
WYKORZYSTANIE BADAÑ AKTYWNOŒCI...
21
LEŒNE PRACE BADAWCZE, 2005, 3: 17–37. Gra¿yna OLSZOWSKA 1 , Józef ZWOLIÑSKI 1 , Irena MATUSZCZYK 1 , Danuta SYREK 1 , Barbara ZWOLIÑSKA 1 , Urszula PAWLAK 1 , Zygmunt KWAPIS 1 , Ma³gorzata DUDZIÑSKA 2 WYKORZYSTANIE BADAÑ AKTYWNOŒCI BIOLOGICZNEJ DO WYZNACZENIA WSKANIKA ¯YZNOŒCI GLEB W DRZEWOSTANACH SOSNOWYCH NA SIEDLISKACH BORU ŒWIE¯EGO I BORU MIESZANEGO ŒWIE¯EGO THE USE OF BIOLOGICAL ACTIVITY STUDIES TO DETERMINE A SOIL FERTILITY INDICATOR IN PINE STANDS ON FRESH CONIFEROUS AND ON MIXED FRESH CONIFEROUS FOREST SITES Abstract. The relations between the site type, soil chemistry and soil biological activity in Scots pine forests were investigated. The aim of the study was to test the biological activity parameters which could be useful in diagnosing quality of forest sites. To evaluate soil quality of the study sites, several biological and two chemical (sum of base cations and base saturation) parameters were cho- sen to calculate the values of Biological Indicator of Soil Fertility (F). Observed significant correlation between the F values and some of the stand taxation fix- tures associated with stand productivity, i.e. d.b.h., height of stand and stand section, suggests that the F indicator provide a valid estimate of forest site quality. Key words: soil chemistry; soil biological activity; fresh coniferous forest; mixed fresh coniferous forest; Scots pine stands; soil fertility indicator. 1 Instytut Badawczy Leœnictwa, Zak³ad Gospodarki Leœnej Rejonów Przemys³owych, ul. Œw. Huberta 35, 40-952 Katowice; e-mail: [email protected] 2 Instytut Badawczy Leœnictwa, Zak³ad Urz¹dzania i Monitoringu Lasu, ul.Bitwy Warszawskiej 1920 r. 3, 00-973 Warszawa; [email protected]
Transcript of WYKORZYSTANIE BADAÑ AKTYWNOŒCI...

LEŒNE PRACE BADAWCZE, 2005, 3: 17–37.
Gra¿yna OLSZOWSKA1, Józef ZWOLIÑSKI1, Irena MATUSZCZYK1, Danuta SYREK1,Barbara ZWOLIÑSKA1, Urszula PAWLAK1, Zygmunt KWAPIS1, Ma³gorzata DUDZIÑSKA2
WYKORZYSTANIE BADAÑ AKTYWNOŒCI BIOLOGICZNEJDO WYZNACZENIA WSKA�NIKA ¯YZNOŒCI GLEBW DRZEWOSTANACH SOSNOWYCH NA SIEDLISKACH BORUŒWIE¯EGO I BORU MIESZANEGO ŒWIE¯EGO
THE USE OF BIOLOGICAL ACTIVITY STUDIES TO DETERMINEA SOIL FERTILITY INDICATOR IN PINE STANDS ON FRESH CONIFEROUSAND ON MIXED FRESH CONIFEROUS FOREST SITES
Abstract. The relations between the site type, soil chemistry and soil biological
activity in Scots pine forests were investigated. The aim of the study was to test
the biological activity parameters which could be useful in diagnosing quality
of forest sites. To evaluate soil quality of the study sites, several biological and
two chemical (sum of base cations and base saturation) parameters were cho-
sen to calculate the values of Biological Indicator of Soil Fertility (F). Observed
significant correlation between the F values and some of the stand taxation fix-
tures associated with stand productivity, i.e. d.b.h., height of stand and stand
section, suggests that the F indicator provide a valid estimate of forest site quality.
Key words: soil chemistry; soil biological activity; fresh coniferous forest;
mixed fresh coniferous forest; Scots pine stands; soil fertility indicator.
1 Instytut Badawczy Leœnictwa, Zak³ad Gospodarki Leœnej Rejonów Przemys³owych, ul. Œw. Huberta 35,40-952 Katowice; e-mail: [email protected]
2 Instytut Badawczy Leœnictwa, Zak³ad Urz¹dzania i Monitoringu Lasu, ul.Bitwy Warszawskiej 1920 r. 3,00-973 Warszawa; [email protected]

1. WSTÊP
Warunki edaficzne, drobnoustroje glebowe oraz szata roœlinna pozostaj¹ wekosystemach l¹dowych w œcis³ej wspó³zale¿noœci. Tworzenie siê odpowiednichzespo³ów drobnoustrojów glebowych i formacji roœlinnych determinowane jestbowiem w³aœciwoœciami fizycznymi i chemicznymi gleby, które jednoczeœniemodyfikowane s¹ dzia³alnoœci¹ tych organizmów. Poza tym, roœliny dostarczaj¹substraty od¿ywcze dla drobnoustrojów glebowych (w postaci obumar³ych czêœcinadziemnych i korzeni oraz ich wydzielin), przetwarzane przez nie w przyswajalnedla roœlin sk³adniki pokarmowe. Mikrobiologiczne procesy mineralizacji materiiorganicznej gwarantuj¹ utrzymanie niezbêdnego dla rozwoju roœlin zapasu do-stêpnych form sk³adników pokarmowych, st¹d uwa¿a siê, ¿e ich aktywnoœæ œciœlewi¹¿e siê z ¿yznoœci¹ i produktywnoœci¹ ekosystemu (Parkinson 1979, Zak i in.1990). Ponadto, biomasa drobnoustrojów jest magazynem i Ÿród³em pokarmu dlaroœlin, stanowi¹c jeden z g³ównych czynników determinuj¹cych ¿yznoœæ gleb(Jenkinson i Ladd 1981, McGill i in. 1986).
W³aœciwe okreœlenie typu siedliskowego lasu, jego zasobnoœci i potencjalnejzdolnoœci produkcyjnej pozwala na optymalny dobór sk³adu gatunkowego drzew,co wp³ywa na prawid³owy przebieg procesów glebowych, a tym samym zapobiegadegradacji siedlisk. W wielu publikacjach naukowych (Galstjan 1963, Gliñski i in.1983, Hoffmann 1955, Koper i Piotrowska 1999a) wykazano, ¿e badania aktyw-noœci biologicznej gleb mog¹ byæ wykorzystane do oceny ¿yznoœci gleb rolnych.W praktyce leœnej jednak nie znalaz³y one szerszego zastosowania, gdy¿ wiêkszoœæz proponowanych dotychczas wskaŸników biologicznych mo¿e byæ wykorzystanaw ograniczonym zakresie, np. do oceny wp³ywu nawo¿enia, zanieczyszczeñ prze-mys³owych lub sposobu uprawy gleby (Balicka 1986, Koper i Piotrowska 1999b),nie odzwierciedlaj¹ one natomiast stanu siedliska, tj. jego ¿yznoœci i produktyw-noœci. Puchalski i Prusinkiewicz (1990) uwa¿aj¹, ¿e w praktyce leœnej wskaŸ-nikiem jakoœci siedliska jest œrednia wysokoœæ drzewostanu, natomiast Sikorska(1999) podaje, ¿e wskaŸnikiem produkcyjnoœci siedlisk mo¿e byæ bonitacja, bo-wiem im korzystniejsze s¹ warunki siedliskowe, tym drzewostany osi¹gaj¹ wiêksz¹wysokoœæ, a tym samym wy¿sz¹ bonitacjê. Na ¿yznoœæ siedlisk mog¹ tak¿e wska-zywaæ inne cechy taksacyjne drzewostanów, takie jak: przeciêtna pierœnica, prze-ciêtny przekrój, pierœnicowe pole przekroju, mi¹¿szoœæ drzewostanu w korze,mi¹¿szoœæ grubizny drzewostanu, wskaŸnik zadrzewienia.
Celem niniejszej pracy* by³o:1. Okreœlenie intensywnoœci przemian biochemicznych i stanu mikrobiologicz-
nego gleb w drzewostanach sosnowych ró¿nej bonitacji, na siedlisku Bœw i BMœw.2. Ustalenie mo¿liwoœci wykorzystania aktywnoœci biochemicznej jako
wskaŸnika ¿yznoœci gleb oraz w szczegó³owej diagnostyce stanu siedlisk leœnych.
18 G. Olszowska i inni
�Pracê wykonano w ramach tematu: 240509, finansowanego przez Ministerstwo Nauki i Informatyzacji.

W latach 2001–2003 prowadzono analizy chemiczne i pomiary aktywnoœcibiologicznej gleb oraz pomiary dendrometryczne drzewostanów sosnowych nawybranych powierzchniach badawczych, za³o¿onych na siedliskach Bœw i BMœw.Wyniki tych badañ wykorzystano do wyznaczenia wartoœci Biologicznego WskaŸ-nika ¯yznoœci Gleb F, okreœlaj¹cego jakoœæ siedlisk.
2. OPIS TERENU BADAÑ
Powierzchnie reprezentuj¹ce siedliska borowe nizinne wybrano w Nadleœ-nictwie W³oszczowa, RDLP w Radomiu, i w Nadleœnictwie Opoczno, RDLP w£odzi. Oba nadleœnictwa po³o¿one s¹ w Krainie Ma³opolskiej – VI krainieprzyrodniczo-leœnej (Trampler T. i in. 1990). Prace badawcze prowadzono na 21powierzchniach z drzewostanami sosny zwyczajnej (Pinus sylvestris L.) w wieku51–73 lat. Na siedlisku boru œwie¿ego za³o¿ono po trzy powierzchnie w drze-wostanach I, II, III i IV bonitacji, a na siedlisku boru mieszanego œwie¿ego po trzypowierzchnie w drzewostanach sosnowych I, II i III klasy bonitacji.
3. METODYKA BADAÑ
W latach 2001–2003, w paŸdzierniku, z ka¿dej powierzchni badawczej pobra-no objêtoœciowe próby ogólne (z 10 punktów równomiernie rozmieszczonych napowierzchni) z poziomów organicznych (Ofh) i mineralnych (A) gleby do analizchemicznych oraz pomiarów mikrobiologicznych i biochemicznych.
Przed wykonaniem analiz mikrobiologicznych, próby glebowe z poziomówOfh i A przesiewano przez sita o œrednicy oczek odpowiednio 4 mm i 2 mm.
Biomasê drobnoustrojów (Cbiom) oznaczano metod¹ indukowanej substratem(glukoz¹) respiracji (Anderson i Domsch 1978), a intensywnoœæ oddychania gleb,mierz¹c iloœæ uwolnionego C-CO2/g gleby·h. Pomiary uwalnianego CO2, nie-zbêdne do oznaczenia biomasy drobnoustrojów i intensywnoœci oddychania glebwykonano na aparacie Warburga, stosuj¹c nawa¿ki 1 g (z poziomu Ofh) i 5 g (zpoziomu A) gleby doprowadzonej do 60% ca³kowitej pojemnoœci wodnej. Po 48godzinach inkubacji w temp. 22 ºC, do nawa¿ek przeznaczonych do oznaczeñ Cbiom
dodawano po 30 mg (Ofh) i 8 mg (A) glukozy w przeliczeniu na 1 g s.m. gleby, poczym prowadzono cogodzinne odczyty uwolnionego CO2 przez 5 godzin.
Iloraz metaboliczny drobnoustrojów (qCO2), odzwierciedlaj¹cy specyficznetempo respiracji biomasy (qCO2 =�g C-CO2/mg Cbiom·h) obliczano, korzystaj¹c zwyników dotycz¹cych intensywnoœci oddychania gleb i wielkoœci biomasy dro-bnoustrojów (Anderson i Domsch 1992).
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 19

Do oznaczeñ w³aœciwoœci chemicznych oraz aktywnoœci enzymatycznej gleb,powietrznie suche próby glebowe przesiano przez sito o œrednicy 2 mm i oznaczono(Ostrowska i in.1991, Instrukcja laboratoryjna dla pracowni gleboznawczo-na-wo¿eniowych 1973):
– odczyn gleby w 1 M KCl i w H2O, metod¹ potencjometryczn¹,– zawartoœæ azotu ogólnego, metod¹ destylacyjn¹ Kjeldahla,– zawartoœæ fosforu przyswajalnego, metod¹ Egnera-Riehma,– zawartoœæ wêgla, na analizatorze Leco SC132,– zawartoœæ zasadowych kationów wymiennych (Na+, K+, Ca2+ i Mg2+) po
ekstrakcji gleby 1 M octanem amonu, metod¹ absorpcji atomowej,– kwasowoœæ hydrolityczn¹, metod¹ Kappena.Z sumy kationów zasadowych S i kwasowoœci hydrolitycznej Hh obliczono
pojemnoœæ sorpcyjn¹ gleb T. Obliczono równie¿ stopieñ wysycenia kompleksusorpcyjnego zasadami V (%).
Badania enzymatyczne obejmowa³y pomiar aktywnoœci enzymów katalizu-j¹cych rozk³ad wêglowodanów, przemianê zwi¹zków azotowych, uwalnianie fo-sforanów nieorganicznych oraz dehydrogenacjê substancji organicznej:
– ureazy i asparaginazy, metod¹ kolorymetryczn¹, w 1 mg NH3 na 10 g gleby(Ga³stian 1978),
– fosfatazy kwaœnej, metod¹ kolorymetryczn¹, w 1 mg fenolu na 10 g gleby(Russel 1972),
– dehydrogenaz, metod¹ kolorymetryczn¹, w 1 mg trójfenyloformazanu (TF)na 10 g gleby (Galstjan 1978, Russel 1972).
Do badañ dendrometrycznych w drzewostanach za³o¿ono 4-arowe powierzch-nie ko³owe. Liczba powierzchni zale¿a³a od liczby drzew. Starano siê, aby ³¹cznaliczba drzew podlegaj¹cych pomiarowi wynosi³a oko³o 100 (Bruchwald 1995).
Na powierzchniach próbnych zmierzono pierœnice wszystkich drzew œred-nicomierzem precyzyjnym z dok³adnoœci¹ do 1 mm. Dodatkowo zmierzono pierœ-nice i wysokoœci 25 drzew znajduj¹cych siê na powierzchni i poza ni¹ (jednaknale¿¹cych do tego samego wydzielenia drzewostanu) w celu opracowania krzywejwysokoœci.
Na podstawie przeprowadzonych w terenie pomiarów dla ka¿dego drzewo-stanu obliczono wa¿niejsze cechy taksacyjne: przeciêtn¹ pierœnicê D, przeciêtn¹wysokoœæ H, liczbê drzew na 1 ha L/ha, przeciêtny przekrój drzewostanu g, pier-œnicowe pole przekroju na 1 ha (G/ha), mi¹¿szoœæ drzewostanu w korze na ha(Vk/ha) i mi¹¿szoœæ grubizny drzewostanu na ha (Vg/ha), zagêszczenie (Zag) izadrzewienie (Zad).
Przy okreœlaniu mi¹¿szoœci drzewostanu pos³u¿ono siê odpowiednimi wzo-rami empirycznymi:
– pierœnicowej liczby kszta³tu drzewostanu w korze:
FD
D
1 4
1
112895 0 90645
=
++ ⋅
⎛⎝⎜
⎞⎠⎟
, ,
20 G. Olszowska i inni

– grubizny drzew drzewostanu:
FD
D
D
Dg =
++ ⋅
⎛⎝⎜
⎞⎠⎟
⋅ −+ ⋅ −
1
112895 0 90645
6
0 4433 0 98194
, ,
, , ( )6
2⎛
⎝⎜⎜
⎞
⎠⎟⎟
Na podstawie analizy wyników pomiarów biochemicznych i mikrobiolo-gicznych wytypowano wskaŸniki aktywnoœci biologicznej gleby, które wyko-rzystano do obliczenia biologicznego wskaŸnika ¿yznoœci siedlisk leœnych,korzystaj¹c z modelu zaproponowanego przez Myœkowa i in. (1996).
Obliczenia statystyczne przeprowadzono za pomoc¹ programu statystycznegoStatistica 5.0. Do statystycznej oceny wp³ywu siedliska i bonitacji drzew na badaneparametry chemiczne, biologiczne oraz dendrometryczne zastosowano analizêwariancji wieloczynnikowej i test Tukeya. Dla scharakteryzowania zwi¹zku po-miêdzy badanymi parametrami chemicznymi i biochemicznymi gleb a cechamidendrometrycznymi zastosowano analizê korelacyjn¹. Przy testowaniu statystycz-nym badanych parametrów przyjêto poziom istotnoœci p=0,05.
4. WYNIKI
4.1. W³aœciwoœci chemiczne gleb
Wyniki analiz, odzwierciedlaj¹ce w³aœciwoœci chemiczne gleb w ca³ym okre-sie badañ (x z lat 2001–2003), przedstawiono w tabeli 1.
Badane powierzchnie charakteryzowa³y siê nieznacznym zró¿nicowaniempod wzglêdem zawartoœci wêgla organicznego. W poziomach organicznych (Ofh)jego zawartoœæ by³a kilkakrotnie wy¿sza ni¿ w poziomach mineralnych (A), na-tomiast w ca³ej badanej warstwie gleby (Ofh+A) by³a nieznacznie (nieistotniestatystycznie) wy¿sza na siedlisku BMœw (27,17%) ni¿ na siedlisku Bœw (26,16%).Nie stwierdzono ¿adnej zale¿noœci pomiêdzy zawartoœci¹ wêgla organicznego abonitacj¹ drzewostanu.
Podobn¹ tendencjê obserwowano w przypadku zawartoœci azotu w glebie. Wca³ym okresie badañ poziomy organiczne gleb by³y kilkukrotnie bardziej zasobnew ten pierwiastek ni¿ poziomy mineralne. Nie stwierdzono zale¿noœci zawartoœciazotu od typu siedliskowego lasu i bonitacji drzewostanu, a œrednia jego zawartoœæw latach 2001–2003 wynosi³a 0,80% w Bœw i 0,83% w BMœw – ró¿nice sta-tystycznie nieistotne.
Poziom organiczny gleb charakteryzowa³ siê wy¿sz¹ zawartoœci¹ fosforuprzyswajalnego ni¿ poziom mineralny. W ca³ym okresie badawczym zawartoœætego pierwiastka by³a wy¿sza na bogatszym siedlisku BMœw (10,55 mg/kg) ni¿ nasiedlisku Bœw (9,31 mg/kg gleby). Nie stwierdzono zale¿noœci pomiêdzy za-wartoœci¹ fosforu i bonitacj¹ drzewostanu, a wystêpuj¹ce ró¿nice by³y statysty-cznie nieistotne.
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 21

22 G. Olszowska i inni
Tab
ela
1.W
³aœc
iwoœ
cich
emic
zne
górn
ych
pozi
omów
gleb
(Ofh
+A
):œr
edni
az
lat
2001
–200
3T
able
1.C
hem
ical
prop
ertie
sof
uppe
rso
ilho
rizo
ns(O
fh+
A)
(mea
nfr
omye
ars
2001
–200
3)
Typ
sied
-lis
kow
yla
suF
ores
tsite
type
Bon
i-ta
cja
Site
inde
x
Nad
-le
œnic
two
Fore
stD
istr
ict
Leœ
nict
wo
Fore
stra
nge
Odd
zia³
podo
ddzi
a³C
ompa
rtm
ent
pHC %
N %P
2O5
mg/
kg
Kom
plek
sso
rbcy
jny
Sorp
tion
com
plex
cmol
(+)k
gB
S %K
Cl
H2O
Na
KC
aM
gSB
CH
wC
EC
Bœw
Fre
shco
nife
rous
fore
st
IW
³osz
czow
a
Goœ
ciêc
in45
f3,
053,
8730
,06
0,89
6,03
0,07
0,45
6,02
0,56
7,10
82,2
589
,35
7,9
Goœ
ciêc
in46
c3,
083,
8129
,36
0,78
7,63
0,05
0,35
5,34
0,45
6,20
79,3
185
,51
7,2
Pêko
wie
c97
g3,
093,
9129
,19
0,90
7,01
0,06
0,39
5,31
0,50
6,27
81,0
887
,35
7,2
x3,
073,
8729
,53
0,86
6,89
0,06
0,40
5,55
0,50
6,52
80,8
887
,40
7,5
IIO
pocz
no
Ro¿
enek
164
d3,
524,
3220
,14
0,74
13,2
80,
070,
505,
660,
686,
9159
,08
65,9
910
,5R
o¿en
ek16
7a
3,41
4,26
27,6
20,
7812
,58
0,07
0,55
6,30
0,78
7,71
63,4
371
,14
10,8
Ro¿
enek
163
a3,
544,
3526
,22
0,74
13,5
00,
060,
527,
680,
789,
0354
,86
63,8
914
,1x
3,49
4,31
24,6
60,
7513
,12
0,06
0,52
6,55
0,75
7,89
59,1
267
,01
11,8
III
Opo
czno
Myœ
libór
z19
1f
3,12
3,89
24,1
10,
9111
,01
0,07
0,50
5,31
0,64
6,52
80,5
487
,07
7,5
Ro¿
enek
165
a3,
273,
9925
,05
0,68
12,3
30,
070,
574,
160,
645,
4464
,60
70,0
47,
8B
ia³a
czka
60g
2,95
3,73
24,9
20,
845,
760,
080,
403,
790,
454,
7188
,30
93,0
15,
1x
3,11
3,87
24,6
90,
819,
700,
070,
494,
420,
575,
5677
,81
83,3
76,
8
IVO
pocz
no
Kró
lów
ka6
i3,
083,
7725
,84
0,75
7,64
0,08
0,46
3,66
0,46
4,67
72,6
877
,35
6,0
Kró
lów
ka4
f3,
003,
7426
,45
0,91
7,22
0,08
0,47
4,20
0,60
5,34
84,5
989
,93
5,9
Kró
lów
ka5f
3,32
4,03
24,9
30,
717,
670,
060,
403,
950,
514,
9270
,29
75,2
06,
5x
3,13
3,85
25,7
40,
797,
510,
080,
443,
930,
524,
9875
,85
80,8
36,
2
BM
œwM
ixed
fres
hco
nife
rous
fore
st
IW
³osz
czow
a
Wol
aŒw
idz.
110
c3,
454,
2731
,32
0,72
8,70
0,05
0,51
8,06
0,73
9,34
55,7
471
,20
13,1
Mot
yczn
a13
3d
3,29
4,25
24,6
60,
939,
150,
060,
429,
140,
6910
,31
60,8
577
,84
13,2
Kur
zeló
w21
d3,
174,
1530
,48
0,87
7,51
0,06
0,40
7,42
0,71
8,59
65,7
776
,52
11,2
x3,
304,
2228
,82
0,84
8,46
0,05
0,44
8,21
0,71
9,41
60,7
975
,19
12,5
IIO
pocz
no
Ro¿
enek
164
j3,
514,
3517
,88
0,83
12,5
90,
050,
588,
490,
8710
,00
59,2
549
,69
20,1
Ro¿
enek
166
d3,
184,
0322
,22
0,88
8,60
0,08
0,60
5,91
0,93
7,52
74,5
158
,45
12,9
Ro¿
enek
169
i3,
204,
0430
,53
0,83
9,95
0,06
0,52
5,80
0,88
7,26
61,0
651
,24
14,2
x3,
304,
1423
,54
0,84
10,3
80,
060,
576,
730,
898,
2664
,94
53,1
315
,7
III
Opo
czno
Myœ
libór
z20
2a
3,74
4,61
27,9
30,
6917
,78
0,03
0,44
7,87
0,98
9,32
41,8
443
,56
21,4
Myœ
libór
z21
2b
3,32
4,14
32,6
60,
8512
,08
0,06
0,69
9,71
1,29
11,7
448
,34
55,7
821
,1M
yœlib
órz
204
c2,
953,
8126
,83
0,89
8,60
0,08
0,56
5,75
0,54
6,92
88,1
366
,80
10,4
x3,
344,
1929
,14
0,81
12,8
20,
060,
567,
770,
949,
3359
,44
55,3
817
,6

Na wszystkich powierzchniach suma kationów zasadowych S by³a zdecy-dowanie wy¿sza w poziomie organicznym (Ofh) ni¿ w poziomie mineralnym (A), aw ca³ym okresie badawczym by³a wiêksza na siedlisku BMœw ni¿ na siedliskuBœw. Wykazywa³a ona tendencjê spadkow¹ wraz ze spadkiem bonitacji – z 6,52cmol(+)/kg w I klasie do 4,98 cmol(+)/kg w IV klasie w Bœw oraz z 9,41cmol(+)/kg w I klasie do 8,26 cmol(+)/kg w II klasie i 9,33 cmol(+)/kg w III klasiew BMœw. Obserwowane ró¿nice by³y statystycznie istotne (F=38,8; p<0,001). Zkolei, gleby na siedlisku Bœw charakteryzowa³y siê statystycznie istotnie wy¿sz¹(F=10,65; p<0,002) kwasowoœci¹ hydrolityczn¹ Hh ni¿ gleby na siedlisku BMœw,tj. odpowiednio 73,42 cmol(+)/kg i 61,72 cmol(+)/kg. Podobn¹ tendencjê obser-wowano w przypadku pojemnoœci kompleksu sorpcyjnego gleb T, która by³aistotnie wy¿sza (F=19,69; p<0,001) na siedlisku Bœw (79,65 cmol(+)/kg) ni¿BMœw (61,23 cmol(+)/kg). Wysycenie kompleksu sorpcyjnego zasadami (V) by³onatomiast istotnie wy¿sze (F=10,9; p<0,001) na siedlisku BMœw (15,28%) ni¿ nasiedlisku Bœw (8,06%).
Na podstawie pomiarów pH prób glebowych okreœlono odczyn gleb, który nawszystkich powierzchniach, niezale¿nie od typu siedliska, jest silnie kwaœny, przyczym poziom Ofh charakteryzowa³ siê ni¿szym pH ni¿ poziom A. Nie stwierdzonostatystycznie istotnych zale¿noœci pomiêdzy ¿yznoœci¹ siedliska i bonitacj¹ a od-czynem badanych gleb. W latach 2001–2003 pH w KCl w poziomie organiczno-mineralnym (Ofh-A) wynios³o œrednio 3,20, a pH w H2O – 3,97 w Bœw i od-powiednio 3,31 i 4,18 w BMœw.
4.2. Stan mikrobiologiczny gleb
Pomiary mikrobiologiczne prowadzono w górnych warstwach gleb (poziomyOfh i A), poniewa¿ reprezentuj¹ one warstwê najwiêkszej aktywnoœci drobno-ustrojów i stanowi¹ g³ówny rezerwuar sk³adników pokarmowych dla roœlin nasiedliskach borowych. Ze wzglêdu na du¿e zró¿nicowanie gleb pod wzglêdemzawartoœci substancji organicznej oraz mi¹¿szoœci poszczególnych poziomów,wyniki oznaczeñ intensywnoœci oddychania gleb (tab. 2) oraz biomasy drobno-ustrojów glebowych (tab. 3) przedstawiono w przeliczeniu na jednostkê powierz-chni (jedn./ha), co uwa¿a siê za bardziej miarodajne przy ocenie stanu mikro-biologicznego gleb ni¿ odniesienie do jednostek wagowych gleby (Federer i in.1993, Aikio i in. 2000).
Na wszystkich powierzchniach intensywnoœæ oddychania gleb w poziomachorganicznych (Ofh) by³a kilkakrotnie wy¿sza ni¿ w poziomach mineralnych (A).Zawieraj¹ one bowiem znacznie wiêcej wêgla organicznego, stanowi¹cego substratwykorzystywany przez drobnoustroje w procesach mineralizacji. Iloœæ wydzie-lonego CO2 z ca³ej badanej warstwy gleby, odzwierciedlaj¹ca potencjaln¹ in-tensywnoœæ mineralizacji wêgla, by³a zró¿nicowana na poszczególnych powierz-chniach. Aktywnoœæ tego procesu (x z lat 2001–2003) by³a jednak wyraŸniewy¿sza na bogatszym siedlisku BMœw (728–846 g C-CO2/ha�h) ni¿ na siedlisku
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 23

24 G. Olszowska i inni
Tab
ela
2.In
tens
ywno
Ͼod
dych
ania
gleb
(gC
-CO
2/ha
·h)
wpo
ziom
ach
orga
nicz
nych
(Ofh
)im
iner
alny
ch(A
)gl
ebT
able
2.So
ilre
spir
atio
n(g
C-C
O2/
ha·h
)in
orga
nic
(Ofh
)an
dm
iner
al(A
)so
ilho
rizo
ns
Sied
lisko
Site
Bon
itac
jaSi
tein
dex
Nr
pow
.P
lotn
o.
Nad
leœn
ictw
oFo
rest
Dis
tric
tL
eœni
ctw
oFo
rest
rang
e
Odd
zia³
Com
part
-m
ent
2001
r.20
02r.
2003
r.20
01–2
003
Ofh
AO
fh+
AO
fhA
Ofh
+A
Ofh
AO
fh+
AO
fh+
A
Bœw
Fres
hco
nife
rous
fore
st
I
1
W³o
szcz
owa
Goœ
ciêc
in45
f52
410
763
171
115
086
157
314
772
0
717
2G
oœci
êcin
46c
598
149
747
686
9878
456
619
375
93
Pêko
wie
c97
g53
314
267
537
316
153
461
312
774
0
x55
213
368
559
013
672
658
415
674
0
II
4
Opo
czno
Ro¿
enek
164d
345
170
515
186
220
406
513
111
624
579
5R
o¿en
ek16
7a52
812
765
550
618
769
358
413
371
76
Ro¿
enek
163a
242
125
367
514
241
755
351
124
475
x37
214
151
3140
221
661
848
312
360
6
III
7
Opo
czno
Myœ
libór
z19
1f47
319
166
542
415
658
056
992
661
679
8R
o¿en
ek16
5a46
612
459
043
619
162
756
515
572
09
Bia
³acz
ów60
g59
673
669
824
102
926
522
149
671
x51
212
964
156
115
071
155
213
268
4
IV
10
Opo
czno
Kró
lów
ka6i
565
261
826
727
150
877
514
154
668
701
11K
róló
wka
4f40
118
158
229
613
443
060
319
980
212
Kró
lów
ka5f
386
165
551
637
159
796
661
113
774
x45
120
265
355
314
870
159
315
574
8
BM
œwM
ixed
fres
hco
nife
rous
fore
st
I
13
W³o
szcz
owa
Wol
aŒw
idz.
110c
403
213
616
619
103
722
637
194
831
728
14M
otyc
zna
133d
425
265
690
557
107
664
759
177
936
15K
urze
lów
21d
417
184
601
669
109
778
596
114
710
x41
522
163
661
510
672
166
416
282
6
II
16
Opo
czno
Ro¿
enek
164j
408
103
511
748
130
878
832
316
1148
846
17R
o¿en
ek16
6d46
518
464
912
7516
614
4187
610
698
218
Ro¿
enek
169i
450
167
617
387
138
525
748
119
867
x44
115
159
280
314
594
881
918
099
9
III
19
Opo
czno
Myœ
libór
z20
2a46
828
675
436
613
450
056
313
269
5
834
20M
yœlib
órz
212b
751
307
1058
813
210
1023
719
268
987
21M
yœlib
órz
204c
678
236
914
419
150
565
858
9395
1
x63
227
690
853
316
569
671
316
489
7

Bœw (579–717 g C-CO2/ha�h). Nie stwierdzono natomiast ¿adnej zale¿noœci po-miêdzy intensywnoœci¹ oddychania gleb a bonitacj¹ drzewostanu.
Podobnie jak w przypadku oddychania gleb, poziomy organiczne gleb (bar-dziej zasobne w substraty od¿ywcze), charakteryzowa³y siê znacznie wiêksz¹biomas¹ drobnoustrojów ani¿eli poziomy mineralne. W ci¹gu ca³ego okresu badañbiomasa drobnoustrojów glebowych by³a wiêksza na siedlisku BMœw ni¿ w Bœw ina obu siedliskach mia³a tendencjê spadkow¹ wraz ze spadkiem bonitacji w Bœw z172 kg Cbiom/ha w I klasie do 121 kg Cbiom/ha w IV klasie, a w BMœw z 195 kgCbiom/ha w I klasie do 146 kg Cbiom/ha w III klasie bonitacji (tab. 3). Powy¿szaobserwacja œwiadczy o wyraŸnym zwi¹zku miêdzy biomas¹ drobnoustrojów ajakoœci¹ siedlisk – maj¹c¹ istotny wp³yw na produkcyjnoœæ drzewostanów.
Jakoœæ siedliska mia³a wyraŸny wp³yw na kszta³towanie siê ilorazu meta-bolicznego (qCO2) drobnoustrojów, odzwierciedlaj¹cego specyficzne tempo re-spiracji biomasy (tab. 3). Œrednia wa¿ona wartoœci qCO2 dla badanych poziomówgleb (Ofh i A) w latach 2001–2003 wzrasta³a wraz z pogarszaniem siê jakoœcisiedlisk: w Bœw z 3,44 w I klasie do 4,40 �g C-CO2/mg Cbiom�h w IV klasie, a wBMœw – z 2,80 w I klasie do 3,79 �g C-CO2/mg Cbiom�h w III klasie bonitacji.Oznaczenia qCO2 s¹ czêsto stosowane przy ocenie efektywnoœci drobnoustrojów wwykorzystywaniu zawartych w glebie substratów od¿ywczych (Anderson i Do-msch 1992). Ni¿sza wartoœæ qCO2 oznacza, ¿e drobnoustroje w wiêkszym stopniuwykorzystuj¹ energiê do biosyntezy ni¿ do procesów katabolicznych (respiracji).Efektem tego jest wiêkszy wzrost biomasy drobnoustrojów i w rezultacie wiêkszyzapas sk³adników pokarmowych w glebie.
4.3. AktywnoϾ enzymatyczna gleb
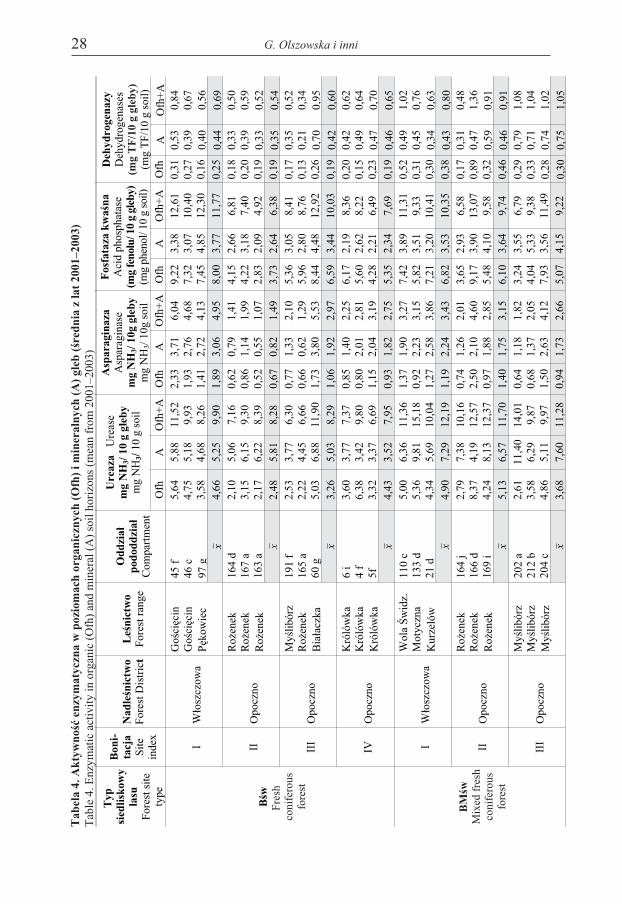
Wykonane w latach 2001–2003 badania wykaza³y, ¿e aktywnoœæ wszystkichbadanych enzymów podlega³a wahaniom sezonowym, wynikaj¹cym prawdopo-dobnie ze zmiennych warunków atmosferycznych oraz iloœci dop³ywaj¹cej do glebmaterii organicznej (ryc. 1). Stwierdzono ponadto, ¿e aktywnoœæ enzymów gle-bowych by³a œciœle zwi¹zana z zawartoœci¹ substancji organicznej: by³a wy¿sza wpoziomie organicznym (Ofh) ni¿ w poziomie mineralnym (A) (tab. 4).
Aktywnoœæ ureazy, enzymu katalizuj¹cego przemianê zwi¹zków azotowych wglebie, by³a zró¿nicowana na poszczególnych powierzchniach. Z badañ wynika, ¿eaktywnoœæ tego enzymu (x z lat 2001–2003) by³a istotnie wy¿sza na bogatszymsiedlisku BMœw – 11,7 mg NH3/10 g gleby ni¿ na siedlisku Bœw – 8,6 mg NH3/10 ggleby (F=2,49; p<0,05). Aktywnoœæ ureazy wykazywa³a tendencjê spadkow¹ wrazze spadkiem bonitacji drzewostanu, wyraŸniej zaznaczon¹ w Bœw, gdzie zmniej-sza³a siê z 9,9 mg NH3/10 g w I klasie do 7,95 mg NH3/10 g gleby w IV klasie,natomiast w BMœw odpowiednio z 12,2 mg NH3/10 g w I klasie do 11,3 mgNH3/10 g gleby w III klasie.
Aktywnoœæ asparaginazy, enzymu bior¹cego udzia³ w hydrolizacji asparaginyna amoniak, by³a zró¿nicowana w zale¿noœci od ¿yznoœci siedliska, lecz ob-
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 25

26 G. Olszowska i inni
Tab
ela
3.B
iom
asa
iilo
raz
met
abol
iczn
y(q
CO
2)dr
obno
ustr
ojów
wpo
ziom
ach
orga
nicz
nych
(Ofh
)im
iner
alny
ch(A
)gl
ebT
able
3.M
icro
bial
biom
ass
and
met
abol
icqu
otie
nt(q
CO
2)in
orga
nic
(Ofh
)an
dm
iner
al(A
)so
ilho
rizo
ns
Sied
-lis
koSi
te
Bon
i-ta
cja
Site
inde
x
Nr
pow
.Pl
otno
.
Bio
mas
adr
obno
ustr
ojów
Mic
robi
albi
omas
s(k
gC
biom
/ha)
qC
O2
2001
r.20
02r.
2003
r.20
01r.
2002
r.20
03r.
Ofh
AO
fh+
AO
fhA
Ofh
+A
Ofh
AO
fh+
AO
fhA
x w*
Ofh
Ax w
*O
fhA
x w*
Bœw
Fres
hco
nife
r-ou
sfo
res
I
113
231
163
108
3914
711
242
154
43,
53,
56,
63,
94,
55,
13,
53,
82
141
4418
515
034
184
131
6219
34,
23,
43,
64,
62,
93,
24,
33,
13,
33
144
5620
092
4313
513
550
185
3,70
2,5
2,7
4,05
3,7
3,8
4,5
2,5
2,8
x13
944
183
117
3915
612
651
177
43,
13,
35,
073,
53,
804,
663,
053,
26
II
476
6414
050
5610
613
233
165
4,5
2,7
2,80
3,72
3,93
3,9
3,9
3,4
3,40
584
5113
573
5312
610
235
137
6,3
2,5
2,8
6,9
3,5
3,80
5,73
3,79
3,98
662
4410
679
6514
468
3410
23,
902,
82,
96,
53,
73,
85,
23,
73,
8x
7453
127
6758
125
101
3413
54,
92,
72,
85,
73,
723,
94,
93,
603,
7
III
773
5412
765
3510
011
024
134
6,5
3,5
3,8
6,5
4,5
4,6
5,2
3,9
48
5043
9398
4714
511
739
156
7,8
2,9
3,4
4,5
4,1
4,1
4,8
44
988
2311
112
329
152
8441
125
6,8
3,2
3,9
6,70
3,5
4,1
6,2
3,6
4,1
x70
4011
095
3713
210
435
138
73,
203,
75,
894
4,3
5,4
3,8
4
IV
1073
5412
792
3913
169
4010
97,
74,
85,
17,
903,
94,
37,
53,
84,
211
6641
107
5133
8497
4614
39,
84,
45
5,80
4,06
4,20
6,22
4,33
4,51
1292
4413
687
4112
896
3012
94,
43,
83,
87,
33,
94,
356,
893,
774,
11x
7746
123
7738
114
8839
127
7,3
4,3
4,7
73,
94,
36,
84
4,3
BM
œwM
ixed
fres
hco
-ni
fero
usfo
rest
I
1386
9518
117
927
206
155
8924
44,
72,
22,
53,
53,
83,
74,
12,
22,
414
9199
190
126
6018
615
473
227
4,7
2,7
2,9
4,4
1,8
24,
92,
42,
615
7289
161
156
3419
013
436
170
4,7
2,1
2,4
4,3
3,21
3,4
4,5
3,2
3,30
x83
8417
715
440
194
148
6621
44,
72,
32,
64,
12,
953,
14,
502,
62,
8
II
1688
4413
280
5413
410
695
201
4,6
2,3
2,5
9,4
2,4
2,90
7,85
3,33
3,57
1712
245
167
193
4423
713
227
159
3,8
4,1
46,
613,
84,
476,
643,
934,
8818
9432
126
9635
131
149
3918
84,
85,
25,
24
3,94
45
3,1
3,20
x10
140
141
123
4416
712
954
183
4,4
3,9
3,9
6,7
3,37
3,8
6,50
3,4
3,9
III
1976
7515
169
3711
610
340
143
6,2
3,8
45,
303,
63,
745,
473,
293,
3920
9471
165
101
5815
910
071
177
83,
33,
68,
13,
63,
97,
23,
84,
0021
7942
121
7350
123
134
2616
08,
63,
44,
15,
73,
003,
36,
403,
64,
1x
8363
146
8152
133
112
4616
07,
63,
503,
96,
43,
43,
76,
43,
63,
8* œr
edni
aw
a¿on
apo
ziom
ówO
fhiA
* wei
ghte
dm
ean
ofO
fhan
dA
hori
zons

serwowane ró¿nice nie by³y istotne statystycznie. Aktywnoœæ tego enzymu w ca³ejbadanej warstwie gleby (x z lat 2001–2003) obni¿a³a siê wraz ze spadkiem bo-nitacji – w Bœw z 4,95 mg NH3/10 g w I klasie do 2,75 mg NH3/10 g w IV klasie, a wBMœw z 3,43 mg NH3/10 g w I klasie do 2,7 mg NH3/10 g w III klasie.
Nie stwierdzono istotnych ró¿nic pomiêdzy siedliskami pod wzglêdem aktyw-noœci fosfatazy kwaœnej, enzymu katalizuj¹cego przemiany fosforanów organicz-
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 27
Ure as e
0,0
4,0
8,0
12,0
16,0
Bœw BMœw
As paraginaza As pa ra gina se
0,0
2,0
4,0
6,0
I II III IV I II IIIBœw BMœw
mg
NH
/10
gg
leb
y3
mg
NH
/10
gs
oil
3
mg
NH
/10
gg
leb
y3
mg
NH
/10
gs
oil
3
Acid phosphata s e
0,0
4,0
8,0
12,0
Bœw BMœw
mg
fen
olu
/10
gg
leb
ym
gp
he
no
l/1
0g
so
il
De hydrog enazy De hydrogenas es
0,0
0,4
0,8
1,2
1,6
I II III IV I II IIIBœw BMœw
Bonitacja, s ie dlis koSite inde x, s ite
mg
TF
/g
gle
by
mg
TF
/g
so
il
2001 20022003 œredniamea n
Ure aza
I II III IV I II III
I II III IV I II III
Fo s fataza kwaœna
Ryc. 1. Aktywnoœæ enzymatyczna gleb borów sosnowych w latach 2001–2003Fig. 1. Enzymatic activity of pine forest soils in years 2001–2003; Bœw – fresh coniferous forest,BMœw – mixed fresh coniferous forest
28 G. Olszowska i inni
Tab
ela
4.A
ktyw
noϾ
enzy
mat
yczn
aw
pozi
omac
hor
gani
czny
ch(O
fh)
imin
eral
nych
(A)
gleb
(œre
dnia
zla
t20
01–2
003)
Tab
le4.
Enz
ymat
icac
tivity
inor
gani
c(O
fh)
and
min
eral
(A)
soil
hori
zons
(mea
nfr
om20
01–2
003)
Typ
sied
lisko
wy
lasu
Fore
stsi
tety
pe
Bon
i-ta
cja
Site
inde
x
Nad
leœn
ictw
oF
ores
tDis
tric
tL
eœni
ctw
oFo
rest
rang
e
Odd
zia³
podo
ddzi
a³C
ompa
rtm
ent
Ure
aza
Ure
ase
mg
NH
3/10
ggl
eby
mg
NH
3/10
gso
il
Asp
arag
inaz
aA
spar
agin
ase
mg
NH
3/10
ggl
eby
mg
NH
3/10
gso
il
Fos
fata
zakw
aœna
Aci
dph
osph
atas
e(m
gfe
nolu
/10
ggl
eby)
(mg
phen
ol/1
0g
soil)
Deh
ydro
gena
zyD
ehyd
roge
nase
s(m
gT
F/1
0g
gleb
y)(m
gT
F/10
gso
il)O
fhA
Ofh
+A
Ofh
AO
fh+
AO
fhA
Ofh
+A
Ofh
AO
fh+
A
Bœw
Fres
hco
nife
rous
fore
st
IW
³osz
czow
a
Goœ
ciêc
in45
f5,
645,
8811
,52
2,33
3,71
6,04
9,22
3,38
12,6
10,
310,
530,
84G
oœci
êcin
46c
4,75
5,18
9,93
1,93
2,76
4,68
7,32
3,07
10,4
00,
270,
390,
67Pê
kow
iec
97g
3,58
4,68
8,26
1,41
2,72
4,13
7,45
4,85
12,3
00,
160,
400,
56x
4,66
5,25
9,90
1,89
3,06
4,95
8,00
3,77
11,7
70,
250,
440,
69
IIO
pocz
no
Ro¿
enek
164
d2,
105,
067,
160,
620,
791,
414,
152,
666,
810,
180,
330,
50R
o¿en
ek16
7a
3,15
6,15
9,30
0,86
1,14
1,99
4,22
3,18
7,40
0,20
0,39
0,59
Ro¿
enek
163
a2,
176,
228,
390,
520,
551,
072,
832,
094,
920,
190,
330,
52x
2,48
5,81
8,28
0,67
0,82
1,49
3,73
2,64
6,38
0,19
0,35
0,54
III
Opo
czno
Myœ
libór
z19
1f
2,53
3,77
6,30
0,77
1,33
2,10
5,36
3,05
8,41
0,17
0,35
0,52
Ro¿
enek
165
a2,
224,
456,
660,
660,
621,
295,
962,
808,
760,
130,
210,
34B
ia³a
czka
60g
5,03
6,88
11,9
01,
733,
805,
538,
444,
4812
,92
0,26
0,70
0,95
x3,
265,
038,
291,
061,
922,
976,
593,
4410
,03
0,19
0,42
0,60
IVO
pocz
no
Kró
lów
ka6
i3,
603,
777,
370,
851,
402,
256,
172,
198,
360,
200,
420,
62K
róló
wka
4f
6,38
3,42
9,80
0,80
2,01
2,81
5,60
2,62
8,22
0,15
0,49
0,64
Kró
lów
ka5f
3,32
3,37
6,69
1,15
2,04
3,19
4,28
2,21
6,49
0,23
0,47
0,70
x4,
433,
527,
950,
931,
822,
755,
352,
347,
690,
190,
460,
65
BM
œwM
ixed
fres
hco
nife
rous
fore
st
IW
³osz
czow
a
Wol
aŒw
idz.
110
c5,
006,
3611
,36
1,37
1,90
3,27
7,42
3,89
11,3
10,
520,
491,
02M
otyc
zna
133
d5,
369,
8115
,18
0,92
2,23
3,15
5,82
3,51
9,33
0,31
0,45
0,76
Kur
zeló
w21
d4,
345,
6910
,04
1,27
2,58
3,86
7,21
3,20
10,4
10,
300,
340,
63x
4,90
7,29
12,1
91,
192,
243,
436,
823,
5310
,35
0,38
0,43
0,80
IIO
pocz
no
Ro¿
enek
164
j2,
797,
3810
,16
0,74
1,26
2,01
3,65
2,93
6,58
0,17
0,31
0,48
Ro¿
enek
166
d8,
374,
1912
,57
2,50
2,10
4,60
9,17
3,90
13,0
70,
890,
471,
36R
o¿en
ek16
9i
4,24
8,13
12,3
70,
971,
882,
855,
484,
109,
580,
320,
590,
91x
5,13
6,57
11,7
01,
401,
753,
156,
103,
649,
740,
460,
460,
91
III
Opo
czno
Myœ
libór
z20
2a
2,61
11,4
014
,01
0,64
1,18
1,82
3,24
3,55
6,79
0,29
0,79
1,08
Myœ
libór
z21
2b
3,58
6,29
9,87
0,68
1,37
2,05
4,04
5,33
9,38
0,33
0,71
1,04
Myœ
libór
z20
4c
4,86
5,11
9,97
1,50
2,63
4,12
7,93
3,56
11,4
90,
280,
741,
02x
3,68
7,60
11,2
80,
941,
732,
665,
074,
159,
220,
300,
751,
05

nych w nieorganiczne. Aktywnoœæ tego enzymu by³a nieznaczne wy¿sza na¿yŸniejszym siedlisku BMœw ni¿ w Bœw. Œrednia aktywnoœæ fosfatazy kwaœnej wlatach 2001-2003 dla ca³ej badanej warstwy gleby (poziomy Ofh+A) zmniejsza³asiê wraz z pogarszaniem siê bonitacji; w Bœw z 11,8 mg fenolu /10 g gleby w Iklasie do 7,7 mg fenolu w IV klasie, a w BMœw z 10,4 mg fenolu /10 g gleby w Iklasie do 9,2 mg fenolu w IV klasie. Obserwowane ró¿nice w aktywnoœci enzymupomiêdzy bonitacjami by³y istotne statystycznie jedynie na siedlisku Bœw (F=3,69;p<0,05).
Aktywnoœæ dehydrogenaz (katalizuj¹cych reakcje utleniania i redukcji), po-dobnie jak poprzednio omawianych enzymów, by³a istotnie wy¿sza (F=10,19;p<0,01) na siedlisku BMœw ni¿ Bœw. Nie stwierdzono natomiast zale¿noœci po-miêdzy aktywnoœci¹ dehydrogenaz a bonitacj¹; notowany na siedlisku BMœwwzrost aktywnoœci tych enzymów wraz ze spadkiem bonitacji (z 0,80 mg TF/ 10 gw I klasie do 1,05 mg TF/10g w III klasie) nie by³ istotny statystycznie.
4.4. Cechy taksacyjne drzewostanów sosnowych
Wiêkszoœæ badanych wskaŸników dendrometrycznych wykazywa³a zwi¹zek zjakoœci¹ badanych siedlisk; wy¿sze wartoœci notowano na bogatszych siedliskachBMœw ni¿ w Bœw (tab. 5).
Przeciêtna pierœnica drzewostanu D na siedlisku BMœw – 25,7 cm by³a istotniewy¿sza (F=111,7; p,<0,001) ni¿ w Bœw – 19,7 cm. Pierœnica drzewostanu zmniej-sza³a siê wraz ze spadkiem bonitacji drzewostanu: z 22,2 cm w I klasie do 18,5 cmw IV klasie w Bœw oraz z 27,3 cm w I klasie do 25,4 cm w III klasie w BMœw.
Przeciêtna wysokoœæ drzew H na siedlisku Bœw by³a istotnie ni¿sza (F=56,6;p<0,001) ni¿ w BMœw i zmniejsza³a siê wraz z bonitacj¹ drzewostanu: w Bœw z21,4 m w I klasie do 15,9 m w IV klasie, a w BMœw z 25,1 m w I klasie do 20,4 m wIII klasie.
Przeciêtny przekrój drzewostanu g by³ na ¿yŸniejszym siedlisku, w BMœw,istotnie wy¿szy (F=131; p<0,001) ni¿ w Bœw. Na obu siedliskach wskaŸnik tenmia³ tendencjê spadkow¹ wraz ze spadkiem bonitacji drzewostanu: z 0,039 m2 w Iklasie do 0,027 m2 w IV klasie w Bœw oraz z 0,059 m2 w I klasie do 0,047 m2 w IIklasie w BMœw.
Na siedlisku BMœw stwierdzono równie¿ istotnie wy¿sz¹ (F=9,98; p<0,01) ni¿w Bœw mi¹¿szoœæ drzewostanu w korze (Vk/ha). Na obu siedliskach wskaŸnik tenmia³ tendencjê spadkow¹ wraz z pogarszaniem siê ich jakoœci – z 442,7 m3/ha w Iklasie do 249,9 m3/ha w III klasie w BMœw oraz z 353,3 m3/ha ( I klasa) do218,1 m3/ha (IV klasa) w Bœw.
Jakoœæ siedliska mia³a tak¿e wyraŸny wp³yw na mi¹¿szoœæ grubizny drzewdrzewostanu (Vg/ha), która z 346,7 m3/ha w I klasie zmniejsza³a siê do 210 m3/ha wIV klasie w Bœw oraz z 440 m3/ha w I klasie do 247,6 m3/ha w III klasie w BMœw.Notowane ró¿nice pomiêdzy siedliskami by³y istotne statystycznie (F=11,52;p<0,01).
Istotnie wiêcej (F=95,9; p<0,001) drzew (L/ha) notowano na siedlisku Bœw ni¿BMœw, przy czym na siedlisku Bœw liczba drzew wzrasta³a wraz z pogarszaniem
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 29

30 G. Olszowska i inni
Tab
ela
5.C
echy
taks
acyj
neba
dany
chdr
zew
osta
nów
(œre
dnia
z10
0dr
zew
)T
able
5.T
axat
ion
feat
ures
ofsu
rvey
edst
ands
(mea
nfr
om10
0tr
ees)
Typ
sied
lis.l
asu
For
ests
itety
peB
onit
acja
Site
inde
xN
adle
œnic
two
Fore
stD
istr
ict
Leœ
nict
wo
Fore
stra
nge
Odd
zia³
Com
partm
ent
Wie
kA
geD
HL
gG
Vk
Vg
Zag
Zad
Bœw
Fres
hco
nife
rous
fore
s
IW
³osz
czow
a
Goœ
ciêc
in45
f54
23,5
23,5
858
0,04
3437
,29
403
397
0,77
1,25
Goœ
ciêc
in46
c51
21,5
20,4
950
0,03
6234
,42
327
320
0,77
1,08
Pêko
wie
c97
g56
21,5
20,3
958
0,03
6434
,93
330
323
0,92
1,02
x22
,17
21,4
092
2,2
0,03
935
,55
353,
334
6,7
0,82
1,12
IIO
pocz
no
Ro¿
enek
164
d52
21,1
19,1
925
0,03
5032
,33
288
282
0,68
1,06
Ro¿
enek
167
a52
19,2
16,3
1017
0,02
8929
,39
226
219
0,74
0,82
Ro¿
enek
163
a52
16,6
15,6
1563
0,02
1633
,67
254
242
1,14
0,91
x18
,97
17,0
011
68,1
0,02
831
,80
256,
024
7,7
0,85
0,93
III
Opo
czno
Myœ
libór
z19
1f
6720
,615
,776
30,
0333
25,4
018
618
20,
720,
67R
o¿en
ek16
5a
5217
,616
,412
580,
0245
30,7
724
223
30,
771,
17B
ia³a
czka
60g
6219
,918
,410
330,
0312
32,2
527
927
20,
861,
09x
19,3
716
,83
1018
,10,
030
29,4
723
5,8
229,
00,
780,
98
IVO
pocz
no
Kró
lów
ka6
i67
20,1
16,2
1008
0,03
1731
,96
243
237
0,80
1,15
Kró
lów
ka4
f67
1916
,498
30,
0284
27,8
821
620
90,
781,
01K
róló
wka
5f62
16,3
15,1
1275
0,02
0826
,50
195
185
0,88
0,98
x18
,47
15,9
010
88,9
0,02
728
,78
218,
121
0,3
0,82
1,05
BM
œwM
ixed
fres
hco
nife
rous
fore
st
IW
³osz
czow
a
Wol
aŒw
idz.
110
c55
25,6
24,5
792
0,05
1540
,75
456
452
0,74
1,40
Mot
yczn
a13
3d
6127
,425
,768
50,
0588
40,2
846
846
60,
771,
30K
urze
lów
21d
6629
25,2
540
0,06
5935
,61
404
403
0,69
1,04
x27
,33
25,1
367
2,2
0,05
938
,88
442,
744
0,3
0,73
1,25
IIO
pocz
no
Ro¿
enek
164
j57
25,9
21,8
569
0,05
2930
,06
298
296
0,49
1,03
Ro¿
enek
166
d52
25,2
19,7
594
0,04
9929
,60
266
263
0,43
0,99
Ro¿
enek
169
i57
22,4
20,6
756
0,03
9429
,80
285
280
0,65
0,98
x24
,50
20,7
063
9,6
0,04
729
,82
283,
127
9,6
0,52
1,00
III
Opo
czno
Myœ
libór
z20
2a
6722
,620
,662
50,
0402
25,1
024
023
60,
590,
87M
yœlib
órz
212
b73
27,8
20,8
475
0,06
0528
,73
271
270
0,52
0,92
Myœ
libór
z20
4c
7225
,719
,851
00,
0521
26,5
523
923
70,
550,
82x
25,3
720
,40
536,
70,
051
26,7
924
9,9
247,
60,
550,
87D
–pr
zeci
êtna
pier
œnic
adr
zew
osta
nu(c
m),
H–
prze
ciêt
naw
ysok
oϾ
drze
wos
tanu
(m),
L–
liczb
adr
zew
(szt
/ha)
,g–
prze
ciêt
nypr
zekr
ójdr
zew
osta
nu(m
2 ),G
–pi
erœn
icow
epo
lepr
zekr
oju
drze
wos
tanu
(m2 /h
a);
Vk
–m
i¹¿s
zoϾ
drze
wos
tanu
wko
rze
(m3 /h
a),
Vg
–m
i¹¿s
zoϾ
grub
izny
drze
wdr
zew
osta
nu(m
3 /ha)
,Z
ag
–za
gêsz
czen
ie,Z
ad
–za
drze
wie
nie
D–
aver
age
D.B
.H.(
cm),
H–
mea
nhe
ight
ofst
and
(m),
L–
stem
s/ha
,g–
mea
nst
and
sect
ion
(m2 ),
G–
basa
lare
a(m
2 /ha)
,Vk
–st
and
volu
me
with
the
bark
(m3 /h
a),
Vg
–st
and
volu
me
ofla
rge
timbe
r(m
3 /ha)
,Zag
–de
nsity
,Zad
–de
gree
ofcr
opde
nsity

siê jakoœci siedliska z 922 w I klasie do 1088 w IV klasie, a na siedlisku BMœwobserwowano spadek liczby drzew wraz ze spadkiem bonitacji z 672 w I klasie do536 w III klasie. Zagêszczenie (Zag) by³o istotnie wy¿sze (F=52; p<0,0001) w Bœwni¿ w BMœw i w tym ostatnim istotnie obni¿a³o siê (F=16,2; p<0,001) wraz zespadkiem bonitacji z 0,73 w I klasie do 0,53 w III klasie. Nie stwierdzono istotnychró¿nic w zadrzewieniu (Zad) pomiêdzy siedliskami, z tym ¿e na siedlisku BMœwwielkoœæ tego wskaŸnika istotnie obni¿a³a siê (F=8,95; p<0,001) wraz ze spadkiembonitacji drzewostanu z 1,25 w I klasie do 0,87 w III klasie.
Œrednie pierœnicowe pole przekroju drzewostanu (G/ha) by³o nieznaczniewy¿sze na siedlisku BMœw ni¿ w Bœw i wykazywa³o tendencjê spadkow¹ wraz zespadkiem bonitacji; w Bœw z 35,6 m2/ha w I klasie do 28,8 m2/ha w IV klasie a wBMœw z 38,9 m2/ha w I klasie do 26,8 m2/ha w III klasie – ró¿nice te by³y istotnestatystycznie (F=37,8; p<0,0001).
4.5. Biologiczny wskaŸnik ¿yznoœci gleb
Do oceny jakoœci siedlisk badanych powierzchni zastosowano biologicznywskaŸnik ¿yznoœci gleb F, obliczany na podstawie parametrów chemicznych,odzwierciedlaj¹cych zasobnoœæ gleb w sk³adniki pokarmowe oraz aktywnoœæ bio-logiczn¹ gleb. Do obliczenia jego wartoœci, zmodyfikowano metodê Myœkowa i in.(1996), korzystaj¹c z równania:
F M S V= + +2 2 2
gdzie:M – aktywnoœæ biologiczna gleb,S – suma kationów zasadowych,V – stopieñ wysycenia kompleksu sorpcyjnego zasadami.Do powy¿szego równania wstawiano standaryzowane wyniki pomiarów (w
jednostkach odchylenia standardowego), przy czym za wartoœæ M przyjêto kolejnojeden z testowanych parametrów aktywnoœci biologicznej gleb, tj. aktywnoœæbadanych enzymów glebowych: dehydrogenaz (D), ureazy (U), asparaginazy (A) ifosfatazy kwaœnej (F-kw), biomasê drobnoustrojów (Cbiom), tempo mineralizacjiwêgla (g C-CO2), iloraz metaboliczny drobnoustrojów (qCO2).
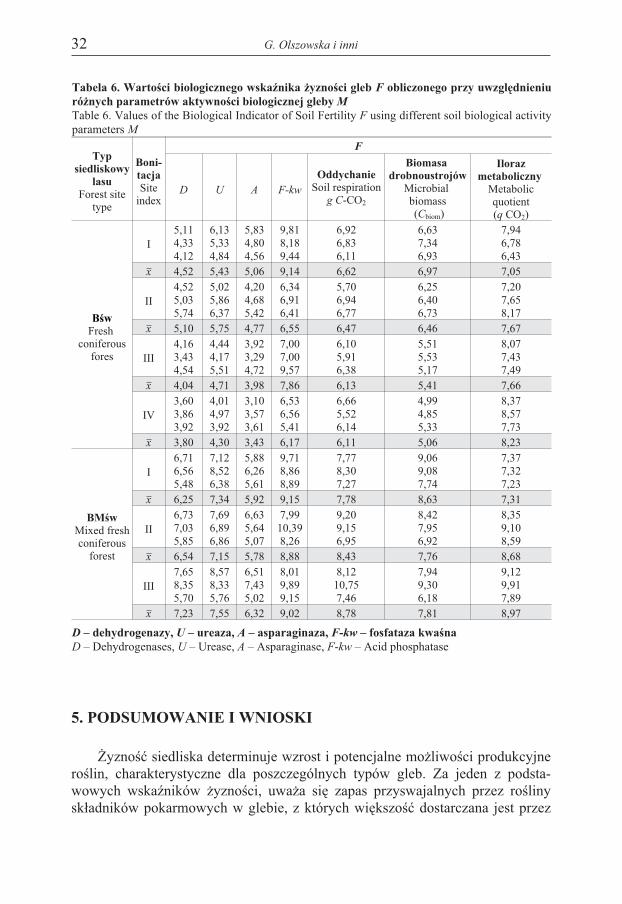
Wartoœci wskaŸnika F dla badanych powierzchni przedstawiono w tabeli 6.Niezale¿nie od u¿ytego w równaniu parametru M, wskaŸnik F by³ istotnie wy¿szyna ¿yŸniejszym siedlisku BMœw ni¿ Bœw. Jego wartoœæ mala³a wraz ze spadkiembonitacji drzewostanu tylko w przypadku siedliska Bœw, gdy za M przyjêto aktyw-noœæ asparaginazy i fosfatazy kwaœnej, intensywnoœæ oddychania oraz biomasêdrobnoustrojów, a wzrasta³a, gdy za M przyjêto wartoœæ ilorazu metabolicznego.Najwy¿sze wartoœci wskaŸnik F osi¹ga³, gdy za M przyjêto aktywnoœæ fosfatazykwaœnej (F-kw), iloraz metaboliczny (qCO2), tempo mineralizacji (g C-CO2) orazbiomasê drobnoustrojów (Cbiom), a najni¿sze, gdy za M przyjêto aktywnoœæ aspara-ginazy (A), dehydrogenaz (D) i ureazy (U).
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 31
5. PODSUMOWANIE I WNIOSKI
¯yznoœæ siedliska determinuje wzrost i potencjalne mo¿liwoœci produkcyjneroœlin, charakterystyczne dla poszczególnych typów gleb. Za jeden z podsta-wowych wskaŸników ¿yznoœci, uwa¿a siê zapas przyswajalnych przez roœlinysk³adników pokarmowych w glebie, z których wiêkszoœæ dostarczana jest przez
32 G. Olszowska i inni
Tabela 6. Wartoœci biologicznego wskaŸnika ¿yznoœci gleb F obliczonego przy uwzglêdnieniuró¿nych parametrów aktywnoœci biologicznej gleby M
Table 6. Values of the Biological Indicator of Soil Fertility F using different soil biological activityparameters M
Typsiedliskowy
lasuForest site
type
Boni-tacjaSite
index
F
D U A F-kw
OddychanieSoil respiration
g C-CO2
Biomasadrobnoustrojów
Microbialbiomass(Cbiom)
Ilorazmetaboliczny
Metabolicquotient(q CO2)
BœwFresh
coniferousfores
I5,11 6,13 5,83 9,81 6,92 6,63 7,944,33 5,33 4,80 8,18 6,83 7,34 6,784,12 4,84 4,56 9,44 6,11 6,93 6,43
x 4,52 5,43 5,06 9,14 6,62 6,97 7,05
II4,52 5,02 4,20 6,34 5,70 6,25 7,205,03 5,86 4,68 6,91 6,94 6,40 7,655,74 6,37 5,42 6,41 6,77 6,73 8,17
x 5,10 5,75 4,77 6,55 6,47 6,46 7,67
III4,16 4,44 3,92 7,00 6,10 5,51 8,073,43 4,17 3,29 7,00 5,91 5,53 7,434,54 5,51 4,72 9,57 6,38 5,17 7,49
x 4,04 4,71 3,98 7,86 6,13 5,41 7,66
IV3,60 4,01 3,10 6,53 6,66 4,99 8,373,86 4,97 3,57 6,56 5,52 4,85 8,573,92 3,92 3,61 5,41 6,14 5,33 7,73
x 3,80 4,30 3,43 6,17 6,11 5,06 8,23
BMœwMixed freshconiferous
forest
I6,71 7,12 5,88 9,71 7,77 9,06 7,376,56 8,52 6,26 8,86 8,30 9,08 7,325,48 6,38 5,61 8,89 7,27 7,74 7,23
x 6,25 7,34 5,92 9,15 7,78 8,63 7,31
II6,73 7,69 6,63 7,99 9,20 8,42 8,357,03 6,89 5,64 10,39 9,15 7,95 9,105,85 6,86 5,07 8,26 6,95 6,92 8,59
x 6,54 7,15 5,78 8,88 8,43 7,76 8,68
III7,65 8,57 6,51 8,01 8,12 7,94 9,128,35 8,33 7,43 9,89 10,75 9,30 9,915,70 5,76 5,02 9,15 7,46 6,18 7,89
x 7,23 7,55 6,32 9,02 8,78 7,81 8,97
D – dehydrogenazy, U – ureaza, A – asparaginaza, F-kw – fosfataza kwaœnaD – Dehydrogenases, U – Urease, A – Asparaginase, F-kw – Acid phosphatase

drobnoustroje glebowe w wyniku rozk³adu substancji organicznej. Istotna roladrobnoustrojów glebowych w kszta³towaniu ¿yznoœci i urodzajnoœci gleb leœnychjest szeroko udokumentowana; szereg prac wskazuje na siln¹ korelacjê miêdzybiomas¹ i aktywnoœci¹ drobnoustrojów a produkcyjnoœci¹ drzewostanów (Myroldi in. 1986, Zak i in. 1994, Kurka i Starr 1997). Potwierdzaj¹ to wyniki badañintensywnoœci oddychania gleb i biomasy drobnoustrojów, których wartoœci by³ywy¿sze na bogatszym siedlisku BMœw ni¿ Bœw. Stwierdzono równie¿, ¿e biomasadrobnoustrojów wykazywa³a tendencjê spadkow¹ wraz ze spadkiem bonitacjidrzew. Z kolei wzrost tempa respiracji biomasy (qCO2), jaki obserwowano wraz zpogarszaniem siê jakoœci siedliska, wskazuje na s³abszy wzrost biomasy drobno-ustrojów, a w nastêpstwie – na mniejszy zapas przyswajalnych form sk³adnikówpokarmowych w glebie.
¯yznoœæ siedlisk zwi¹zana jest tak¿e z aktywnoœci¹ katalizowanych przezenzymy procesów rozk³adu i przemiany substancji organicznej w glebach (Russel iKobus 1974, Myœków 1981, Gliñski i in. 1983, Myœków i in. 1996). Wskazuj¹ na totak¿e przeprowadzone badania, które wykaza³y spadek aktywnoœci ureazy, as-paraginazy, fosfatazy kwaœnej i dehydrogenaz wraz ze spadkiem jakoœci siedliskleœnych. Ponadto aktywnoœæ ureazy, asparaginazy i fosfatazy kwaœnej wykazywa³ytendencjê spadkow¹ wraz z obni¿aniem siê bonitacji drzewostanu.
Jak donosz¹ Puchalski i Prusinkiewicz (1990), Garbaye i Bonneau (1997) orazRanger i Turpault (1999), ¿yznoœæ siedliska uwarunkowana jest iloœci¹ dostêpnychdla drobnoustrojów sk³adników pokarmowych w glebie. Stwierdzony w przed-stawianych badaniach gorszy stan mikrobiologiczny gleb, a tak¿e ni¿sza aktyw-noœæ badanych enzymów w Bœw ni¿ w BMœw, œwiadcz¹ o mniej intensywnymprocesie rozk³adu substancji organicznej i w rezultacie mniej efektywnym uwal-nianiu przyswajalnych sk³adników pokarmowych w glebie. Potwierdzi³y to prze-prowadzone badania chemiczne, wskazuj¹ce ¿e zasobnoœæ gleb w podstawowesk³adniki od¿ywcze by³a wy¿sza na ¿yŸniejszym siedlisku BMœw ni¿ Bœw, ztendencj¹ spadkow¹ wraz ze spadkiem bonitacji drzewostanu.
Za miarodajny wskaŸnik ¿yznoœci siedlisk uwa¿a siê w³aœciwoœci gleb, cha-rakteryzowane m.in. sk³adem chemicznym, aktywnoœci¹ biologiczn¹ i enzyma-tyczn¹ (Burns 1982, Bruchwald i Kliczkowska 1997, Zaguralskaja 1998). Wprowadzonych badaniach zastosowano biologiczny wskaŸnik ¿yznoœci gleby F,stosowany wczeœniej do oceny jakoœci gleb rolnych i wykazuj¹cy istotn¹ korelacjêz plonami kukurydzy i ziemniaków (Myœkow i in. 1996). WskaŸnik F przyjmowa³ni¿sze wartoœci na siedlisku ubo¿szym Bœw ni¿ BMœw, a prawid³owoœæ ta wy-stêpowa³a niezale¿nie od tego, który z parametrów aktywnoœci biologicznej (D, U,A, F-kw, qCO2, Cbiom, g C-CO2) przyjêto w obliczeniach.
W praktyce leœnej, jako wskaŸniki ¿yznoœci siedliska mog¹ s³u¿yæ cechytaksacyjne drzewostanów (Bruchwald 1995, 1997, Sikorska 1999). Przeprowa-dzona analiza korelacyjna (tab. 7) wykaza³a istotn¹ zale¿noœæ pomiêdzy wskaŸ-nikiem F a parametrami dendrometrycznymi, tj. przeciêtn¹ pierœnic¹ D, przeciêtn¹wysokoœci¹ H i przeciêtnym przekrojem drzewostanu g. Wskazuje to na miaro-
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 33
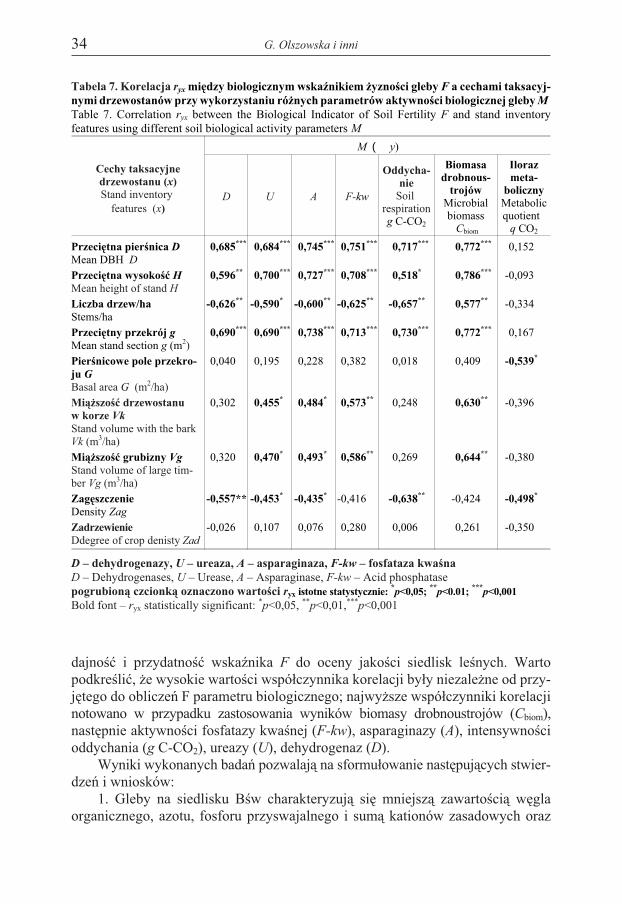
dajnoœæ i przydatnoœæ wskaŸnika F do oceny jakoœci siedlisk leœnych. Wartopodkreœliæ, ¿e wysokie wartoœci wspó³czynnika korelacji by³y niezale¿ne od przy-jêtego do obliczeñ F parametru biologicznego; najwy¿sze wspó³czynniki korelacjinotowano w przypadku zastosowania wyników biomasy drobnoustrojów (Cbiom),nastêpnie aktywnoœci fosfatazy kwaœnej (F-kw), asparaginazy (A), intensywnoœcioddychania (g C-CO2), ureazy (U), dehydrogenaz (D).
Wyniki wykonanych badañ pozwalaj¹ na sformu³owanie nastêpuj¹cych stwier-dzeñ i wniosków:
1. Gleby na siedlisku Bœw charakteryzuj¹ siê mniejsz¹ zawartoœci¹ wêglaorganicznego, azotu, fosforu przyswajalnego i sum¹ kationów zasadowych oraz
34 G. Olszowska i inni
Tabela 7. Korelacja ryx miêdzy biologicznym wskaŸnikiem ¿yznoœci gleby F a cechami taksacyj-nymi drzewostanów przy wykorzystaniu ró¿nych parametrów aktywnoœci biologicznej gleby M
Table 7. Correlation ryx between the Biological Indicator of Soil Fertility F and stand inventoryfeatures using different soil biological activity parameters M
Cechy taksacyjnedrzewostanu (x)Stand inventory
features �x)
M ( y)
D U A F-kw
Oddycha-nie
Soilrespirationg C-CO2
Biomasadrobnous-
trojówMicrobialbiomass
Cbiom
Ilorazmeta-
bolicznyMetabolicquotient
q CO2
Przeciêtna pierœnica D
Mean DBH D
0,685*** 0,684*** 0,745*** 0,751*** 0,717*** 0,772*** 0,152
Przeciêtna wysokoœæ H
Mean height of stand H
0,596** 0,700*** 0,727*** 0,708*** 0,518* 0,786*** -0,093
Liczba drzew/haStems/ha
-0,626** -0,590* -0,600** -0,625** -0,657** 0,577** -0,334
Przeciêtny przekrój g
Mean stand section g (m2)0,690*** 0,690*** 0,738*** 0,713*** 0,730*** 0,772*** 0,167
Pierœnicowe pole przekro-ju G
Basal area G (m2/ha)
0,040 0,195 0,228 0,382 0,018 0,409 -0,539*
Mi¹¿szoœæ drzewostanuw korze Vk
Stand volume with the barkVk (m3/ha)
0,302 0,455* 0,484* 0,573** 0,248 0,630** -0,396
Mi¹¿szoœæ grubizny Vg
Stand volume of large tim-ber Vg (m3/ha)
0,320 0,470* 0,493* 0,586** 0,269 0,644** -0,380
ZagêszczenieDensity Zag
-0,557** -0,453* -0,435* -0,416 -0,638** -0,424 -0,498*
ZadrzewienieDdegree of crop denisty Zad
-0,026 0,107 0,076 0,280 0,006 0,261 -0,350
D – dehydrogenazy, U – ureaza, A – asparaginaza, F-kw – fosfataza kwaœnaD – Dehydrogenases, U – Urease, A – Asparaginase, F-kw – Acid phosphatasepogrubion¹ czcionk¹ oznaczono wartoœci ryx istotne statystycznie: *
p<0,05; **p<0.01; ***
p<0,001Bold font – ryx statistically significant: *p<0,05, **p<0,01,***p<0,001

wiêksz¹ kwasowoœci¹ hydrolityczn¹ ni¿ gleby na siedlisku BMœw. Wartoœci po-wy¿szych parametrów wykazywa³y wyraŸny zwi¹zek z bonitacj¹ drzewostanu,zw³aszcza na siedlisku Bœw.
2. WyraŸny wp³yw na stan mikrobiologiczny gleb mia³a jakoœæ siedlisk.Intensywnoœæ oddychania gleb i biomasa drobnoustrojów by³y wyraŸnie wy¿sze nabogatszym siedlisku BMœw ni¿ Bœw. Z kolei wartoœæ ilorazu metabolicznego(qCO2) wzrasta³a wraz z pogarszaniem siê jakoœci siedliska i bonitacji, co oznaczani¿sz¹ wydajnoœæ wzrostu drobnoustrojów, efektem czego jest mniejsza biomasadrobnoustrojów i mniejszy zapas sk³adników pokarmowych w glebie.
3. Jakoœæ siedliska mia³a wp³yw na aktywnoœæ niektórych badanych enzymówglebowych, tj. ureazy, fosfatazy kwaœnej i dehydrogenaz. Ich wartoœci by³y ni¿szew Bœw ni¿ w BMœw i spada³y wraz z obni¿aniem siê bonitacji drzewostanu.
4. Na miarodajnoœæ zastosowanego do oceny jakoœci siedlisk biologicznegowskaŸnika ¿yznoœci gleb F wskazuje istotna korelacja jego wartoœci z niektórymicechami taksacyjnymi, charakteryzuj¹cymi produkcyjnoœæ drzewostanu: przeciêt-n¹ pierœnic¹ D, przeciêtn¹ wysokoœci¹ H i przeciêtnym przekrojem drzewostanu g.
5. Najwy¿sze wartoœci wspó³czynnika korelacji pomiêdzy wskaŸnikiem F acechami taksacyjnymi drzewostanu uzyskano, gdy za parametr biologiczny przy-jêto biomasê drobnoustrojów (Cbiom), intensywnoœæ oddychania (g C-CO2) orazaktywnoœæ fosfatazy kwaœnej (F-kw) i asparaginazy (A).
6. Biologiczny wskaŸnik ¿yznoœci gleb F zosta³ opracowany dla borów sosno-wych, st¹d dla innych siedlisk jego poprawnoœæ i przydatnoœæ powinny byæ zwe-ryfikowane.
Praca zosta³a z³o¿ona 21.01.05 r. i przyjêta przez Komitet Redakcyjny 22.02.05 r.
THE USE OF BIOLOGICAL ACTIVITY STUDIES TO DETERMINEA SOIL FERTILITY INDICATOR IN PINE STANDS ON FRESH CONIFEROUSAND ON MIXED FRESH CONIFEROUS FOREST SITES
Summary
In 2001–2003, soil chemistry, soil biological activity, and pine stands characteristics alonga gradient of forest site quality were investigated. The research plots were set up in Scots pinestands growing on fresh coniferous and mixed fresh coniferous forest sites, located in W³osz-czowa and Opoczno Forest Districts. The aim of the study was to test the soil biological activityparameters which could be useful in diagnosing quality of forest sites.
The soils on fresh coniferous forest sites were characterized by lower concentrations of car-bon, nitrogen, phosphorus, and base cations and were more acidic, compared to those on mixedfresh coniferous forest sites. The values of above parameters were found to be closely related tostand site indices, especially on fresh coniferous forest sites.
Soil biological activity appeared to be dependent on site quality. Soil respiration, microbialbiomass and activity of some investigated enzymes (urease, acid phosphatase and dehydroge-
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 35

nases) were positively correlated with site index, and thus were distinctly higher on more fertilemixed fresh coniferous sites than on fresh coniferous sites. To evaluate site quality of the studiedplots, several chemical and biological parameters were chosen to calculate the values ofBiological Indicator of Soil Fertility (F), defined as:
F M S V= + +2 2 2
where: M – soil biological activity, S – sum of base cations, V – base saturation.As a M – one of the tested soil biological activity parameters were taken into account: dehy-
drogenases, urease, asparaginase, acid phosphatase, microbial biomass (Cbiom), metabolic quo-tient (qCO2) or soil respiration. F – values turned out to be lower on fresh coniferous sites than onmixed fresh coniferous sites, irrespective of which biological parameters was taken into F calcu-lation. The significant correlation found between the F – values and some stand taxation featuresassociated with stand productivity (d.b.h., height of stand and stand section) imply, that the F in-dex, calculated on the base of biological parameters, can provide a valid estimate of forest sitequality.
(transl. M. T.)
LITERATURA
Aikio S., Väre H., Strömmer R. 2000: Soil microbial activity and biomass in the primary successionof a dry heath forest. Soil Biol. Biochem., 32: 1091-1100.
Anderson J. P. E., Domsch K. H. 1978: A physiological method for quantitative measurementof microbial biomass in soil. Soil Biol. Biochem., 10: 215-21.
Anderson T. H., Domsch K. H. 1992: The metabolic quotient for CO2 (qCO2) as specific activityparameter to assess the effect of environment condition, such as pH, on the microbial biomassof forest soils. Soil Biol. Biochem., 25: 393-395.
Balicka N. 1986: Wykorzystanie wskaŸników mikrobiologicznych w analizie œrodowiska glebowego.Post. Mikrob. 25, 3/4: 289-291.
Bruchwald A. 1995: Dendrometria. Wydawnictwo SGGW. Warszawa, ss.450.Bruchwald A., Kliczkowska A. 1997: Kszta³towanie siê bonitacji dla drzewostanów sosnowych
Polski. Prace Inst. Bad. Leœ., A, 838: 63-73.Burns R. G. 1982: Enzyme activity in soil: location and a possible role on microbial ecology. Soil.
Biol. Biochem., 34: 423-427.Federer C. A., Turcotte D. , Smith C. T. 1993: The organic fraction – bulk density relationship and the
expression of nutrient content in forest soils. Can. J. For. Res., 23: 1026-1032.Galstjan A. Š. 1963: K ocenke stepeni plodorodija poèvy fermentativnymi reakcijami. Mikro-
organizmy v selskom chozjajstve. Izd. MGU: 327-335.Galstjan A. Š. 1978: Opredelenie aktivnosti fermentov poèv – metodièeskie ukazanija. Erevan'.Garbaye J., Bonneau M. 1997: Assuring sufficient nutrient supply for trees – a basic condition of
sustainable forests. IUFRO Occas., 9: 28-31.Gliñski J., Stêpniewski W., £abuda S. 1983: Pobieranie tlenu i wydzielanie dwutlenku wêgla w
œrodowisku glebowym. Probl. Agrofiz., 39: 3-72.Hofmann E. 1955: Die Enzyme im Boden und ihre Bedeutung für seine Bilogie und Fruchtbarkeit. Z.
Acker u. Pflanzenbau., 100: 31-35.Instrukcja laboratoryjna dla pracowni gleboznawczo-nawo¿eniowych (red. A. Kowalkowski), 1973,
Warszawa - Sêkocin.Jenkinson D. S., Ladd J. N. 1981: Microbial biomass in soil: measurement and turover. [W:] Soil
Biochemistry, (eds: E. A. Paul and J. N. Ladd), Marcel Dekker, New York, 5: 415-471.
36 G. Olszowska i inni

Koper J., Piotrowska A. 1999a: Biochemiczne wskaŸniki ¿yznoœci gleby ukszta³towane w wynikuwieloletniego nawo¿enia organiczno-mineralnego. Zesz. Nauk. ART Bydgoszcz, 220: 151-158.
Koper J., Piotrowska A. 1999b: Aktywnoœæ enzymatyczna gleb jako parametr jej ¿yznoœci wywo³anysystemem uprawy. Zesz. Probl. Post. Nauk Rol., 467(I): 127-134.
Kurka A. M., Starr M. 1997: Relationship between decomposition of cellulose in the soil and treestand characteristics in natural boreal forests. Plant and Soil, 197: 167-175.
McGill W. B., Cannon K. R., Robertson J. A., Cook F. D. 1986: Dynamics of soil microbial biomassand water soluble organic C in Breton L after 50 years of cropping to two rotations. Can. J. SoilSci., 66:1-19.
Myrold D. D., Matson P. A., Peterson D. L. 1989: Relationships between soil microbial properties andabove-ground stand characteristics of conifer forests in Oregon. Biogeochemistry, 8: 265-281.
Myœków W. 1981: Próby wykorzystania wskaŸników aktywnoœci mikrobiologicznej do oceny ¿yz-noœci gleb. Post. Mikrob., XX, 3/4: 173-192.
Myœków W., Stachyra A., Ziêba S., Masiak D. 1996: Aktywnoœæ biologiczna gleby jako wskaŸnik jej¿yznoœci i urodzajnoœci. Rocz. Glebozn., XLVII: 89-99.
Ostrowska A., Gawliñski S., Szczubia³ka Z. 1991: Metody analizy i oceny w³aœciwoœci gleb i roœlin.Instytut Ochrony Œrodowiska, Warszawa, ss.334.
Parkinson D. 1979: Aspects of the microbial ecology of forest ecosystems. [W:] Forests: freshperspectives from ecosystem analysis (ed. R.Waring). Proc. of the 40th Annual BiologyColloquium, Oregon State University, Corvallis, 109-117.
Puchalski T., Prusinkiewicz Z. 1990: Ekologiczne podstawy siedliskoznawstwa leœnego. Wyd. IIPWRiL, Warszawa, ss. 619.
Ranger J., Turpault M. 1999: Input-output nutrient budgets as a diagnostic tool for sustainable forestmanagement. For. Ecol. Manag., 122, 1/2: 139-154.
Russel S. 1972: Metody oznaczania enzymów glebowych. PTG Komisja Biologii Gleby. Warszawa,ss 64.
Russel S., Kobus J., 1974: Aktywnoœæ dehydrogenaz w ró¿nych typach gleb polskich. Pr. Kom. Biol.Gleby PTG, 12: 65-66.
Sikorska E. 1999: Siedliska leœne. Cz. I. Siedliska obszarów ni¿owych . Wyd. AR Kraków, ss. 136.Trampler T., M¹kosa K., Gir¿da A., B¹kowski J., Dmyterko E. 1990: Siedliskowe podstawy hodowli
lasu. PWRiL Warszawa, ss.197.Zaguralskaja L. M. 1998: Biologièeskaja aktivnost poèv kak pokazatel' uslovij rosta lesnych na-
sa�denij. Lesovedenie, 1: 24-29.Zak D. R., Grigal D. F., Gleeson S., Tilman D. 1990: Carbon and nitrogen cycling during old-field
succession: constrains on plant and microbial biomass. Biogeochemistry, 11: 111-129.Zak D. R., Tilman D., Parmenter R. R., Rice C. W., Fisher F. M., Vose J., Milchanus D., Martin C. W.
1994: Plant production and soil microorganisms in late-successional ecosystems: a continentalstudy. Ecology, 75: 2333-2347.
Wykorzystanie badañ aktywnoœci biologicznej do wyznaczenia wskaŸnika ¿yznoœci 37


















