
Wchłanianie, przemiany metaboliczne i wydalanie nikotyny u ...
12
Postępy Biochemii 59 (1) 2013 33 Robert Sobkowiak Andrzej Lesicki Zakład Biologii Komórki, Instytut Biologii Eks- perymentalnej, Uniwersytet im. Adama Mic- kiewicza, Poznań Zakład Biologii Komórki, Instytut Biologii Eksperymentalnej, Uniwersytet im. Adama Mickiewicza, ul. Umultowska 89, 61-614 Po- znań; tel.: (61) 829 58 22, e-mail: robsob@amu. edu.pl Artykuł otrzymano 3 lipca 2012 r. Artykuł zaakceptowano 11 września 2012 r. Słowa kluczowe: nikotyna, kotynina, meta- bolizm, wchłanianie, wydalanie, enancjomer, cytochrom P450 Wykaz skrótów: CYP (ang. cytochrome P450 oxidase) — oksydaza cytochromu P450; CY- P2A6, CYP2B6, CYP2E1, CYP3A4 — odmiany oksydazy cytochromu P450; FMO (ang. flavin containing monooxygenase 3) — monooksyge- naza flawinowa; UGT (ang. uridine diphosphate glucuronosyltransferase) — transferaza urydy- nodifosfoglukuronowa Wchłanianie, przemiany metaboliczne i wydalanie nikotyny u człowieka STRESZCZENIE N ikotyna jest alkaloidem występującym w wielu roślinach z rodziny psiankowatych. Jest zwykle mieszaniną dwóch izomerów optycznych, z których zdecydowanie domi- nuje lewoskrętny enancjomer (S). Nikotyna najczęściej przenika do organizmu człowieka, jako jeden ze składników dymu tytoniowego. Tempo przenikania nikotyny przez błony bio- logiczne wzrasta w środowisku alkalicznym. Niemal 90% nikotyny pobranej przez organizm człowieka metabolizowane jest w wątrobie. Nikotyna może również być metabolizowana w nerkach, płucach, mózgu oraz błonach śluzowych dróg oddechowych. Nikotyna w orga- nizmie człowieka ulega bardzo skomplikowanym przemianom. Kluczową rolę w metabo- lizmie nikotyny odgrywają oksydazy cytochromu P450 (przede wszystkim ich forma CY- P2A6). Poza nimi w procesie rozkładu nikotyny zaangażowane są transferazy urydynodifos- foglukuronowe, cytosolowe oksydazy aldehydowe, N-metylotransferazy aminowe oraz mo- nooksygenazy flawinowe typu 3. Zidentyfikowano sześć głównych metabolitów nikotyny, spośród których procentowo najliczniejszym jest kotynina. Z niej powstaje trans-3’-hydrok- sykotynina, związek wydalany przede wszystkim wraz z moczem. Na tempo metabolizmu nikotyny ma wpływ zróżnicowana aktywność polimorficznych enzymów uczestniczących w jej przemianach, dieta, płeć oraz stan fizjologiczny organizmu. WPROWADZENIE Nikotyna jest organicznym związkiem chemicznym z grupy alkaloidów piry- dynowych zawartym przede wszystkim w różnych gatunkach tytoniu oraz in- nych roślin z rodziny psiankowatych (Solanaceae), takich jak ziemniaki, pomido- ry, papryka czy bakłażan. Suche liście tytoniu szlachetnego Nicotiana tabacum za- wierają od 2 do 8% nikotyny, natomiast tytoń indiański („dziki tytoń”) Nicotiana rustica może zawierać nawet do 18% tego alkaloidu [1,2]. Nikotyna została wy- izolowana po raz pierwszy w 1828 r. [3]. Struktura nikotyny, 3-(1-metylo-2-piro- lidyno) pirydyna, C 10 H 14 N 2 , została zaproponowana w 1892 roku i potwierdzona w 1895 roku w wyniku syntezy [3]. Czysta nikotyna to klarowny płyn o charak- terystycznym, intensywnym zapachu, brunatniejący na powietrzu [1]. Nikotyna może występować w postaci wolnej zasady lub jako sól dwuchlorowodorkowa, salicylowa, siarczanowa lub dwuwinianowa. Szczegółowy opis właściwości fi- zyko-chemicznych nikotyny można odnaleźć w opracowaniu O’Neil i wsp. [1]. Hydrofobowe właściwości pirydyny i pierścienia pirolidynowego w cząsteczce nikotyny nadają jej niską polarność, co powoduje dobrą rozpuszczalność tego alkaloidu w lipidach. Nikotyna jest aminą zbudowaną z dwóch pierścieni heterocyklicznych, piry- dyny i pirolidyny, której atom węgla w pozycji 2 stanowi centrum chiralne [3,4]. Stąd występują dwa izomery optyczne (enancjomery) nikotyny, które skręcają światło spolaryzowane w przeciwnych kierunkach (Ryc. 1) [3]. Poza tym więk- szość własności fizycznych oraz chemicznych dla obu enancjomerów jest niemal identyczna. Szczegółowe informacje można znaleźć w bazach danych np. Pub- Chem (http://pubchem.ncbi.nlm.nih.gov) lub Chemical Entities of Biological Interest (ChEBI) (http://www.ebi.ac.uk/chebi). W przypadku nikotyny natural- ny produkt jest zdominowany przez lewoskrętny enancjomer (S) (ChEBI:17688) tradycyjnie nazywany L- lub (-)-nikotyną, natomiast zawartość prawoskrętnego (R)-enancjomeru nikotyny (określa- nego również jako D- lub (+)-nikoty- na, ChEBI:39162) zwykle nie prze- kracza kilku pro- cent. Zmiany pH wywołują szereg Rycina 1. Enancjomery nikotyny. A) (S)-nikotyna B) (R)-nikotyna
Transcript of Wchłanianie, przemiany metaboliczne i wydalanie nikotyny u ...

Postępy Biochemii 59 (1) 2013 33
Robert Sobkowiak
Andrzej Lesicki
Zakład Biologii Komórki, Instytut Biologii Eks-perymentalnej, Uniwersytet im. Adama Mic-kiewicza, Poznań
Zakład Biologii Komórki, Instytut Biologii Eksperymentalnej, Uniwersytet im. Adama Mickiewicza, ul. Umultowska 89, 61-614 Po-znań; tel.: (61) 829 58 22, e-mail: [email protected]
Artykuł otrzymano 3 lipca 2012 r.Artykuł zaakceptowano 11 września 2012 r.
Słowa kluczowe: nikotyna, kotynina, meta-bolizm, wchłanianie, wydalanie, enancjomer, cytochrom P450
Wykaz skrótów: CYP (ang. cytochrome P450 oxidase) — oksydaza cytochromu P450; CY-P2A6, CYP2B6, CYP2E1, CYP3A4 — odmiany oksydazy cytochromu P450; FMO (ang. flavin containing monooxygenase 3) — monooksyge-naza flawinowa; UGT (ang. uridine diphosphate glucuronosyltransferase) — transferaza urydy-nodifosfoglukuronowa
Wchłanianie, przemiany metaboliczne i wydalanie nikotyny u człowieka
STRESZCZENIE
Nikotyna jest alkaloidem występującym w wielu roślinach z rodziny psiankowatych. Jest zwykle mieszaniną dwóch izomerów optycznych, z których zdecydowanie domi-
nuje lewoskrętny enancjomer (S). Nikotyna najczęściej przenika do organizmu człowieka, jako jeden ze składników dymu tytoniowego. Tempo przenikania nikotyny przez błony bio-logiczne wzrasta w środowisku alkalicznym. Niemal 90% nikotyny pobranej przez organizm człowieka metabolizowane jest w wątrobie. Nikotyna może również być metabolizowana w nerkach, płucach, mózgu oraz błonach śluzowych dróg oddechowych. Nikotyna w orga-nizmie człowieka ulega bardzo skomplikowanym przemianom. Kluczową rolę w metabo-lizmie nikotyny odgrywają oksydazy cytochromu P450 (przede wszystkim ich forma CY-P2A6). Poza nimi w procesie rozkładu nikotyny zaangażowane są transferazy urydynodifos-foglukuronowe, cytosolowe oksydazy aldehydowe, N-metylotransferazy aminowe oraz mo-nooksygenazy flawinowe typu 3. Zidentyfikowano sześć głównych metabolitów nikotyny, spośród których procentowo najliczniejszym jest kotynina. Z niej powstaje trans-3’-hydrok-sykotynina, związek wydalany przede wszystkim wraz z moczem. Na tempo metabolizmu nikotyny ma wpływ zróżnicowana aktywność polimorficznych enzymów uczestniczących w jej przemianach, dieta, płeć oraz stan fizjologiczny organizmu.
WPROWADZENIE
Nikotyna jest organicznym związkiem chemicznym z grupy alkaloidów piry-dynowych zawartym przede wszystkim w różnych gatunkach tytoniu oraz in-nych roślin z rodziny psiankowatych (Solanaceae), takich jak ziemniaki, pomido-ry, papryka czy bakłażan. Suche liście tytoniu szlachetnego Nicotiana tabacum za-wierają od 2 do 8% nikotyny, natomiast tytoń indiański („dziki tytoń”) Nicotiana rustica może zawierać nawet do 18% tego alkaloidu [1,2]. Nikotyna została wy-izolowana po raz pierwszy w 1828 r. [3]. Struktura nikotyny, 3-(1-metylo-2-piro-lidyno) pirydyna, C10H14N2, została zaproponowana w 1892 roku i potwierdzona w 1895 roku w wyniku syntezy [3]. Czysta nikotyna to klarowny płyn o charak-terystycznym, intensywnym zapachu, brunatniejący na powietrzu [1]. Nikotyna może występować w postaci wolnej zasady lub jako sól dwuchlorowodorkowa, salicylowa, siarczanowa lub dwuwinianowa. Szczegółowy opis właściwości fi-zyko-chemicznych nikotyny można odnaleźć w opracowaniu O’Neil i wsp. [1]. Hydrofobowe właściwości pirydyny i pierścienia pirolidynowego w cząsteczce nikotyny nadają jej niską polarność, co powoduje dobrą rozpuszczalność tego alkaloidu w lipidach.
Nikotyna jest aminą zbudowaną z dwóch pierścieni heterocyklicznych, piry-dyny i pirolidyny, której atom węgla w pozycji 2 stanowi centrum chiralne [3,4]. Stąd występują dwa izomery optyczne (enancjomery) nikotyny, które skręcają światło spolaryzowane w przeciwnych kierunkach (Ryc. 1) [3]. Poza tym więk-szość własności fizycznych oraz chemicznych dla obu enancjomerów jest niemal identyczna. Szczegółowe informacje można znaleźć w bazach danych np. Pub-Chem (http://pubchem.ncbi.nlm.nih.gov) lub Chemical Entities of Biological Interest (ChEBI) (http://www.ebi.ac.uk/chebi). W przypadku nikotyny natural-ny produkt jest zdominowany przez lewoskrętny enancjomer (S) (ChEBI:17688) tradycyjnie nazywany L- lub (-)-nikotyną, natomiast zawartość prawoskrętnego
(R)-enancjomeru nikotyny (określa-nego również jako D- lub (+)-nikoty-na, ChEBI:39162) zwykle nie prze-kracza kilku pro-cent.
Zmiany pH wywołują szereg Rycina 1. Enancjomery nikotyny. A) (S)-nikotyna B) (R)-nikotyna

34 www.postepybiochemii.pl
zmian struktury cząsteczkowej nikotyny. Równowaga ter-modynamiczna pomiędzy poszczególnymi strukturami molekularnymi nikotyny ustala się samorzutnie zgodnie z prawami termodynamiki (Ryc. 2) [5].
Znaczący postęp w badaniach nad metabolizmem niko-tyny, obserwowany od końca lat osiemdziesiątych ubiegłe-go wieku, związany był z udoskonaleniem metod oczysz-czania enzymów i uzyskiwania wysoce specyficznych prze-ciwciał, a także z rozwojem technik immunochemicznych i biochemicznych oraz z upowszechnieniem wysokospraw-nej chromatografii cieczowej (HPLC). Przemiany nikotyny w organizmach żywych obejmują wiele skomplikowanych procesów, które rozpoczynają się tuż po przedostaniu się alkaloidu do organizmu poprzez błony komórkowe. Celem niniejszej pracy jest przedstawienie farmakokinetyki ni-kotyny, ze szczególnym uwzględnieniem jej metabolizmu oraz czynników mogących wpływać na omawiane procesy.
WCHŁANIANIE NIKOTYNY
Przypuszcza się, że przenikalność przez błonę komór-kową, wynikająca z dobrej rozpuszczalności w jej lipido-wych warstwach, jest taka sama w przypadku różnych enancjomerów nikotyny [4], natomiast tempo przenikania nikotyny poprzez błonę jest zależne od jej stężenia oraz pH środowiska [4,7]. Maleje wraz ze wzrostem stężenia niko-tyny, jednak jej transport przez błonę nie polega tylko na prostej dyfuzji. Badania, w których wykazano, że nikotyna hamuje przenoszenie kationów organicznych przez trans-portery należące do rodziny OCT (ang. organic cation trans-porter) i OCTN (ang. organic cation/carnitine transporter) [8,9], sugerują, że nikotyna może być transportowana przez tego rodzaju przenośniki [9,10]. Przepuszczalność błony komór-kowej dla nikotyny rośnie wraz ze wzrostem pH (w zakre-sie pH 5,5–8,1) [7,11,12], gdyż nikotyna jako słaba zasada w środowisku o obniżonej zawartości H+ (zasadowym) staje się cząsteczką niezjonizowaną i dlatego łatwiej przenika przez niepolarną błonę komórkową. Obecność w cząstecz-
ce nikotyny grupy pirolidinowej oraz azotu pirydynowego decyduje o jej dwuzasadowym charakterze.
Nikotyna najczęściej przenika do organizmu człowieka, jako jeden ze składników dymu papierosowego. Wydajność absorpcji nikotyny w pęcherzykach płucnych zależy od jej stężenia w dymie. Jednak tylko 10% nikotyny zawartej w dymie przyswajanych jest przez organizm człowieka [13]. Jeden papieros zawiera około 1-2% nikotyny (ok. 0,8-1,9 mg) [13]. Przy średniej wadze człowieka 70 kg jeden papie-ros dostarcza nikotyny w dawce około 10-30 µg/kg masy ciała. W obrębie pęcherzyków płucnych pH wynosi około 7,4, a w tych warunkach około 31% nikotyny jest w formie niezjonizowanej. Nikotyna z łatwością przedostaje się przez błonę komórkową pneumocytów i dalej do krwiobiegu [4].
Wchłanianie nikotyny może także zachodzić poprzez błonę śluzową jamy ustnej lub nosa (np. nie tylko podczas palenia, ale też żucia tytoniu lub zażywania tabaki). Szyb-kość oraz ilość wchłoniętej nikotyny zależy od rodzaju zaży-wanego środka zawierającego ten alkaloid oraz od sposobu jego podawania [10,14-16]. Po eksperymentalnym podaniu nikotyny przez błonę śluzową nosa, obserwowano wzrost jej poziomu we krwi prawie tak szybki, jak po wypaleniu papierosa [4]. W przeciwieństwie do dymu papierosowe-go, który zwykle ma kwaśne pH (ok. 5,5), dym pochodzą-cy z cygar, a także tytoń do żucia, tabaka i nikotynizowane gumy do żucia są łagodnie zasadowe. Ich producenci wpro-wadzają buforujące dodatki, takie jak amoniak, podnoszące ich pH do ok. 8,5, zwiększając tym samym siłę działania tej samej dawki nikotyny, by przy wyższym pH była wydajniej wchłaniana do organizmu [15,17]. Efekt ten nie został jed-noznacznie potwierdzony, bowiem kwaśny lub zasadowy odczyn dymu papierosowego lub cygar może być neutra-lizowany przez buforujące właściwości płynu pokrywają-cego nabłonek oddechowy pęcherzyków płucnych [17,18]. Wchłanianie nikotyny przez błonę śluzową jamy ustnej jest ułatwione, bo w zasadowym pH, jak już wspomniano, jest ona w dużej mierze niezjonizowana. Tempo wzrostu stęże-nia nikotyny we krwi podczas żucia tytoniu lub nikotyni-
Rycina 2. Strukturalne przekształcenia (S)-nikotyny wywołane zmianami pH środowiska. A) możliwe struktury cząsteczkowe (S)-nikotyny. B) zmiany energii swobodnej oraz stałe dysocjacji poszczególnych form (S)-nikotyny, pKa — ujemny logarytm dziesiętny ze stałej dysocjacji (Ka); ΔG — zmiana energii swobodnej; k — szybkość reakcji. Na podstawie (B) [5] oraz (A) [6], za zgodą Amerykańskiego Towarzystwa Chemicznego (copyright 2005, 2011).

Postępy Biochemii 59 (1) 2013 35
zowanej gumy jest wolniejsze niż w przypadku palenia pa-pierosów, chociaż osiągana w końcu maksymalna wartość stężenia związku we krwi jest zbliżona [10], a podwyższone stężenie we krwi utrzymuje się dłużej [14].
Absorpcja nikotyny w dalszych odcinkach układu pokar-mowego jest silnie uzależniona od panującego w nich pH [19]. Nikotyna połykana ze śliną, w kwaśnym środowisku soku żołądkowego, przekracza błony bardzo wolno, gdyż znaczna część cząsteczek nikotyny jest silnie zjonizowana [3,10]. W ciągu 15 min przy pH 1 absorbowane jest jedynie 3% połkniętej nikotyny [19]. Dobrze natomiast wchłaniana jest w jelicie cienkim, które ma lekko zasadowe pH i dużą powierzchnię absorpcji [4].
Duże ilości nikotyny mogą być wprowadzane do orga-nizmu człowieka w trakcie nikotynowych terapii zastęp-czych, w których stosuje się gumy do żucia lub naklejane na skórę plastry nikotynowe [13]. Do absorpcji nikotyny przez skórę może także dochodzić podczas ręcznego zbioru liści tytoniu. Kolejnym miejscem wnikania nikotyny do organizmu człowieka może być pęcherz moczowy [3], z któ-rego wchłaniana jest zwrotnie, a tempo wchłaniania zależy od pH moczu. Przenikanie nikotyny przez błonę śluzową pęcherza moczowego zwiększa się, gdy alkaloid jest niezjo-nizowany, czyli wtedy, kiedy pH moczu mieści się w grani-cach 8,0 do 9,0. Nikotyna nie ulega resorpcji z moczu, gdy pH moczu jest poniżej 6,0.
Nikotyna po pokonaniu powłok ciała wraz z krwią roz-prowadzana jest po całym organizmie, gdzie poprzez błony plazmatyczne wnika do komórek. Wykazano, że stężenie nikotyny w osoczu biernych palaczy wynosi około 2,5-8,0 ng/ml, podczas gdy poziom nikotyny w osoczu u osób pa-lących wynosi 30-40 ng/ml [13,15]. W fizjologicznym pH krwi (ok. 7,4) nikotyna w 69% jest zjonizowana. Ocenia się, że około 5 % nikotyny wiąże się z albuminami osocza [14] lub innymi białkami krwi (do 20 %) [19]. Maksymalne stę-żenie nikotyny w krwi tętniczej palacza tuż po wypaleniu papierosa może wynosić nawet 100 ng/ml (0,6 µM) [13]. Po około 20 minutach stężenie nikotyny we krwi szybko spada w wyniku przenikania nikotyny do tkanek. W ciągu 8-20 sekund nikotyna dociera do mózgu, gdzie łatwo przenika przez barierę krew-mózg i wnika wydajnie do komórek nerwowych [1,17]. Akumulacja nikotyny w innych tkan-kach zachodzi w różnym stopniu [18]. Najwyższe powino-wactwo do nikotyny wykazują: wątroba, nerki, śledziona i płuca, a najniższe tkanka tłuszczowa. Poziom nikotyny w mięśniach szkieletowych jest podobny jak we krwi. Aku-mulację nikotyny obserwuje się również w komórkach za-wierających melaninę np. w melanocytach [20]. Nikotyna łatwo pokonuje też łożysko, jej stężenie w krwi pępowino-wej wynosi od 0,5 do 25 ng/ml, wnika do krwi płodu oraz do płynu owodniowego, gdzie jej stężenie może się wahać w zakresie 1,5 do 23 ng/ml [13,19]. Koncentracja nikotyny w mleku palącej tytoń matki jest około 3 krotnie większa niż we krwi [13]. Stężenie nikotyny w płynie pozakomórko-wym poszczególnych tkanek, może znacznie się różnić od stężenia wewnątrz komórek.
BIOLOGICZNE WŁAŚCIWOŚCI ENANCJOMERóW NIKOTYNY
Działanie nikotyny intensywnie badano w warunkach zarówno in vitro, jak i in vivo na organizmach zwierząt, a także człowieka. Pojawienie się nikotyny w organizmie wy-wołuje przyspieszenie tętna oraz wzrost ciśnienia krwi, pro-wadzi do podwyższenia poziomu wolnych kwasów tłusz-czowych, glukozy oraz katecholamin [3]. Silne działanie ni-kotyny zaobserwowano także na poziomie komórkowym, głównie poprzez receptory nikotynowe [3,21,22].
Liczne badania wykazały, że działanie farmakologicz-ne (R)-nikotyny, choć jakościowo podobne, jest ilościowo słabsze niż (S)-nikotyny. Prawie identyczne właściwości fizykochemiczne obu enancjomerów skutkują jednakową kinetyką wchłaniania S-nikotyny i R-nikotyny, niezależną od sposobu podania alkaloidu do organizmu. Jednak oba enancjomery wykazują odmienne powinowactwo do róż-nych tkanek i narządów. Różnice te najprawdopodobniej wynikają z obecności różnych receptorów acetylocholiner-gicznych wiążących nikotynę w błonach komórkowych określonych typów komórek [21] oraz z aktywacji charak-terystycznych dla nich szlaków metabolicznych, w których uczestniczą białka wykazujące powinowactwo do nikotyny. Wydaje się, że czynniki te decydują o różnej cytotoksyczno-ści obu enancjomerów [3,4].
Badania przeprowadzone in vivo na mózgach szczurów z wykorzystaniem znakowanych trytem lub węglem C14 izomerów nikotyny sugerują, że obydwie formy nikotyny mogą wiązać się do różnych typów receptorów nikotyno-wych [4,23]. Szacuje się, że średnie powinowactwo (R)-ni-kotyny do receptorów N-acetylocholinergicznych jest około dziesięć razy mniejsze niż (S)-nikotyny, jednak sposób wią-zania (R)-nikotyny i (S)-nikotyny przez receptory choliner-giczne typu N może się różnić tylko nieznacznie [4].
W trakcie wieloletnich badań okazało się, że enancjomery nikotyny wykazują zróżnicowaną aktywność biologiczną [3]. Ostra toksyczność i działanie farmakologiczne (R)-ni-kotyny zostały zbadane tylko na kilku gatunkach zwierząt. Porównawczo analizowano także działanie (S)-nikotyny. Dawka nikotyny, obliczana w miligramach na kilogram masy ciała, potrzebna do uśmiercenia 50% badanej popula-cji (LD50) po podawaniu dożylnym (R)-nikotyny jest około 18 razy większa (6,15 mg/kg) niż (S)-nikotyny (0,33 mg/kg) [3,23]. Różnice w farmakologicznej aktywności izomerów nikotyny zaobserwowano u małp w zapisie fal mózgowych (EEG), ciśnieniu krwi, częstości akcji serca, skurczu komó-rek gładkich jelita cienkiego i innych [23]. Badania beha-wioralne wykazały, że subiektywne hedonistyczne efekty odczuwane przez palaczy wywołane przez (R)-nikotynę są porównywalne do tych wywołanych przez (S)-nikotynę [4].
METABOLITY NIKOTYNY
Nikotyna pobierana przez nabłonek oddechowy pęche-rzyków płucnych, przez błonę śluzową nosa i jamy ustnej oraz przez skórę lub wchłaniana zwrotnie z moczu trafia bezpośrednio do krążenia i za jego pośrednictwem do po-szczególnych narządów, w tym do wątroby. Natomiast ni-

36 www.postepybiochemii.pl
kotyna wchłonięta w jelitach przenika do żylnego krążenia wrotnego, skąd bezpośrednio trafia do wątroby i tylko w niewielkim stopniu do pozostałych narządów [3,4].
Wątroba jest głównym miejscem przemian nikotyny w organizmie człowieka, bo blisko 90% zaabsorbowanej nikotyny jest metabolizowane w komórkach wątroby, he-patocytach [19]. Szlaki przemian metabolicznych nikotyny wykazują duży stopień złożoności [3,10,14,24,25]. Pomimo stosunkowo prostej budowy chemicznej nikotyna w orga-nizmie człowieka ulega bardzo skomplikowanym prze-mianom (Ryc. 3, Tab. 1). Nikotyna po przedostaniu się do wnętrza hepatocytu, tak jak inne ksenobiotyki, podlega procesom metabolicznym prowadzącym do jej unieszkodli-wienia, zwiększenia rozpuszczalności i w konsekwencji wy-dalenia. Cząsteczka nikotyny w swojej strukturze zawiera pierścień aromatyczny, alifatyczny węgiel oraz dwa atomy azotu. Istnieje zatem szereg miejsc, gdzie cząsteczka nikoty-ny może ulec reakcji utlenienia i koniugacji [3,10]. W czasie kilkudziesięcioletnich badań nad detoksykacją nikotyny zi-dentyfikowano wiele jej metabolitów wtórnych. Często sto-sowano różne nazwy dla tej samej pochodnej. Najczęściej używane alternatywne nazwy nikotyny oraz wielu jej meta-bolitów podano w tabeli 1.
W wątrobie nikotyna przekształcana jest do kilku związ-ków (Ryc. 3). Zidentyfikowano sześć głównych produktów przemian nikotyny [3,10,14,25]. Kluczową rolę w metaboli-zmie nikotyny, podobnie jak innych ksenobiotyków, odgry-wają oksydazy cytochromu P450 (CYP) [26]. Jedną z form tych oksydaz jest enzym określany jako CYP2A6, który przeprowadza reakcje: 5’-hydroksylacji, 2’-hydroksylacji i N-demetylacji (Ryc. 3).
Ilościowo najważniejszym metabolitem nikotyny u większości gatunków ssaków jest jej pochodna laktamo-wa, kotynina (Ryc. 3 i 4). U ludzi około 70-80% nikotyny przekształcana jest do kotyniny. Proponowany mechanizm przekształcenia nikotyny do kotyniny polega na jej hydrok-sylacji przez enzymy mikrosomalne hepatocytów, następ-nie konwersji do odpowiedniego aldehydu oraz produkcji kotyniny przez enzymy cytoplazmatyczne (Ryc. 3). W pro-ponowanym mechanizmie transformacji nikotyny w koty-ninę wyróżnić można kilka etapów. Pierwszy z nich kata-lizuje jedna z oksydaz cytochromu P450, określana obecnie jako CYP2A6 (dawniej nazywana hydroksylazą kumaryno-wą). W wyniku tej reakcji 5’-hydroksylacji z nikotyny po-wstaje jon Δ1’(5’) iminowy nikotyny (Ryc. 3) oraz pozostająca z nim w równowadze 5’-hydroksynikotyna [27]. W drugim etapie jest zaangażowana cytoplazmatyczna oksydaza alde-hydowa. Substratem dla niej jest jon Δ1’(5’)iminowy nikotyny, który wykazuje silne właściwości alkilacyjne, mogące od-grywać istotną rolę w farmakokinetyce nikotyny [3,14]. Za-zwyczaj proces metabolizmu ksenobiotyków po utlenieniu przez oksydazę cytochromu P450 obejmuje etap koniugacji ze związkiem wprowadzającym do metabolizowanego kse-nobiotyka polarne grupy funkcyjne. W tym etapie bardzo często dochodzi do sprzężenia ksenobiotyku z kwasem glu-kuronowym albo grupami tiolowymi glutationu bądź cy-steiny [23]. Jednakże, w niektórych przypadkach, w proce-sie eliminacji ksenobiotyków, mogą powstawać reaktywne półprodukty o właściwościach toksycznych lub rakotwór-czych [23].
Przyjmuje się, że mechanizm reakcji prowadzonej przez oksydazę cytochromu P450 polega na utlenieniu węgla trze-ciorzędowej aminy w pozycji α. Cytochrom P450 katalizu-je przeniesienie elektronu z niezwiązanej pary elektronów
Rycina 3. Kluczowe enzymy biorące udział w metabolizmie nikotyny: oksydaza cytochromu P450 (CYP), transferaza urydynodifosfoglukuronowa (UGT), oksydaza aldehydowa, N–metylotransferaza aminowa, monooksygenaza flawinowa 3 (FMO). Na podstawie [10], za zgodą Amerykańskiego Towarzystwa Farmakologii i Terapii Eksperymentalnej (copyright 2005).

Postępy Biochemii 59 (1) 2013 37
Tabela 1. Niektóre często używane alternatywne nazwy nikotyny oraz jej metabolitów (na podstawie [1,6,10,14,24,37,38]).
Polska nazwa związku Angielska nazwa związku Alternatywna nazwa w języku angielskim
nikotyna nicotine(54-11-5)a
(S)-nicotine(–)-nicotinel-nicotine3-(1-methyl-2-pyrrolidinyl)pyridineCAb: pyridine, 3-[(2S)-1-methyl-2-pyrrolidinyl]-
kotynina cotinine(486-56-6)
(S)-cotinine(–)-cotinine1-methyl-5-(3-pyridinyl)-2-pyrrolidinoneCA: 2-pyrrolidinone, 1-methyl-5-(3-pyridinyl)-, (5S)-
trans-3’-hydroksykotynina trans-3’-hydroxycotinine(34834-67-8)
hydroxycotinine3’-hydroxycotinine3-hydroxy-1-methyl-5-(3-pyridinyl)-2-pyrrolidinoneCA: 2-pyrrolidinone, 3-hydroxy-1-methyl-5-(3-pyridinyl)-, (3R,5S)-
N-tlenek nikotyny nicotine N’-oxide(491-26-9; trans: 51095-86-4)
nicotine 1’-oxidenicotine 1’-N-oxide1’(S)-2’(S)-trans-nicotine-N’-oxideCA: pyridine, 3-[(2S)-1-methyl-1-oxido-2-pyrrolidinyl]-trans, CA: pyridine, 3-[(1S,2S)-1-methyl-1-oxido-2-pyrrolidinyl]-
nornikotyninanornicotine(S-Isomer: 494-97-3; Racemic: 5746-86-1)
S-isomer: (–)-nornicotinel-nornicotineS-isomer, CA: pyridine, 3-(2S)-2-pyrrolidinyl-racemic: (±)-nornicotine(RS)-nornicotineracemic, CA: pyridine, 3-(2-pyrrolidinyl)-
norkotyninanorcotinine(S-isomer: 5980-06-3; racemic: 17708-87-1)
demethylcotinineS-isomer: (–)-demethylcotinineS-(–)-norcotinineS-isomer, CA: 2-pyrrolidinone, 5-(3-pyridinyl)-, (5S)-racemic: (±)-demethylcotinine(±)-Norcotinine(RS)-Norcotinineracemic, CA: 2-Pyrrolidinone, 5-(3-pyridinyl)-
N-tlenek kotyniny cotinine N-oxide(36508-80-2) CA: 2-pyrrolidinone, 1-methyl-5-(1-oxido-3-pyridinyl)-, (5S)-
5-hydroksykotynina5’-hydroxycotinine(Racemic: 75919-05-0; 5_(R): 61192-50-5)
allohydroxycotinineracemic, CA: 2-pyrrolidinone, 5-hydroxy-1-methyl-5-(3-pyridinyl)- 5’(R), CA: 2-pyrrolidinone, 5-hydroxy-1-methyl-5-(3-pyridinyl)-, (5R)-
4-okso-4-(3-pirydylo)-N-metylobutanamid
4-oxo-4-(3-pyridyl)-N-methylbutanamide(713-05-3)
γ-(3-Pyridyl)-γ-oxo-N-methylbutyramideCA: 3-pyridinebutanamide, N-methyl-γ-oxo-
5’-hydroksynorkotynina 5’-hydroxynorcotinine(118995-82-7)
allohydroxynorcotinineCA: 2-pyrrolidinone, 5-hydroxy-5-(3-pyridinyl)-
4-okso-4-(3-pirydylo)-butanamid 4-oxo-4-(3-pyridyl)-butanamide(61192-49-2)
4-(3-pyridyl)-4-oxobutyramideCA: 3-pyridinebutanamide, γ-oxo-
kwas 4-okso-4-(3-pirydylo)- butanowy
4-oxo-4-(3-pyridyl)butanoic acid(4192-31-8)
γ-(3-pyridyl)-γ-oxobutyric acid4-(3-pyridyl)-4-oxobutyric acidCA: 3-pyridinebutanoic acid, γ-oxo-
kwas 4-hydroksy-4-(3-pirydylo)- butanowy
4-hydroxy-4-(3-pyridyl)butanoic acid(15569-97-8)
γ-(3-pyridyl)-γ-hydroxybutyric acid4-(3-pyridyl)-4-hydroxybutyric acidCA: 3-pyridinebutanoic acid, γ-hydroxy-
jon izometoniowy nikotyny nicotine isomethonium ion(21446-46-8)
N-methylnicotinium ionCA: pyridinium, 1-methyl-3-[(2S)-1-methyl-2-pyrrolidinyl]-, iodide
jon metoniowy kotyniny cotinine methonium ion(33952-07-7)
N-methylcotininium ionCA: pyridinium, 1-methyl-3-(1-methyl-5-oxo-2-pyrrolidinyl)-, iodide, (S)-
N-glukuronid nikotyny nicotine N-glucuronide
N-glukuronid kotyniny cotinine N-glucuronide
O-glukuronid trans-3’-hydroksykotyniny
trans-3’-hydroxycotinine O-glucuronide
a — w nawiasie podano nr katalogowy w rejestrze Chemical Abstractsb — CA, nazwa w indeksie bazy danych Chemical Abstracts
38 www.postepybiochemii.pl
azotu na mający niedobór elektronów atom tlenu związany z hemem. W przypadku nikotyny szlak ten prowadzi do powstania produktu pośredniego — aminiowego rodnika kationowego [23]. Ulega on kolejnemu jednoelektronowe-mu utlenieniu, co prowadzi do odłączenia atomu wodoru od węgla 5’ lub 2’ albo ewentualne tworzenie z grupy N--metylowej odpowiednich kationów iminowych [23]. Po-równanie energii swobodnych aktywacji reakcji ujawniło, że 5’-hydroksylacja jest znacząco faworyzowana względem reakcji N-demetylacji w stosunku 18:1 [28]. Oksydacyjna N--demetylacja jest często istotnym etapem metabolizmu wie-
lu ksenobiotyków. Jednak w przemianach nikotyny u czło-wieka proces ten nie odgrywa znaczącej roli [10].
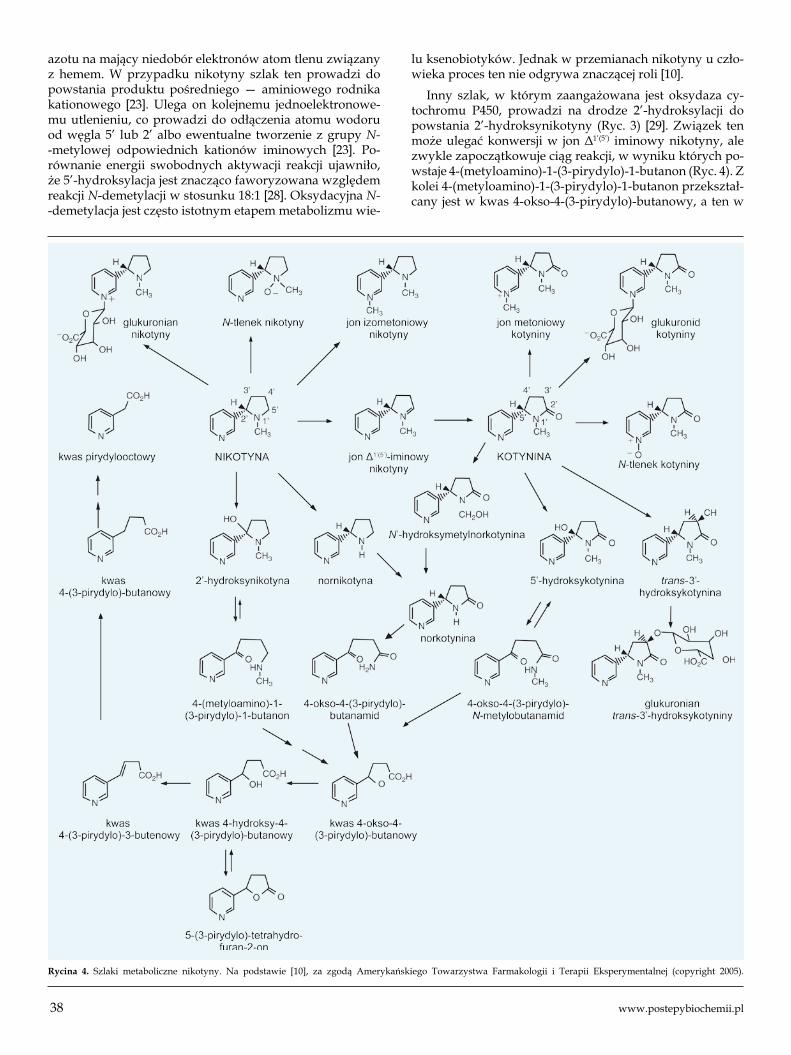
Inny szlak, w którym zaangażowana jest oksydaza cy-tochromu P450, prowadzi na drodze 2’-hydroksylacji do powstania 2’-hydroksynikotyny (Ryc. 3) [29]. Związek ten może ulegać konwersji w jon Δ1’(5’) iminowy nikotyny, ale zwykle zapoczątkowuje ciąg reakcji, w wyniku których po-wstaje 4-(metyloamino)-1-(3-pirydylo)-1-butanon (Ryc. 4). Z kolei 4-(metyloamino)-1-(3-pirydylo)-1-butanon przekształ-cany jest w kwas 4-okso-4-(3-pirydylo)-butanowy, a ten w
Rycina 4. Szlaki metaboliczne nikotyny. Na podstawie [10], za zgodą Amerykańskiego Towarzystwa Farmakologii i Terapii Eksperymentalnej (copyright 2005).

Postępy Biochemii 59 (1) 2013 39
kwas 4-hydroksy-4-(3-pirydylo)-butanowy, pozostający w równowadze z 5-(3-pirydylo)-tetrahydrofuranem-2-on.
U człowieka enzym CYP2A6 katalizuje również po-wstawanie norkotyniny i 5’-hydroksykotyniny z kotyniny (Ryc. 4) [10]. Z 5’-hydroksykotyniny syntetyzowany jest 4-okso-4-(3-pirydylo)-N-metylobutanamid. Jest on praw-dopodobnie prekursorem wielu metabolitów nikotyny, pochodnych powstających w wyniku degradacji pierście-nia pirolidyny. Należą do nich ketokwas 4-okso-4-(3-pi-rydylo)-butanowy i produkty jego redukcji, kolejno kwas 4-hydroxy-4-(3-pirydylo)-butanowy, hydroksykwas, który pozostaje w równowadze z laktonem 5-(3-pirydylo)-tetra-hydrofuranu oraz kwasem 3-pirydylooctowym. Ostatnie z wymienionych związków określa się mianem terminalnych metabolitów nikotyny, które nie podlegają dalszym prze-mianom. Jak wspomniano wcześniej, metabolity te są praw-dopodobnie syntetyzowane jednak przede wszystkim na drodze 2’-hydroksylacji nikotyny (Ryc. 4) [29]. Modelowanie molekularnego oddziaływania nikotyny z centrum aktyw-nym CYP2A6 wskazuje, że utlenianie enzymatyczne niko-tyny przez P450 jest ściśle kontrolowane stereochemiczne. Wiązania wodorowe oraz oddziaływania π-π orientują ni-kotynę względem enzymu w ten sposób, że faworyzowane jest utlenianie alkaloidu w pozycji 5’ (stereo selektywność na poziomie 89-97%) [6]. Alternatywne szlaki metabolizmu nikotyny przy udziale cytochromu P450 wymagają rotacji układu pierścieniowego pirolidyny. Są one mniej korzystne energetycznie niż tworzenie kotyniny przez 5’-utlenianie [23,30]. Najmniejsze bariery energii swobodnej obliczone dla reakcji trans-5’-hydroksylacji oraz cis-5’-hydroksylacji wynoszą odpowiednio 14,1 i 14,4 kcal/mol [6]. Kalkulacje przeprowadzone przez Liu i wsp. [11] wskazują, że w reak-cji tej powstają trans-5’-hydroksynikotyna i cis-3’-hydroksy-nikotyna w stosunku ~36:1. Produkty takich reakcji można zidentyfikować doświadczalnie.
W toku detoksykacji nikotyny przy udziale oksydaz CY-P2A6 i CYP2B6 [31] dochodzi do powstawania nornikotyny, a z niej 4-(3-pirydylo)-4-okso-N-metylobutanamidu. Związ-ki te ulegają dalszym przemianom metabolicznym. Mimo że energetycznie faworyzowana jest reakcja 5’ hydroksylacji, nornikotyna i metylobutanamid jako produkty 2’-utlenia-nia, stają się bezpośrednimi prekursorami rakotwórczych metabolitów nikotyny. Wspomniany wyżej 4-(metyloami-no)-1-(3-pirydylo)-1-butanon może być konwertowany do rakotwórczych NNK (ang. nicotine-derived nitrosamine ketone, 4-(metylonitrozoamino)-1-(3-pirydylo)-1-butanon), natomiast nornikotyna może być prekursorem karcynogen-nych NNN (N’-nitrozonornikotyna), które są uważane za kluczowe w wywoływaniu raka płuc w palaczy [23,29,32].
Oprócz oksydazy cytochromu P450 i oksydazy aldehy-dowej zaangażowany jest cały szereg innych enzymów bio-rących udział w metabolizmie nikotyny (Ryc. 3).
N-tlenek nikotyny jest kolejnym podstawowym meta-bolitem nikotyny, chociaż jedynie 4-7% alkaloidu dostają-cego się do organizmu jest metabolizowane w ten sposób. Reakcję przekształcenia nikotyny do jej N-tlenku katalizuje monooksygenaza flawinowa 3 (FMO3). Produktem reak-cji są dwa diastereoizomery N-tlenku nikotyny, tj. izomer
1’-(R)-2’-(S)-cis oraz izomer 1’-(S)-2’-(S)-trans (Ryc. 3). Szlak ten u ludzi jest bardziej selektywny i w przeważającej części powstaje izomer trans [33]. Wydaje się, że N-tlenek nikoty-ny nie jest dalej metabolizowany. W jelitach może ulegać jednak ponownej redukcji do nikotyny. Obniżenie poziomu N-tlenku u ludzi odbywa się za pośrednictwem aktywności metabolicznej bakterii w jelicie grubym [10].
Poza powyżej opisanymi reakcjami chemicznym pole-gającymi na utlenianiu pierścienia pirolidyny, nikotyna jest metabolizowana poprzez dwa nieoksydacyjne szlaki. Jeden z nich polega na metylacji azotu pirydyny przy udziale en-zymu N-metylotransferazy aminowej, w wyniku czego po-wstaje jon izometoniowy nikotyny. Badania nad szlakami N-metylacji prowadzone z wykorzystaniem modeli zwie-rzęcych i ludzkich, wykorzystujące homogenaty wątroby, wskazały, że S-adenozylo-L-metionina jest źródłem grupy metylowej w reakcji katalizowanej przez N-metylotrans-ferazę aminową [10]. Wyizolowane z wątroby człowieka enzymy cytosolowe przeprowadzały metylację obu enan-cjomerów nikotyny, ale (R)-izomer metylowały szybciej niż (S)-izomer [10].
Drugim sposobem nieoksydacyjnego metabolizmu niko-tyny jest N- i O- glukuronidacja jej pochodnych. Reakcja ta jest katalizowana przez transferazę urydynodwufosfoglu-kuronową (UGT), biorącą udział w koniugacji metabolitów nikotyny z kwasem glukuronowym (Ryc. 3 i 4) [34-36]. W N-glukuronidację zaangażowane są enzymy UGT2B10, UGT2B17, UGT1A4 i częściowo UGT1A9 [37-41]. Powsta-jące produkty wykazują większą rozpuszczalność w po-równaniu do związków macierzystych. W wyniku przy-łączenia reszty cukrowcowej do nikotyny i kotyniny oraz ich pochodnych powstają N-glukuronidy np. glukuronidy nikotyny (3-5%), kotyniny (12-17%) i N-tlenku nikotyny (0,1-0,4%) oraz glukuroniany N-tlenku kotyniny (0,1-0,4%), kwasu 4-hydroksy-4-(3-pirydylo)-butanowego (0,6-0,7%), nornikotyny (0,1%) i norkotyniny (0,1%) [10,37]. Natomiast w przypadku 3’-hydroksykotyniny jej koniugaty to O-glu-kuronidy. Powstawanie O-glukuronianiu trans-3’-hydrok-sykotyniny (7-9%) katalizują enzymy UGT2B7 i częściowo UGT1A9 [14,37,38].
Oba enancjomery nikotyny, z pewnymi tylko wyjątkami, metabolizowane są w tych samych szlakach biochemicznych [23]. Różnice energii niektórych wiązań chemicznych, mają pewien wpływ na zachowanie termodynamiczne enancjo-merów. Tempo metabolizmu (R)-nikotyny jest ok. 1,4 razy szybsze niż (S)-nikotyny. Najprawdopodobniej wynika to z różnicy w energii swobodnej wiązania obu enancjomerów (ok. 1,4 kJ/mol wyższa dla (R)-nikotyny) do cytochromu P450 [6,28]. Na podstawie danych doświadczalnych, teo-retycznie wyznaczono najbardziej stabilną konformację kompleksu substrat-enzym (nikotyna — cytochrom P450), faworyzującą najszybsze powstanie produktu w obrębie grupy 5’-metylenowej. Reorientacja tej konformacji dopro-wadza ostatecznie do powstania innych metabolitów (np. 4-(3-pirydylo)-4-okso-N-metylobutanamidu), wywołane różnicą energii swobodnego wiązania (R)-nikotyny i (S)-ni-kotyny. W rezultacie, w przypadku (R)-nikotyny obserwu-jemy znacznie niższy poziom toksyczności i rakotwórczości powstających metabolitów. Biorąc pod uwagę ogólną cyto-

40 www.postepybiochemii.pl
toksyczność związku i jego metabolitów, wydaje się, że (R)--nikotyna jest około osiemdziesiąt razy mniej cytotoksyczna niż (S)-nikotyna [3,23].
U ludzi stwierdzono występowanie kilku różnych me-tabolitów kotyniny. Należą do nich 3’-hydroksykotynina, 5’-hydroksykotynina (nazywaną także allohydroksykoty-niną, występująca w równowadze ze swoją tautomeryczną pochodną z otwartym pierścieniem 4-okso-4-(3-pirydylo)--N-metylobutanamidem), N-tlenek kotyniny, jon metonio-wy kotyniny oraz, wymienione już wcześniej, norkotynina (nazywaną także demetylokotyniną) i glukuronid kotyniny (Ryc. 4) [14,42]. Norkotynina może powstawać w dwóch szlakach: z kotyniny w wyniku jej demetylacji lub proce-sach metabolizmu oksydacyjnego nornikotyny (Ryc. 4). Ba-dania na zwierzętach wykazały istnienie obu tych ścieżek. W mikrosomach hepatocytów szczurów norkotynina jest dalej metabolizowana do pozostających w równowadze 5’-hydroksynorkotyniny (allohydroksydemetylokotyniny) i 4-okso-4-(3-pirydylo)-butanamidu, który jest następnie przekształcany do kwasu 4-okso-4-(3-pirydylo)-butano-wego. Jego odwodnienie prowadzi do powstania enaminy Δ2’(3’) norkotyniny [10].
U ludzi przekształcenie kotyniny do 3’-hydroksykotyni-ny jest wysoce stereospecyficzne z wyraźną preferencją izo-meru typu trans [43]. W moczu ludzkim wykrywa się mniej niż 5% cis-3’-hydroksykotyniny.
POZAWĄTROBOWY METABOLIZM NIKOTYNY
U zwierząt wykazano, że nikotyna może również być metabolizowana poza wątrobą w nerkach, płucach, mó-zgu i błonach śluzowych nosa [10]. Wykorzystując techniki immunologiczne oraz prowadząc analizę ekspresji mRNA kodującego enzymy zaangażowane w proces metabolizmu nikotyny stwierdzono ich obecność (głównie CYP2A) w na-błonku płuc płodów, w nabłonku oskrzeli, w błonach ślu-zowych nosa, płuc, krtani, przełyku, skóry, naczyń wieńco-wych, gruczołów mlecznych, mózgu, jelitach oraz nerkach [10]. CYP2A13 bierze udział w metabolizmie nikotyny w drogach oddechowych, zwłaszcza w błonach śluzowych nosa, natomiast CYP2B6, CYP2D6 i CYP2E1 najprawdo-podobniej uczestniczą w metabolizmie nikotyny w mózgu [10,24,44]. W hepatocytach oksydaza CYP2E1 uczestniczy również w utlenianiu etanolu [44,45].
Aktywność oksydazy aldehydowej zlokalizowano w płucach, nerkach i nadnerczach [46]. Pozawątrobową ak-tywność FMO3 wykazano w mózgu, zwłaszcza w istocie czarnej [47]. Wysoką aktywność N-metylotransferazy zaob-serwowano u człowieka w tarczycy, nadnerczach i płucach [48]. Aktywność enzymów przeprowadzających reakcje glukuronidacji (UGT1A4) wykryto w jelicie grubym, żołąd-ku i drogach żółciowych [10]. Najwyższy poziom syntezy UGT1A9 (odpowiedzialnej bezpośrednio za proces glu-kuronidacji nikotyny) wykryto w nerkach, jelicie cienkim, wątrobie, żołądku, przełyku, płucach, jądrach, jajnikach i gruczołach piersiowych [10]. Pomimo wielonarządowej re-prezentacji enzymów, potencjalnie mogących konwertować nikotynę, u ludzi metabolizm pozawątrobowy tego alkalo-idu ma marginalne znaczenie w aspekcie ilościowym.
TEMPO METABOLIZMU NIKOTYNY
Obserwuje się znaczną zmienność osobniczą w szybkości eliminacji nikotyny i kotyniny u ludzi [10,14,24]. Zidentyfi-kowano liczne przypadki polimorfizmu genów kodujących enzymy metabolizujące nikotynę i jej pochodne [13,14,24]. Obserwowana duża zmienność osobnicza tempa metaboli-zmu nikotyny [13,14] może być spowodowana przez jeszcze dotąd niezidentyfikowane czynniki genetyczne lub być na-stępstwem obecności dodatkowych substancji tzn. aktywa-torów lub inhibitorów metabolizmu nikotyny.
Jak już wspomniano kluczowym enzymem w metabo-lizmie nikotyny i kotyniny jest zlokalizowana w mikroso-mach hepatocytów oksydaza cytochromu P450 CYP2A6 [49]. U człowieka zidentyfikowano liczne polimorficzne for-my CYP2A6 [21,50,51]. Nomenklaturę ludzkich alleli oksy-dazy cytochromu P450 można prześledzić w odpowiednich bazach danych (http://www.cypalleles.ki.se/). Obecność określonego allozymu CYP2A6 prowadzi do wolniejszego lub szybszego metabolizmu nikotyny [52]. Brak aktywności oksydazy CYP2A6 wynika z obecności allozymów CYP2A6 * 2 i * 5, z kolei występowanie allozymów CYP2A6 * 6, * 7, * 9, * 10, * 11 i * 12 prowadzi jedynie do obniżenia aktyw-ności enzymatycznej. Istnieją jednak sprzeczne opinie na temat różnic w aktywności metabolicznej tych form oraz ich związku z występowaniem raka płuc u palaczy [3,10,24]. W obrębie genów kodujących CYP2B6 zidentyfikowano rów-nież kilka alleli, lecz nie wykazano jak do tej pory ich istot-nego wpływu na zmiany w tempie metabolizmu nikotyny.
Na ogólną aktywność CYP2A6 wpływ mają czynniki działające zarówno na poziomie ekspresji genów kodują-cych oksydazy P450 jak i na etapie regulacji ich aktywno-ści enzymatycznej. Induktory takie jak leki przeciwdrgaw-kowe, ryfampicyna oraz doustne środki antykoncepcyjne, poprzez zwiększenie poziomu ekspresji genu kodującego CYP2A6, przyspieszają metabolizm nikotyny [14,53,54]. Poprzez podniesienie liczby cząsteczek CYP2A6, mogą wy-zwalać zapotrzebowanie na zwiększone przyjmowanie ni-kotyny i utrudniać zerwanie z nałogiem. Inhibitory, obniża-jąc aktywność enzymatyczną CYP2A6, mogą być pomocne w ograniczeniu palenia. Utlenianie nikotyny do kotyniny może być hamowane przez kumaryny, które są kompeten-cyjnym inhibitorem CYP2A6 [10,14]. Mentol jest szeroko stosowany jako środek zapachowy w żywności, do płuka-nia jamy ustnej, zawarty jest w pastach do zębów oraz pa-pierosach. Wykazano, że palenie papierosów mentolowych wydłuża okres półtrwania kotyniny u kobiet. Odnotowano również umiarkowane hamowanie aktywności CYP2A6 w mikrosomach wątroby człowieka przez mentol i związki mu pokrewne [10]. Palenie papierosów zawierających men-tol hamuje metabolizm nikotyny do kotyniny oraz proces glukuronidacji nikotyny w porównaniu z paleniem papie-rosów bez tego dodatku [55]. Sok grejpfrutowy ma silny ha-mujący wpływ na CYP3A4, co często prowadzi do istotnych klinicznie oddziaływań [10]. Hamuje on również CYP2A6, czego dowodem jest spowalnianie metabolizmu kumaryny in vivo [10]. Z kolei kwas askorbinowy zwiększa wydalanie kotyniny i nikotyny [3].
Niewiele jest danych na temat polimorfizmu innych ge-nów zawierających informację dla enzymów metabolizmu

Postępy Biochemii 59 (1) 2013 41
nikotyny. Zaobserwowano go w przypadku genów kodu-jących FMO3 [13,14]. Konsekwencją obecności polimorficz-nych form tej monooksygenazy było obniżenie tempa utle-niania nikotyny [13,14]. Większość polimorfizmów FMO3 występuje stosunkowo rzadko w populacji ludzkiej.
Szybkość metabolizmu nikotyny zależy od przepływu krwi przez wątrobę. Przepływ ten zmienia się w wyniku spożywanych posiłków, wraz z postawą ciała, wykonywa-niem ćwiczeń i pobieraniem leków. Czynnikami wpływa-jącymi na tempo metabolizmu nikotyny u ludzi są także: wiek, płeć, przynależność etniczna oraz stan zdrowia [10,14]. Okres półtrwania nikotyny w organizmie noworodków, narażonych na wdychanie dymu tytoniowego, jest 3 do 4 razy dłuższy niż u osób dorosłych [10,14]. Wykazano, że estrogeny mogą indukować ekspresje genów CYP2A6 [56]. U kobiet, które zażywają doustne środki antykoncepcyjne, tempo metabolizmu nikotyny i kotyniny są odpowiednio o 30% i 33% wyższe, w porównaniu do tych, które nie stosują takich środków. Dodatkowo, w trakcie ciąży, obserwuje się wyraźne przyspieszenie metabolizmu zarówno nikotyny (wzrost o 60%) jak i kotyniny (wzrost o 140%) w porów-naniu z poziomem zmierzonym po porodzie [10,14,57]. U populacji azjatyckich obserwuje się wolniejsze tempo me-tabolizmu nikotyny niż u populacji europejskich. Przynaj-mniej częściowo przyczynę tego zjawiska można upatrywać w występowaniu alleli genu CYP2A6, które fenotypowo ob-jawiają się zmniejszeniem lub brakiem aktywności enzymu [10,24,58,59]. Nie ma istotnych różnic w tempie metabolizmu nikotyny między populacją kaukaską a latynoską, jednak u afroamerykanów metabolizm nikotyny i kotyniny przebie-ga wolniej niż u grup wymienionych wcześniej [10,14,24]. Przewlekłe palenie zmniejsza szybkość metabolizmu niko-tyny, prawdopodobnie przez zmniejszenie ekspresji genu CYP2A6 [13]. Wiele schorzeń wątroby, takich jak zmiany wywołane przez alkohol oraz wirusowe zapalenie wątroby typu A, prowadzi do obniżenia tempa metabolizmu nikoty-ny związanego ze spadkiem aktywności CYP2A6 [24]. Nato-miast infekcja wywołana pasożytniczą motylicą wątrobową wzmaga aktywność CYP2A6 [10,60].
WYDALANIE NIKOTYNY
Ilościowy aspekt wydalanych metabolitów nikotyny zo-stał dość dobrze poznany u ludzi (Ryc. 5). Około 90% przy-jętej przez organizm dawki nikotyny jest wykrywane w moczu. Wykazano, że nikotyna może być usuwana także z kałem (ok. 1-5%), żółcią, śliną, sokiem żołądkowym, potem i mlekiem [3,10]. Od 8 do 10 % nikotyny wydalane jest przez nerki w formie niezmienionej (Ryc. 5). Reszta wchłoniętej przez organizm nikotyny jest w znacznym stopniu zmeta-bolizowana. Czas połowicznej eliminacji nikotyny z organi-zmu człowieka wynosi od 100 min (osoby niepalące) do 150 min (palacze).
W nerkach nikotyna jest wydalana na drodze przesą-czania kłębuszkowego. Stopień wydalania nikotyny zależy od pH moczu oraz jego wydalanej objętości [10,14]. Klirens nikotyny zależny jest w wysokim stopniu od wieku, płci, cech dziedzicznych i stanu zdrowia. Klirens nerkowy dla nikotyny średnio wynosi około 35 do 90 ml/min [10]. Jak już wspomniano, w kwaśnym moczu, nikotyna występuje
głównie w formie zjonizowanej, co minimalizuje zwrotne wchłanianie kanalikowe. Przy pH moczu równym 4,4 kli-rens nerkowy może być bardzo wydajny i może wynosić nawet 600 ml/min, natomiast przy pH 7,0 klirens nerkowy wynosi ok. 17 ml/min, gdyż znaczna część nikotyny wy-stępuje w formie niezjonizowanej, co ułatwia jej zwrotne wchłanianie [10]. Na podstawie badań z jednoczesnym wle-wem nikotyny i kotyniny, udało się ustalić, że chociaż od 70 do 80 % nikotyny u ludzi przekształcane jest do kotyniny, to jedynie 10-15% wchłoniętej przez palaczy nikotyny pojawia się w moczu w postaci niezmienionej kotyniny. Okres pół-trwania kotyniny wynosi od 770 min do 1130 min [10,13,61] i dlatego jest doskonałym wskaźnikiem oceny narażenia na dym tytoniowy u palaczy aktywnych i biernych. Klirens nerkowy kotyniny jest znacznie niższy niż nikotyny. Koty-nina ulega wydajnej resorpcji w kanalikach nerkowych. Na wydalanie kotyniny w znacznie mniejszym stopniu wpływ ma kwasowość moczu, ponieważ w fizjologicznym zakre-sie pH kotynina występuje w formie niezjonizowanej, przez co łatwo jest resorbowana. Kotynina jest wydalana z mo-czem w niewielkim stopniu (10-15% nikotyny). Głównym metabolitem wykrywanym w moczu palaczy jest trans-3’--hydroksykotynina (33-40%, Ryc. 5). Od około 4 do 7% ni-kotyny jest wydalane w postaci N-tlenku nikotyny, a 3-5% jako glukuronid nikotyny. Pozostała część to metabolity wtórne, przede wszystkim glukuronid kotyniny (12-17%) oraz glukuronid trans-3’-hydroksykotyniny (7-9%) (Ryc. 5) [3,10]. Norkotyninę wykryto w moczu palaczy w ilości 1 do 2% całości wchłoniętej nikotyny [10].
Usuwanie nikotyny przez nerki u osób starszych w wie-ku 65-76 lat jest o 23% wolniejsze niż u dorosłych palaczy w młodszym wieku (22-43 lat). Najprawdopodobniej jest to spowodowane wieloma czynnikami, wśród których wy-mienia się, wspominane wcześniej, niższą aktywność me-taboliczną CYP2A6 oraz zmniejszony przepływ krwi przez wątrobę i nerki. Zahamowanie aktywności CYP2A6 lub ob-niżenie ekspresji jej genu w wątrobie może prowadzić do 50% zmniejszenia klirensu nikotyny. Również przewlekła choroba nerek często prowadzi do zmniejszenia klirensu ni-kotyny i kotyniny. U kobiet wydalanie nikotyny i kotyniny przebiega szybciej niż u mężczyzn. Klirens tych związków jest u kobiet odpowiednio wyższy o 13% i 26% [10,14].
PODSUMOWANIE
Wraz z dymem wypalanego papierosa do krwi palacza przedostaje się nikotyna. Roznoszona po całym organizmie dociera do wszystkich narządów. Szczególnie istotną rolę w procesie eliminacji nikotyny pełni wątroba. W trakcie inten-sywnych badań metabolizmu nikotyny w ostatnich kilku dziesięcioleciach dość dobrze poznano szlaki metaboliczne nikotyny oraz enzymy zaangażowane w ten proces. Jed-nak nie wszystkie enzymy uczestniczące w metabolizmie nikotyny zostały do tej pory zidentyfikowane i scharakte-ryzowane. Badania polimorfizmu genetycznego enzymów biorących udział w przemianach nikotyny ujawniły, że ist-nieją duże różnice osobnicze w reakcjach na ten alkaloid, szczególnie w aspekcie ilości przyswajanej nikotyny, popa-dania w uzależnienie oraz indukcji raka płuc [24,44]. Ilość i częstotliwość dostarczania nikotyny do organizmu może wywoływać znaczące zmiany na poziomie regulacji aktyw-

42 www.postepybiochemii.pl
ności kluczowych białek, takich jak receptory nikotynowe w neuronach lub aktywności enzymów detoksykacyjnych w hepatocytach, np. oksydaz cytochromu P450 (CYP2A6). Umiarkowane powinowactwo (R)-nikotyny do choliner-gicznych receptorów typu N w połączeniu ze znacznie niższym poziomem toksyczności powodują, że (R)-niko-tyna może być użytecznym środkiem terapeutycznym dla chorób neurodegeneracyjnych i uzależnienia od palenia tytoniu [23]. Inhibitory CYP2A6 wykorzystywane w tera-pii nikotynozastępczej mogą przyczyniać się do redukcji palenia tytoniu, co w konsekwencji zmniejsza narażenie na rakotwórcze działanie metabolitów nikotyny i być może ograniczać ryzyko zachorowania na raka [10]. W codziennej praktyce lekarskiej istotne byłoby uświadomienie pacjen-tom, że jeśli wypalają oni nawet niewielką liczbę papiero-sów, to ze względu na omówione czynniki, które wpływają na metabolizm nikotyny, szkodliwe działanie tego alkalo-idu może być znaczne, a ta sama dawka nikotyny na każde-go z nas może działać z różną siłą.
PIŚMIENNICTWO1. O’Neil MJ, Smith A, Heckelman PE, Obenchain JR Jr, Gallipeau JAR,
D’Arecca MA, Budavari S (2001) The Merck Index: An Encyclopedia of
Chemicals, Drugs, and Biologicals, 13th ed., Merck Research Laborato-ries Division of Merck & Co., Inc.; Monograph Number: 6551, White-house Station, NJ
2. Modnicki D (2007) Tytoń bakun (Nicotiana rustica L.) niedostrzegane zagrożenie? Przegl Lek 64: 906-907
3. Yildiz D (2004) Nicotine, its metabolism and an overview of its biologi-cal effects. Toxicon 43: 619-632
4. Pogocki D, Ruman T, Danilczuk M, Celuch M, Walajtys-Rode E (2007) Application of nicotine enantiomers, derivatives and analogues in therapy of neurodegenerative disorders. Eur J Pharmacol 563: 18-39
5. Huang X, Zheng F, Crooks PA, Dwoskin LP, Zhan CG (2005) Model-ing multiple species of nicotine and deschloroepibatidine interacting with α4β2 nicotinic acetylcholine receptor: from microscopic binding to phenomenological binding affinity. J Am Chem Soc 127: 14401-14414
6. Li D, Huang X, Han K, Zhan C (2011) Catalytic mechanism of cyto-chrome P450 for 5′-hydroxylation of nicotine: fundamental reaction pathways and stereoselectivity. J Am Chem Soc 133: 7416–7427
7. Nielsen HM, Rassing MR (2002) Nicotine permeability across the buc-cal TR146 cell culture model and porcine buccal mucosa in vitro: effect of pH and concentration. Eur J Pharm Sci 16: 151-157
8. Fukada A, Saito H, Inui K (2002) Transport mechanisms of nicotine across the human intestinal epithelial cell line Caco-2. J Pharmacol Exp Ther 302: 532-538
Rycina 5. Udział najważniejszych metabolitów nikotyny w moczu człowieka. Na podstawie [10], za zgodą Amerykańskiego Towarzystwa Farmakologii i Terapii Ekspe-rymentalnej (copyright 2005).

Postępy Biochemii 59 (1) 2013 43
9. You G, Morris ME, Wang B (2007) Drug transporters: molecular char-acterization and role in drug disposition, John Wiley & Sons, Inc., New Jersey
10. Hukkanen J, Jacob P 3rd, Benowitz NL (2005) Metabolism and disposi-tion kinetics of nicotine. Pharmacol Rev 57: 79-115
11. Wu HJ, Chi CW, Liu TY (2005) Effects of pH on nicotine-induced DNA damage and oxidative stress. J Toxicol Environ Health A 68: 1511-1523
12. Tomar SL, Henningfield JE (1997) Review of the evidence that pH is a determinant of nicotine dosage from oral use of smokeless tobacco. Tob Control 6: 219-225
13. Matta SG, Balfour DJ, Benowitz NL, Boyd RT, Buccafusco JJ, Caggiula AR, Craig CR, Collins AC, Damaj MI, Donny EC, Gardiner PS, Grady SR, Heberlein U, Leonard SS, Levin ED, Lukas RJ, Markou A, Marks MJ, McCallum SE, Parameswaran N, Perkins KA, Picciotto MR, Quik M, Rose JE, Rothenfluh A, Schafer WR, Stolerman IP, Tyndale RF, Wehner JM, Zirger JM (2007) Guidelines on nicotine dose selection for in vivo research. Psychopharmacology (Berl) 190: 269-319
14. Benowitz NL, Hukkanen J, Jacob P 3rd (2009) Nicotine chemistry, me-tabolism, kinetics and biomarkers. Handb Exp Pharmacol 192: 29-60
15. Salomon ME (2006) Nicotine and tobacco preparations, W: Flomen-baum N, Goldfrank L, Hoffman R, Howland MA, Lewin N and Nel-son L (red) Goldfrank’s Toxicologic Emergencies, McGraw-Hill Com-panies, Inc., New York, 1221-1230
16. Shiffman S, Fant RV, Buchhalter AR, Gitchell JG, Henningfield JE (2005) Nicotine delivery systems. Expert Opin Drug Deliv 2: 563-577
17. Seeman JI (2007) Possible role of ammonia on the deposition, reten-tion, and absorption of nicotine in humans while smoking. Chem Res Toxicol 20: 326-343
18. van Amsterdam J, Sleijffers A, van Spiegel P, Blom R, Witte M, van de Kassteele J, Blokland M, Steerenberg P, Opperhuizen A (2011) Effect of ammonia in cigarette tobacco on nicotine absorption in human smok-ers. Food Chem Toxicol 49: 3025-3030
19. McGuigan MA (2004) Nicotine, W: Dart RC (red) Medical Toxicology, Lippincott Williams & Wilkins, Philadelphia, str. 601-604
20. Yerger VB, Malone RE (2006) Melanin and nicotine: A review of the literature. Nicotine Tob Res 8: 487-498
21. Sobkowiak R, Lesicki A (2011) Komórkowe szlaki sygnalizacyjne akty-wowane przez nikotynę. Post Biol Kom 38: 581-596
22. Sobkowiak R, Lesicki A (2009) Genotoxicity of nicotine in cell culture of Caenorhabditis elegans evaluated by the comet assay. Drug Chem Toxicol 32: 252-257
23. Pogocki D (2007) Application of nicotine enantiomers, derivatives and analogues in therapy of neurodegenerative disorders. Eur J Pharmacol 563: 18-39
24. Tricker AR (2003) Nicotine metabolism, human drug metabolism polymorphisms, and smoking behaviour. Toxicology 183: 151-173
25. Tutka P, Mosiewicz J, Wielosz M (2005) Pharmacokinetics and me-tabolism of nicotine. Pharmacol Rep 57: 143-153
26. Niemira M, Wiśniewska A, Mazerska Z (2009) Rola polimorfizmu i zróżnicowanej ekspresji genów cytochromów P450 w metabolizmie ksenobiotyków. Postepy Biochem 55: 279-289
27. von Weymarn L, Retzlaff C, Murphy S (2012) CYP2A6 and CYP2A13-catalyzed metabolism of the nicotine Δ5›(1›)iminium ion. J Pharmacol Exp Ther DOI:10.1124/jpet.112.195255:
28. Li D, Wang Y, Han K, Zhan C (2010) Fundamental reaction pathways for cytochrome P450-catalyzed 5′-hydroxylation and N-demethylation of nicotine. J Phys Chem B 114: 9023-9030
29. Hecht SS, Hochalter JB, Villalta PW, Murphy SE (2000) 2’-Hydrox-ylation of nicotine by cytochrome P450 2A6 and human liver micro-somes: formation of a lung carcinogen precursor. Proc Natl Acad Sci USA 97: 12493-12497
30. Murphy SE, Raulinaitis V, Brown KM (2005) Nicotine 5’-oxidation and methyl oxidation by P450 2A enzymes. Drug Metab Dispos 33: 1166-1173
31. Yamanaka H, Nakajima M, Fukami T, Sakai H, Nakamura A, Katoh M, Takamiya M, Aoki Y, Yokoi T (2005) CYP2A6 and CYP2B6 are involved in nornicotine formation from nicotine in humans: interin-
dividual differences in these contributions. Drug Metab Dispos 33: 1811-1818
32. Schuller HM (2007) Nitrosamines as nicotinic receptor ligands. Life Sci 80: 2274-2280
33. Cashman JR, Park SB, Yang ZC, Wrighton SA, Jacob P 3rd, Benowitz NL (1992) Metabolism of nicotine by human liver microsomes: stere-oselective formation of trans-nicotine N’-oxide. Chem Res Toxicol 5: 639-646
34. Kaivosaari S, Toivonen P, Hesse LM, Koskinen M, Court MH, Finel M (2007) Nicotine glucuronidation and the human UDP-glucuronosyl-transferase UGT2B10. Mol Pharmacol 72: 761-768
35. Kuehl GE, Murphy SE (2003) N-glucuronidation of nicotine and coti-nine by human liver microsomes and heterologously expressed UDP-glucuronosyltransferases. Drug Metab Dispos 31: 1361-1368
36. Fedejko B, Mazerska Z (2011) UDP-glukuronylotranferazy w meta-bolizmie detoksykacyjnym i aktywacyjnym związków endogennych oraz ksenobiotyków. Postepy Biochem 57: 49-62
37. Rangiah K, Hwang WT, Mesaros C, Vachani A, Blair IA (2011) Nico-tine exposure and metabolizer phenotypes from analysis of urinary nicotine and its 15 metabolites by LC-MS. Bioanalysis 3: 745-761
38. Nakajima M, Yokoi T (2005) Interindividual variability in nicotine me-tabolism: C-oxidation and glucuronidation. Drug Metab Pharmacoki-net 20: 227-235
39. Berg JZ, von Weymarn LB, Thompson EA, Wickham KM, Weisensel NA, Hatsukami DK, Murphy SE (2010) UGT2B10 genotype influences nicotine glucuronidation, oxidation, and consumption. Cancer Epide-miol Biomarkers Prev 19: 1423-1431
40. Chen G, Giambrone NE Jr, Dluzen DF, Muscat JE, Berg A, Gallagher CJ, Lazarus P (2010) Glucuronidation genotypes and nicotine meta-bolic phenotypes: importance of functional UGT2B10 and UGT2B17 polymorphisms. Cancer Res 70: 7543-7552
41. Berg JZ, Mason J, Boettcher AJ, Hatsukami DK, Murphy SE (2010) Nicotine metabolism in African Americans and European Americans: variation in glucuronidation by ethnicity and UGT2B10 haplotype. J Pharmacol Exp Ther 332: 202-209
42. Kędziora H, Florek E, Piekoszewski W, Bręborowicz GH, Anholcer A, Gomółka E, Kulza M, Stanek A (2007) Stężenie kotyniny i trans-3’-hy-droksykotyniny oraz ich glukuronidów w moczu kobiet ciężarnych palących tytoń. Przegl Lek 64: 740-746
43. Piekoszewski W, Florek E, Kulza M, Wilimowska J, Loba U (2009) Opracowanie metody oznaczanie metabolitow nikotyny w moczu. Przegl Lek 66: 593-597
44. Wall TL, Schoedel K, Ring HZ, Luczak SE, Katsuyoshi DM, Tyndale RF (2007) Differences in pharmacogenetics of nicotine and alcohol me-tabolism: review and recommendations for future research. Nicotine Tob Res 9 Suppl 3: 459-474
45. Lieber CS (2004) The discovery of the microsomal ethanol oxidizing system and its physiologic and pathologic role. Drug Metab Rev 36: 511-529
46. Moriwaki Y, Yamamoto T, Takahashi S, Tsutsumi Z, Hada T (2001) Widespread cellular distribution of aldehyde oxidase in human tis-sues found by immunohistochemistry staining. Histol Histopathol 16: 745-753
47. Cashman JR, Zhang J (2002) Interindividual differences of human fla-vin-containing monooxygenase 3: genetic polymorphisms and func-tional variation. Drug Metab Dispos 30: 1043-1052
48. Thompson MA, Moon E, Kim UJ, Xu J, Siciliano MJ, Weinshilboum RM (1999) Human indolethylamine N-methyltransferase: cDNA clon-ing and expression, gene cloning, and chromosomal localization. Ge-nomics 61: 285-297
49. Al Koudsi N, Hoffmann EB, Assadzadeh A, Tyndale RF (2010) Hepa-tic CYP2A6 levels and nicotine metabolism: impact of genetic, physio-logical, environmental, and epigenetic factors. Eur J Clin Pharmacol 66: 239-251
50. Benowitz NL, Swan GE, Jacob P 3rd, Lessov-Schlaggar CN, Tyndale RF (2006) CYP2A6 genotype and the metabolism and disposition kine-tics of nicotine. Clin Pharmacol Ther 80: 457-467

44 www.postepybiochemii.pl
51. Mwenifumbo JC, Tyndale RF (2009) Molecular genetics of nicotine metabolism. Handb Exp Pharmacol 2009: 235-259
52. Mwenifumbo JC, Lessov-Schlaggar CN, Zhou Q, Krasnow RE, Swan GE, Benowitz NL, Tyndale RF (2008) Identification of novel CYP2A-6*1B variants: the CYP2A6*1B allele is associated with faster in vivo nicotine metabolism. Clin Pharmacol Ther 83: 115-121
53. Berlin I, Gasior MJ, Moolchan ET (2007) Sex-based and hormonal con-traception effects on the metabolism of nicotine among adolescent to-bacco-dependent smokers. Nicotine Tob Res 9: 493-498
54. Benowitz NL, Lessov-Schlaggar CN, Swan GE, Jacob P 3rd (2006) Fe-male sex and oral contraceptive use accelerate nicotine metabolism. Clin Pharmacol Ther 79: 480-488
55. Benowitz NL, Herrera B, Jacob P, 3rd (2004) Mentholated cigaret-te smoking inhibits nicotine metabolism. J Pharmacol Exp Ther 310: 1208-1215
56. Higashi E, Fukami T, Itoh M, Kyo S, Inoue M, Yokoi T, Nakajima M (2007) Human CYP2A6 is induced by estrogen via estrogen receptor. Drug Metab Dispos 35: 1935-1941
57. Dempsey D, Jacob P 3rd, Benowitz NL (2002) Accelerated metabolism of nicotine and cotinine in pregnant smokers. J Pharmacol Exp Ther 301: 594-598
58. Schoedel KA, Hoffmann EB, Rao Y, Sellers EM, Tyndale RF (2004) Eth-nic variation in CYP2A6 and association of genetically slow nicotine metabolism and smoking in adult Caucasians. Pharmacogenetics 14: 615-626
59. Bloom J, Hinrichs AL, Wang JC, von Weymarn LB, Kharasch ED, Bierut LJ, Goate A, Murphy SE (2011) The contribution of common CYP2A6 alleles to variation in nicotine metabolism among European--Americans. Pharmacogenet Genomics 21: 403-416
60. Satarug S, Lang MA, Yongvanit P, Sithithaworn P, Mairiang E, Mair-iang P, Pelkonen P, Bartsch H, Haswell-Elkins MR (1996) Induction of cytochrome P450 2A6 expression in humans by the carcinogenic para-site infection, opisthorchiasis viverrini. Cancer Epidemiol Biomarkers Prev 5: 795-800
61. Benowitz NL (1996) Cotinine as a biomarker of environmental tobacco smoke exposure. Epidemiol Rev 18: 188-204
Absorption, metabolism and excretion of nicotine in humansRobert Sobkowiak, Andrzej Lesicki
Department of Cell Biology, Institute of Experimental Biology, Adam Mickiewicz University, 89 Umultowska St., 61-614 Poznań, Polande-mail: [email protected]
Key words: nicotine, cotinine, metabolism, absorption, excretion, enantiomer, cytochrome P450
ABSTRACTNicotine is an alkaloid present in many plants of Solanaceae family. The levorotatory enantiomer (S) is a naturally occurring form. Nico-tine enters the human body as a component of tobacco smoke. In alkaline environment the rate of nicotine permeation through biological membranes is increased. Almost 90% of nicotine absorbed by the body is metabolized in the liver. Nicotine may also be metabolized in the kidneys, lungs, brain, and respiratory epithelium membranes. The nicotine undergoes many transformations. Key role in the metabolism of nicotine is played by cytochrome P450 oxidases (mainly CYP2A6). Apart from them, UDP-glucuronosyltransferases, cytosolic aldehyde oxidase, amine N-methyltransferase, and flavin-containing monooxygenase 3 are involved in the decomposition of nicotine. Six major me-tabolites of nicotine have been identified. One of the most important metabolite is cotinine, from which is formed of trans-3’-hydroxycotinine — the compound which is excreted in the largest amount within the urine. The rate of nicotine metabolism is affected by diversified activity of polymorphic enzymes involved in this process, diet, gender and physiological condition of the organism.


















