mogą fotosyntetyzować… Jeśli owoce sa częściowo tracone …
… przełączenie wcześniej … przełączenie później
czas
1
Tts
mar
gin
al v
alu
e

Alokacja zasobów roślin jednorocznych
( )P w
( )uP w
(1 ) ( )u P w
wzrost
reprodukcja
relokacja
( )endP w
w= body mass

Marginal valuesRelokacja z części wegetatywnych do generatywnych
opóźnia przełączenie
Iwasa 2000, Kozłowski i Wiegert 1980
bez relokacji
z relokacją
mar
gin
al v
alu
e
1
ts
1
Tts
bez relokacji
z relokacją
Tczas czas
Jeśli tempo produkcji zależy liniowo od masy wegetatywnej
Jeśli tempo produkcji rośnie wolniej niż liniowo z masą
wegetatywną
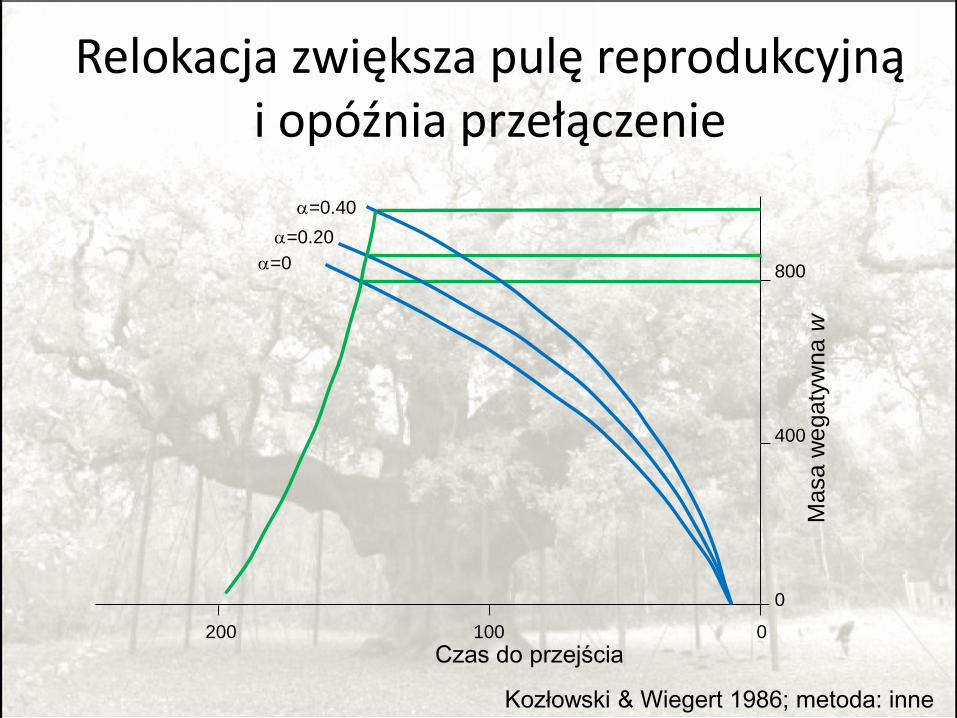
Relokacja zwiększa pulę reprodukcyjną i opóźnia przełączenie
Czas do przejścia200 100 0
800
400
0
Masa w
egaty
wna w
=0.40
=0.20
=0
Kozłowski & Wiegert 1986; metoda: inne

Czy wielokrotne przełączenia mogą być optymalne u roślin jednorocznych?
• Tak, jeśli:
– Duże straty masy wegetatywnej wcześnie w sezonie
– Głęboki spadek produkcji w środku sezonu
• Jeśli rozwijające się owoce fotosyntetyzują, wielokrotne przełączenia mniej prawdopodobne.
King and Roughgarden 1982; metoda: PMP
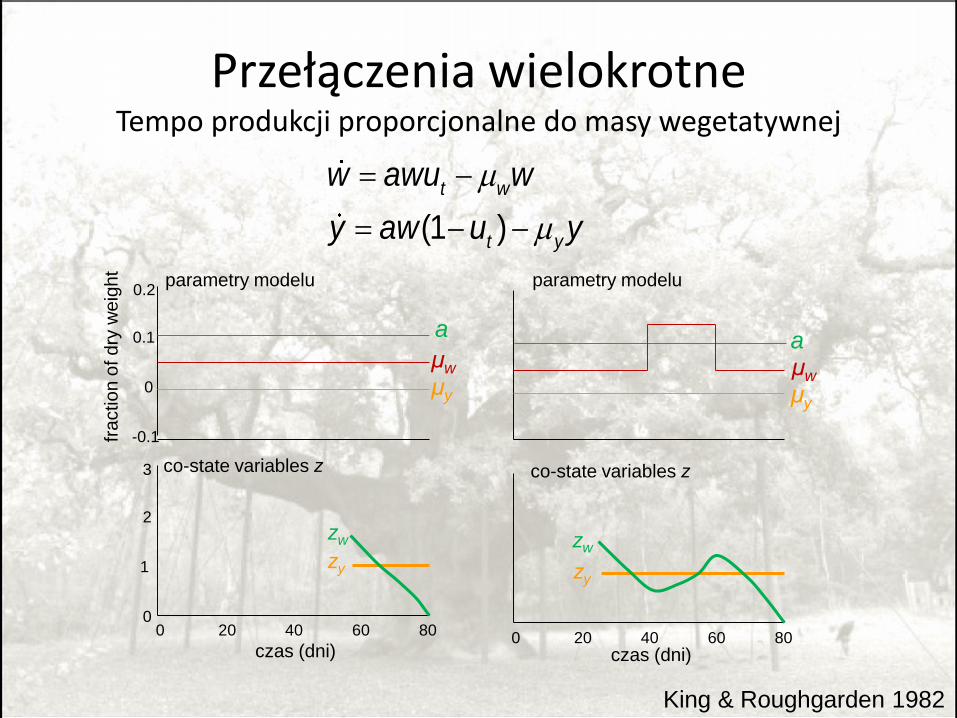
Przełączenia wielokrotneTempo produkcji proporcjonalne do masy wegetatywnej
0 20 40 60 80
czas (dni) czas (dni)0 20 40 60 80
0.2
0.1
0
-0.1
3
2
1
0
fra
ctio
n o
f d
ry w
eig
ht
parametry modelu parametry modelu
co-state variables z co-state variables z
aa
μw μwμy μy
zw zwzy zy
King & Roughgarden 1982
(1 )
t w
t y
w awu w
y aw u y
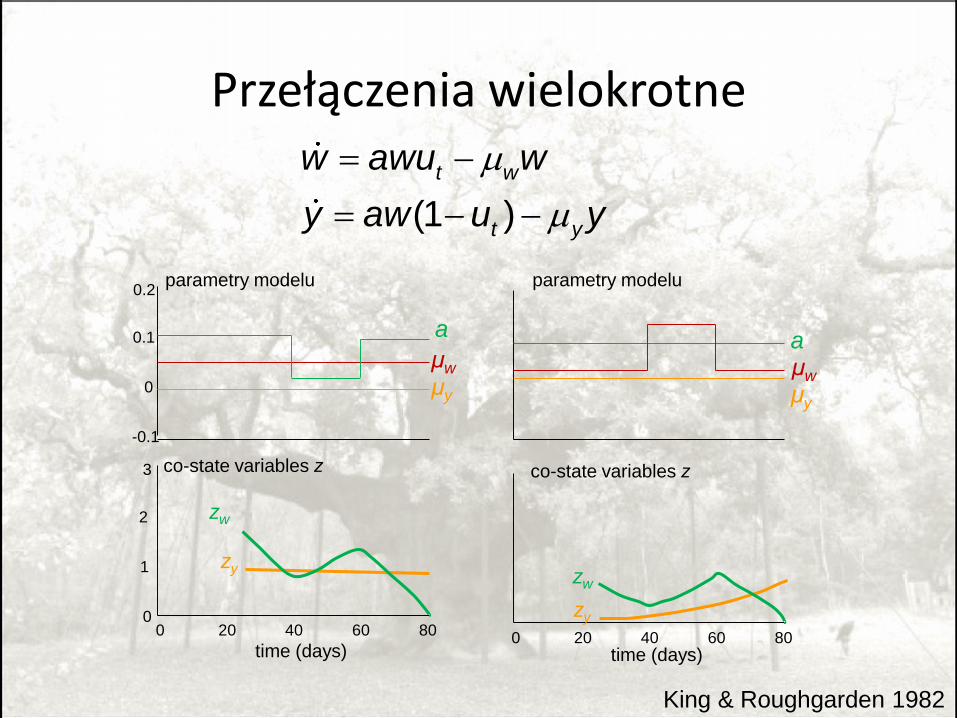
Przełączenia wielokrotne
(1 )
t w
t y
w awu w
y aw u y
0 20 40 60 80
time (days) time (days)0 20 40 60 80
0.2
0.1
0
-0.1
3
2
1
0
parametry modelu parametry modelu
co-state variables z co-state variables z
aa
μw μwμy μy
zw
zw
zy
zy
King & Roughgarden 1982

Przełączenia wielokrotne
(1 )
t w
t y
w awu w
y aw u y
0 20 40 60 80
time (days) time (days)0 20 40 60 80
0.2
0.1
0
-0.1
3
2
1
0
model parameters model parameters
co-state variables z co-state variables z
aa
μw μwμy μy
zw
zw
zy
zy
King & Roughgarden 1982

Czy odkładanie zapasów może być optymalne u roślin jednorocznych?
• Tak, jeśli:
– Organy zapasowe są mniej eksploatowane przez roślinożerców niż owoce (Chiariello & Roughgarden 1984);
– Najlepszy czas na produkcję wegetatywną i produkcję nasion jest różny (Schaffer et al. 1982).

Alokacja zasobów u wieloletnich roślin zielnych
• Przełączenie z wzrostu na odkładanie zapasów zawsze zero-jedynkowe
• ut=0 or ut=1.
• Istotna jest proporcja zasobów zużywanych na reprodukcję, nie dokładny timing.
( )P w
( )tu P w
(1 ) ( )tu P w
growth
storagew= body mass
S= ilość zapasów
odrastanie na początku
następnego sezonu
(1-zt)ST
reprodukcja w
tym samym
sezonie
ztST

Wieloletnie rośliny zielnebez reprodukcji wegetatywnej
lato
wio
sna
jesie
ń
wio
sna
zim
a
jesie
ń
zim
a
krytyczny stan zapasów
rlato
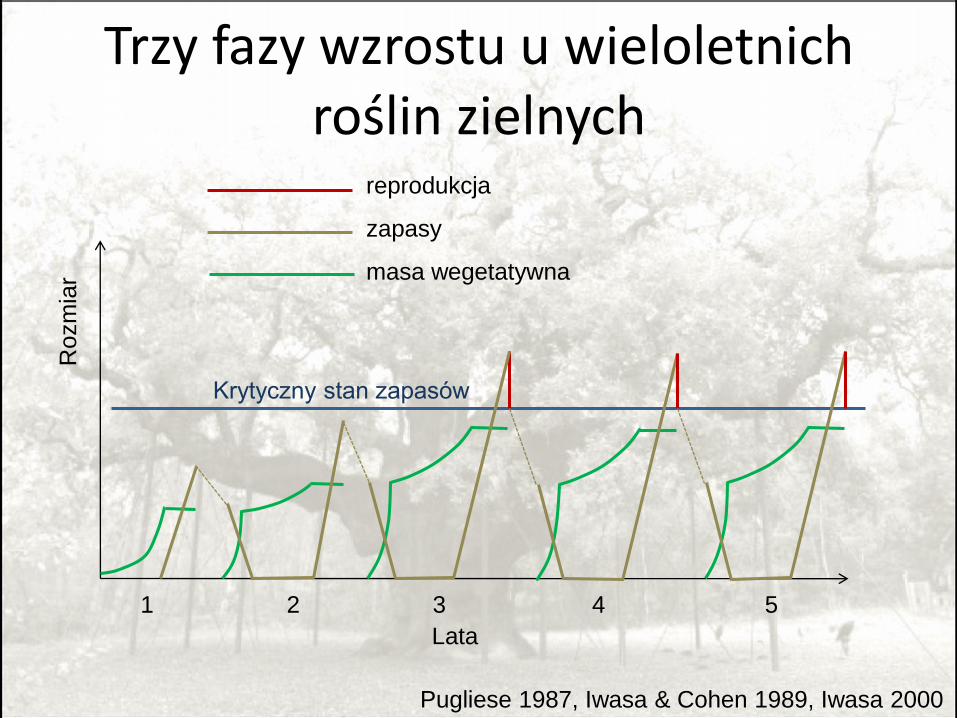
Trzy fazy wzrostu u wieloletnich roślin zielnych
Rozm
iar
Lata
1 2 3 4 5
Krytyczny stan zapasów
reprodukcja
zapasy
masa wegetatywna
Pugliese 1987, Iwasa & Cohen 1989, Iwasa 2000
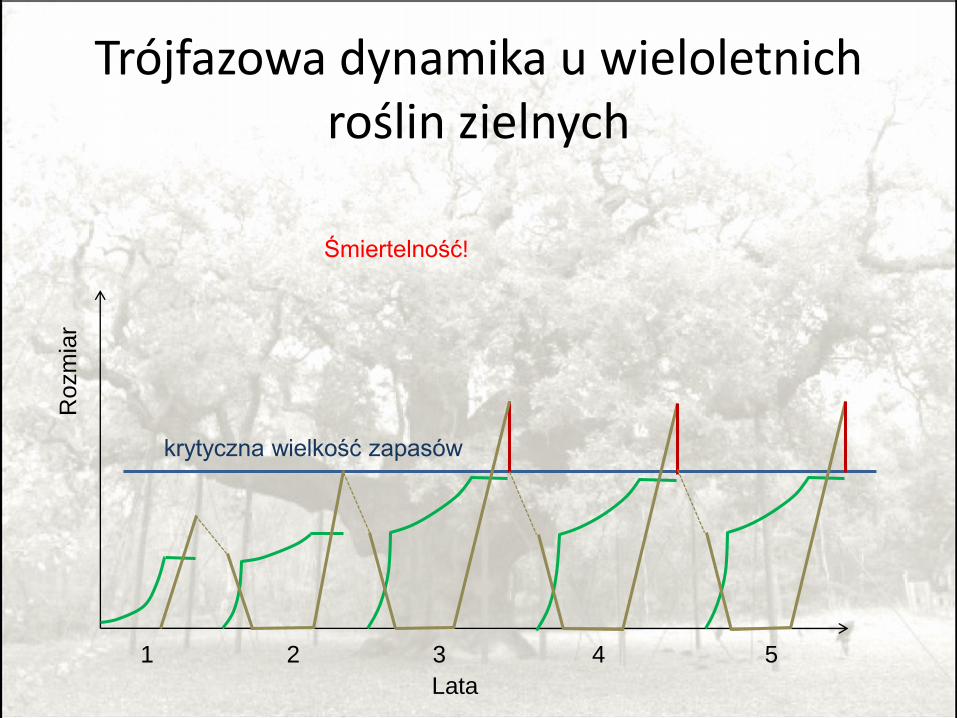
Trójfazowa dynamika u wieloletnich roślin zielnych
Rozm
iar
Lata
1 2 3 4 5
krytyczna wielkość zapasów
Śmiertelność!

Trójfazowa dynamika u wieloletnich roślin zielnych
• Krytyczny poziom zasobów zależy od śmiertelności.
• Przy bardzo wysokiej śmiertelności krytyczny poziom zasobów spada do zera = preferowana jednoroczność.
Rozm
iar
Lata1 2 3 4 5
Krytyczny poziom zasobów
Pugliese 1987; inne metody; Iwasa 1989; PMP
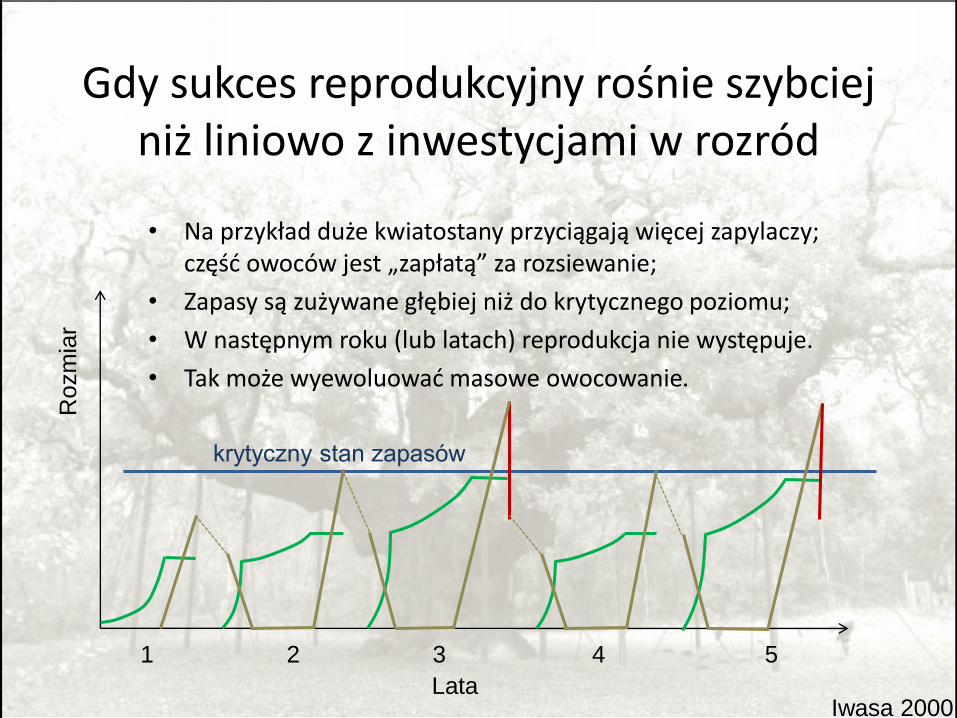
Gdy sukces reprodukcyjny rośnie szybciej niż liniowo z inwestycjami w rozród
• Na przykład duże kwiatostany przyciągają więcej zapylaczy; część owoców jest „zapłatą” za rozsiewanie;
• Zapasy są zużywane głębiej niż do krytycznego poziomu;
• W następnym roku (lub latach) reprodukcja nie występuje.
• Tak może wyewoluować masowe owocowanie.
Rozm
iar
Lata
1 2 3 4 5
krytyczny stan zapasów
Iwasa 2000

Optymalność rozmnażanie się raz w ciągu życia
• Paradoks Cole’s (1954) paradox:– Pytanie: Dlaczego powszechne jest rozmnażanie się wiele razy w życiu?
Wystarczy wyprodukować jednego więcej potomka, by zastąpić siebie.
– Odpowiedź (Charnov & Schaffer 1973): ponieważ śmiertelność osobników młodych jest większa niż dorosłych.
• Odwrócony paradoks Cole’s:– Prawdopodobieństwo dożycia od stadium nasienia do dorosłej rośliny
jest bardzo niskie. Dlaczego nie oszczędzić trochę zasobów, przeżyć i rozmnażać się wielokrotnie?
• Prawdopodobna odpowiedź (Klinkhamer et al. 1997):– Warunek konieczny: sukces reprodukcyjny rośnie szybciej niż liniowo z
nakładami na reprodukcję: trzeba wielu lat by odtworzyć zapasy.
– Rośliny kwitnące muszą manifestować swą obecność, co przyciąga też wrogów; patogeny i roślinożercy obniżają przeżywanie dorosłych po kwitnieniu/owocowaniu.

Rośliny jednoroczne: Dlaczego przełączenie zwykle nie jest zero-jedynkowe?
• Nieliniowość kontroli? Mało prawdopodobny kierunek nieliniowości.
• Rozwiązanie osobliwe (singular solution)? Mało prawdopodobne w dłuższym przedziale czasu
• Stochastyczna długość sezonu? Prawdopodobnie możliwe, ale problem słabo rozpracowany teoretycznie.
• „Ograniczenie konstrukcyjne”? Najbardziej prawdopodobne, tłumaczy też podobne zjawisko u roślin wieloletnich.
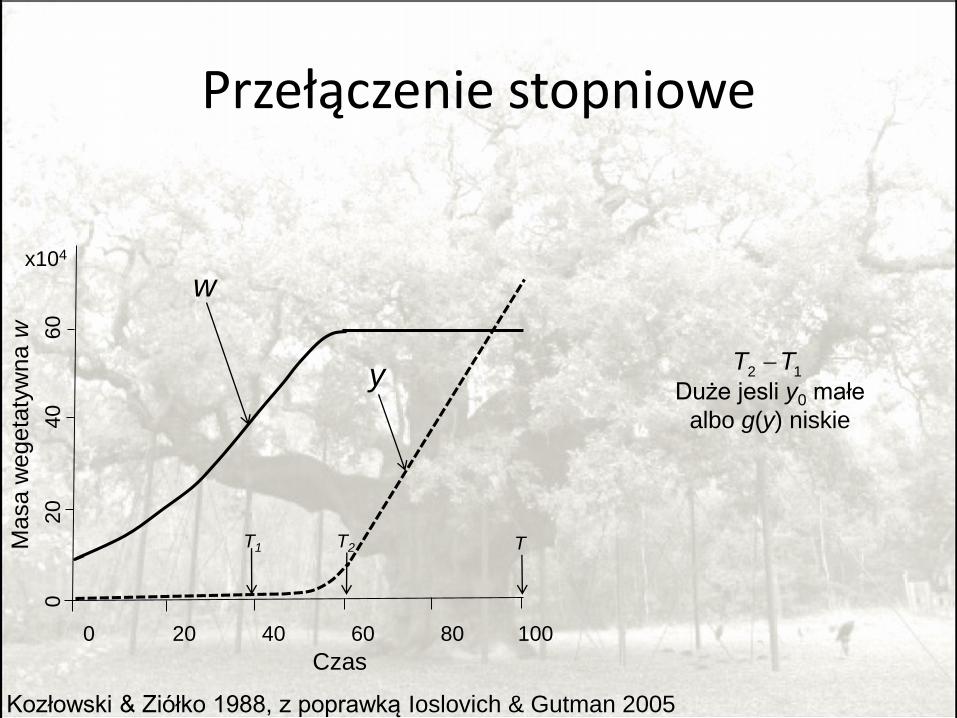
Przełączenie stopniowe
Kozłowski & Ziółko 1988, z poprawką Ioslovich & Gutman 2005
Duże jesli y0 małe
albo g(y) niskie
2 1T T
Czas
0 20 40 60 80 100
TT2T1Masa w
egeta
tyw
na w
0
20
40
60
x104
w
y
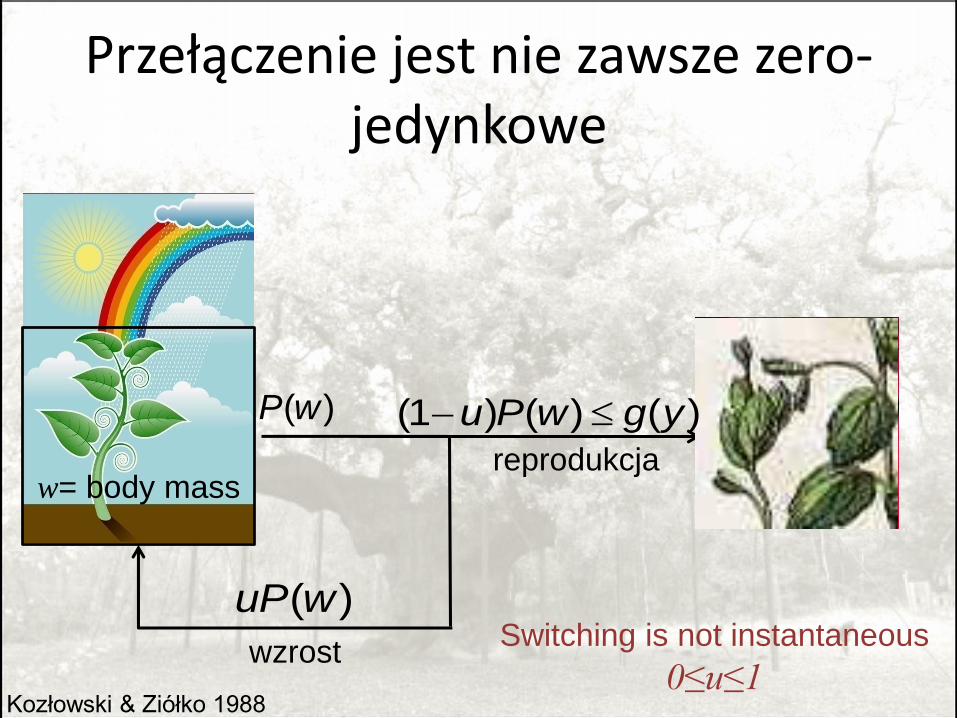
Przełączenie jest nie zawsze zero-jedynkowe
( )P w
( )uP w
wzrost
reprodukcjaw= body mass
Switching is not instantaneous
0≤u≤1Kozłowski & Ziółko 1988
(1 ) ( ) ( )u P w g y

Dlaczego istnieją rośliny zielne, krzewinki, krzewy i drzewa?

Zwierzęta
okres alokacji w
reprodukcję
rozm
iar
wiek/czas
year 1 year 2 year 3 year 4
w
i
n
t
e
r
w
i
n
t
e
r
w
i
n
t
e
r
w
i
n
t
e
r
wzrost asymptotyczny
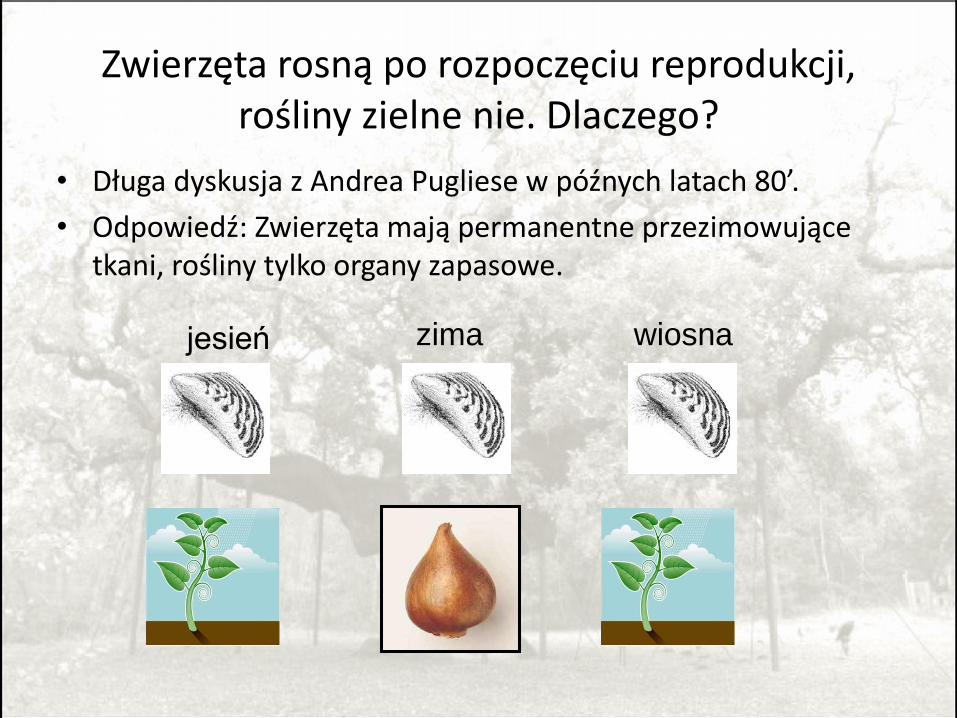
Zwierzęta rosną po rozpoczęciu reprodukcji, rośliny zielne nie. Dlaczego?
• Długa dyskusja z Andrea Pugliese w późnych latach 80’.
• Odpowiedź: Zwierzęta mają permanentne przezimowujące tkani, rośliny tylko organy zapasowe.
jesień zima wiosna

Gdy tylko część tkanek jest permanentna
• Istnieje krytyczna frakcja permanentnych tkanek q, poniżej tylko dorastanie do poprzednich rozmiarów, powyżej powiększanie rozmiarów.Do puli wchodzą też zasoby wycofywane z traconych na zimę liści.
• Krytyczna frakcja zależy od:
– prawdopodobieństwa przeżycia
– kształtu krzywej produkcji
optymalny wzrost
po dojrzewaniu
stały rozmiar po
dojrzewaniu
Pugliese and Kozlowski 1990prawdopodobieństwo przeżycia
kry
tyczne q
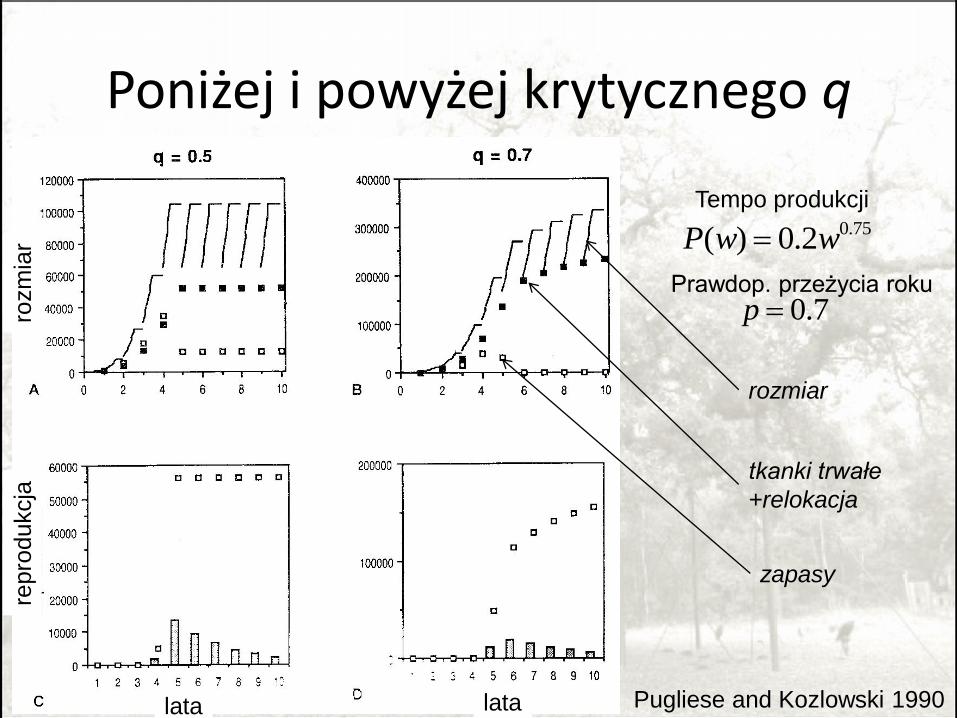
Poniżej i powyżej krytycznego q
Pugliese and Kozlowski 1990
0.75( ) 0.2P w w
0.7p Prawdop. przeżycia roku
Tempo produkcji
rozmiar
tkanki trwałe
+relokacja
zapasy
rozm
iar
repro
dukcja
lata lata

0
0,2
0,4
0,6
0,8
1
0 0,2 0,4 0,6 0,8 1
crit
ical
q
survival probability p
b=1
b=0.67
b=0.50
Krytyczne q a ilość zdrewniałych tkanek
• Więcej zdrewniałych tkanek = większy udział permanentnej masy
• Więcej zdrewniałych tkanek = bardziej wypukła krzywa produkcji
Pugliese and Kozlowski 1990
rośliny zielne
drzewa
prawdopodobieństwo przeżycia
kry
tyczne q

Zgryzanie i obrona
• h – tempo zgryzania
• w – masa części wegetatywnych
• z – ilość (nie stężenie) substancji obronnych
Janczur 2009
1
2
3
1 2 3
( )
( )
( )
1
w u P w hw
y u P w
z u P w hz
u u u

• Zmiana jednego założenia:
– Asymilowana masa trafia najpierw do zapasów, a stamtąd jest dystrybuowana do części wegetatywnych, generatywnych lub pozostaje
• Powstał model ogólny, obejmujący szereg omówionych modeli
– Sezonowa zmiana warunków środowiskowych ma charakter ciągły

Mironchenko & Kozłowski 2014Układ równań
części wegetatywne
części generatywne
zapasy
Zmienne sterujące:
v(t) = frakcja maksymalnego tempa zużywania zasobów
v1(t) = frakcja lokowana we wzrost masy wegetatywnej
v2(t) = [v(t) - v1(t)] = frakcja lokowana w rozród
Zmienne sezonowe:
f, μ, ω = funkcje zależne od sezonu
Maksymalizowany funkcjonał: Metoda PMP
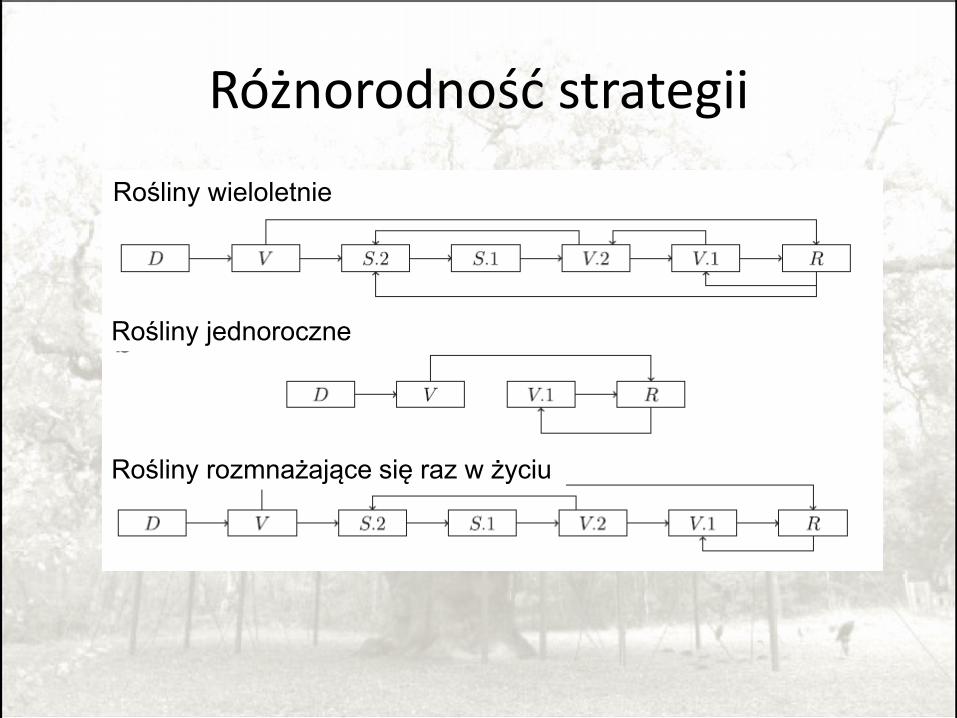
Różnorodność strategii
Rośliny wieloletnie
Rośliny rozmnażające się raz w życiu
Rośliny jednoroczne
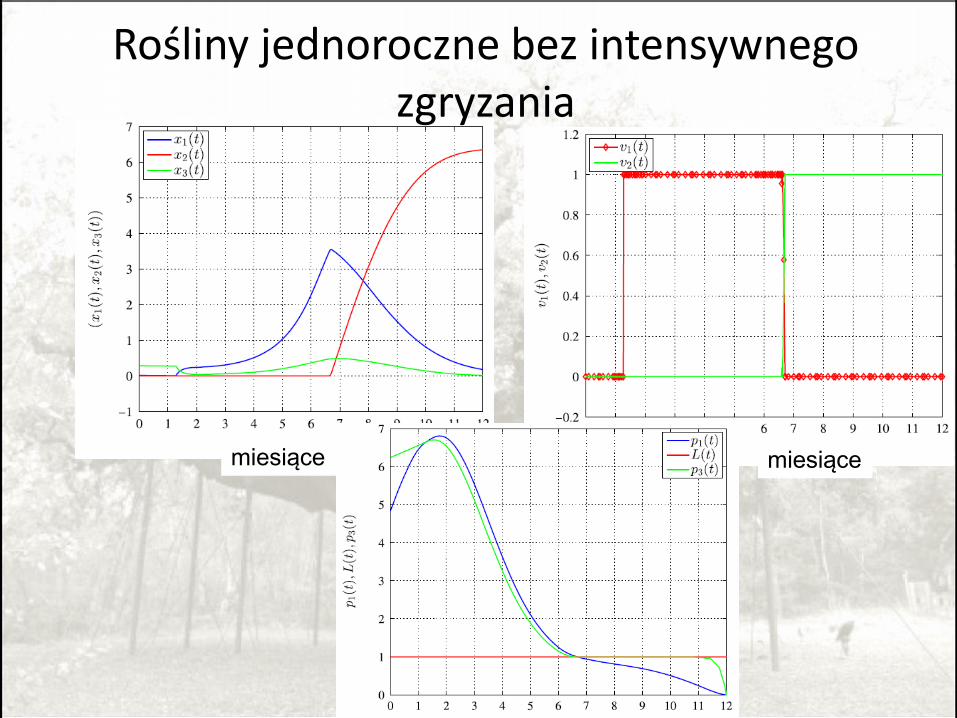
Rośliny jednoroczne bez intensywnego zgryzania
miesiące miesiące
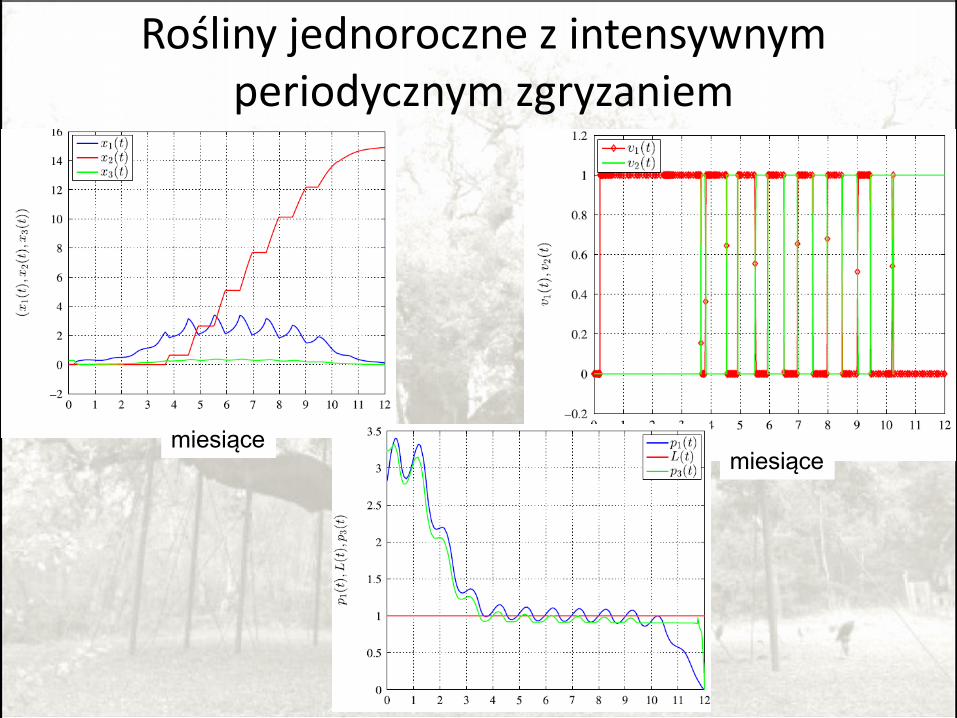
Rośliny jednoroczne z intensywnym periodycznym zgryzaniem
miesiącemiesiące
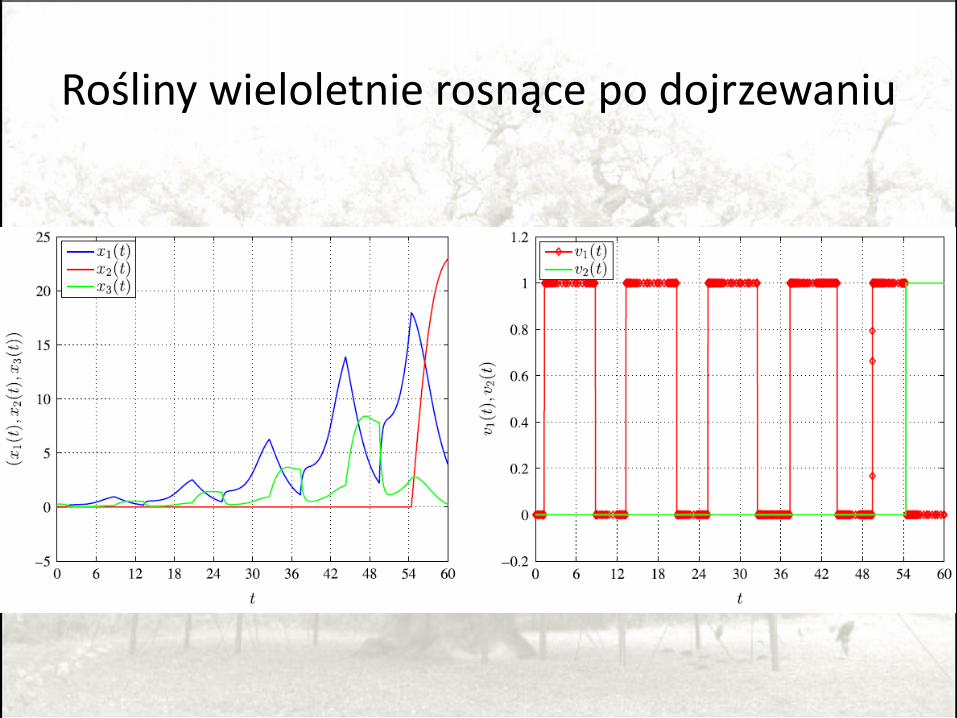
Rośliny wieloletnie rosnące po dojrzewaniu
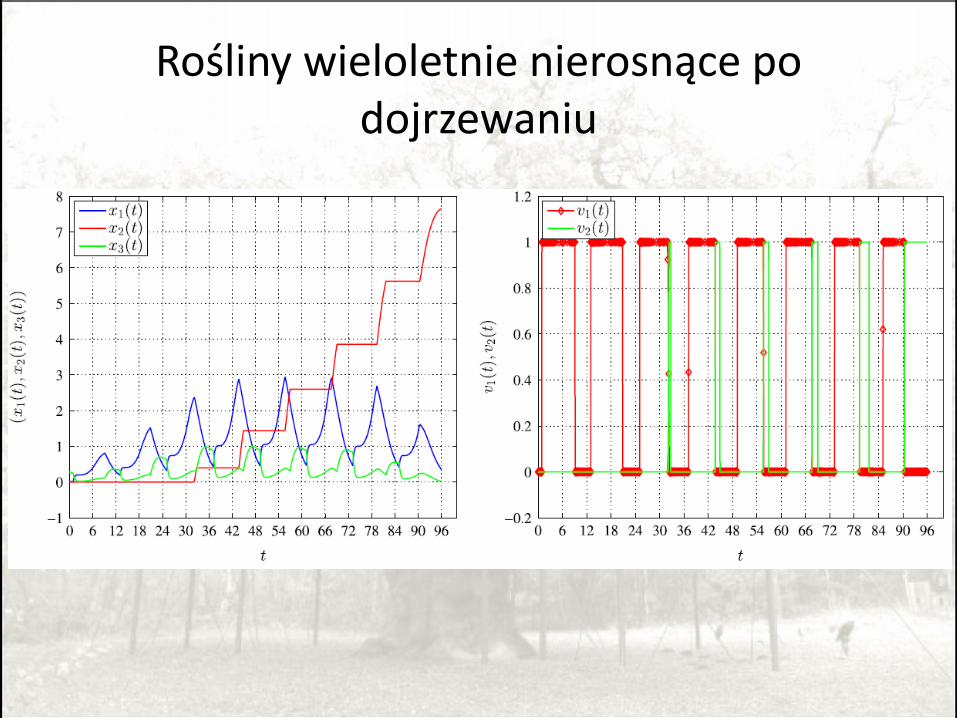
Rośliny wieloletnie nierosnące po dojrzewaniu
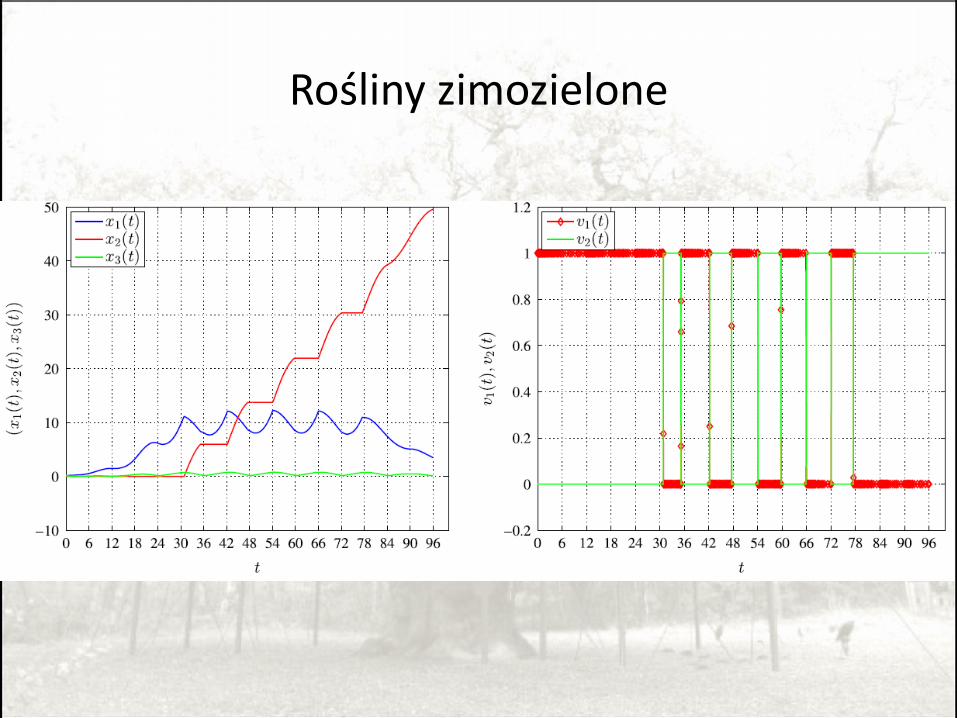
Rośliny zimozielone

Alokacja w rozród wegetatywny i płciowy
• Nasiona mają minimalne szanse wygrać konkurencję o miejsce z rametami;
• Nasiona muszą znaleźć puste miejsce by wykiełkować.
Olejniczak 2001
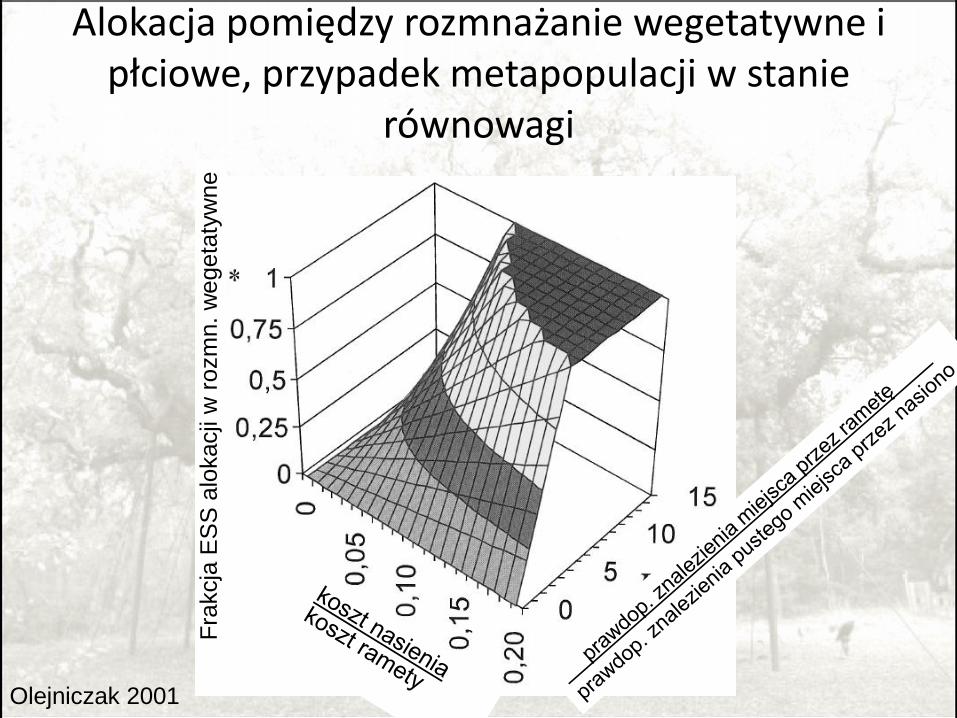
Alokacja pomiędzy rozmnażanie wegetatywne i płciowe, przypadek metapopulacji w stanie
równowagi
Olejniczak 2001
Fra
kcja
ES
S a
lokacji
w r
ozm
n.
wegeta
tyw
ne

Alokacja w rozród wegetatywny i płciowy, efekt lokalnego zagęszczenia ramet
Olejniczak 2003
lokalne zagęszczenie ramet N
= krytyczne zagęszczenie ramet
Op
tym
aln
a a
lokacja
w
rozró
d w
egeta
tyw
ny
N
N zależy od: koszt nasieniakoszt ramety
prawdopodobieństwa rekrutacji ramety
,
,

Dziękuję!
Dąb Robin Hooda
Wikipedia
Masa oceniana na 23 ton, obwód10m, wiek 800-1000 lat


















