
Wybór systemu ekspresyjnego - pg.gda.pl · Bacillus spp. -zalety Organizm GRAS (generally regarded...
73
Wybór systemu ekspresyjnego
Transcript of Wybór systemu ekspresyjnego - pg.gda.pl · Bacillus spp. -zalety Organizm GRAS (generally regarded...

Wybór systemu ekspresyjnego

Bakterie

Escherichia coli
Zalety:
wysoki poziom ekspresji (do 500 mg/l hodowli)
dobrze poznany system
proste technologie
niskie koszty
Wady:
brak modyfikacji potranslacyjnych
ograniczona wielkość klonowanego DNA
różnice w „codon usage”

Bacillus spp. - zalety
Organizm GRAS (generally regarded as safe)
Możliwość produkcji wielu białek do pożywki
Bakterie dobrze scharakteryzowane (znany genom B. subtilis
i B. licheniformis)
Rośnie na ubogich podłożach
Wysoki poziom ekspresji (10 mg/litr)

Bacillus spp. - ograniczenia
Preferowana ekspresja homologiczna:
Geny pochodzące z Bacillus sp. produkowane z dużą wydajnością (skala
produkcji białek gram/litr)
Niewydajna ekspresja białek eukariotycznych (skala produkcji białek
mg/litr)

Grzyby

Saccharomyces cerevisiae
ZALETY
Średni poziom ekspresji
Glikozylacja białek
Białka kierowane do
pożywki
Dobrze poznany proces
fermentacji
Łatwa liza komórek
WADY
Mała wydajność
transformacji
Ograniczona ilość wektorów

Pichia pastoris
Modyfikacje potranslacyjne białek
Poziom ekspresji genów heterogenicznych jest z reguły od 10- do 100- razy wyższy niż w drożdżachSaccharomyces cerevisiae

Pichia pastoris
Produkowane białka mogą być kierowane do: cytoplazmy
na zewnątrz komórki (mała ilość białek Pichii kierowanych na zewnątrz komórki)
Produkcję białek rekombinowanych w Pichia pastoris powinno prowadzić się w odpowiednich bioreaktorach (pełne wykorzystanie „potencjału ekspresyjnego P. pastoris)
Glikozylacja białek posiadających w swej sekwencji motyw Asn-X-Ser/Thr może prowadzić do utraty aktywności enzymatycznej lub antygenowych eksprymowanego białka heteregonicznego w Pichia pastoris

Kluyveromyces lactis
Wysoki poziom ekspresji
Możliwość zakupu gotowych kompetentnych komórek K.
lactis
Proste protokoły (dla osób bez doświadczenia w pracy z
drożdżami)
Możliwość wykorzystania systemów dla celów
komercyjnych (system sublicencjonowania)
Możliwość produkcji białek na zewnątrz lub wewnątrz
komórek
http://www.neb.com/nebecomm/products/productE1000.asp

Kluyveromyces lactis
Ekspresja w drożdżach zachodzi spod silnego promotora LAC4, który NIE umożliwia ekspresji w E. coli (geny kodujące białka toksyczne dla E. coli mogą być klonowane do plazmidu pKLAC1 w bakteriach przed ich ekspresją w systemach drożdżowych)
Białka produkowane w K. lactis ulegają modyfikacjom (glikozylacja)
Selekcja odbywa się bez użycia antybiotyków – acetamidaza (amdS) z pKLAC1 pozwala transformowanym komórkom na użycie acetamidu, jako źródła azotu

Hansenula polymorpha
Do 150 kopii klonowanego genu na komórkę
Możliwość koekspresjii do czterech różnych genów
Tanie pożywki
Wysoki poziom ekspresji do13.5 g/L
Stabilność białek - niewielka aktywność proteolityczna
http://www.artes-biotechnology.com/expres/hansenula/hansenula01.jsp

Trichoderma reesei
Wysoki poziom ekspresji
Glikozylacja białek
Możliwość ekspresji
zewnątrzkomórkowej
Mała wydajność
transformacji
Trudny proces fermentacji
Ograniczony wybór
wektorów
Specyficzny codon usage

Komórki owadzie

Baculovirus system, komórki owadzie Sf9
Glikozylacja
Proste zwiększanie skali
Trudne warunki hodowli
Powolny wzrost
www.invitrogen.com

Hodowle komórkowe

Komórki zwierzęce
Zalety Glikozylacja
Mogą być z nich wyprowadzane ciągłe linie komórkowe
Niektóre komórki rosną w zawiesinie (e.g., CHO cells)
Tempo wzrostu bardzo wysokie(e.g., HeLa cells)
Ekspresja zewnątrzkomórkowa
Wady Niektóre komórki rosną
przytwierdzone do podłoża
Skomplikowana hodowla
Wolny wzrost niektórych linii komórkowych (15-25h)
Bardzo wrażliwe na kontaminację
Bardzo wrażliwe na wstrząsy

Hodowla komórek roślinnych
Zalety
Hodowla w zawiesinie
Małe wymagania
napowietrzania
Białka w wakuolach
Wady
Mała ilość systemów
Agrobacterium
Specyficzne wirusy
Powolny wzrost
Wrażliwe na wstrząsy

Inne systemy
Zwierzęta
Transformacja zarodków
Użycie tkankowo-specyficznych promotorów (np. produkcja
w mleku)

ESCHERICHIA COLI

Plazmid - definicja
Plazmidy – autonomiczne, pozachromosomalne elementy genetyczne występujące (zwykle w postaci kolistych, lub rzadziej w postaci liniowych cząsteczek DNA) w bardzo wielu organizmach prokariotycznych (i niektórych eukariotycznych)

Plazmidy - charakterystyczne cechy
Zdolność do trwałego utrzymywania się w komórce
Zdolność replikowania się w komórce w kontrolowany sposób
Plazmidy nie kodują funkcji, które byłyby niezbędne do życia komórki (w „normalnych” warunkach)

Wybór systemów ekspresyjnych w E. coli
Zalety:wysoki poziom ekspresji (do 500 mg/l hodowli)dobrze poznany systemproste technologieniskie koszty
Wady:brak modyfikacji posttranslacyjnychograniczona wielkość klonowanego DNAróżnice w „codon usage”

Wektory prokariotyczne
Zawierają tzw. markery selekcyjne, nadające komórkom gospodarza łatwy
do testowania fenotyp (np. oporność na antybiotyk)
Posiadają unikalne miejsca rozpoznawane przez enzymy restrykcyjne w
obrębie rejonów, w które można wprowadzić obcy DNA, bez naruszania
zdolności wektora do replikacji i możliwości selekcji zrekombinowanych
cząstek
Są niezdolne do przeżycia poza komórką gospodarza, namnażają się w
niewielu szczepach i nie posiadają zdolności koniugacyjnych

Wektory prokariotyczne - podział
Wektory autonomiczne – autonomicznie replikują się w
komórkach gospodarza
Wektory integracyjne – włączają się do genomu gospodarza
(duża stabilność, mała liczba kopii)

Wektory ekspresyjne - podział
Wektory ekspresyjne z silnym promotorem - jeżeli klonowany
gen nie posiada efektywnego promotora, ale zawiera własny
rejon RBS to włączany jest do wektora tuż za silnym
promotorem (fuzja genowa transkrypcyjna)

Wektory ekspresyjne
Wektory ekspresyjne z sygnałami translacyjnymi – konieczność
modyfikacji sekwencji genu w miejscu kodonu inicjatorowego
translacji (ATG) (fuzja genowa translacyjna)

Wektory ekspresyjne
Wektory ekspresyjne umożliwiające produkcję białek
hybrydowych – białka hybrydowe z dodatkowymi
aminokwasami w części N-terminalnej (fuzje proksymalne) lub
w części C-terminalnej (fuzje dystalne) (fuzja translacyjnie
złożona)

Wybór systemu
Wybór odpowiedniego systemu
stopień tolerancji komórek gospodarza na obecność obcego białka
zasada: siła promotora determinuje możliwy do osiągnięcia stan jego
represji (słabszy promotor jest ściślej kontrolowany, natomiast silny
promotor jest gorzej regulowany)
Wybór odpowiedniego wektora:
ilość kopii wektora na komórkę
promotory bakteriofagowe (PR i PL z faga λ oraz Φ10 z faga T7), promotory
bakteryjne (PLAC) oraz eukariotyczne (PCMV)

Składniki systemu ekspresyjnego
DNA kodujace białko docelowe mRNA
cDNA
genomowe DNA
Wektor (plazmid) pET
pQE
pGEX
Szczep ekspresyjny (E. coli ) BL21(DE3)
Rosetta(DE3) plus pRARE
Tuner(DE3)
ADA494(DE3)trxB mutant
Origami(DE3) trxB and gor mutations

System nadprodukcji białek bakteriofaga T7
(Tabor-Studier)
genom E.coli Komórka gospodarza
pET
gen kodujący lizozym
lizozym T7
pLysS
Inaktywacja
polimeraza RNA T7
DE3
gen T7
lac Opromotor lac
represor lacI
gen kodujący represor
lacI
polimeraza RNA E.coli
Indukcja IPTG
represor lacI
gen kodujący represor
lacI
lac Opromotor T7
polimeraza RNA T7
Indukcja IPTG
gen docelowy
http://www.pg.gda.pl/chem/Katedry/Mikrobiologia/inzgen.html

System pET (Novagen)
Możliwość klonowania genów, które chcemy eksprymować w E. coli, pod silny promotor bakeriofaga T7, który nie jest rozpoznawany przez polimerazę RNA E. coli
Brak ekspresji genów spod promotora T7 w komórkach E. colidopóki nie pojawi się w nich polimeraza RNA faga T7

System pET –części składowe
Plazmidowy wektor serii pET umożliwiający ekspresjęwklonowanego do niego genu pod kontrolą silnychsygnałów transkrypcyjnych (promotor) i translacyjnych(RBS) pochodzących z bakteriofaga T7
Szczep E. coli pozwalający na indukcję na ściślekontrolowanym poziomie polimerazy RNA bakteriofaga T7spod wybranego promotora E. coli np. pLacUV5
Induktor IPTG regulujący poziom ekspresji genu z plazmidupET w komórkach E. coli

Wektory ekspresyjne serii pET (Novagen)

Wektory ekspresyjne serii pET - wektory
transkrypcyjne
Dostępne trzy wektory transkrypcyjne: pET21(+), pET23(+) ipET24(+) umożliwiają silną transkrypcję mRNA wklonowanegogenu z promotora T7
Brak sekwencji RBS w obrębie transkryptu powstającego spodpromotora T7
W celu uzyskania białka z genu wklonowanego do tychwektorów należy klonować go z własną sekwencją RBSumożliwiającą translację w E. coli


Wektory ekspresyjne serii pET - wektory
translacyjne
Umożliwiają silną transkrypcję mRNA wklonowanego genu zpromotora T7
Posiadają sekwencje RBS w obrębie transkryptu powstającegospod promotora T7 zapewniającą wydajną translacje
Większość zawiera miejsca restrykcyjne pozwalająceklonować w jedną z trzech możliwych ramek odczytu spodRBS znajdującego się na wektorze


Wektory ekspresyjne rodziny pET
Większość wektorów serii pET poza typowym ori replikacyjnym zawiera również sekwencję origin replikacji faga f1 umożliwiającą uzyskanie jednoniciowej formy DNA tych plazmidów np. w celu przeprowadzenia mutagenezy ukierunkowanej w obrębie ich sekwencji
Wszystkie wektory z origin f1 zawierają w nazwie sufix (+)

pET System – trzy podstawowe kryteria
wyboru wektora ekspresyjnego
1. Ilość białka, którą chcemy wyprodukować:
„małe” ilości białka (techniki badania aktywności białek,
badania oddziaływania uzyskanego białka z wybranymi
ligandami, uzyskiwanie przeciwciał przeciwko tym białkom)
„duże” ilości białka (uzyskanie stężonych lub wysoce
aktywnych preparatów – badania strukturalne, zastosowania
przemysłowe)

pET System – 3 podstawowe kryteria
wyboru wektora ekspresyjnego
2. Możliwość lub konieczność uzyskania białka w postaci fuzji z wybranymi domenami fuzyjnymi mającemu na celu np.:
• ułatwienie procedury oczyszczania eksprymowanego białka
• zwiększenie rozpuszczalności uzyskanego białka w cytozolu E. coli
• transport eksprymowanego białka do przestrzeni periplazmatycznej E. coli

pET System – 3 podstawowe kryteria
wyboru wektora ekspresyjnego
3. Strategia klonowania czyli wybór wektora pod
względem miejsc restrykcyjnych obecnych w MCS
oraz pod względem wyboru jednej z trzech
możliwych ramek odczytu

pET System – trzy podstawowe kryteria
wyboru wektora ekspresyjnego
Jeżeli możliwość klonowania do wektorów
ekspresyjnych jest ograniczona można zastosować
„klonowanie bez ligazy” (wektory LIC serii pET np.
pET30LIC)

Klonowanie bez użycia ligazy

Domeny fuzyjne- różnorodność wektorów
plazmidowych pET
Domeny fuzyjne („tag”) umożliwiają ekspresję tego samego genu w postaci białek fuzyjnych różniących się właściwościami wynikającymi z ich sekwencji
Domeny fuzyjne mogą składać się z kilku reszt aminokwasowych np. 6 reszt aminokwasowych His-Tag lub mogą być to sekwencje reszt aminokwasowych białek np. 495 reszt aa. białka NusA z Nus-Tag

Domeny fuzyjne - różnorodność
wektorów plazmidowych pET
Domeny fuzyjne umożliwiające łatwą detekcję białek fuzyjnych za pomocą techniki Western blotting:
S•Tag, T7•Tag, GST•Tag, His•Tag, Nus•Tag, HSV•Tag
Domeny fuzyjne umożliwiające „proste” oszacowanie poziomu ekspresji białka nawet w surowych ekstraktach komórkowych:
S•Tag, GST•Tag

Domeny fuzyjne - różnorodność
wektorów plazmidowych pET
Domeny fuzyjne umożliwiające łatwe oczyszczanie
białek fuzyjnych za pomocą różnego typu
chromatografii powinowactwa:
His•Tag, GST•Tag, S•Tag, T7•Tag
Domeny fuzyjne umożliwiające transport
eksprymowanego białka do przestrzeni
periplazmatycznej E. coli np.
sekwencja sygnalna ompT/pelB w wektorach pET12a-c


Domeny fuzyjne - różnorodność
wektorów plazmidowych pET
Sekwencje rozpoznania dla wybranych enzymów
proteolitycznych umożliwiające łatwe odcięcie
domeny fuzyjnej po wykorzystaniu ich w oczyszczaniu
docelowego białka typu dzikiego za pomocą wybranej
metody chromatografii powinowactwa:
LeuValProArg*GlySer – trombina
AspAspAspAspLys* - enterokinaza
IleGluGlyArg* - czynnik X
* - miejsce cięcia

Domeny fuzyjne - różnorodność
wektorów plazmidowych pET
Domeny fuzyjne zwiększające rozpuszczalność białek
fuzyjnych i ułatwiające powstawanie właściwych
mostków disiarczkowych:
W cytoplazmie szczepów E. coli mutantów trxB- - Nus•Tag,
Trx•Tag i GST•Tag
W przestrzeni periplazmatycznej E. coli oraz wspomagające
„folding” białek fuzyjnych
DsbA•Tag
DsbC•Tag

Domeny fuzyjne - różnorodność
wektorów plazmidowych pET
Domena fuzyjna zmniejszająca rozpuszczalność białek fuzyjnych
i ułatwiające powstawanie ciał inkluzyjnych tzw „kamień
białkowy”:
KSI-Tag
silnie eksprymowana silnie hydrofobowa domena fuzyjna zbudowana
ze 125 aa
użyteczna także przy ekspresji małych białek a nawet peptydów
zabezpieczająca je przed proteolizą w cytoplazmie

His•Tag
Sekwencja domeny fuzyjnej
Ligand wiążący domenę
fuzyjną:
Elutant:
• Zbuforowane roztwory imidazolu (pH=7,9)
• Poprzez powolne obniżanie pH
Np. HisHisHisHisHisHisMetXXXX

His•Tag
Zalety Możliwość oczyszczania
białkaMożliwość oczyszczania
białka w warunkach denaturujących (6M Gu-HCl lub 6M mocznik)
Wady Należy unikać buforów
zawierających związki: redukujące takie jak: -merkaptoetanol, ditiotreitol oraz chelatujące tj. EDTA
www.invitrogen.com

Plazmidy serii pETBlue
Powodem skonstruowania plazmidów serii pETBlue była
chęć połączenia zalet plazmidów serii pET tj.
wysokokopijność i ekspresja wklonowanych genów spod
silnego promotora T7 z prostą selekcją
rekombinantowych klonów E. coli opartą o znany z
plazmidów pUC18/19 system selekcji białe/niebieskie
kolonie na podłożach z X-Gal i IPTG

Plazmidy serii pETBlue oraz pET
Plazmidy serii pET
Plazmidy serii pETBlue
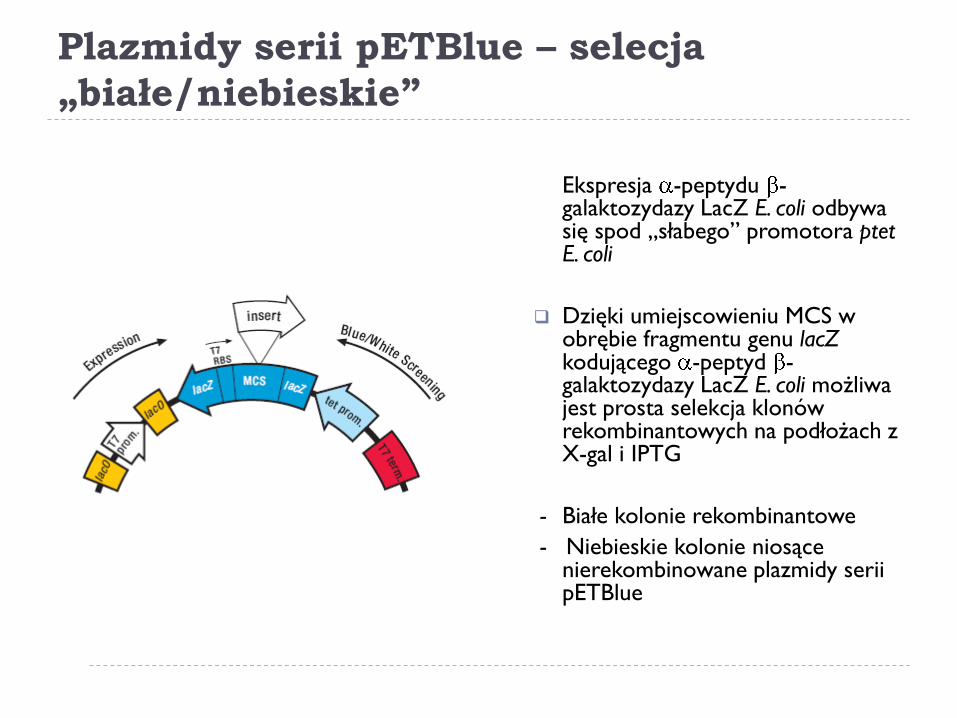
Plazmidy serii pETBlue – selecja
„białe/niebieskie”
Ekspresja -peptydu -galaktozydazy LacZ E. coli odbywa się spod „słabego” promotora ptet E. coli
Dzięki umiejscowieniu MCS w obrębie fragmentu genu lacZ kodującego -peptyd -galaktozydazy LacZ E. coli możliwa jest prosta selekcja klonów rekombinantowych na podłożach z X-gal i IPTG
- Białe kolonie rekombinantowe
- Niebieskie kolonie niosące nierekombinowane plazmidy serii pETBlue

Plazmidy serii pETBlue – selecja
„białe/niebieskie”
Selekcja „białe/niebieskie” dla plazmidów pETBlue jest możliwa do przeprowadzenia tylko w szczepach E. coli, które nie produkują natywnej beta-galaktozydazy E. coli

Plazmidy serii pETBlue – selekcja
„białe/niebieskie”

Plazmidy serii pETBlue – ekspresja spod
promotora T7
W plazmidach serii pETBlue
znajdują się dwie sekwencje
operatorowe lacO (miejsca
wiązania dla represora lacI)

Plazmidy serii pETBlue – ekspresja spod promotora T7
Plazmidy serii pETBlue
nie niosą własnej kopii
genu lacI represora (w
przeciwieństwie do
plazmidów serii pET)

Plazmidy serii pETBlue – ekspresja spod
promotora T7
Plazmidy serii pETBlue są wykorzystywane do ekspresji białek w warunkach ściślejszej kontroli represorem lacI niż ma to miejsce w przypadku plazmidów serii pET

Szczepy ekspresyjne E. coli dla plazmidów
pETBlue
www.novagen.com

Szczepy ekspresyjne E. coli dla plazmidów
pETBlue
www.novagen.com

Szczepy ekspresyjne E. coli dla plazmidów
pETBlue
Istniej możliwość prowadzenia ekspresji w komórkach E.
coli NovaBlue transformowanych plazmidem serii pETBlue
po ich zakażeniu fagiem CE6

Plazmidy serii pETBlue
www.novagen.com

Koekspresja dwóch i więcej genów w
systemie pET
Koekspresja dwóch (lub więcej genów) w E. coli może
mieć na celu np.:
Wyprodukowanie dużej liczby multimerycznych kompleksów
białek enzymatycznych
Produkcję określonych białek wraz ze specyficznymi dla nich
białkami opiekuńczymi

Koekspresja dwóch i więcej genów w
systemie pET
Produkcja multi-podjednostowych kompleksów białek służy:
Badaniu oddziaływań pomiędzy poszczególnymi podjednostkami
kompleksu
Badaniu właściwości enzymatycznych multimerycznego kompleksu
enzymatycznego
Koekspresja z białkiem opiekuńczym jest często jedyną drogą
otrzymania niektórych białek w postaci natywnej z
zachowaniem ich „pierwotnej” funkcji

Koekspresja dwóch i więcej genów w
systemie pET
Koekspresja dwóch i więcej genów w E. coli może być
osiągnięta:
Poprzez wklonowanie dwóch lub więcej genów do
jednego wektora plazmidowego
Poprzez transformację komórek E. coli dwoma lub więcej
plazmidami posiadającymi kompatybilne ori replikacji oraz
niosącymi różne geny oporności na antybiotyki

Koekspresja dwóch i więcej genów w
systemie pET

Koekspresja dwóch i więcej genów w systemie
pET

Koekspresja dwóch i więcej genów w
systemie pET
Istotny jest odpowiedni wybór wektora i szczepu ze
względu na kompatybilność ori replikacji i genów
oporności na antybiotyki wektorów (oporność „własna”
szczepów ekspresyjnych na antybiotyki)

Koekspresja dwóch i więcej genów w
systemie pET
DuetSystem System składa się z czterech wektorów plazmidowych
umożliwiających jednoczesną ekspresję nawet ośmiu różnych białek w E. coli
Każdy z tych wektorów pozwala na jednoczesną ekspresję do dwóch różnych genów spod niezależnych promotorów T7lac
Wektory te umożliwiają ekspresję wklonowanych białek w postaci natywnej lub fuzji z His-Tag lub S-Tag w celu detekcji i łatwego oczyszczania powstałych białek fuzyjnych

Koekspresja dwóch i więcej genów w
systemie pET
Duet System
System ten pozwala na stosowanie innych wektorów serii
pET, pETBlue, pETcoco wymiennie z poszczególnymi
wektorami Duet System



![Strona główna - Wojskowa Akademia Techniczna4)130.pdf · tion, linking two main liquid crystal (LC) technologies [1 ]. The first one was used in conventional LCD and regarded the](https://static.fdocuments.pl/doc/165x107/5f447ca5d1f4361da6102dc7/strona-gwna-wojskowa-akademia-techniczna-4130pdf-tion-linking-two-main.jpg)



![The Effect of Whole-Body Cryotherapy at Different ...downloads.hindawi.com/journals/omcl/2018/2157496.pdf · generally accompanies increased levels of oxidative stress markers [5].](https://static.fdocuments.pl/doc/165x107/5f87f69f5e09f21917510f9c/the-effect-of-whole-body-cryotherapy-at-different-generally-accompanies-increased.jpg)




