Regulacja procesu glikozylacji białek przez kaskadę cAMP · Jedną z głównych klas glikomu...
8
Postępy Biochemii 60 (3) 2014 305 Magdalena Surman Marcelina Janik * Zakład Biochemii Glikokoniugatów, Instytut Zoologii, Uniwersytet Jagielloński, Kraków * Zakład Biochemii Glikokoniugatów, Instytut Zoologii, Uniwersytet Jagielloński, ul. Gronostajowa 9, 30-386 Kraków; tel.: (12) 664 64 67, faks: (12) 664 51 01; e-mail: marcelina. [email protected] Artykuł otrzymano 28 lutego 2014 r. Artykuł zaakceptowano 9 czerwca 2014 r. Słowa kluczowe: glikozylacja, kaskada cAMP, syntaza mannozylofosfodolicholu, białka G, cyklaza adenylanowa Wykaz skrótów: AC — cyklaza adenylano- wa; cAMP — cykliczny adenozynomonofos- foran; CMP — cytozynomonofosforan; Dol- -P — fosforan dolicholu; DPMS — syntaza mannozylofosfodolicholu; Gal — galaktoza, GDP — guanozynodifosforan; GEF — czynnik wymiany nukleotydów guaninowych; GlcNAc — N-acetyloglukozoamina; GPCR — recepto- ry sprzężone z białkiem G; GTP — guanozy- notrifosforan; LLO — jednostka oligosachary- dowa związana z nośnikiem lipidowym; Man — mannoza; PKA — kinaza białkowa A; RER — szorstkie retikulum endoplazmatyczne; UDP — urydynodifosforan Regulacja procesu glikozylacji białek przez kaskadę cAMP STRESZCZENIE G likozylacja jest jedną z najczęściej występujących modyfikacji potranslacyjnych. Spo- śród znanych jej form najpowszechniejsze są N- i O-glikozylacja. Inna z potranslacyj- nych modyfikacji białek, fosforylacja jest powszechnym sposobem regulacji aktywności bia- łek enzymatycznych. Regulacji tej podlegają także enzymy prowadzące reakcje na wczesnych etapach procesów N- i O-glikozylacji. Jednym z nich jest syntaza mannozylofosfodolicholu prowadząca reakcję dołączania pierwszej reszty mannozy do fosfodolicholu i tym samym rozpoczynająca syntezę struktury oligosacharydowej w procesie N-glikozylacji. Indukowa- na przez kaskadę cAMP, a realizowaną przez kinazę białek A fosforylacja syntazy manno- zylofosfodolicholu, powoduje wzmacnianie jej aktywności. Innym enzymem wczesnych etapów N-glikozylacji, regulowanym przez cAMP jest cis-prenyltransferaza, katalizująca syntezę fosfodolicholu, a regulacja opiera się o zmiany ekspresji genu tego białka. Wpływ cAMP na przebieg O-glikozylacji opisany został w odniesieniu syntezy struktur O-GlcNAc. Fosforylacja reguluje aktywność syntetyzującej glukozaminę amidotransferazy glutamina fruktozo-6-fosforan i prowadzi do ograniczenia syntezy O-GlcNAc. Obserwacje te sugerują istotną rolę czynników oddziałujących poprzez GPCR na syntezę N- i O-glikanów. WPROWADZENIE Cukry to jedna z największych grup związków organicznych, występujących w każdym żywym organizmie. Pełnią one kluczową rolę w przebiegu większo- ści procesów życiowych, będąc głównym materiałem budulcowym i energetycz- nym. Ich struktura, synteza oraz biologiczna funkcja w stanach fizjologicznych oraz w przebiegu procesów patologicznych są przedmiotem badań glikobiolo- gii. Całość składnika cukrowcowego, zarówno odrębnych cząsteczek węglowo- danów, jak i tych budujących bardziej złożone struktury, określa się mianem gli- komu. W sposób systematyczny jego różnymi aspektami (m. in. genetycznym, fizjologicznym, patologicznym) zajmuje się glikomika, stanowiąca jedną z gałęzi glikobiologii. Jedną z głównych klas glikomu stanowią węglowodany komplek- sowe, czyli glikokoniugaty, które w swej strukturze zawierają co najmniej jeden łańcuch oligosacharydowy. Wszystkie glikokoniugaty są produktami procesu glikozylacji, czyli enzymatycznego tworzenia wiązań kowalencyjnych między łańcuchami oligosacharydowymi a łańcuchami białek (w przypadku glikopro- tein i proteoglikanów) lub lipidów (w glikolipidach). Biosynteza struktur oligosacharydowych, czy to bezpośrednio na łańcuchu polipeptydowym, czy na lipidowym nośniku, odbywa się przy udziale gliko- zylotransferaz, dodających wybrane reszty cukrowe do rozbudowywanej struk- tury oraz glikozydaz, które usuwają określone monosacharydy ze struktury cukrowca. Są to reakcje charakterystyczne dla filogenetycznie szerokiego spek- trum organizmów żywych, począwszy od Archea i Eubacteria, po Eukaryota [1]. W przypadku białek glikozylacja jest najczęstszą potranslacyjną modyfikacją, w wyniku której różnorodne oligosacharydy zostają przyłączone nawet do 50% powstających łańcuchów polipeptydowych [2]. Niewątpliwie stanowi ona jeden z głównych czynników decydujących o chemicznej i funkcjonalnej różnorod- ności białek, i wpływa na ich biologiczną aktywność. Jak stwierdzono, wzory glikozylacji są charakterystyczne dla konkretnych łańcuchów białkowych syn- tetyzowanych w określonych typach komórek bądź w konkretnych tkankach. Obserwacje te sugerują, że w przypadku glikoprotein to nie komponenta biał- kowa, ale właśnie część cukrowa stanowi podstawę złożoności ich struktur, a przez to modyfikuje ich funkcje [3]. Zróżnicowanie glikoprotein wynikać może z modyfikacji, do jakich dochodzi w trakcie syntezy, a które odnoszą się do róż- nych aspektów ich struktury, począwszy od typu i miejsca utworzenia wiązania glikozydowego, poprzez strukturę łańcucha oligosacharydowego (rodzaje reszt cukrowych) i jego długość, skończywszy na obecności lub braku rozgałęzień. Zdolność monomerów budujących oligosacharydy do łączenia się ze sobą na wiele sposobów gwarantuje chemiczną różnorodność. Jej źródłem jest przede
Transcript of Regulacja procesu glikozylacji białek przez kaskadę cAMP · Jedną z głównych klas glikomu...

Postępy Biochemii 60 (3) 2014 305
Magdalena Surman
Marcelina Janik*
Zakład Biochemii Glikokoniugatów, Instytut Zoologii, Uniwersytet Jagielloński, Kraków
*Zakład Biochemii Glikokoniugatów, Instytut Zoologii, Uniwersytet Jagielloński, ul. Gronostajowa 9, 30-386 Kraków; tel.: (12) 664 64 67, faks: (12) 664 51 01; e-mail: [email protected]
Artykuł otrzymano 28 lutego 2014 r.Artykuł zaakceptowano 9 czerwca 2014 r.
Słowa kluczowe: glikozylacja, kaskada cAMP, syntaza mannozylofosfodolicholu, białka G, cyklaza adenylanowa
Wykaz skrótów: AC — cyklaza adenylano-wa; cAMP — cykliczny adenozynomonofos-foran; CMP — cytozynomonofosforan; Dol--P — fosforan dolicholu; DPMS — syntaza mannozylofosfodolicholu; Gal — galaktoza, GDP — guanozynodifosforan; GEF — czynnik wymiany nukleotydów guaninowych; GlcNAc — N-acetyloglukozoamina; GPCR — recepto-ry sprzężone z białkiem G; GTP — guanozy-notrifosforan; LLO — jednostka oligosachary-dowa związana z nośnikiem lipidowym; Man — mannoza; PKA — kinaza białkowa A; RER — szorstkie retikulum endoplazmatyczne; UDP — urydynodifosforan
Regulacja procesu glikozylacji białek przez kaskadę cAMP
STRESZCZENIE
Glikozylacja jest jedną z najczęściej występujących modyfikacji potranslacyjnych. Spo-śród znanych jej form najpowszechniejsze są N- i O-glikozylacja. Inna z potranslacyj-
nych modyfikacji białek, fosforylacja jest powszechnym sposobem regulacji aktywności bia-łek enzymatycznych. Regulacji tej podlegają także enzymy prowadzące reakcje na wczesnych etapach procesów N- i O-glikozylacji. Jednym z nich jest syntaza mannozylofosfodolicholu prowadząca reakcję dołączania pierwszej reszty mannozy do fosfodolicholu i tym samym rozpoczynająca syntezę struktury oligosacharydowej w procesie N-glikozylacji. Indukowa-na przez kaskadę cAMP, a realizowaną przez kinazę białek A fosforylacja syntazy manno-zylofosfodolicholu, powoduje wzmacnianie jej aktywności. Innym enzymem wczesnych etapów N-glikozylacji, regulowanym przez cAMP jest cis-prenyltransferaza, katalizująca syntezę fosfodolicholu, a regulacja opiera się o zmiany ekspresji genu tego białka. Wpływ cAMP na przebieg O-glikozylacji opisany został w odniesieniu syntezy struktur O-GlcNAc. Fosforylacja reguluje aktywność syntetyzującej glukozaminę amidotransferazy glutamina fruktozo-6-fosforan i prowadzi do ograniczenia syntezy O-GlcNAc. Obserwacje te sugerują istotną rolę czynników oddziałujących poprzez GPCR na syntezę N- i O-glikanów.
WPROWADZENIE
Cukry to jedna z największych grup związków organicznych, występujących w każdym żywym organizmie. Pełnią one kluczową rolę w przebiegu większo-ści procesów życiowych, będąc głównym materiałem budulcowym i energetycz-nym. Ich struktura, synteza oraz biologiczna funkcja w stanach fizjologicznych oraz w przebiegu procesów patologicznych są przedmiotem badań glikobiolo-gii. Całość składnika cukrowcowego, zarówno odrębnych cząsteczek węglowo-danów, jak i tych budujących bardziej złożone struktury, określa się mianem gli-komu. W sposób systematyczny jego różnymi aspektami (m. in. genetycznym, fizjologicznym, patologicznym) zajmuje się glikomika, stanowiąca jedną z gałęzi glikobiologii. Jedną z głównych klas glikomu stanowią węglowodany komplek-sowe, czyli glikokoniugaty, które w swej strukturze zawierają co najmniej jeden łańcuch oligosacharydowy. Wszystkie glikokoniugaty są produktami procesu glikozylacji, czyli enzymatycznego tworzenia wiązań kowalencyjnych między łańcuchami oligosacharydowymi a łańcuchami białek (w przypadku glikopro-tein i proteoglikanów) lub lipidów (w glikolipidach).
Biosynteza struktur oligosacharydowych, czy to bezpośrednio na łańcuchu polipeptydowym, czy na lipidowym nośniku, odbywa się przy udziale gliko-zylotransferaz, dodających wybrane reszty cukrowe do rozbudowywanej struk-tury oraz glikozydaz, które usuwają określone monosacharydy ze struktury cukrowca. Są to reakcje charakterystyczne dla filogenetycznie szerokiego spek-trum organizmów żywych, począwszy od Archea i Eubacteria, po Eukaryota [1]. W przypadku białek glikozylacja jest najczęstszą potranslacyjną modyfikacją, w wyniku której różnorodne oligosacharydy zostają przyłączone nawet do 50% powstających łańcuchów polipeptydowych [2]. Niewątpliwie stanowi ona jeden z głównych czynników decydujących o chemicznej i funkcjonalnej różnorod-ności białek, i wpływa na ich biologiczną aktywność. Jak stwierdzono, wzory glikozylacji są charakterystyczne dla konkretnych łańcuchów białkowych syn-tetyzowanych w określonych typach komórek bądź w konkretnych tkankach. Obserwacje te sugerują, że w przypadku glikoprotein to nie komponenta biał-kowa, ale właśnie część cukrowa stanowi podstawę złożoności ich struktur, a przez to modyfikuje ich funkcje [3]. Zróżnicowanie glikoprotein wynikać może z modyfikacji, do jakich dochodzi w trakcie syntezy, a które odnoszą się do róż-nych aspektów ich struktury, począwszy od typu i miejsca utworzenia wiązania glikozydowego, poprzez strukturę łańcucha oligosacharydowego (rodzaje reszt cukrowych) i jego długość, skończywszy na obecności lub braku rozgałęzień. Zdolność monomerów budujących oligosacharydy do łączenia się ze sobą na wiele sposobów gwarantuje chemiczną różnorodność. Jej źródłem jest przede

306 www.postepybiochemii.pl
wszystkim duża liczba naturalnie występujących monosa-charydów i ich izomerów, anomeryczność wiązania gliko-zydowego, tworzenie rozgałęzień, ale także możliwe mody-fikacje reszt cukrowych poprzez przyłączanie grup siarcza-nowych, acetylowych lub fosforanowych [4].
Do syntezy łańcuchów oligosacharydowych w komór-kach eukariotycznych dochodzi w świetle szorstkiej siatecz-ki endoplazmatycznej (RER), a poszczególne reszty cukro-we są usuwane bądź dodawane podczas transportu gliko-proteiny z RER, poprzez aparat Golgiego, na powierzchnię komórki. Obecność reszt cukrowych w strukturze białka zmienia jego właściwości fizyczne, w tym rozmiar, kształt, stopień sfałdowania, rozpuszczalność i ładunek elektrycz-ny. Pod względem biologicznym, białka poddane glikozy-lacji mogą różnić się poziomem sekrecji, aktywnością czy immunogennością. Przykładem wpływu glikozylacji na stabilność cząsteczki są hormony ssaków, będące glikopro-teinami, które najczęściej występują w formie dimerycznej, łańcuchy oligosacharydowe umożliwiają tu połączenie mię-dzy dwiema cząsteczkami białka i zapewniają odporność na proteolizę. Ponadto glikozylacja jest istotna dla mecha-nizmu wzajemnego rozpoznawania się komórek (np. pod-czas tworzenia przerzutów nowotworowych) czy procesu zapłodnienia (częściowo zależnego od wiązania się łańcu-chów oligosacharydowych znajdujących się na powierzchni oocytu i plemnika) [5].
Spośród ok. 200 naturalnie występujących monosachary-dów, zaledwie 8 powszechnie występuje w łańcuchach oli-gosacharydowych glikoprotein [1]. Co istotne w większości reakcji biosyntezy glikoprotein nie biorą udziału wolne, czy ufosforylowane formy cukrów lecz wykorzystywane są odpowiadające im nukleotydocukry. Większość z nich po-wstaje w cytoplazmie z nukleotydów, takich jak urydynodi-fosforan (UDP), guanozynodifosforan (GDP) bądź cytozy-nomonofosforan (CMP). Wyjątkiem są pochodne kwasów sjalowych, które związane z CMP wytwarzane są w jądrze [6]. Zaktywowane cukry zostają włączone w struktury łań-cuchów oligosacharydowych pod warunkiem dostępności swoistych glikozylotransferaz. Większość z tych enzymów wymaga do pełnej aktywności obecności dwuwartościowe-go kationu (głównie jonu Mn2+) i jest wysoce specyficzna względem akceptora, oligosacharydu, cukru nukleotydo-wego oraz rodzaju połączenia cukru z akceptorem [7], i co istotne, większość z nich (poza kilkoma wyjątkami) należy do glikoprotein [8].
Kolejność dodawania reszt cukrowych do tworzonego łańcucha oligosacharydowego, a tym samym jego struktu-ra, zależy również od rozmieszczenia glikozylotransferaz w poszczególnych przedziałach komórkowych. Wybrana reszta cukrowa jest włączana w strukturę oligosacharydu tylko wtedy, gdy znajdzie się on w przedziale, w którym występuje właściwa glikozylotransferaza. Pierwsze resz-ty cukrowe lub prekursorowe łańcuchy oligosacharydowe są przyłączane do łańcuchów polipeptydowych w obrębie RER. Proces ten może zachodzić zarówno w trakcie trans-lacji jak i po jej zakończeniu, a utworzona w jego wyniku cząsteczka podlega dalszym modyfikacjom w aparacie Gol-giego [8]. Rozmieszczenie glikozylotransferaz w jego po-szczególnych przedziałach jest charakterystyczne, enzymy
biorące udział w początkowych etapach syntezy struktur oligosacharydowych przeważają ilościowo w części cis i cy-sternach pośrednich, natomiast enzymy właściwe dla eta-pów późniejszych są zgromadzone w części trans [9].
Wytworzenie wiązania pomiędzy pierwszą resztą cu-krową a aminokwasem jest kluczowe dla biosyntezy łańcu-chów oligosacharydowych w procesie glikozylacji, urucha-mia ono złożoną serię potranslacyjnych reakcji enzymatycz-nych. Dotychczas zidentyfikowano 13 różnych monosacha-rydów (uwzględniając izomery) i 8 reszt aminokwasowych zdolnych do utworzenia 41 różnych wiązań między resztą cukrową a resztą aminokwasową w glikoproteinach. Na podstawie ich szczegółowej struktury wyróżnia się 5 typów glikozylacji: O-glikozylację, N-glikozylację, C-glikozylację, glipiację i fosfoglikozylację [1]. Najpowszechniej występu-jące N- i O-glikoproteiny charakteryzują połączenia oligo-sacharydów z resztami aminokwasowymi w łańcuchu po-lipeptydowym poprzez Asn w przypadku N-oligosachary-dów i Ser/Thr w przypadku O-oligosacharydów.
MECHANIZM SYNTEZY JEDNOSTEK OLIGOSACHARYDOWYCH ZWIĄZANYCH Z NOŚNIKIEM LIPIDOWYM W PRZEBIEGU N-GLIKOZYLACJI
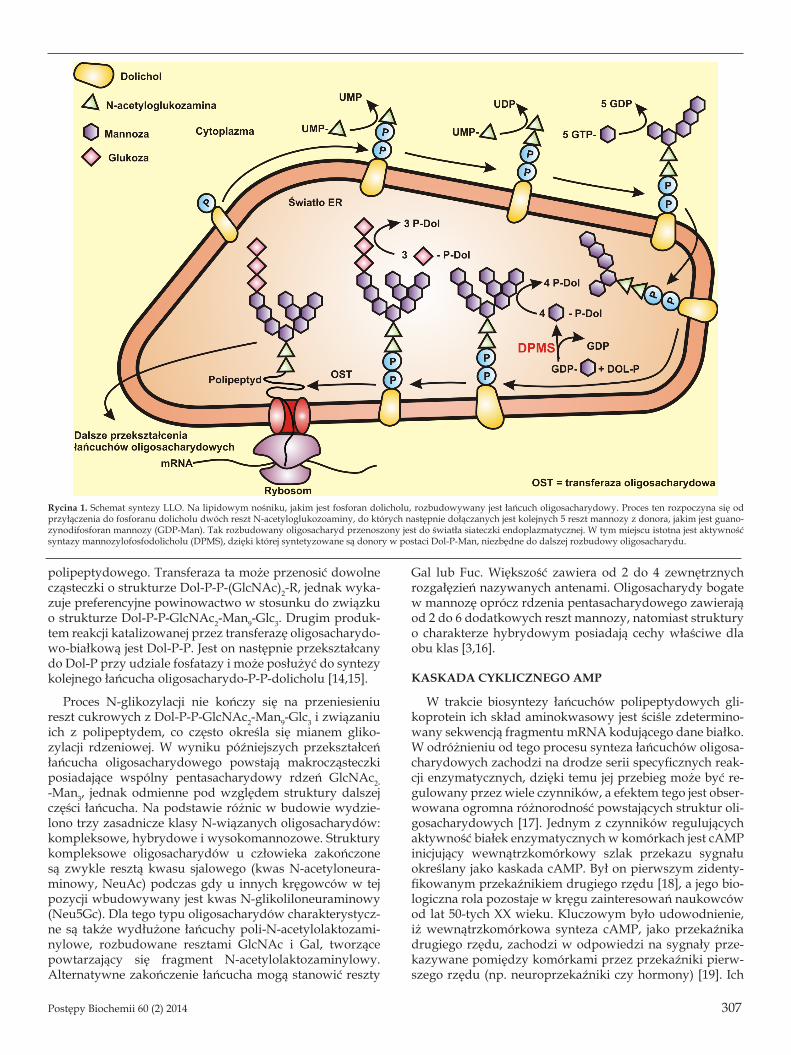
Dotychczas opublikowane wyniki badań nad regulacją procesu glikozylacji białek przez kaskadę cyklicznego ade-nozynomonofosforanu (cAMP), dotyczyły w przeważającej mierze procesu N-glikozylacji. Powstałe w jego wyniku gli-koproteiny posiadają oligosacharydy połączone wiązaniem N--glikozydowym z Asn w łańcuchu polipeptydowym o wspól-nym rdzeniu składającym się z pięciu reszt cukrowych: Man-α(1,3)-Man-α(1,6)-Man-β(1,4)-GlcNAc-β(1,4)-GlcNAc-β1. Typowa struktura rdzenia powstaje w efekcie wspólnego dla wszystkich N-glikanów pierwszego etapu ich biosyn-tezy, który nie wymaga obecności łańcuchów białkowych lecz odbywa się z wykorzystaniem związanego z błoną ER lipidowego nośnika, fosforanu dolicholu (Dol-P). Dol-P stanowi podstawę do syntezy prekursorowej jednostki oli-gosacharydowej związanej z nośnikiem lipidowym (LLO, ang. lipid linked oligosaccharide) (Ryc. 1), który następnie jest przenoszony na atom azotu grupy amidowej Asn w strukturze docelowego białka [10]. Dolichol to jeden z naj-dłuższych, naturalnie występujących alkoholi, zawierający około 17-20 jednostek izoprenowych (C5). W pierwszych reakcjach procesu N-glikozylacji ulega on fosforylacji do Dol-P, kinaza dolicholowa katalizuje reakcję przeniesienia reszty fosforanowej z adenozynotrifosforanu (ATP) na doli-chol. Ufosforylowana forma tworzy wiązanie z łańcuchem oligosacharydowym poprzez końcową grupę fosforanową [11,12]. Reszty cukrowe są przyłączane do Dol-P kolejno od cytosolowej, a następnie od luminarnej strony RER. W wyniku kolejnych reakcji powstaje jednostka oligosacha-rydowa LLO, składająca się z dwóch reszt GlcNAc, dzie-więciu reszt Man i trzech reszt Glc (GlcNAc2-Man9-Glc3). Związany z difosfodolicholem łańcuch oligosacharydowy jest następnie przenoszony w całości na nowo zsyntetyzo-wane białko związane z błoną RER [13]. Tam przy udziale transferazy oligosacharydowo-białkowej, związanej z błoną wewnętrzną RER, wytwarzane jest wiązanie N-glikozydo-we z udziałem atomu azotu grupy amidowej Asn łańcucha
Postępy Biochemii 60 (2) 2014 307
polipeptydowego. Transferaza ta może przenosić dowolne cząsteczki o strukturze Dol-P-P-(GlcNAc)2-R, jednak wyka-zuje preferencyjne powinowactwo w stosunku do związku o strukturze Dol-P-P-GlcNAc2-Man9-Glc3. Drugim produk-tem reakcji katalizowanej przez transferazę oligosacharydo-wo-białkową jest Dol-P-P. Jest on następnie przekształcany do Dol-P przy udziale fosfatazy i może posłużyć do syntezy kolejnego łańcucha oligosacharydo-P-P-dolicholu [14,15].
Proces N-glikozylacji nie kończy się na przeniesieniu reszt cukrowych z Dol-P-P-GlcNAc2-Man9-Glc3 i związaniu ich z polipeptydem, co często określa się mianem gliko-zylacji rdzeniowej. W wyniku późniejszych przekształceń łańcucha oligosacharydowego powstają makrocząsteczki posiadające wspólny pentasacharydowy rdzeń GlcNAc2--Man3, jednak odmienne pod względem struktury dalszej części łańcucha. Na podstawie różnic w budowie wydzie-lono trzy zasadnicze klasy N-wiązanych oligosacharydów: kompleksowe, hybrydowe i wysokomannozowe. Struktury kompleksowe oligosacharydów u człowieka zakończone są zwykle resztą kwasu sjalowego (kwas N-acetyloneura-minowy, NeuAc) podczas gdy u innych kręgowców w tej pozycji wbudowywany jest kwas N-glikoliloneuraminowy (Neu5Gc). Dla tego typu oligosacharydów charakterystycz-ne są także wydłużone łańcuchy poli-N-acetylolaktozami-nylowe, rozbudowane resztami GlcNAc i Gal, tworzące powtarzający się fragment N-acetylolaktozaminylowy.Alternatywne zakończenie łańcucha mogą stanowić reszty
Gal lub Fuc. Większość zawiera od 2 do 4 zewnętrznych rozgałęzień nazywanych antenami. Oligosacharydy bogate w mannozę oprócz rdzenia pentasacharydowego zawierają od 2 do 6 dodatkowych reszt mannozy, natomiast struktury o charakterze hybrydowym posiadają cechy właściwe dla obu klas [3,16].
KASKADA CYKLICZNEGO AMP
W trakcie biosyntezy łańcuchów polipeptydowych gli-koprotein ich skład aminokwasowy jest ściśle zdetermino-wany sekwencją fragmentu mRNA kodującego dane białko. W odróżnieniu od tego procesu synteza łańcuchów oligosa-charydowych zachodzi na drodze serii specyficznych reak-cji enzymatycznych, dzięki temu jej przebieg może być re-gulowany przez wiele czynników, a efektem tego jest obser-wowana ogromna różnorodność powstających struktur oli-gosacharydowych [17]. Jednym z czynników regulujących aktywność białek enzymatycznych w komórkach jest cAMP inicjujący wewnątrzkomórkowy szlak przekazu sygnału określany jako kaskada cAMP. Był on pierwszym zidenty-fikowanym przekaźnikiem drugiego rzędu [18], a jego bio-logiczna rola pozostaje w kręgu zainteresowań naukowców od lat 50-tych XX wieku. Kluczowym było udowodnienie, iż wewnątrzkomórkowa synteza cAMP, jako przekaźnika drugiego rzędu, zachodzi w odpowiedzi na sygnały prze-kazywane pomiędzy komórkami przez przekaźniki pierw-szego rzędu (np. neuroprzekaźniki czy hormony) [19]. Ich
Rycina 1. Schemat syntezy LLO. Na lipidowym nośniku, jakim jest fosforan dolicholu, rozbudowywany jest łańcuch oligosacharydowy. Proces ten rozpoczyna się od przyłączenia do fosforanu dolicholu dwóch reszt N-acetyloglukozoaminy, do których następnie dołączanych jest kolejnych 5 reszt mannozy z donora, jakim jest guano-zynodifosforan mannozy (GDP-Man). Tak rozbudowany oligosacharyd przenoszony jest do światła siateczki endoplazmatycznej. W tym miejscu istotna jest aktywność syntazy mannozylofosfodolicholu (DPMS), dzięki której syntetyzowane są donory w postaci Dol-P-Man, niezbędne do dalszej rozbudowy oligosacharydu.

308 www.postepybiochemii.pl
cząsteczki nie wnikają do komórek, lecz wiążą się ze spe-cyficznymi receptorami obecnymi w błonie komórkowej, których aktywacja prowadzi do cyklizacji ATP. Wewnątrz-cząsteczkowe przejście prowadzące do utworzenia wiąza-nia fosfodiestrowego w cząsteczce ATP z jednoczesnym uwolnieniem pirofosforanu katalizuje cyklaza adenylano-wa (AC), integralne białko błonowe o masie 120 kDa i wielu segmentach transbłonowych [20-23]. Stężenie powstającego cAMP podlega regulacji dzięki silnie egzoergicznej reakcji hydrolizy tej cząsteczki do 5’-AMP katalizowanej przez specyficzną fosfodiesterazę cAMP. W efekcie dochodzi do zahamowania przekazu sygnału wewnątrzkomórkowego [20,24,25].
Kaskada cAMP jest modelowym przykładem systemu sygnalizacji inicjowanego przez receptory sprzężone z biał-kami G (GPCRs, ang. G-protein-coupled receptors). Synteza cAMP następuje zazwyczaj na skutek aktywacji receptorów, które w odpowiedzi na bodziec docierający do komórki, stymulują lub hamują AC za pośrednictwem heterotrime-rycznych białek G (Ryc. 2). Jest to najliczniejsza i najbardziej zróżnicowana pod względem funkcjonalnym grupa recep-torów dla hydrofilowych związków o charakterze przekaź-ników chemicznych. Zalicza się do niej białka receptorowe swoiście wiążące neuroprzekaźniki, neuromodulatory i hormony (receptory adrenergiczne, receptory dopamino-we, receptory serotoninowe, receptory histaminowe, błono-we receptory melatoninowe), a także rodopsynę, receptor dla światła [26-28]. Wszystkie GPCRs zlokalizowane są w błonie komórkowej, a czas od związania agonisty do uzy-skania odpowiedzi układu efektorowego (kanału jonowe-go czy enzymu) w ich przypadku jest rzędu sekund [28]. Wszystkie GPCRs mają ten sam schemat budowy i mecha-nizm działania, łańcuch białka receptorowego siedmiokrot-nie przechodzi przez błonę komórkową, w wyniku czego powstaje siedem α-helikalnych transbłonowych odcinków zbudowanych z 20–25 reszt hydrofobowych aminokwasów, które połączone są pętlami wewnątrz- i zewnątrzkomórko-wymi; N-koniec zlokalizowany jest na zewnątrz komórki, a C-koniec łańcucha polipeptydowego znajduje się w cytozo-lu i zawiera sekwencje mogące ulegać fosforylacji. Miejsce wiązania się liganda z receptorem znajduje się w obrębie hydrofobowej „kieszeni” utworzonej w błonie komórkowej przez transbłonowe α-helisy, natomiast białka G łączą się z receptorem w pobliżu błony komórkowej za pośrednictwem cytoplazmatycznej, najbliższej C-końcowi pętli [28]. Funk-cjonalna wszechstronność systemów sygnalizacji komór-kowej wykorzystujących białko G w roli mediatora, opiera się na jego modułowej strukturze (zbudowane jest z trzech podjednostek: α, β i γ) i istnieniu wielu jego podtypów [29]. Prowadzone w ostatnich dekadach badania wykazały, że powstające w wyniku dysocjacji zaktywowanego białka G podjednostki Gα i Gβγ są w stanie niezależnie wpływać na działanie kilku układów efektorowych, a efekt tego oddzia-ływania jest synergistyczny bądź przeciwstawny [30].
GPCR po związaniu liganda odgrywa rolę czynnika wy-miany nukleotydów guaninowych (GEF, ang. guanine nuc-leotide exchange factor). Po przejściu guanozynodifosforanu (GDP) w guanozynotrifosforan (GTP), heterotrimeryczny kompleks białka G dysocjuje na podjednostki Gβγ i Gα. Kompleks Gαs•GTP aktywuje AC, podczas gdy kompleks
Gαi•GTP jest jej inhibitorem. Ponadto podjednostka Gα posiada aktywność GTPazy, która doprowadza do hydro-lizy GTP do GDP, co hamuje jej oddziaływanie na AC [23]. Mechanizm cyklizacji związanego przez AC ATP do cAMP wymaga ponadto obecności dwóch jonów Mg2+, które wią-żą się z AC oraz z powstałym kompleksem AC i ATP [31].
Cykliczny AMP, jako element wielu szlaków sygnalizacji wewnątrzkomórkowej, bierze udział w regulacji różnych procesów fizjologicznych. Do jego efektorów zalicza się białka Epac 1 i 2 (EPAC, ang. exchange protein directly acti-vated by cAMP), które są białkowymi GEF aktywowanymi przez cAMP, a oddziałującymi przede wszystkim z GTPa-zami Rap 1 i 2 [32]. Efektorami cAMP są również kanały jonowe bramkowane cyklicznymi nukleotydami, które są szczególnie istotne dla funkcjonowania narządów zmysłów (zapachu i smaku). Większość efektów wywoływanych w komórkach eukariotycznych przez cAMP zachodzi jednak w wyniku aktywacji pojedynczej kinazy białkowej, określa-nej jako kinaza białkowa A (PKA). Utworzony przy udziale AC cAMP oddysocjowuje od błony do cytosolu, a gdy jego stężenie osiągnie wartość rzędu 10 nM, aktywuje PKA [33]. PKA zbudowana jest z dwóch rodzajów podjednostek, di-merycznej podjednostki regulatorowej oraz dwóch podjed-nostek katalitycznych. Podjednostki katalityczne pozostają związane z podjednostką regulacyjną, tworząc nieaktywną formę holoenzymu. W odpowiedzi na aktywację AC przez GPCR i wytworzenie drugorzędowego przekaźnika w po-
Rycina 2. Aktywacja kaskady cAMP przez receptor sprzężony z białkiem G (GPCR). GPCR związane z białkiem G, w skład którego wchodzi podjednostka Gsα, po związaniu liganda aktywują cyklazę adenylanową (AC). W efekcie tej aktywacji dochodzi do przekształcenia ATP do formy cyklicznego AMP, które następnie łącząc się z regulatorowymi (R) podjednostkami kinazy białkowej A (PKA), powoduje ich zmiany konformacyjne, co wywołuje oddysocjowanie pod-jednostek katalitycznych (C), umożliwiając im prowadzenie reakcji fosforylacji na wybranych substratach białkowych.

Postępy Biochemii 60 (3) 2014 309
staci cAMP, cztery jego cząsteczki wiążą się z podjednostką regulatorową PKA. Dochodzi wówczas do zmian konfor-macyjnych w obrębie podjednostki regulatorowej PKA, co prowadzi do zmniejszenia jej powinowactwa do podjed-nostek katalitycznych (do 104–105), które oddysocjowując z kompleksu uzyskują zdolność do fosforylacji białkowego substratu [34]. Fosforylacja oznacza przeniesienie przez ki-nazy białkowe końcowej grupy fosforanowej z ATP na resz-tę hydroksylową specyficznego aminokwasu (seryny lub treoniny, w przypadku PKA, bądź tyrozyny). Niezmody-fikowana forma białka może być regenerowana dzięki ak-tywności fosfataz białkowych. Katalizują one reakcje hydro-lizy grup fosforanowych połączonych z białkami, w wyniku których dochodzi do odwrócenia skutków działania kinaz poprzez odtworzenie grupy hydroksylowej łańcucha bocz-nego i powstanie ortofosforanu. W przypadku białek enzy-matycznych fosforylacja przez PKA prowadzi do zmiany ich aktywności [2].
REGULACJA PROCESU GLIKOZYLACJI BIAŁEK PRZEZ KASKADĘ cAMP
Większość dotychczas prowadzonych badań nad regula-cyjnym wpływem kaskady cAMP na przebieg proces gliko-zylacji dotyczyła jednej z pierwszych reakcji zachodzących w trakcie syntezy N-oligosacharydów, syntezy Dol-P-Man, będącego donorem części reszt mannozy podczas syntezy LLO. Reakcję syntezy Dol-P-Man z GDP-Man i Dol-P, któ-ra zachodzi po cytosolowej stronie RER, katalizuje syntaza mannozylofosfodolicholu (DPMS) i to właśnie aktywność tego enzymu podlega regulacji przez kaskadę cAMP [35-39]. Dol-P-Man ulega następnie translokacji do światła RER, gdzie stanowi jedyny donor mannozy w procesie glikozy-lacji, w związku z brakiem transporterów dla GDP-Man. W efekcie DMPS jest silnie zaangażowana w syntezę LLO, któ-ry następnie przenoszony jest na łańcuch polipeptydowy w procesie N-glikozylacji [40].
Badania struktury DMPS ssaków pokazały, że składa się ona z trzech białek: DPM1, DPM2, DPM3. DMP1 odpo-wiada bezpośrednio za katalizowanie reakcji syntezy Dol--P-Man (Ryc. 3). Rolę dwóch pozostałych białek ustalono na podstawie obserwacji linii komórek z zahamowaną ich syntezą (linie komórek z jajnika chomika chińskiego: Lec15 z deficytem DPM2 i CHO2.38 z deficytem DPM3). Wszyst-kie trzy białka wpływają na aktywność katalityczną DPMS. DPM1 wiąże się bezpośrednio z DPM3, a poprzez DPM3
także pośrednio z DPM2. DPM2, tworząc specyficzne wa-runki przestrzenne, ułatwia wiązanie Dol-P, co podnosi wydajność DPMS, natomiast DPM3 stabilizuje kompleks DPMS w błonie RER. Jak zaobserwowano brak DPM3 po-wodował związanie DPM1 z białkiem CHIP, a tak wyzna-kowany kompleks przeznaczony był do degradacji w pro-teasomie [40].
Odpowiedzialne za aktywność katalityczną DPMS biał-ko DPM1 jest kodowane przez gen strukturalny (determi-nujący pierwotną sekwencję aminokwasową tworzonego białka) Dpm1, po raz pierwszy wyizolowany i opisany u Saccharomyces cerevisiae [41]. Na przestrzeni lat odnalezio-no analogiczne sekwencje w genomie przedstawicieli Pro-tozoa, grzybów, a także w genomie mysim i ludzkim. Na podstawie różnic w strukturze, białka DMP1 dzieli się na dwie klasy, z których pierwszą charakteryzuje hydrofobo-wy region na C-końcu łańcucha (u S. cerevisiae, U. maydis, T. brucei, Leishmania mexicana), a druga jest go pozbawiona i dzięki temu pozostaje rozpuszczalna (u S. pombe, C. brigg-siae, Trichoderma reesei i ludzi) [42]. Należące do pierwszej z klas, białko DMP1 jest zdolne do samodzielnego katalizo-wania syntezy Dol-P-Man, podczas gdy DMP1 pozbawione hydrofobowego regionu wymaga obecności dodatkowych elementów (DMP2 i DMP3) umożliwiających zakotwicze-nie jej w błonie ER tak by mogła efektywniej oddziaływać z obecnym tam substratem reakcji, Dol-P. Obecnie za me-chanizm katalityczny charakterystyczny dla DPMS uznaje się inwersję konfiguracji anomerycznej. DPM1, jako kom-ponent katalityczny, wykorzystuje GDP-Man jako donor i Dol-P jako akceptor reszty Man w syntezie Dol-P-Man [40]. Symulacje in silico oraz analizy wykorzystujące zjawisko re-zonansowego przeniesienia energii (FRET, ang. Förster Reso-nance Energy Transfer) pozwoliły stworzyć model struktury 3D DPMS, uwzględniający stopień i sposób sfałdowania białek oraz ukazujący lokalizację miejsca aktywnego i wyja-śniający mechanizm zachodzącej w nim katalizy [42].
Jak pokazano aktywność DPMS, a co za tym idzie do-stępność LLO, jest regulowana poprzez zależny od PKA cykl fosforylacji/defosforylacji. Jest to zjawisko uniwersal-ne, gdyż zostało zaobserwowane u różnych organizmów (od grzybów po ssaki) [35-39,43]. W efekcie ufosforylowa-nia enzymu ilość powstającego produktu była wyższa o 40–80% w przypadku komórek izolowanych ze ślinianek szczura [38], do nawet 40–380% u E. histolityca [43]. Ponie-waż wyższa aktywność katalityczna enzymów może wyni-kać zarówno ze zwiększenia ilości białka syntezowanego de novo jak i aktywacji endogennego enzymu poprzez jego kowalencyjną modyfikację, konieczne było jednoznaczne wykazanie molekularnego mechanizmu odpowiadającego za aktywację DPMS.
Badania aktywności DPMS prowadzono z wy-korzystaniem izoproterenolu, agonisty receptorów β-adrenergicznych, będących typowymi GPCR. W hodo-wanych w obecności izoproterenolu komórkach ślinianek przyusznych szczurów [38] oraz komórkach śródbłonka naczyń włosowatych krowy [39] wykazano, że regulacja N--glikozylacji zachodziła za pośrednictwem kaskady cAMP, której czynnikiem efektorowym była PKA prowadząca fosforylację docelowego enzymu. W efekcie obserwowano
Rycina 3. Struktura syntazy mannozylofosfodolicholu (DPMS). DPMS zbudowa-na jest z 3 podjednostek: DPM1, DPM2, DPM3. Pierwsza z nich posiada aktyw-ność katalityczną, podczas gdy dwie pozostałe odpowiednio: kotwiczą ją w bło-nie siateczki endoplazmatycznej (ER) — DPM3 oraz ułatwiają wiązanie z Dol-P w przebiegu reakcji enzymatycznej — DPM2.

310 www.postepybiochemii.pl
zarówno wzrost szybkości syntezy Dol-P-Man [38], jak i wynikający z niego wzrost ilości tworzonego produktu, bio-rącego później udział w syntezie LLO [39]. Ponadto w przy-padku komórek śródbłonka zaobserwowano wzrost ilości włączanych w trakcie syntezy LLO reszt mannozy (wydłu-żanie Man5GlcNAc2-P-P-Dol do Man9GlcNAc2-P-P-Dol). Analogiczne obserwacje przeprowadzono w tych komór-kach po ich inkubacji w obecności innych związków zwięk-szających wewnątrzkomórkowe stężenie cAMP takich jak: forskolina, toksyna cholery czy prostaglandyna E1 [39]. Otrzymane wyniki sugerują, że w efekcie wzrostu stężenia cAMP i aktywacji kaskady przekazu sygnału prowadzącej do aktywacji PKA następował wzrost syntezy LLO. Anali-za puli N-glikanów w komórkach po stymulacji izoprote-renolem potwierdziła ogólny wzrost ilości tych struktur. Pokazano także na przykładzie czynnika VIIIC, który jest glikoproteiną zaangażowaną w proces angiogenezy i stale obecną w komórkach śródbłonka naczyń włosowatych, iż stymulacja syntezy cAMP przez izoproterenol spowodowa-ła wzrost ilości struktur oligosacharydowych włączonych w strukturę tego konkretnego białka [39]. W celu wyjaśnienia mechanizmu prowadzącego do wzmożonej produkcji N--glikanów w komórkach, w których stymulowana była syn-teza cAMP wykluczono tezę iż wzrost aktywności DPMS jest wynikiem zwiększonej ekspresji genów dla jego pod-jednostek. Po dodaniu aktynomycyny (antybiotyku z gru-py cytostatyków, blokującego syntezę DNA, RNA i białek) zwiększona na skutek działania izoproterenolu aktywność enzymu pozostawała nie zmieniona [44].
Hipotezę iż cząsteczki zdolne do zwiększania wewnątrz-komórkowego stężenia cAMP mogą regulować proces N--glikozylacji białek na drodze fosforylacji/defosforylacji z udziałem PKA potwierdziły również wyniki inkubacji frak-cji mikrosomalnych lub komórek w obecności czynników sprzyjających fosforylacji tj.: ATP oraz podjednostek kata-litycznych PKA. W tym układzie in vitro na skutek fosfory-lacji białek obserwowano zwiększoną aktywność enzyma-tyczną DPMS we frakcjach mikrosomalnych izolowanych z komórek patogennego grzyba Candida albicans [35] oraz z komórek ślinianek przyusznych szczurów [38], a także wzrost aktywności rekombinowanego białka enzymatycz-nego z Saccharomyces cerevisiae [37]. Wykorzystując DMPS wyizolowany z komórek Saccharomyces cerevisiae określo-no również konsensusową sekwencję, która może ulegać fosforylacji, Tyr-Arg-Arg-Val-Ile-Ser. W obrębie cząstecz-ki DPMS pierwotnym miejscem fosforylacji przez PKA w odpowiedzi na sygnalizację indukowaną cAMP jest reszta Ser141, występująca w rejonie o strukturze podobnej do białka DPM1 [37].
Potwierdzeniem zaangażowania PKA w regulację pro-cesu N-glikozylacji były wyniki badań na linii komórek wyizolowanych z jajników samic chomika chińskiego (linia CHO). Do komórek tych wprowadzono trzy różne mutacje hamujące ekspresję genu kodującego PKA, a grupę kontro-lną stanowiły komórki o fenotypie dzikim (z prawidłową ekspresją genu kodującego PKA). W przypadku komórek z mutacjami warunkującymi deficyt PKA zaobserwowano obniżony stopień N-glikozylacji białek, po inkubacji komó-rek w obecności 8Br-cAMP indeks glikozylacji (stosunek włączonej 3H-mannozy do 14C-leucyny w strukturę gliko-
protein) w przypadku komórek o fenotypie dzikim wyniósł 120%, przy zaledwie 7–23% dla komórek z mutacjami. Po-nadto fosforylacja enzymu pochodzącego z komórek z mu-tacjami prowadzona w warunkach in vitro przez egzogenną PKA skutkowała przywróceniem aktywności katalitycznej w stopniu zbliżonym do tego obserwowanego w komór-kach o fenotypie dzikim [36].
Obserwacje związane z wpływem cAMP na N-gliko-zylację prowadzono także na komórkach raka kosmówki (ang. choriocarcinoma) JEG-3 [45,46]. Komórki hodowane w obecności 8-bromo-cAMP wykazywały podwyższony po-ziom syntezy glikoprotein, ale efekt ten obserwowany był tylko przy wydłużonych czasach inkubacji. Sugerowało to, że mechanizmem odpowiedzialnym za ten efekt jest raczej wzmożona produkcja białek enzymatycznych zaangażowa-nych w przebieg procesów N-glikozylacji, a konkretnie w syntezę LLO, a nie ich fosforylacja [45]. Jak pokazano en-zymem, którego aktywność była w tym wypadku regulo-wana przez wewnątrzkomórkowe stężenia cAMP jest cis--prenyltransferaza. Katalizuje ona syntezę Dol-P poprzez przyłączenie 14–17 reszt izopentynylowych do difosforanu-farnezylu. W komórkach JEG-3 stymulowanych 8-bromo--cAMP, wzrastała aktywność tego enzymu, co powodowało zwiększoną syntezę Dol-P, kluczowego substratu pierwsze-go etapu N-glikozylacji. Jak pokazano wzrost aktywności cis-prenyltransferazy, nie wynikał z fosforylacji białka en-zymatycznego, lecz był efektem wzmożonej jego syntezy, wywołanej przez cAMP [46].
Regulacyjny wpływ cAMP został opisany także w sto-sunku do przebiegu O-glikozylacji, a konkretnie do procesu, w wyniku którego białka zostają zmodyfikowane poprzez dołączenie wiązaniem O-glikozydowym N-acetylogluko-zoaminy (O-GlcNAc) do reszt Ser lub Thr w łańcuchu poli-peptydowym [47-49]. Modyfikacji takiej podlegają głównie białka jądrowe oraz czynniki transkrypcyjne, w tym czynnik Sp1 [47] odpowiadający za aktywację genów transkrybowa-nych w każdej komórce [50]. Obserwacje wpływu kaskady cAMP na liczbę struktur O-GlcNAc związanych z białkami prowadzono w hodowlach mysich neuronów móżdżku [48] oraz w linii fibroblastów nerki szczura [47,49]. Jak stwierdzo-no aktywacja kaskady cAMP powodowała ogólne obniże-nie liczby struktur O-GlcNAc związanych z białkami cyto- szkieletu w przypadku neuronów móżdżku myszy [48] oraz prowadziła do częściowej lub całkowitej deglikozylacji białka Sp1 szczurzych fibroblastów [47]. W przypadku fibroblastów nasilenie efektu deglikozylacji obserwowano w hodowlach dodatkowo głodzonych (hodowanych przy braku glukozy). Elementem bezpośrednio regulowanym przez kaskadę była amidotransferaza glutamina:fruktozo-6-fosforan (ang. hu-man glutamine:fructose-6-phosphate amidotransferase, hGFAT) ludzka, enzym prowadzący reakcję syntezy glukozoaminy. Dodatkowo prowadzona przez PKA fosforylacja wyizolowa-nej aktywnej formy enzymu powodowała zahamowanie jego aktywności. Postulowany charakter oddziaływania kaskady cAMP na hGFAT powoduje, że w sytuacji braku dostępnej glukozy głównym szlakiem regulującym jej przemiany jest tor heksozaminowy. W przypadku ograniczonej dostępności glukozy hormony działające poprzez kaskadę cAMP umoż-liwiają wykorzystanie istniejącej glukozy jako źródła energii, ograniczając wykorzystanie jej do syntezy glikoprotein [49].

Postępy Biochemii 60 (3) 2014 311
PODSUMOWANIE
Glikoproteiny pełnią kluczową rolę w przebiegu wielu procesów biologicznych, takich jak adhezja komórkowa, kierowanie białek do odpowiednich przedziałów komórko-wych lub ich sekrecja, aktywacja receptorów, przekaz sy-gnałów czy endocytoza [5]. Co za tym idzie ich znaczenie dla utrzymania stanu fizjologicznego komórek oraz rola w stanach patologicznych jest niepodważalna. Wykazany w różnych układach badawczych, stymulujący wpływ wzro-stu wewnątrzkomórkowego stężenia cAMP na syntezę LLO, realizowany poprzez aktywację syntazy mannozylo-fosfodolicholu na drodze jej fosforylacji zależnej od PKA oraz wywołanych sygnalizacją indukowaną przez cAMP zmian w syntezie cis-prenyltransferazy czy też wpływ fos-forylacji przez PKA na prowadzoną przez hGFAT syntezę glukozoaminy, sugeruje znaczący wpływ czynników hu-moralnych, będących ligandami receptorów sprzężonych z białkami G, na przebieg procesu N-glikozylacji, a także O-glikozylacji. Te obserwacje stanowią istotny krok ku zro-zumieniu roli czynników wpływających na biosyntezę łań-cuchów cukrowych na samym jej początku, a fizjologiczne i medyczne implikacje tego mechanizmu regulacji i jego rola w rozwoju chorób związanych z zaburzeniami glikozylacji powinny stać się przedmiotem przyszłych badań.
Ponieważ dotychczasowe eksperymenty przeprowadza-ne były w większości metodami in vitro, na białkach enzyma-tycznych izolowanych z pojedynczych komórek, nie można jednoznacznie stwierdzić czy aktywacja kaskady cAMP (np. przez podanie agonistów receptorów β-adrenergicznych) w podobnym bądź równym stopniu podnosiłaby lub hamo-wała aktywność enzymu w warunkach in vivo. Niezbędne są zatem dodatkowe badania z wykorzystaniem żywych organizmów, które jednoznacznie potwierdziłyby zało-żenie, że cykl fosforylacji/defosforylacji w znaczący spo-sób reguluje aktywność DPMS i cis-prenyltransferazy (i w efekcie syntezę LLO w procesie N-glikozylacji) [35-46] czy hGFAT [47-48] w warunkach przyżyciowych. Co istotne, wyniki opisanych badań sugerują, że kaskada cAMP może być uniwersalnym mechanizmem regulującym proces gli-kozylacji białek także w innych tkankach organizmów zwierzęcych niż te, z których pochodziły wykorzystane do badań komórki, np. DPMS znajduje się bowiem w komór-kach narządów nawet tak odmiennych, jak jajowód kury i mózg krowy [39]. Warto zaznaczyć, że przedmiotem badań były tylko wybrane enzymy z początkowych etapów proce-sów N- i O-glikozylacji, dlatego potencjalnie interesującym kierunkiem badawczym wydaje się być analiza możliwości regulacji tych procesów na innych etapach (wpływ kaska-dy cAMP na inne glikozylotransferazy). Interesujące byłoby także zbadanie regulacyjnego wpływu kaskady cAMP na przebieg pozostałych, znanych procesów glikozylacji.
PIŚMIENNICTWO1. Spiro RG (2002) Protein glycosylation: Nature, distribution, enzymatic
formation, disease implications of glycopeptide bonds. Glycobiology 12: 43-56
2. Ryšlava H, Doubnerova V, Kavan D, Vanĕk O (2013) Effect of post-translational modifications on enzyme function and assembly. J Pro-teomics 91: 80-109
3. Przybyło M (1998) Budowa i synteza łańcuchów cukrowych glikopro-tein. Kosmos 47: 69-82
4. Mariño K, Bones J, Kattla JJ, Rudd PM (2010) A systematic approach to protein glycosylation analysis: a path through the maze. Nat Chem Biol 6: 713-723
5. Ohtsubo K, Marth JD (2006) Glycosylation in cellular mechanisms of health and disease. Cell 126: 855-867
6. Jimenez del Val I, Kyriakopoulos S, Polizzi KM, Kontoravdi C (2013) An optimized method for extraction and quantification of nucleotides and nucleotide sugars from mammalian cells. Anal Biochem 443: 172-180
7. Breton C, Fournel-Gigleux S, Palcic MM (2012) Recent structures, evo-lution and mechanisms of glycosyltransferases. Curr Opin Struct Biol 22: 540-549
8. Czlapinski JL, Bertozzi CR (2006) Synthetic glycobiology: exploits in the Golgi compartment. Curr Opin Chem Biol 10: 645-651
9. Opat AS, van Vilet C, Gleeson PA (2001) Trafficking and localization of resident Golgi glycosylation enzymes. Biochimie 83: 763-773
10. Banerjee DK (2012) N-glycans in cell survival and death: Cross-talk between glycosyltransferases. Biochim Biophys Acta 1820: 1338-1346
11. Haeuptle MA, Hülsmeier AJ, Hennet T (2009) HPLC and mass spec-trometry analysis of dolichol-phosphates at the cell culture scale. Anal Biochem 396: 133-138
12. Hartley MD, Imperiali B (2012) At the membrane frontier: A prospec-tus on the remarkable evolutionary conservation of polyprenols and polyprenyl-phosphates. Arch Biochem Biophys 517: 83-97
13. Burda P, Aebi M (1999) The dolichol pathway of N-linked glycosyla-tion. Biochim Biophys Acta 1426: 239-257
14. Schwarz F, Aebi M (2011) Mechanism and principles of N-linked pro-tein glycosylation. Curr Opin Struci Biol 21: 576-582
15. Aebi M (2013) N-linked protein glycosylation in the ER. Biochim Bio-phys Acta 1833: 2430-2437
16. Trombetta ES (2003) The contribution of N-glycans and their process-ing in the endoplasmic reticulum to glycoprotein biosyntheis. Glyco-biology 13: 77R-91R
17. Przybyło M (1999) Czynniki wpływające na proces biosyntezy N-gli-kanów w glikoproteinach. Kosmos 48: 95-103
18. Sutherland EW, Rall TW (1958) Fractionation and characterization of cyclic ribonucleotide formed by tissue particles. J Biol Chem 232: 1077-1091
19. Sutherland EW, Robinson GA, Butcher RW (1968) Some Aspects of the Biological Role of Adenosine 3’,5’-monophosphate (Cyclic AMP). Circulation 37: 279-306
20. Antoni FA (2012) New paradigms in cAMP signalling. Mol Cell En-docrinol 353: 3-9
21. Cooper DM (2003) Regulation and organization of adenylyl cyclases and cAMP. Biochem J 375: 517-529
22. Linder JU, Schultz JE (2008) Versatility of signal transduction encoded in dimeric adenylyl cyclases. Curr Opin Struct Biol 18: 667-672
23. Antoni FA (2000) Molecular diversity od cyclic AMP signalling. Front Endocriniol 21: 103-132
24. Bender AT, Beavo JA (2006) Cyclic nucleotide phosphodiesterases: molecular regulation to clinical use. Pharmacol Rev 58: 488-520
25. Essayan DM (2001) Cyclic nucleotide phosphodiesterases. J Allergy Clin Immunol 108: 671-680
26. Heng BC, Aubel D, Fussenegger M (2013) An overview of the diverse roles of G-protein coupled receptors (GPCRs) in the pathophysiology of various human diseases. Biotechnol Adv 31: 1676-1694
27. Ward RJ, Milligan G (2014) Structural and biophysical characterisation of G protein-coupled receptor ligand binding using resonance energy transfer and fluorescent labelling techniques. Biochim Biophys Acta 1838: 3-14
28. Vázquez-Prado J, Casas-González P, García-Sáinz JA (2003) G protein coupled receptor cross-talk: pivotal roles of protein phosphorylation and protein-protein interactions. Cell Signal 15: 549-557
29. Cotton M, Claing A (2009) G protein-coupled receptors stimulation and the control of cell migration. Cell Signal 21: 1045-1053

312 www.postepybiochemii.pl
cAMP cascade in regulation of protein glycosylationMagdalena Surman, Marcelina Janik*
Department of Glycoconjgate Biochemistry, Institute of Zoology, Jagiellonian University, 9 Gronostajowa St., 30-386 Kraków, Poland*e-mail: [email protected]
Key words: glycosylation, cAMP cascade, dolichol phosphate mannose synthase, G proteins, adenylyl cyclase
ABSTRACTO- and N-glycosylation are the most common and complex of the post-translational modifications. Both are enzymatic processes and it was suggested that both could be regulated by cAMP cascade at the early stages. N-glycosylation starts with the formation of lipid-linked oligosac-charides and this process is catalysed by crucial glycosyltransferase — dolichol phosphate mannose synthase. The results of several studies strongly suggest that the cAMP acting through a cAMP-dependent protein kinase A-mediated protein phosphorylation/dephosphorylation cycle may modulate activation of this enzyme. It was shown that cAMP can also up regulate another enzyme involved in phosphodolichole synthesis — cis-prenyltransferase. The mechanism acting here is the alteration of the rate of its gene expression. cAMP cascade is also in-volved in regulation of O-glycosylation since phosphorylation of human glutamine:fructose-6-phosphate amidotransferase results in deple-tion of O-GlcNAc structure formation. These observation suggested an important role of GPCRs and their ligand in regulation of N- and O-glycan synthesis.
30. Wettschureck N, Offermans S (2005) Mammalian G proteins and their cell type specific functions. Physiol Rev 85: 1159-1204
31. Hurley JH (1999) Structure, mechanism, and regulation of mammalian adenylyl cyclases. J Biol Chem 274: 7599-7602
32. Bos JL (2006) Epac proteins: multi-purpose cAMP targets. Trends Bio-chem Sci 31: 680-686
33. Sassone-Corsi P (2012) The Cyclic AMP Pathway. Cold Spring Harb Perspect Biol 4: 1-4
34. Taylor SS, Kim C, Cheng CY, Brown SHJ, Wu J, Kannan N (2008) Sig-naling through cAMP and cAMP-dependent protein kinase: Diverse strategies for drug design. Biochim Biophys Acta 1784: 16-26
35. Arroyo-Flores BL, Calvo-Méndez C, Flores-Carreón A, López-Romero E (2005) Biosynthesis of glycoproteins in the pathogenic fungus Can-dida albicans: Activation of dolichol phosphate mannose synthase by cAMP-mediated protein phosphorylation. FEMS Immunol Med Mi-crobiol 45: 429-434
36. Banerjee DK, Aponte E, Dasilva JJ (2004) Low expression of lip-id-linked oligosaccharide due to a functionally altered Dol-P-Man synthase reduces protein Glycosylation in cAMP-dependent protein kinase deficient Chinase hamster ovary cells. Glycoconj J 21: 479-86
37. Banerjee DK, Carrasquillo EA, Hughey P, Schutzbach JS, Martínez JA, Baksi K (2005) In Vitro Phosphorylation by cAMP-dependent Protein Kinase Up-regulates recombinant Saccharomyces cerevisiae Mannosyl-phosphodolichol Synthase. J Biol Chem 280: 4174-4181
38. Banerjee DK, Kousvelari EE, Baum BJ (1987) cAMP-mediated protein phosphorylation of microsomal membranes increases mannosylphos-podolichol synthase activity. Proc Natl Acad Sci USA 84: 6389-6393
39. Martínez JA, Tavárez JJ, Oliveira CM, Banerjee DK (2006) Potentation of angiogenic switch in capillary endothelial cells by cAMP: A cross-talk between up-regulated LLO biosynthesis and the HSP-70 expres-sion. Glycoconj J 23: 209-220
40. Maeda Y, Kinoshita T (2008) Dolicholphosphate mannose synthase: Structure, function and regulation. Biochim Biophys Acta 1780: 861-868
41. Orlean P, Albright CH, Robbins PW (1988) Cloning and sequencing of the least gene for dolichol phosphate mannose synthase, an essentials protein. J Biol Chem 263: 17499-17507
42. Lamani E, Mewbourne RB, Fletcher DS, Maltsev SD, Danilov LL, Veselovsky VV, Lozanova AV, Grigorieva NY, Pinsker OA, Xing J, Forsee WT, Cheung HC, Schutzbach JS, Shibaev VN, Jedrzejas MJ (2006) Structural studies and mechanism of Saccharomyces cerevisiae dolichol-phosphate-mannose synthase: insights into the initial step of synthesis of dolichylphosphate-linked oligosaccharide chains in mem-branes of endoplasmic retikulum. Glycobiology 16: 666-678
43. Villagómez-Castro JC, Calvo-Méndez C, Flores-Carreón A, López-Romero E (2000) Partial purification and characterization of dolichol phosphate mannose synthase from Entameba histolytica. Gly-cobiology 10: 1311-1316
44. Baksi K, Tavárez-Pagán JJ, Martínez JA, Benerjee DK (2008) Unique structural motif supports mannosylphosphodolichol synthase: an im-portant angiogenesis regulator. Curr Drug Targets 9: 262-71
45. Konrad M, Merz WE (1994) Regulation of N-glycosylation. Long-term effect of cyclic AMP mediates enhances synthesis of the dolichol pyro-phosphate core oligosaccharide. J Biol Chem 269: 8659-8666
46. Konrad M, Merz WE (1996) Long-term effect of cyclic AMP on N-gly-cosylation is caused by an increase in the activity of the cis-prenyl-transferase. Biochem J 316: 575-581
47. Chang Q, Su K, Baker JR, Yang X, Patterson AJ, Kudlow JE (2004) Phosphorylation of human glutamine-fructose-6-phosphate amido-transferase by cAMP-dependent protein kinase at serine 205 blocks the enzyme activity. J Biol Chem 275: 21981-21987
48. Griffith LS, Schmitz B (1999) O-linked N-acetyloglucosamine levels in cerebellar neurons respond reciprocally to perturbations of phoshory-lation. Eur J Biochem 262: 824-831
49. Han I, Kudlow JE (1997) Reduced O-glycosylation of Sp-1 is associated with increased proteasome susceptibility. Mol Cell Biol 17: 2550-2558
50. Czyż M (2001) Sp1 transcription factor. Postepy Biochem 47: 10-8


















