
Rola mikroRNA w komórkach śródbłonka — regulacja ... · Rola mikroRNA w komórkach...
10
Postępy Biochemii 59 (4) 2013 405 Witold Norbert Nowak 1,2 Urszula Florczyk 1 Alicja Józkowicz 1 Józef Dulak 1, 1 Zakład Biotechnologii Medycznej, Wy- dział Biochemii, Biofizyki i Biotechno- logii, Uniwersytet Jagielloński, Kraków 2 Jagiellońskie Centrum Rozwoju Leków, Uni- wersytet Jagielloński, Kraków Zakład Biotechnologii Medycznej, Wydział Biochemii, Biofizyki i Biotechnologii, Uniwersytet Jagielloński, ul. Gronostajowa 7, 30-387 Kraków; tel.: (12) 66 46 375, faks: (12) 66 46 918, e-mail: [email protected] Artykuł otrzymano 5 listopada 2013 r. Artykuł zaakceptowano 6 listopada 2013 r. Słowa kluczowe: mikroRNA, śródbłonek, an- giogeneza, różnicowanie, zarodkowe komórki macierzyste Wykaz skrótów: CD (ang. cluster of differentia- tion) — antygen różnicowania komórkowego; DGCR8 — (ang. DiGeorge syndrome critical re- gion 8); ESC (ang. embryonic stem cells) — za- rodkowe komórki macierzyste; EGF (ang. epi- dermal growth factor) — nabłonkowy czynnik wzrostu; FGF-2 (ang. fibroblast growth factor-2) — czynnik wzrostu fibroblastów 2; HIF (ang. hypoxia inducible factor) — czynnik indukowa- ny przez hipoksję; HMEC (ang. human micro- vascular endothelial cells) — ludzkie mikrowa- skularne komórki śródbłonka; HUVEC (ang. human umbilical vein endothelial cells) — ludzkie komórki śródbłonka żyły pępowinowej; IGF-1 (ang. insulin-like growth factor-1) — insulinopo- dobny czynnik wzrostu 1; iPSC (ang. induced pluripotent stem cells) – indukowane pluripo- tencjalne komórki macierzyste; miR — mi- kroRNA; PMA — octan mirystynianu forbo- lu; PDGF (ang. platelet-derived growth factor) — płytkopochodny czynnik wzrostu; shRNA (ang. small/short hairpin RNA) — krótkie RNA o strukturze spinki do włosów; VEGF (ang. vascular endothelial growth factor) — czynnik wzrostu śródbłonka naczyniowego Podziękowania: Badania autorów artykułu były ze środków przeznaczonych na realizację projektu POIG 01.01.02-069/09, NCN Opus 2012/07/B/NZ1/02881 oraz NCN Harmonia 2012/06/M/NZ1/00008. Rola mikroRNA w komórkach śródbłonka — regulacja różnicowania i angiogenezy STRESZCZENIE M odyfikacja różnicowania i funkcji śródbłonka, w kontekście jego wpływu na procesy regeneracji tkanek, rozwoju miażdżycy czy wzrostu nowotworów, stanowi atrakcyjny cel terapeutyczny. Czynnikami zdolnymi w sposób plejotropowy, a jednocześnie precyzyjny regulować całe szlaki sygnałowe, są mikroRNA, krótkie cząsteczki RNA, które rozpoznają i hamują ekspresję określonych transkryptów. Liczne badania pozwoliły na scharakteryzo- wanie mikroRNA o działaniu pobudzającym lub hamującym powstawanie nowych naczyń krwionośnych, regulującym ekspresję genów białek adhezyjnych czy syntazy tlenku azo- tu i modulującym w ten sposób funkcje komórek śródbłonka. Ponadto, mikroRNA mogą wpływać na różnicowanie zarodkowych oraz indukowanych pluripotencjalnych komórek macierzystych do komórek śródbłonkowych oraz na specjalizację dojrzewających komórek w kierunku śródbłonków typowych dla poszczególnych odcinków łożyska naczyniowego. Prezentowany artykuł opisuje wpływ mikroRNA na własności angiogenne i różnicowanie komórek śródbłonkowych. WPROWADZENIE Śródbłonek naczyniowy jest pojedynczą warstwą komórek wyścielających naczynia krwionośne. U dorosłych ludzi całkowita powierzchnia śródbłonka może stanowić około 350 m 2 [1]. Jednocześnie, komórki śródbłonka ulegają podziałom stosunkowo rzadko [2]. Główną rolą śródbłonka jest gwarantowanie stabilnego przepływu krwi w obrębie naczyń krwionośnych. Komórki śródbłonkowe odpowiedzialne są między innymi za utrzymanie odpowiedniego napięcia naczyniowego, re- gulację aktywności płytek krwi i odpowiedzi zapalnej oraz za inicjację two- rzenia nowych naczyń krwionośnych (angiogenezę) [3]. Angiogeneza odgry- wa ważną rolę w gojeniu ran [4], ale również we wzroście nowotworów [5], oba te procesy są spowolnione w przypadku niedostatecznego ukrwienia. Dlatego zrozumienie regulacji procesów związanych z funkcjami śródbłon- ka jest kluczowe dla opracowania nowych metod leczenia nie tylko chorób naczyniowych, ale także hamowania rozwoju nowotworów czy poprawy re- generacji uszkodzonych tkanek. Ponadto, łatwość uzyskania indukowanych komórek pluripotencjalnych (iPSC) otwiera możliwość uzyskania komórek śródbłonka w procesie różnicowania in vitro. Pozwala to na zbadanie mecha- nizmów powstawania komórek śródbłonka oraz wykorzystania tej wiedzy w medycynie regeneracyjnej. Funkcje śródbłonka regulowane są przez roz- liczne szlaki sygnałowe, na które wpływ mają także krótkie cząsteczki RNA, mikroRNA. MikroRNA są niekodującymi cząsteczkami RNA o długości około 22 nukleotydów. Główną funkcją mikroRNA jest potranskrypcyjna regulacja mRNA. Synteza oraz dojrzewanie mikroRNA jest procesem wieloetapowym, który zachodzi w jądrze komórkowym, a następnie cytoplazmie. Sekwencje mikroRNA kodowane są w obrębie ich własnych genów, często policistro- nowych, posiadających samodzielne promotory, lub w obrębie intronów lub egzonów innych genów. Transkrypcja genów mikroRNA, w wyniku której powstaje pri-mikroR- NA, prowadzona jest zazwyczaj przez polimerazę RNA II. Cząsteczki pri- -mikroRNA o długości około 70 nukleotydów i strukturze spinki do włosów ulegają intensywnej obróbce, tzw. dojrzewaniu. Na początku, cząsteczki pri- -mikroRNA są rozpoznawane przez białko jądrowe DGCR8 (Pasha) związa- ne z enzymem Drosha (tworzących razem tzw. kompleks mikroprocesora) przecinającym pierwotny transkrypt i uwalniającym struktury spinek, które stanowią pre-mikroRNA. W przypadku mikroRNA powstających z intro- nów innych genów tzw. mirtronów, obróbka pierwotnego transkryptu za
-
Upload
nguyenhuong -
Category
Documents
-
view
218 -
download
0
Transcript of Rola mikroRNA w komórkach śródbłonka — regulacja ... · Rola mikroRNA w komórkach...

Postępy Biochemii 59 (4) 2013 405
Witold Norbert Nowak1,2
Urszula Florczyk1
Alicja Józkowicz1
Józef Dulak1,
1Zakład Biotechnologii Medycznej, Wy-dział Biochemii, Biofizyki i Biotechno-logii, Uniwersytet Jagielloński, Kraków 2Jagiellońskie Centrum Rozwoju Leków, Uni-wersytet Jagielloński, Kraków
Zakład Biotechnologii Medycznej, Wydział Biochemii, Biofizyki i Biotechnologii, Uniwersytet Jagielloński, ul. Gronostajowa 7, 30-387 Kraków; tel.: (12) 66 46 375, faks: (12) 66 46 918, e-mail: [email protected]
Artykuł otrzymano 5 listopada 2013 r.Artykuł zaakceptowano 6 listopada 2013 r.
Słowa kluczowe: mikroRNA, śródbłonek, an-giogeneza, różnicowanie, zarodkowe komórki macierzyste
Wykaz skrótów: CD (ang. cluster of differentia-tion) — antygen różnicowania komórkowego; DGCR8 — (ang. DiGeorge syndrome critical re-gion 8); ESC (ang. embryonic stem cells) — za-rodkowe komórki macierzyste; EGF (ang. epi-dermal growth factor) — nabłonkowy czynnik wzrostu; FGF-2 (ang. fibroblast growth factor-2) — czynnik wzrostu fibroblastów 2; HIF (ang. hypoxia inducible factor) — czynnik indukowa-ny przez hipoksję; HMEC (ang. human micro-vascular endothelial cells) — ludzkie mikrowa-skularne komórki śródbłonka; HUVEC (ang. human umbilical vein endothelial cells) — ludzkie komórki śródbłonka żyły pępowinowej; IGF-1 (ang. insulin-like growth factor-1) — insulinopo-dobny czynnik wzrostu 1; iPSC (ang. induced pluripotent stem cells) – indukowane pluripo-tencjalne komórki macierzyste; miR — mi-kroRNA; PMA — octan mirystynianu forbo-lu; PDGF (ang. platelet-derived growth factor) — płytkopochodny czynnik wzrostu; shRNA (ang. small/short hairpin RNA) — krótkie RNA o strukturze spinki do włosów; VEGF (ang. vascular endothelial growth factor) — czynnik wzrostu śródbłonka naczyniowego
Podziękowania: Badania autorów artykułu były ze środków przeznaczonych na realizację projektu POIG 01.01.02-069/09, NCN Opus 2012/07/B/NZ1/02881 oraz NCN Harmonia 2012/06/M/NZ1/00008.
Rola mikroRNA w komórkach śródbłonka — regulacja różnicowania i angiogenezy
STRESZCZENIE
Modyfikacja różnicowania i funkcji śródbłonka, w kontekście jego wpływu na procesy regeneracji tkanek, rozwoju miażdżycy czy wzrostu nowotworów, stanowi atrakcyjny
cel terapeutyczny. Czynnikami zdolnymi w sposób plejotropowy, a jednocześnie precyzyjny regulować całe szlaki sygnałowe, są mikroRNA, krótkie cząsteczki RNA, które rozpoznają i hamują ekspresję określonych transkryptów. Liczne badania pozwoliły na scharakteryzo-wanie mikroRNA o działaniu pobudzającym lub hamującym powstawanie nowych naczyń krwionośnych, regulującym ekspresję genów białek adhezyjnych czy syntazy tlenku azo-tu i modulującym w ten sposób funkcje komórek śródbłonka. Ponadto, mikroRNA mogą wpływać na różnicowanie zarodkowych oraz indukowanych pluripotencjalnych komórek macierzystych do komórek śródbłonkowych oraz na specjalizację dojrzewających komórek w kierunku śródbłonków typowych dla poszczególnych odcinków łożyska naczyniowego. Prezentowany artykuł opisuje wpływ mikroRNA na własności angiogenne i różnicowanie komórek śródbłonkowych.
WPROWADZENIE
Śródbłonek naczyniowy jest pojedynczą warstwą komórek wyścielających naczynia krwionośne. U dorosłych ludzi całkowita powierzchnia śródbłonka może stanowić około 350 m2 [1]. Jednocześnie, komórki śródbłonka ulegają podziałom stosunkowo rzadko [2].
Główną rolą śródbłonka jest gwarantowanie stabilnego przepływu krwi w obrębie naczyń krwionośnych. Komórki śródbłonkowe odpowiedzialne są między innymi za utrzymanie odpowiedniego napięcia naczyniowego, re-gulację aktywności płytek krwi i odpowiedzi zapalnej oraz za inicjację two-rzenia nowych naczyń krwionośnych (angiogenezę) [3]. Angiogeneza odgry-wa ważną rolę w gojeniu ran [4], ale również we wzroście nowotworów [5], oba te procesy są spowolnione w przypadku niedostatecznego ukrwienia. Dlatego zrozumienie regulacji procesów związanych z funkcjami śródbłon-ka jest kluczowe dla opracowania nowych metod leczenia nie tylko chorób naczyniowych, ale także hamowania rozwoju nowotworów czy poprawy re-generacji uszkodzonych tkanek. Ponadto, łatwość uzyskania indukowanych komórek pluripotencjalnych (iPSC) otwiera możliwość uzyskania komórek śródbłonka w procesie różnicowania in vitro. Pozwala to na zbadanie mecha-nizmów powstawania komórek śródbłonka oraz wykorzystania tej wiedzy w medycynie regeneracyjnej. Funkcje śródbłonka regulowane są przez roz-liczne szlaki sygnałowe, na które wpływ mają także krótkie cząsteczki RNA, mikroRNA.
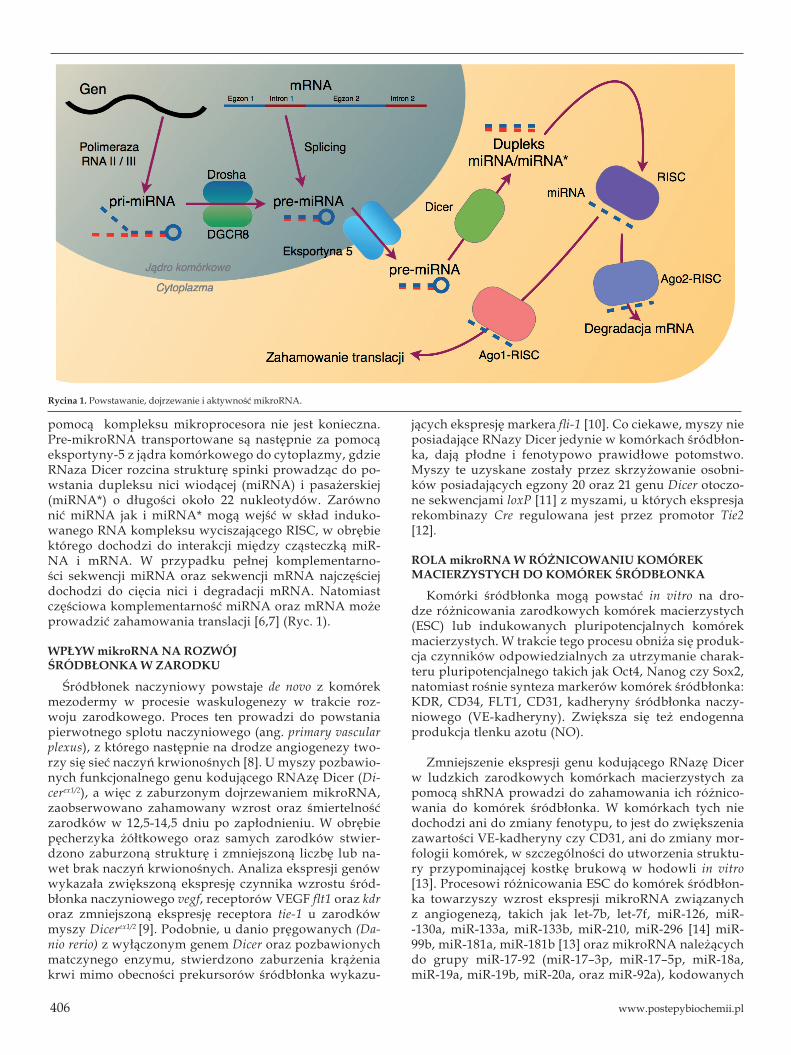
MikroRNA są niekodującymi cząsteczkami RNA o długości około 22 nukleotydów. Główną funkcją mikroRNA jest potranskrypcyjna regulacja mRNA. Synteza oraz dojrzewanie mikroRNA jest procesem wieloetapowym, który zachodzi w jądrze komórkowym, a następnie cytoplazmie. Sekwencje mikroRNA kodowane są w obrębie ich własnych genów, często policistro-nowych, posiadających samodzielne promotory, lub w obrębie intronów lub egzonów innych genów.
Transkrypcja genów mikroRNA, w wyniku której powstaje pri-mikroR-NA, prowadzona jest zazwyczaj przez polimerazę RNA II. Cząsteczki pri--mikroRNA o długości około 70 nukleotydów i strukturze spinki do włosów ulegają intensywnej obróbce, tzw. dojrzewaniu. Na początku, cząsteczki pri--mikroRNA są rozpoznawane przez białko jądrowe DGCR8 (Pasha) związa-ne z enzymem Drosha (tworzących razem tzw. kompleks mikroprocesora) przecinającym pierwotny transkrypt i uwalniającym struktury spinek, które stanowią pre-mikroRNA. W przypadku mikroRNA powstających z intro-nów innych genów tzw. mirtronów, obróbka pierwotnego transkryptu za
406 www.postepybiochemii.pl
pomocą kompleksu mikroprocesora nie jest konieczna. Pre-mikroRNA transportowane są następnie za pomocą eksportyny-5 z jądra komórkowego do cytoplazmy, gdzie RNaza Dicer rozcina strukturę spinki prowadząc do po-wstania dupleksu nici wiodącej (miRNA) i pasażerskiej (miRNA*) o długości około 22 nukleotydów. Zarówno nić miRNA jak i miRNA* mogą wejść w skład induko-wanego RNA kompleksu wyciszającego RISC, w obrębie którego dochodzi do interakcji między cząsteczką miR-NA i mRNA. W przypadku pełnej komplementarno-ści sekwencji miRNA oraz sekwencji mRNA najczęściej dochodzi do cięcia nici i degradacji mRNA. Natomiast częściowa komplementarność miRNA oraz mRNA może prowadzić zahamowania translacji [6,7] (Ryc. 1).
WPŁYW mikroRNA NA ROZWÓJ ŚRÓDBŁONKA W ZARODKU
Śródbłonek naczyniowy powstaje de novo z komórek mezodermy w procesie waskulogenezy w trakcie roz-woju zarodkowego. Proces ten prowadzi do powstania pierwotnego splotu naczyniowego (ang. primary vascular plexus), z którego następnie na drodze angiogenezy two-rzy się sieć naczyń krwionośnych [8]. U myszy pozbawio-nych funkcjonalnego genu kodującego RNAzę Dicer (Di-cerex1/2), a więc z zaburzonym dojrzewaniem mikroRNA, zaobserwowano zahamowany wzrost oraz śmiertelność zarodków w 12,5-14,5 dniu po zapłodnieniu. W obrębie pęcherzyka żółtkowego oraz samych zarodków stwier-dzono zaburzoną strukturę i zmniejszoną liczbę lub na-wet brak naczyń krwionośnych. Analiza ekspresji genów wykazała zwiększoną ekspresję czynnika wzrostu śród-błonka naczyniowego vegf, receptorów VEGF flt1 oraz kdr oraz zmniejszoną ekspresję receptora tie-1 u zarodków myszy Dicerex1/2 [9]. Podobnie, u danio pręgowanych (Da-nio rerio) z wyłączonym genem Dicer oraz pozbawionych matczynego enzymu, stwierdzono zaburzenia krążenia krwi mimo obecności prekursorów śródbłonka wykazu-
jących ekspresję markera fli-1 [10]. Co ciekawe, myszy nie posiadające RNazy Dicer jedynie w komórkach śródbłon-ka, dają płodne i fenotypowo prawidłowe potomstwo. Myszy te uzyskane zostały przez skrzyżowanie osobni-ków posiadających egzony 20 oraz 21 genu Dicer otoczo-ne sekwencjami loxP [11] z myszami, u których ekspresja rekombinazy Cre regulowana jest przez promotor Tie2 [12].
ROLA mikroRNA W RÓŻNICOWANIU KOMÓREK MACIERZYSTYCH DO KOMÓREK ŚRÓDBŁONKA
Komórki śródbłonka mogą powstać in vitro na dro-dze różnicowania zarodkowych komórek macierzystych (ESC) lub indukowanych pluripotencjalnych komórek macierzystych. W trakcie tego procesu obniża się produk-cja czynników odpowiedzialnych za utrzymanie charak-teru pluripotencjalnego takich jak Oct4, Nanog czy Sox2, natomiast rośnie synteza markerów komórek śródbłonka: KDR, CD34, FLT1, CD31, kadheryny śródbłonka naczy-niowego (VE-kadheryny). Zwiększa się też endogenna produkcja tlenku azotu (NO).
Zmniejszenie ekspresji genu kodującego RNazę Dicer w ludzkich zarodkowych komórkach macierzystych za pomocą shRNA prowadzi do zahamowania ich różnico-wania do komórek śródbłonka. W komórkach tych nie dochodzi ani do zmiany fenotypu, to jest do zwiększenia zawartości VE-kadheryny czy CD31, ani do zmiany mor-fologii komórek, w szczególności do utworzenia struktu-ry przypominającej kostkę brukową w hodowli in vitro [13]. Procesowi różnicowania ESC do komórek śródbłon-ka towarzyszy wzrost ekspresji mikroRNA związanych z angiogenezą, takich jak let-7b, let-7f, miR-126, miR--130a, miR-133a, miR-133b, miR-210, miR-296 [14] miR-99b, miR-181a, miR-181b [13] oraz mikroRNA należących do grupy miR-17-92 (miR-17–3p, miR-17–5p, miR-18a, miR-19a, miR-19b, miR-20a, oraz miR-92a), kodowanych
Rycina 1. Powstawanie, dojrzewanie i aktywność mikroRNA.

Postępy Biochemii 59 (4) 2013 407
przez jeden policistronowy gen [15]. Ekspresja miR-17, miR-18, miR-19 oraz miR-20 zwiększa się w trakcie róż-nicowania i osiąga najwyższy poziom siódmego dnia. Co ciekawe, ekspresja miR-92a podobnie jak pri-miR-17-92 ulega zmniejszeniu. Jednocześnie, zahamowanie w za-rodkowych komórkach macierzystych zarówno ekspresji poszczególnych mikroRNA należących do grupy miR-17-92 jak i wszystkich jej składowych, nie wpływa ani na charakter pluripotencjalny ESC ani na ich różnicowanie do śródbłonka [15]. Zwiększenie ekspresji miR-99b, miR--181a, miR-181b powoduje natomiast wzrost poziomu CD31 oraz VE-kadheryny oraz produkcji tlenku azotu w różnicowanych zarodkowych komórkach macierzystych. Z drugiej, strony obniżenie ekspresji miR-99b, miR-181a, miR-181b nie wpływa na poziom markerów śródbłonko-wych, natomiast zmniejsza produkcję NO przez komórki śródbłonka wywodzące się z komórek ESC [13].
Co ciekawe, mikroRNA regulują także różnicowanie komórek śródbłonka wywodzących się z zarodkowych komórek macierzystych do poszczególnych podtypów śródbłonka naczyniowego. Zwiększona ekspresja miR--99b prowadzi do podniesienia ekspresji markera śród-błonka tętniczego Hey2 oraz limfatycznego FLT4, przy jednoczesnym obniżeniu ekspresji podoplaniny, mar-kera śródbłonka naczyń limfatycznych. Co zaskakujące, również obniżenie ekspresji miR-99b zwiększa ekspresję Hey2 i Jagged-1 (Jag-1, typowych dla śródbłonka tętni-czego) jednocześnie jednak podnosi poziom podoplani-ny i zmniejsza ekspresję markerów śródbłonka żylnego, takich jak ephB4 i NR2F2. Z kolei nadekspresja miR-181a prowadzi do podniesienia poziomu transkryptów cha-rakterystycznych dla komórek śródbłonkowych wyście-łających tętnice i naczynia limfatyczne, zaś jego wycisze-nie zwiększa poziom markera limfatycznego Prox1 i ha-muje ekspresję efryny B2 typowej dla tętnic. Zwiększony poziom miR-181b prowadzi do nadekspresji markerów naczyń limfatycznych oraz obniża poziom markerów tętniczych NRP1 oraz efryny B1 [13]. Fish i współpra-cownicy wykazali ponadto, że nadekspresja miR-126 w zarodkowych komórkach macierzystych poddanych na-stępnie różnicowaniu śródbłonkowemu nie wpływa na ekspresję Flk1, NOS3 (eNOS, śródbłonkowa syntaza NO), Tie2 ani CD31, co sugeruje, że ekspresja tego mikroRNA nie wystarcza by pobudzić różnicowanie [16]. Procesowi różnicowania ESC do komórek śródbłonka towarzyszy także spadek ekspresji miR-7641, hamującego ekspresję proangiogennej chemokiny CXCL1 [17].
WPŁYW mikroRNA NA DOJRZAŁE KOMÓRKI ŚRÓDBŁONKA
MikroRNA regulują rozliczne funkcje pełnione przez dojrzałe komórki śródbłonka. Zmniejszenie całkowitej puli mikroRNA poprzez obniżenie poziomu białek Dro-sha i Dicer za pomocą siRNA w ludzkich komórkach śródbłonka pochodzących z żyły pępowinowej (HUVEC) prowadzi do ograniczenia liczby wypustek utworzonych przez sferoidy zbudowane ze modyfikowanych komó-rek w trójwymiarowych hodowlach in vitro oraz zmniej-szenia długości struktur naczyniopodobnych w teście in vitro na matriżelu. W teście in vivo, komórki HUVEC z
obniżoną syntezą Drosha wykazują mniejszą aktywność angiogenną, a obniżenie poziomu RNazy Dicer ogranicza ich potencjał migracyjny. Zahamowanie ekspresji obu ge-nów nie ma natomiast wpływu na żywotność komórek HUVEC [18]. Podobne efekty zaobserwowano po zmniej-szeniu syntezy Dicer w ludzkich mikrowaskularnych ko-mórkach śródbłonka (HMEC). Towarzyszyło temu rów-nież zwiększenie syntezy czynnika VEGF, oraz wzrost produkcji reaktywnych form tlenu (ROS) w odpowie-dzi na stymulację czynnikiem martwicy nowotworów-α (TNFα) czy octanem mirystynianu forbolu (PMA) [19]. U dorosłych myszy, u których komórki śródbłonka pozba-wione były enzymu Dicer także zaobserwowano zmniej-szoną angiogenezę w odpowiedzi na VEGF oraz w trak-cie gojenia ran, regeneracji przepływu krwi po zabiegu niedokrwienia kończyny czy w trakcie wzrostu guzów nowotworowych [12].
Obniżenie ekspresji białka argonauty 2 (Ago2), od-powiedzialnego za cięcie mRNA w przypadku pełnej komplementarności nici mikroRNA z nicią mRNA oraz przy hamowaniu ekspresji za pomocą siRNA, prowadzi do zmniejszenia potencjału angiogennego oraz wzrostu komórek HUVEC in vitro [20]. McCall i współpracownicy przeprowadzili analizę ekspresji mikroRNA w ludzkich komórkach śródbłonka pochodzących z różnych odcin-ków łożyska naczyniowego, dzięki której zidentyfikowali 164 mikroRNA występujące w śródbłonku. Trzy spośród rozpoznanych mikroRNA tj. miR-99b, miR-20b oraz let--7b istotnie różniły się między analizowanymi rodzajami komórek śródbłonka, dzięki czemu można było podzielić je na trzy grupy: pierwszą, charakteryzującą się wysoką ekspresją miR-20b stanowiły komórki śródbłonka pocho-dzące z żyły pępowinowej oraz mózgowe komórki mi-krowaskularne; do drugiej grupy, o wysokiej ekspresji miR-99b, należały komórki śródbłonka z tętnic płucnych oraz komórki z naczyń wieńcowych; do trzeciej, o wyso-kiej ekspresji let-7b, komórki aortalne oraz mikrowasku-larne płuc i skóry [21]. Kuehbacher i współpracownicy zidentyfikowali w komórkach HUVEC wysoką ekspresję mikroRNA należących do rodziny let-7, miR-21, miR-126, miR-221 oraz miR-222 [18].
ROLA POSZCZEGÓLNYCH mikroRNA W REGULACJI FUNKCJI ŚRÓDBŁONKA
miR-17-92Grupa mikroRNA miR-17-92, w skład której wcho-
dzą miR-17–3p, miR-17–5p, miR-18a, miR-19a, miR-19b, miR-20a, oraz miR-92a kodowana jest w ludzkim geno-mie na długim ramieniu 13. chromosomu [22]. MiR-17-92 hamują produkcję czynników przeciwangiogennych, takich jak trombospondyna-1 (Tsp-1, głównie regulowana przez miR-19a), wydzielane kwaśne białko bogate w cy-steinę (SPARC), a także czynnik wzrostu tkanki łącznej (CTGF, regulowany przez miR-18a), który może zarówno hamować jak i pobudzać wzrost naczyń krwionośnych. W komórkach nowotworowych ekspresja miR-17-92 jest nasilana przez c-myc [23] (Ryc. 2A). MiR-19a może także regulować cykl komórkowy zmniejszając syntezę cykliny D1 [24].

408 www.postepybiochemii.pl
W komórkach HUVEC ekspresja miR-17-92 zwięk-sza się w odpowiedzi na stymulację czynnikiem VEGF. Nadekspresja miR-17-5p, miR-18a oraz miR-20a w śródbłonkach pozbawionych RNazy Dicer przywraca ich potencjał proliferacyjny oraz zdolność do tworze-nia struktur naczyniopodobnych na matriżelu, co może wiązać się ze zmniejszeniem ekspresji Tsp1 [12]. Z dru-giej jednak strony Doebele i współpracownicy wykazali, że zwiększenie ekspresji miR-17, miR-18a, miR-19a oraz miR-20a w komórkach HUVEC ogranicza zdolności an-giogenne komórek śródbłonka w testach in vitro oraz (dla miR-17 i -20a) in vivo. Ponadto, zahamowanie miR-17 oraz -20a nie zmniejszało unaczynienia guzów nowotwo-rowych [25].
Wśród genów regulowanych przez miR-17 i miR-20a zidentyfikowano p21 – odpowiedzialny za regulację cy-klu komórkowego, oraz kinazę tyrozynową Jak1 [25] (Ryc. 2A). Zwiększenie poziomu pre-miR-92a w ludzkich komórkach śródbłonka ograniczyło ich migrację, zdol-ność przylegania do fibronektyny czy tworzenia wypu-stek w teście sferoidowym in vitro oraz liczbę funkcjo-nalnych naczyń krwionośnych po podaniu w matriżelu in vivo. Zahamowanie ekspresji miR-92a za pomocą an-tagomirów zwiększyło natomiast potencjał angiogen-ny komórek śródbłonka oraz przyspieszyło regenerację przepływu krwi w niedokrwionych kończynach myszy. Transkryptami regulowanymi przez miR-92a, pełniącymi rolę w angiogenezie, są integryna α5 (ITGA5) oraz kina-za kinaz aktywowanych mitogenem 4 (MKK4) [26] (Ryc. 2A).
miR-21
Jednym z mikroRNA o bardzo wysokiej ekspresji w komórkach śródbłonka jest miR-21 [18]. Stymulacja ko-mórek HUVEC płodową surowicą bydlęcą lub zasado-wym czynnikiem wzrostu fibroblastów (FGF-2) obniża ekspresję miR-21. Transfekcja komórek śródbłonka za pomocą pre-miR-21 prowadzi do zaburzenia ich zdolno-
ści angiogennych in vitro oraz upośledza migrację. Przeciw-ny efekt uzyskano w przypad-ku zablokowania miR-21 za pomocą anty-miR-21 [27].
Wśród transkryptów, któ-rych ekspresja jest hamowa-na przez miR-21 rozpoznano Sprouty1 (Spry1), który może regulować aktywację kinaz ERK oraz RhoB, kluczowego regulatora cytoszkieletu, wpły-wającego między innymi na migrację komórek śródbłonka oraz ich właściwości angio-genne [27] (Ryc. 2B). Z drugiej jednak strony, zwiększona synteza miR-21 w komórkach nowotworowych stymuluje ekspresję HIF1α oraz VEGF poprzez zahamowanie funkcji PTEN, przyczyniając się w ten
sposób do zwiększenia unaczynienia guzów [28] (Ryc. 2B). Podobnie, zwiększenie poziomu miR-21 oraz towarzyszą-ce mu obniżenie syntezy PTEN zaobserwowano w komór-kach HUVEC poddanych działaniu sił ścinających (ang. shear stress). Komórki HUVEC z wysoką ekspresją miR-21 charakteryzowały się obniżoną apoptozą oraz zwiększo-ną produkcją tlenku azotu [29]. Ponadto, wyższa ekspre-sja miR-21 oraz miR-10A* w komórkach progenitorowych śródbłonka prowadzi do obniżenia poziomu białka grupy wysokiej mobilności A (HmgA) co przyspiesza starzenie się progenitorów [30] (Ryc. 2B).
miR-23-27-24
Ludzkie mikroRNA należące do grupy miR-23-27-24 kodowane są przez zduplikowane geny. MiR-23a, miR--27a oraz miR-24-1 kodowane są na długim ramieniu chromosomu 19, natomiast miR-23b, miR-27b oraz miR-24-2 w obrębie intronu genu znajdującego się na chromo-somie 9 [22]. Komórki HUVEC oraz mysie krążki aortalne transfekowane modyfikowanymi anty-miR-23a/b i anty--miR-27a/b wykazują obniżony potencjał angiogenny w testach in vitro. Odwrotny trend zaobserwowano przy zwiększeniu ekspresji miR-23a/b i miR-27a/b.
Transkryptami rozpoznawanymi przez miR-23a/b oraz miR-27a/b są SPROUTY2 oraz semaforyny 6A oraz 6D (Sema6A, Sema6D) (Ryc. 3). Białka Sprouty hamują szlak sygnałowy Ras/Raf/ERK. Stąd, obniżenie ekspresji miR-23a/b oraz miR-27a/b skutkuje obniżeniem fosfory-lacji ERK1/2, kluczowej dla angiogenezy, w odpowiedzi na stymulację komórek HUVEC czynnikiem VEGF. Z ko-lei semaforyna 6A hamuje fosforylację receptora VEGFR2 oraz kinazy ERK1/2 [31].
MiR-27b reguluje funkcje komórek wierzchołkowych (ang. tip cells) oraz rozgałęzianie naczyń krwionośnych wpływając na szlak sygnałowy Notch. Transkryptem docelowymi miR-27b jest Dll4, który, konkurując z Jag-
Rycina 2. Regulacja funkcji komórek śródbłonka przez rodzinę miR-17-92 (A), miR-21 (B), oraz miR-424 (C).

Postępy Biochemii 59 (4) 2013 409
1 o wiązanie z receptorem Notch hamuje powstawanie komórek wierzchołkowych (Ryc. 3). MiR-27b wpływa także na specyfikację żylną komórek śródbłonka oraz ich proliferację i migrację, obniżając poziom białka Spry2, które hamuje szlak sygnałowy kinaz MAP oraz reguluje ekspresję markera komórek żylnych ephB4 oraz markera śródbłonka limfatycznego receptora Flt4 [32] (Ryc. 3).
Poziom ekspresji miR-24 może być regulowany przez hi-poksję i czynnik transkrypcyjny HIF1, który ulega stabiliza-cji w sercowych komórkach śródbłonka po niedokrwieniu serca. Hamując syntezę GATA2, czynnika transkrypcyjne-go charakterystycznego dla komórek hematopoetycznych i śródbłonkowych, oraz kinazy seroninowo-treoninowej PAK4, miR-24 zmniejsza potencjał angiogenny komórek śródbłonka. GATA2 pobudza bowiem syntezę błonowego inhibitora BMP oraz aktywiny BAMBI, cząsteczki specyficz-nej dla komórek śródbłonka-1 (ESM1), sirtuiny-1 (SIRT1) i oksygenazy hemowej-1 (HMOX1) (Ryc. 3), enzymu rozkła-dającego hem do biliwerdyny, tlenku węgla i jonów Fe2+, który działa cytoprotekcyjnie, przeciwzapalnie i proangio-gennie [33]. Kinaza PAK4 fosforyluje natomiast proapopto-tyczne białko BAD, hamując jego aktywność. Stąd miR-24 wykazuje działanie przeciwangiogenne oraz zwiększające apoptozę.
miR-100
Niedokrwienie kończyny prowadzi do silnego niedo-tlenienia i gwałtownych zmian w ekspresji licznych ge-nów oraz mikroRNA. Jednym z mikroRNA, których po-ziom obniża się jest miR-100. Jego obecność potwierdzo-no w komórkach śródbłonka, pericytach oraz komórkach mięśni gładkich i szkieletowych. Ekspresja miR-100 jest niższa w rosnących naczyniach krwionośnych, w których obecne są dzielące się komórki, wykazujące obecność markera Ki67. Zwiększona ekspresja miR-100 nie zabu-rza żywotności komórek śródbłonka, natomiast obniża ich proliferację oraz właściwości angiogenne w testach in vitro. Obniżenie ekspresji miR-100 u myszy poddanych zabiegowi podwiązania tętnicy udowej przyspiesza re-
generację przepływu krwi i zwiększa unaczynienie nie-dokrwionej kończyny. MiR-100 hamuje ekspresję genu FRAP1 kodującego ssaczy cel dla rapamycyny (mTOR). Jednoczesne podanie rapamycyny oraz zahamowanie ekspresji miR-100 w mysim modelu in vivo spowalnia re-waskularyzację niedokrwionej kończyny [34].
miR-126
Najlepiej poznanym mikroRNA związanym ze śród-błonkiem naczyniowym jest proangiogenny miR-126. Jego ekspresja została pierwotnie wykazana w mysim sercu [35] oraz sercu i naczyniach krwionośnych Danio rerio [36]. Spo-śród wszystkich analizowanych mikroRNA, miR-126 wyka-zał najwyższy wzrost ekspresji w mysich komórkach CD31+ wywodzących się z różnicowanych zarodkowych komórek macierzystych [16]. Nić pasażerska miR-126* ulega niskiej ekspresji [37]. miR-126 oraz miR-126* kodowane są w ob-rębie siódmego intronu genu domeny podobnej do nabłon-kowego czynnika wzrostu 7 (Egfl7), która jest chemoatrak-tantem dla komórek śródbłonka, wpływającym na rozwój i integralność naczyń krwionośnych [38]. U myszy pozba-wionych miR-126, przy zachowanej ekspresji Egfl7, zaob-serwowano zwiększoną śmiertelność noworodków oraz zarodków, u których występowały obrzęki i liczne krwa-wienia. W sytuacji odwrotnej, gdy poziom miR-126 jest za-chowany, natomiast nie ma ekspresji Egfl7 nie obserwuje się zmian w żywotności i fenotypie myszy w porównaniu do myszy kontrolnych [39].
Ekspresja zarówno Egfl7, jak i miR-126 może być induko-wana przez czynnik transkrypcyjny Ets-1 [37] (Ryc. 4) oraz w odpowiedzi na siły ścinające przez czynnik transkryp-cyjny klf2a [40]. Brak miR-126 ogranicza wzrost komórek śródbłonka z krążków aortalnych w teście in vitro oraz ob-niża unaczynienie podskórnych implantów matriżelowych zawierających FGF-2. Ponadto, myszy pozbawione miR-126 gorzej przeżywają niedokrwienie serca [37]. Obniżenie eks-presji miR-126 oraz miR126* u danio pręgowanego, prowa-dzi do zaburzenia stabilności i szczelności naczyń krwiono-śnych przy zachowaniu ich morfologii [37].
Zahamowanie ekspresji miR-126 w komórkach HU-VEC ogranicza ich zdolność do migracji w odpowiedzi na VEGF i FGF-2 oraz do tworzenia struktur naczynio-podobnych na matriżelu [16]. Wśród genów, których eks-presja jest regulowana w komórkach HUVEC przez miR-126 zidentyfikowano Egfl7, co świadczy o ujemnej pętli zwrotnej regulacji ekspresji miR-126, oraz geny białka spokrewnionego z białkiem Sprouty, zawierającego domenę EZH (SPRED1), białko adhezyjne komórek na-czyń 1 (VCAM1) [16,37] i podjednostkę regulacyjną ki-nazy trifosforanu inozytolu 2 (PIK3R2) [16] (Ryc. 4). Re-gulacja syntezy VCAM1, potwierdzona w niezależnym badaniu, sugeruje iż miR-126 może wpływać na adhezję leukocytów do śródbłonka i reakcję zapalną [41]. SPRED1 hamuje przekaz części sygnału od receptorów czynników wzrostu takich jak VEGF, FGF-2 czy czynnik wzrostu nabłonka (EGF) negatywnie regulując aktywność kinaz MAP. PIK3R2 hamuje natomiast aktywność kinazy trifos-foranu inozytolu, co wpływa między innymi na migrację komórek HUVEC pod wpływem VEGF [16].
Rycina 3. Regulacja funkcji komórek śródbłonka przez rodzinę miR-23-27-24.

410 www.postepybiochemii.pl
miR-130A
Poziom miR-130a w komórkach śródbłonka rośnie po stymulacji czynnikami wzrostu takimi jak FGF-2 lub VEGF, co prowadzi do obniżenia ekspresji przeciwangiogennego genu homeotycznego zatrzymania wzrostu GAX oraz genu homeotycznego HOXA5 (Ryc.4). GAX hamuje wzrost na-czyń krwionośnych poprzez obniżenie aktywności NFκB oraz zatrzymuje cykl komórkowy w fazie G0 poprzez in-dukcję p21WAF1/CIP1, niezależną od p53 [42]. HOXA5 zwięk-sza natomiast syntezę przeciwangiogennej trombospondy-ny-2 oraz hamuje ekspresję podjednoski α zależnego od nie-dotlenienia czynnika transkrypcyjnego HIF1 [43] (Ryc. 4).
miR-132
MiR-132, którego sekwencja kodowana jest w obrębie re-jonu międzygenowego, znajdującego się na długim ramie-niu chromosomu 11 [22], jest indukowany w komórkach nerwowych przez CREB (białko wiążące się z elementem odpowiedzi na cAMP) po stymulacji BDNF (czynnik neu-rotroficzny pochodzenia mózgowego) [44]. W komórkach śródbłonka w stanie spoczynku ekspresja miR-132 nie jest wykrywalna, rośnie natomiast po stymulacji czynnikami VEGF oraz FGF-2 lub mediami kondycjonowanymi znad komórek nowotworowych. Dalsze badania przeprowadzo-ne na modelu zwierzęcym pozwoliły stwierdzić, iż zahamo-wanie ekspresji miR-132 in vivo w rozwijających się oczach mysich noworodków prowadziło do obniżenia waskula-ryzacji siatkówki [45]. Transkryptem regulowanym przez miR-132 jest Rasa1 kodujący białko p120RasGAP [45], które zwiększając wewnętrzną aktywność GTPazy białka Ras po-woduje zahamowanie jego aktywności [46], w tym pobu-dzania angiogenezy w odpowiedzi na VEGF [47].
miR-200b oraz miR-200c
MikroRNA należące do grupy miR-200, takie jak miR--200a, miR-200b oraz miR-200c, odgrywają istotną rolę w re-
gulacji przejścia nabłonkowo-mezenchymalnego w trakcie rozwoju nowotworów, bezpośrednio wpływając na poziom czynników ZEB1 oraz ZEB2 [48]. W ludzkich mikrowasku-larnych komórkach śródbłonka ekspresja miR-200b zmniej-sza się przy niskim stężeniu tlenu w wyniku stabilizacji czynnika transkrypcyjnego HIF1α.
Obniżenie ekspresji miR-200b zwiększa potencjał an-giogenny śródbłonka. Proangiogennym czynnikiem, którego ekspresja regulowana jest przez miR-200b bez-pośrednio, niezależnie od ZEB1 oraz ZEB2, jest Ets-1, po-budzający syntezę między innymi MMP1 (metaloprote-inazy macierzy-1) oraz receptora VEGFR-2 [49] (Ryc. 4). Choi i współpracownicy zidentyfikowali ponadto VEGF oraz jego receptory VEGFR-1 i VEGFR-2 jako transkryp-ty bezpośrednio regulowane przez miR-200b w linii raka płuc A549. Transfekcja komórek śródbłonka HUVEC za pomocą miR-200b obniża ich zdolność do tworzenia struktur naczyniopodobnych na matriżelu oraz hamuje aktywację kinazy ERK1/2 w odpowiedzi na stymulację VEGF [50]. Z kolei stres oksydacyjny stymuluje w komór-kach HUVEC ekspresję miR-200c, który może hamować podziały komórkowe oraz zwiększać apoptozę komórek śródbłonka. Wyższej ekspresji miR-200c towarzyszy też zwiększony poziom SA-β galaktozydazy oraz białka p21 odpowiedzialnego za zatrzymanie wzrostu i starzenie się komórek. Transkryptem, którego zahamowanie po-woduje zmiany obserwowane w komórkach śródbłonka wykazujących zwiększony poziom miR-200c, jest ZEB1 [51] (Ryc. 4).
miR-210
Obniżenie stężenia tlenu w tkankach, np. w wyniku niedokrwienia tkanki lub w trakcie wzrostu nowotworów, aktywuje kaskadę sygnałową pozwalającą między innymi na zwiększenie proliferacji oraz migracji komórek śród-błonka, a co się z tym wiąże, na pobudzenie procesu angio-genezy. Jednym z mikroRNA, których ekspresja ulega sil-
Rycina 4. Sieć regulacji funkcji śródbłonka przez miR-126, miR-130a, miR-200b/c, miR-210 oraz miR-221/222.

Postępy Biochemii 59 (4) 2013 411
nym zmianom w warunkach niedotlenienia jest miR-210. Wzrost jego poziomu w komórkach HUVEC hodowanych w atmosferze zawierającej 1% tlenu wynika z aktywacji czynnika HIF1α (Ryc. 4).
Zahamowanie ekspresji miR-210 zmniejsza właściwości angiogenne komórek HUVEC zarówno w warunkach nie-dotlenienia jak i przy normalnym stężeniu tlenu. Fasanaro i współpracownicy zidentyfikowali efrynę A3 jako trans-krypt bezpośrednio regulowany przez miR-210 w warun-kach obniżonego stężenia tlenu (Ryc. 4). Dokładna rola efryny A3 w regulacji angiogenezy nie jest znana, jednak jej obniżenie przez miR-210 jest konieczne dla stymulacji chemotaksji i tworzenia struktur naczyniopodobnych przez komórki śródbłonka w odpowiedzi na VEGF [52].
miR-214
MiR-214 ulega silnej ekspresji w większości typów doj-rzałych komórek śródbłonka. Zwiększona ekspresja miR-214 ogranicza liczbę wypustek tworzonych przez mysie zarodkowe komórki śródbłonka w teście sferoidowym oraz struktur naczyniopodobnych przez komórki HUVEC na matriżelu. Wyciszenie ekspresji miR-214 in vivo powoduje zwiększenie waskularyzacji podskórnych implantów ma-triżelowych oraz siatkówki u mysich noworodków. Trans-kryptem odpowiedzialnym za wpływ miR-214 na angioge-nezę jest Quaking (QKI), którego obniżenie zmniejsza pro-dukcję czynników proangiogennych, w tym VEGF, FGF-2 czy PDGF. Zahamowanie ekspresji miR-214 w ludzkich ko-mórkach śródbłonka mikrowaskularnego HMEC zwiększa produkcję VEGF, FGF-2, EGF oraz IGF-1 (insulinopodobny czynnik wzrostu-1) [53].
miR-217
Wraz ze starzeniem się ludzkich komórek HUVEC ro-śnie w nich ekspresja miR-217, który bezpośrednio hamuje syntezę zależnej od NAD deacetylazy sirtuiny-1 (SiRT1). Obniżenie poziomu SiRT1 w komórkach HUVEC na niskim pasażu wiąże się ze wzrostem zawartości β-galaktozydazy związanej ze starzeniem się (SA-βgal) oraz prowadzi do osłabienia ich potencjału angiogennego in vitro i obniżenia poziomu NOS3 [54].
miR-221/222
Kolejnymi mikroRNA ulegającymi wysokiej ekspresji w komórkach śródbłonka są miR-221 oraz miR-222 [55], ko-dowane przez wspólny policistronowy gen znajdujący się na krótkim ramieniu chromosomu X [22] oraz posiadające identyczną sekwencję docelową (ang. seed sequence) warun-kującą rozpoznawanie regulowanych mRNA.
Jednym z genów regulowanych przez miR-221/222 w ludzkich komórkach śródbłonka HUVEC jest CKIT, który koduje receptor dla czynnika wzrostu komórek macierzy-stych SCF (Ryc. 4). Nadekspresja miR-221/222 powoduje obniżenie poziomu białka c-kit, zaburzając zdolność ko-mórek HUVEC do migracji i tworzenia struktur naczynio-podobnych w odpowiedzi na SCF [55]. Innym transkryp-tem regulowanym przez miR-221/222 jest czynnik trans-
krypcyjny Ets-1, który pobudza produkcję między innymi VCAM-1, MCP-1 (czynnik chemotaktyczny monocytów) czy podjednostki p47phox oksydazy NAD(P)H, wpływając w ten sposób na stan zapalny w naczyniach krwionośnych [56] (Ryc. 4).
Mimo takiej samej sekwencji docelowej miR-221 oraz miR-222 regulują ekspresję różnych genów, co może mieć związek z różnicami w pozostałych częściach ich sekwencji. MiR-221 wykazuje działanie przeciwangiogenne poprzez zahamowa-nie genu ZEB2 zaangażowanego w przejście epitelialno-me-zenchymalne, oraz genu GAX kodującego represor inhibitora angiogenezy [57] (Ryc. 4). Z drugiej jednak strony, miR-221 ulega wysokiej ekspresji u rozwijających się zarodków danio pręgowanego, a obniżenie jego poziomu powoduje zaburze-nia rozwoju unaczynienia. Danio pręgowane pozbawione genu miR-221 mają fenotyp podobny do ryb pozbawionych kinazy tyrozynowej spokrewnionej z Fms (flt4) czyli recepto-ra dla czynników wzrostu VEGF-C i VEGF-D.
Brak miR-221 zaburza migrację komórek wierzchołko-wych nowo tworzonych naczyń, kluczowych dla prawidło-wej angiogenezy, ale także zapobiega nadmiernemu two-rzeniu naczyń krwionośnych przy niedoborze sygnałów ścieżki Notch. Co ciekawe, ekspresja miR-221 ulega obni-żeniu przez Notch, co hamuje w konsekwencji proliferację i migrację komórek wierzchołkowych. Do genów regulo-wanych przez miR-221 u danio pręgowanego należy także zależny od cykliny inhibitor kinazy 1b (cdkn1b), który może hamować cykl komórkowy, oraz podjednostka regulatoro-wa p85α kompleksu kinazy trifosforanu inozytolu (pik3r1) [58].
Ekspresja miR-222 obniża się w komórkach HUVEC trak-towanych FGF-2 i IL-3, czyli w odpowiedzi na pobudzenie angiogenezy związanej z zapaleniem. Genem regulowanym bezpośrednio przez miR-222 jest STAT5A, kodujący czyn-nik transkrypcyjny regulujący proliferację i przeżywalność komórek śródbłonka. Ponadto, ekspresja miR-222 ujemnie koreluje z ekspresją STAT5A w komórkach śródbłonka izo-lowanych z naczyń z płytką miażdżycową [59]. Co cieka-we, zaobserwowano niższą ekspresję miR-222 u samic niż u samców myszy C57BL/6, co wiązało się z wyższą zawarto-ścią Ets-1, która prowadziła do podniesienia poziomu NOS3 i w konsekwencji wyższej produkcji tlenku azotu [60].
miR-378
Angiogeneza związana ze wzrostem nowotworów jest kluczowym procesem dla ich rozwoju oraz przerzutowania. Jednym z mikroRNA istotnie wpływających na unaczynie-nie guzów jest miR-378, kodowany w obrębie genu PGC1B, kodującego koaktywator 1β receptora PPARγ. Zwiększona ekspresja miR-378 w ludzkiej linii glejaka wielopostaciowe-go U87 poprawia przeżywalność komórek oraz nasila ich właściwości angiogenne in vitro i in vivo poprzez obniżenie poziomu ekspresji supresora białek fuzyjnych Sufu oraz Fus-1 [61]. Co ciekawe, zwiększona synteza oksygenazy hemowej-1 w mioblastach [62] lub liniach niedrobnoko-mórkowego raka płuc NCI-H292 [63] powoduje obniżenie poziomu miR-378. Z kolei zwiększenie ekspresji miR-378 w linii NCI-H292 obniża poziom HMOX1, a jednocześnie

412 www.postepybiochemii.pl
podwyższa produkcję czynnika VEGF, interleukiny 8, an-giopoetyny 1 i mucyny 5AC, wpływając w ten sposób na właściwości proangiogenne komórek nowotworowych [63]. Ponadto, w komórkach raka jamy nosowo gardłowej CNE podwyższony poziom miR-378 prowadzi do zwiększenia produkcji VEGF poprzez konkurencję z miR-125a o miejsce docelowe w 3’ UTR VEGF [64]. Rola miR-378 w komórkach śródbłonka jest słabo poznana, ale ze względu na jego za-angażowanie (zarówno formy miR-378 jak i miR-378*) w regulację ekspresji wielu różnych genów związanych z an-giogenezą, przypuszczać można, iż działanie to przejawia się także bezpośrednio w śródbłonku.
miR-424
Obniżenie stężenia tlenu prowadzi w ludzkich komór-kach śródbłonka do zwiększenia ekspresji miR-424, które-go mysim homologiem jest miR-322. Zwiększenie ekspresji miR-424 w komórkach HUVEC, przy zachowaniu normal-nego stężenia tlenu, stabilizuje podjednostki HIF1α oraz HIF2α czynnika transkrypcyjnego HIF, zwiększa aktywność promotora VEGF oraz nasila syntezę transportera glukozy GLUT-1 i erytropoetyny. Dzieje się tak, gdyż transkryptem regulowanym przez miR-424 jest kullina-2 (CUL2), element kompleksu VCBCR (Ryc. 2), który odpowiedzialny jest za ubikwitynację podjednostek HIF1α oraz HIF2α kierującą je do degradacji w proteasomie. Nadekspresja miR-424 zwięk-sza migrację i potencjał angiogenny komórek śródbłonka in vitro oraz zwiększa unaczynienie guzów nowotworowych in vivo. U myszy poddanych zabiegowi podwiązania tętnicy udowej ekspresja miR-332 kolokalizuje z naczyniami krwio-nośnymi a jego poziom koreluje z unaczynieniem niedo-krwionych mięśni. Ekspresja miR-424 regulowana jest przez należący do rodziny Ets czynnik transkrypcyjny PU.1, indu-kowany przez czynniki Runx-1 oraz C/EBPα [65] (Ryc. 2).
PODSUMOWANIE
Śródbłonek naczyniowy odgrywa ważną rolę między in-nymi w regulacji gojenia ran oraz wzroście nowotworów. Stąd dokładna regulacja jego funkcji, a w szczególności procesu tworzenia nowych naczyń krwionośnych, stanowi atrakcyjny cel terapii wielu chorób. Czynnikami, które re-gulują szlaki sygnałowe zaangażowane w kontrolę aktyw-ności komórek śródbłonka są cząsteczki mikroRNA. Każda z nich może wpływać na ekspresję wielu genów co umoż-liwia dokładne dostrajanie odpowiedzi na zmieniające się warunki.
Różnicowaniu śródbłonkowemu towarzyszą zmiany ekspresji wielu mikroRNA, z których miR-99b, miR-181a oraz miR-181b bezpośrednio wpływają na zmiany ekspre-sji markerów śródbłonkowych oraz specyfikację komórek śródbłonka. MikroRNA zaangażowane są także zarówno w proces hamowania jak i pobudzania angiogenezy, zarówno w warunkach niedotlenienia jak i w odpowiedzi na stymu-lację czynnikami wzrostu. Do mikroRNA o dość dobrze po-znanych właściwościach proangiogennych należą między innymi miR-126, miR-1301, miR-132, miR-378 oraz miR-214. Rola odgrywana przez inne microRNA, w szczególności miR-221 oraz -222, nie jest jednoznaczna. Niemniej, funkcja mikroRNA w komórkach śródbłonka jest ciekawym tema-
tem badań, szczególnie w kontekście terapeutycznej regula-cji ekspresji genów.
PIŚMIENNICTWO1. Pries AR, Secomb TW, Gaehtgens P (2000) The endothelial surface lay-
er. Pflügers Archiv 440: 653-6662. Hobson B, Denekamp J (1984) Endothelial proliferation in tumours and
normal tissues: continuous labelling studies. Br J Cancer 49: 405-4133. Vita JA (2011) Endothelial function. Circulation 124: e906-9124. Tonnesen MG, Feng X, Clark RA (2000) Angiogenesis in wound heal-
ing. J Investig Dermatol Symp Proc 5: 40-465. Folkman J (2002) Role of angiogenesis in tumor growth and metastasis.
Semin Oncol 29: 29(6 Suppl 16): 15-186. Winter J, Jung S, Keller S, Gregory RI, Diederichs S (2009) Many roads
to maturity: microRNA biogenesis pathways and their regulation. Na-ture Cell Biol 11: 228-234
7. Gu S, Kay MA (2010) How do miRNAs mediate translational repres-sion? Silence 1: 11
8. Vokes SA, Krieg PA (2002) Endoderm is required for vascular endothe-lial tube formation, but not for angioblast specification. Development 129: 775-785
9. Yang WJ (2004) Dicer is required for embryonic angiogenesis during mouse development. J Biol Chem 280: 9330-9335
10. Giraldez AJ, Cinalli RM, Glasner ME, Enright AJ, Thomson JM, Bask-erville S, Hammond SM, Bartel DP, Schier AF (2005) MicroRNAs reg-ulate brain morphogenesis in zebrafish. Science 308: 833-838
11. Cobb BS, Nesterova TB, Thompson E, Hertweck A, O’Connor E, God-win J, Wilson CB, Brockdorff N, Fisher AG, Smale ST, Merkenschlager M (2005) T cell lineage choice and differentiation in the absence of the RNase III enzyme Dicer. J Exp Med 201: 1367-1373
12. Suárez Y, Fernández-Hernando C, Yu J, Gerber SA, Harrison KD, Pober JS, Iruela-Arispe ML, Merkenschlager M, Sessa WC (2008) Di-cer-dependent endothelial microRNAs are necessary for postnatal an-giogenesis. Proc Natl Acad Sci 105: 14082-14087
13. Kane NM, Howard L, Descamps B, Meloni M, McClure J, Lu R, Mc-Cahill A, Breen C, Mackenzie RM, Delles C, Mountford JC, Milligan G, Emanueli C, Baker AH (2012) Role of microRNAs 99b, 181a, and 181b in the differentiation of human embryonic stem cells to vascular endothelial cells. Stem Cells 30: 643-654
14. Kane NM, Meloni M, Spencer HL, Craig MA, Strehl R, Milligan G, Houslay MD, Mountford JC, Emanueli C, Baker AH (2010) Deriva-tion of endothelial cells from human embryonic stem cells by directed differentiation: analysis of microRNA and angiogenesis in vitro and in vivo. Arterioscler Thromb Vasc Biol 30: 1389-1397
15. Tréguer K, Heinrich E-M, Ohtani K, Bonauer A, Dimmeler S (2012) Role of the microRNA-17–92 cluster in the endothelial differentiation of stem cells. J Vasc Res 49: 447-460
16. Fish JE, Santoro MM, Morton SU, Yu S, Yeh RF, Wythe JD, Ivey KN, Bruneau BG, Stainier DY, Srivastava D (2008) miR-126 regulates angio-genic signaling and vascular integrity. Dev Cell 15: 272-284
17. Yoo JK, Jung HY, Kim CH, Son WS, Kim JK (2013) miR-7641 modu-lates the expression of CXCL1 during endothelial differentiation de-rived from human embryonic stem cells. Arch Pharm Res 36: 353-358
18. Kuehbacher A, Urbich C, Zeiher AM, Dimmeler S (2007) Role of Dicer and Drosha for endothelial microRNA expression and angiogenesis. Circ Res 101: 59-68
19. Shilo S, Roy S, Khanna S, Sen CK (2008) Evidence for the involvement of miRNA in redox regulated angiogenic response of human micro-vascular endothelial cells. Arterioscler Thromb Vasc Biol 28: 471-477
20. Asai T, Suzuki Y, Matsushita S, Yonezawa S, Yokota J, Katanasaka Y, Ishida T, Dewa T, Kiwada H, Nango M, Oku N (2008) Disappearance of the angiogenic potential of endothelial cells caused by Argonaute2 knockdown. Biochem Biophys Res Commun 368: 243-248
21. McCall MN, Kent OA, Yu J, Fox-Talbot K, Zaiman AL, Halushka MK (2011) MicroRNA profiling of diverse endothelial cell types. BMC Med Genomics 4: 78

Postępy Biochemii 59 (4) 2013 413
22. Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M, Croce CM (2004) Human microRNA genes are frequently located at fragile sites and genomic regions involved in cancers. Proc Natl Acad Sci USA 101: 2999-3004
23. Dews M, Homayouni A, Yu D, Murphy D, Sevignani C, Wentzel E, Furth EE, Lee WM, Enders GH, Mendell JT, Thomas-Tikhonenko A (2006) Augmentation of tumor angiogenesis by a Myc-activated mi-croRNA cluster. Nat Genet 38: 1060-1065
24. Yin R, Bao W, Xing Y, Xi T, Gou S (2012) MiR-19b-1 inhibits angiogen-esis by blocking cell cycle progression of endothelial cells. Biochem Biophys Res Commun 417: 771-776
25. Doebele C, Bonauer A, Fischer A, Scholz A, Reiss Y, Urbich C, Hofmann WK, Zeiher AM, Dimmeler S (2010) Members of the mi-croRNA-17-92 cluster exhibit a cell-intrinsic antiangiogenic function in endothelial cells. Blood 115: 4944-4950
26. Bonauer A, Carmona G, Iwasaki M, Mione M, Koyanagi M, Fischer A, Burchfield J, Fox H, Doebele C, Ohtani K, Chavakis E, Potente M, Tjwa M, Urbich C, Zeiher AM, Dimmeler S (2009) MicroRNA-92a con-trols angiogenesis and functional recovery of ischemic tissues in mice. Science 324: 1710-1713
27. Sabatel C, Malvaux L, Bovy N, Deroanne C, Lambert V, Gonzalez ML, Colige A, Rakic JM, Noël A, Martial JA, Struman I (2011) MicroR-NA-21 exhibits antiangiogenic function by targeting RhoB expression in endothelial cells. PLoS ONE 6: e16979
28. Liu L-Z, Li C, Chen Q, Jing Y, Carpenter R, Jiang Y, Kung HF, Lai L, Jiang BH (2011) MiR-21 induced angiogenesis through AKT and ERK activation and HIF-1α expression. PLoS ONE 6: e19139
29. Weber M, Baker MB, Moore JP, Searles CD (2010) MiR-21 is induced in endothelial cells by shear stress and modulates apoptosis and eNOS activity. Biochem Biophys Res Commun 393: 643-648
30. Zhu S, Deng S, Ma Q, Zhang T, Jia C, Zhuo D, Yang F, Wei J, Wang L, Dykxhoorn DM, Hare JM, Goldschmidt-Clermont PJ, Dong C (2013) MicroRNA-10A* and MicroRNA-21 modulate endothelial progenitor cell senescence via suppressing high-mobility group A2. Circ Res 112: 152-164
31. Zhou Q, Gallagher R, Ufret-Vincenty R, Li X, Olson EN, Wang S (2011) Regulation of angiogenesis and choroidal neovascularization by mem-bers of microRNA-23~27~24 clusters. Proc Natl Acad Sci 108: 8287-8292
32. Biyashev D, Veliceasa D, Topczewski J, Topczewska JM, Mizgirev I, Vinokour E, Reddi AL, Licht JD, Revskoy SY, Volpert OV (2012) miR-27b controls venous specification and tip cell fate. Blood 119: 2679-2687
33. Dulak J, Deshane J, Jozkowicz A, Agarwal A (2008) Heme oxygenase-1 and carbon monoxide in vascular pathobiology: focus on angiogene-sis. Circulation 117: 231-241
34. Grundmann S, Hans FP, Kinniry S, Heinke J, Helbing T, Bluhm F, Sluijter JP, Hoefer I, Pasterkamp G, Bode C, Moser M (2011) MicroR-NA-100 regulates neovascularization by suppression of mammalian target of rapamycin in endothelial and vascular smooth muscle cells. Circulation 123: 999-1009
35. Lagos-Quintana M, Rauhut R, Yalcin A, Meyer J, Lendeckel W, Tuschl T (2002) Identification of tissue-specific microRNAs from mouse. Curr Biol 12: 735-739
36. Wienholds E (2005) MicroRNA expression in Zebrafish embryonic de-velopment. Science 309: 310-311
37. Wang S, Aurora AB, Johnson BA, Qi X, McAnally J, Hill JA, Richard-son JA, Bassel-Duby R, Olson EN (2008) The endothelial-specific mi-croRNA miR-126 governs vascular integrity and angiogenesis. Devel-op Cell 15: 261-271
38. Nichol D, Stuhlmann H (2012) EGFL7: a unique angiogenic signaling factor in vascular development and disease. Blood 119: 1345-1352
39. Kuhnert F, Mancuso MR, Hampton J, Stankunas K, Asano T, Chen CZ, Kuo CJ (2008) Attribution of vascular phenotypes of the murine Egfl7 locus to the microRNA miR-126. Development 135: 3989-3993
40. Nicoli S, Standley C, Walker P, Hurlstone A, Fogarty KE, Lawson ND (2010) MicroRNA-mediated integration of haemodynamics and Vegf signalling during angiogenesis. Nature 464: 1196-1200
41. Harris TA, Yamakuchi M, Ferlito M, Mendell JT, Lowenstein CJ (2008) MicroRNA-126 regulates endothelial expression of vascular cell adhe-sion molecule 1. Proc Natl Acad Sci USA 105: 1516-1521
42. Chen Y, Gorski DH (2007) Regulation of angiogenesis through a mi-croRNA (miR-130a) that down-regulates antiangiogenic homeobox genes GAX and HOXA5. Blood 111: 1217-1226
43. Zhu Y, Cuevas IC, Gabriel RA, Su H, Nishimura S, Gao P, Fields A, Hao Q, Young WL, Yang GY, Boudreau NJ (2009) Restoring transcrip-tion factor HoxA5 expression inhibits the growth of experimental hemangiomas in the brain. J Neuropathol Exp Neurol 68: 626-632
44. Vo N, Klein ME, Varlamova O, Keller DM, Yamamoto T, Goodman RH, Impey S (2005) A cAMP-response element binding protein-in-duced microRNA regulates neuronal morphogenesis. Proc Natl Acad Sci USA 102: 16426-16431
45. Anand S, Majeti BK, Acevedo LM, Murphy EA, Mukthavaram R, Scheppke L, Huang M, Shields DJ, Lindquist JN, Lapinski PE, King PD, Weis SM, Cheresh DA (2010) MicroRNA-132–mediated loss of p120RasGAP activates the endothelium to facilitate pathological an-giogenesis. Nat Med 16: 909-914
46. McCormick F (1989) ras GTPase activating protein: signal transmitter and signal terminator. Cell 56: 5-8
47. Meadows KN, Bryant P, Vincent PA, Pumiglia KM (2004) Activated Ras induces a proangiogenic phenotype in primary endothelial cells. Oncogene 23: 192-200
48. Korpal M, Kang Y (2008) The emerging role of miR-200 family of mi-croRNAs in epithelial-mesenchymal transition and cancer metastasis. RNA Biol 5: 115-119
49. Chan YC, Khanna S, Roy S, Sen CK (2011) miR-200b targets Ets-1 and is down-regulated by hypoxia to induce angiogenic response of endo-thelial cells. J Biol Chem 286: 2047-2056
50. Choi Y-C, Yoon S, Jeong Y, Yoon J, Baek K (2011) Regulation of vas-cular endothelial growth factor signaling by miR-200b. Mol Cells 32: 77-82
51. Magenta A, Cencioni C, Fasanaro P, Zaccagnini G, Greco S, Sarra-Fer-raris G, Antonini A, Martelli F, Capogrossi MC (2011) miR-200c is up-regulated by oxidative stress and induces endothelial cell apoptosis and senescence via ZEB1 inhibition. Cell Death Differ 18: 1628-1639
52. Fasanaro P, D’Alessandra Y, Di Stefano V, Melchionna R, Romani S, Pompilio G, Capogrossi MC, Martelli F (2008) MicroRNA-210 modula-tes endothelial cell response to hypoxia and inhibits the receptor tyro-sine kinase ligand Ephrin-A3. J Biol Chem 283: 15878-15883
53. van Mil A, Grundmann S, Goumans MJ, Lei Z, Oerlemans MI, Jaksani S, Doevendans PA, Sluijter JP (2012) MicroRNA-214 inhibits angio-genesis by targeting Quaking and reducing angiogenic growth factor release. Cardiovasc Res 93: 655-665
54. Menghini R, Casagrande V, Cardellini M, Martelli E, Terrinoni A, Amati F, Vasa-Nicotera M, Ippoliti A, Novelli G, Melino G, Lauro R, Federici M (2009) MicroRNA 217 modulates endothelial cell senes-cence via silent information regulator 1. Circulation 120: 1524-1532
55. Poliseno L, Tuccoli A, Mariani L, Evangelista M, Citti L, Woods K, Mercatanti A, Hammond S, Rainaldi G (2006) MicroRNAs modulate the angiogenic properties of HUVECs. Blood 108: 3068-3071
56. Zhu N, Zhang D, Chen S, Liu X, Lin L, Huang X, Guo Z, Liu J, Wang Y, Yuan W, Qin Y (2011) Endothelial enriched microRNAs regulate angiotensin II-induced endothelial inflammation and migration. Ath-erosclerosis 215: 286-293
57. Chen Y, Banda M, Speyer CL, Smith JS, Rabson AB, Gorski DH (2010) Regulation of the expression and activity of the antiangiogenic homeo-box gene GAX/MEOX2 by ZEB2 and microRNA-221. Mol Cel Biol 30: 3902-3913
58. Nicoli S, Knyphausen C-P, Zhu LJ, Lakshmanan A, Lawson ND (2012) miR-221 is required for endothelial tip cell behaviors during vascular development. Dev Cell 22: 418-429
59. Dentelli P, Rosso A, Orso F, Olgasi C, Taverna D, Brizzi MF (2010) microRNA-222 controls neovascularization by regulating signal transducer and activator of transcription 5A expression. Arterioscler Thromb Vasc Biol 30: 1562-1568

414 www.postepybiochemii.pl
Role of microRNA in endothelial cells — regulation of differentiation and angiogenesisWitold Norbert Nowak1,2, Urszula Florczyk1, Alicja Józkowicz1, Józef Dulak1,
1Department of Medical Biotechnology, Faculty of Biochemistry, Biophysics and Biotechnology, Jagiellonian University, 7 Gronostajowa St., 30-387 Krakow, Poland 2Jagiellonian Centre for Experimental Therapeutics, Jagiellonian University, 14 Bobrzyńskiego St., 30-348 Krakow, Polande-mail: [email protected]
Key words: microRNA, endothelium, angiogenesis, differentiation, embryonic stem cells
ABSTRACTEndothelium has an immense impact on the tissue regeneration, regulation of atherosclerosis and tumour growth. Therefore, modification of endothelial cell differentiation and function seems a promising target for many therapies. MicroRNAs are small RNA molecules, which recognize and inhibit specific mRNAs. In that way, they can regulate and orchestrate whole signalling pathways. It has been shown that mi-croRNAs can fine-tune endothelial cell functions since they have either pro- and antiangiogenic activity, regulate expression of e.g. adhesion molecules or nitric oxide synthase. Furthermore, microRNAs modulate differentiation of embryonic stem cells to endothelial cells and their further specialization towards specific vascular bed. This review focuses mainly on the influence of microRNA on the angiogenesis and en-dothelial cell differentiation.
60. Evangelista AM, Deschamps AM, Liu D, Raghavachari N, Murphy E (2013) miR-222 contributes to sex-dimorphic cardiac eNOS expression via ets-1. Physiol Genomics 45: 493-498
61. Lee DY, Deng Z, Wang C-H, Yang BB (2007) MicroRNA-378 promotes cell survival, tumor growth, and angiogenesis by targeting SuFu and Fus-1 expression. Proc Natl Acad Sci USA 104: 20350-20355
62. Kozakowska M, Ciesla M, Stefanska A, Skrzypek K, Was H, Jazwa A, Grochot-Przeczek A, Kotlinowski J, Szymula A, Bartelik A, Mazan M, Yagensky O, Florczyk U, Lemke K, Zebzda A, Dyduch G, Nowak W, Szade K, Stepniewski J, Majka M, Derlacz R, Loboda A, Dulak J, Jozko-wicz A (2012) Heme oxygenase-1 inhibits myoblast differentiation by targeting myomirs. Antioxid Redox Signal 16: 113-127
63. Skrzypek K, Tertil M, Golda S, Ciesla M, Weglarczyk K, Collet G, Guichard A, Kozakowska M, Boczkowski J, Was H, Gil T, Kuzdzal J,
Muchova L, Vitek L, Loboda A, Jozkowicz A, Kieda C, Dulak J (2013) Interplay between heme oxygenase-1 and miR-378 affects non-small cell lung carcinoma growth, vascularization, and metastasis. Antioxid Redox Signal 19: 644-660
64. Hua Z, Lv Q, Ye W, Wong CK, Cai G, Gu D, Ji Y, Zhao C, Wang J, Yang BB, Zhang Y (2006) MiRNA-directed regulation of VEGF and other angiogenic factors under hypoxia. PLoS ONE 1: e116
65. Ghosh G, Subramanian IV, Adhikari N, Zhang X, Joshi HP, Basi D, Chandrashekhar YS, Hall JL, Roy S, Zeng Y, Ramakrishnan S (2010) Hypoxia-induced microRNA-424 expression in human endothelial cells regulates HIF-α isoforms and promotes angiogenesis. J Clin In-vest 120: 4141-4154


















