
Produkcja roœlinnych metabolitów wtórnych w kulturach ... · BIOTECHNOLOGIA 4 (75) 124-135 2006...
12
Produkcja roœlinnych metabolitów wtórnych w kulturach organów transformowanych Halina Wysokiñska, Aleksander Chmiel Katedra Biologii i Biotechnologii Farmaceutycznej, Uniwersytet Medyczny, £ódŸ Secondary metabolites production in cultures of transformed plant organs Summary Plant cell cultures in vitro produce secondary metabolites with varied effec- tiveness. Despite industrial application of only few cell suspension cultures, considerable progress in research on plant cell biotechnology has been made over the last few years. Transformed organ cultures, especially hairy roots, seem to be an interesting model for stable production of plant metabolites with high yield. In this paper, cultures of hairy roots of Salvia sclarea, S. officinalis, S. miltiorrhiza, S. przewalskii, and Centaurium erythraea, as well as transformed shoots and plants of C. erythraea are presented. Also, production of secondary metabolites in these cultures is discussed. Key words: plant cultures, transformed shoots, transformed roots, secondary metabo- lite production. 1. Wstêp Roœlinne metabolity wtórne s¹ szeroko wykorzystywane jako farmaceutyki, substancje zapachowe i aromatyzuj¹ce, barwniki, biopestycydy i dodatki do ¿ywnoœci. Szacuje siê, ¿e 25% leków stosowanych w krajach uprzemys³owionych zawiera zwi¹zki, które bezpoœrednio lub poœrednio (semisynteza) pochodz¹ z roœ- lin (1). Metabolity wtórne s¹ niskocz¹steczkowymi zwi¹zkami organicznymi o bardzo ró¿norodnej strukturze chemicznej. Nie PRACE PRZEGL¥DOWE Adres do korespondencji Halina Wysokiñska, Katedra Biologii i Biotechnologii Farmaceutycznej, Uniwersytet Medyczny, ul. Muszyñskiego 1, 90-151 £ódŸ; e-mail: [email protected] 4 (75) 124–135 2006
Transcript of Produkcja roœlinnych metabolitów wtórnych w kulturach ... · BIOTECHNOLOGIA 4 (75) 124-135 2006...

Produkcja roœlinnych metabolitówwtórnych w kulturach organówtransformowanych
Halina Wysokiñska, Aleksander Chmiel
Katedra Biologii i Biotechnologii Farmaceutycznej,Uniwersytet Medyczny, £ódŸ
Secondary metabolites production in cultures of transformed plantorgans
S u m m a r y
Plant cell cultures in vitro produce secondary metabolites with varied effec-tiveness. Despite industrial application of only few cell suspension cultures,considerable progress in research on plant cell biotechnology has been madeover the last few years. Transformed organ cultures, especially hairy roots,seem to be an interesting model for stable production of plant metabolites withhigh yield. In this paper, cultures of hairy roots of Salvia sclarea, S. officinalis,S. miltiorrhiza, S. przewalskii, and Centaurium erythraea, as well as transformedshoots and plants of C. erythraea are presented. Also, production of secondarymetabolites in these cultures is discussed.
Key words:
plant cultures, transformed shoots, transformed roots, secondary metabo-lite production.
1. Wstêp
Roœlinne metabolity wtórne s¹ szeroko wykorzystywane jakofarmaceutyki, substancje zapachowe i aromatyzuj¹ce, barwniki,biopestycydy i dodatki do ¿ywnoœci. Szacuje siê, ¿e 25% lekówstosowanych w krajach uprzemys³owionych zawiera zwi¹zki,które bezpoœrednio lub poœrednio (semisynteza) pochodz¹ z roœ-lin (1). Metabolity wtórne s¹ niskocz¹steczkowymi zwi¹zkamiorganicznymi o bardzo ró¿norodnej strukturze chemicznej. Nie
P R A C E P R Z E G L ¥ D O W E
Adres do korespondencji
Halina Wysokiñska,Katedra Biologiii BiotechnologiiFarmaceutycznej,Uniwersytet Medyczny,ul. Muszyñskiego 1,90-151 £ódŸ;e-mail:[email protected]
4 (75) 124–135 2006

odgrywaj¹ one istotnej roli w podstawowych procesach ¿yciowych roœlin (wzrost,ró¿nicowanie, rozmna¿anie), ale mog¹ spe³niaæ inne wa¿ne funkcje, np. w interakcjiroœlina – œrodowisko. Liczba roœlinnych metabolitów wtórnych szacowana jest na100 000 (2) do 200 000 (3). Ka¿dego roku wykrywa siê oko³o 1600 nowych zwi¹z-ków (4), co jest zrozumia³e zwa¿ywszy, ¿e znanych jest oko³o 400 000 gatunkówroœlin wy¿szych, a dotychczas jedynie oko³o 10% z nich zbadano fitochemicznie. Je-den tylko gatunek Arabidopsis thaliana mo¿e wytwarzaæ 5000 ró¿nego typu metabo-litów (5,6). Dla porównania, liczba poznanych metabolitów wtórnych syntetyzowa-nych przez drobnoustroje oceniana jest na ponad 20 000, a przez zwierzêta naoko³o 2500 (7,8).
Metabolity roœlinne pozyskiwane s¹ najczêœciej przez ekstrakcjê z surowca roœ-linnego pochodz¹cego z uprawy. Jednak zawartoœæ tych zwi¹zków w roœlinach jestzazwyczaj niska (czêsto poni¿ej 1% suchej masy) i zmienna, uzale¿niona od warun-ków œrodowiska. Niedogodnoœci¹ mo¿e byæ tak¿e ograniczona dostêpnoœæ surowcaroœlinnego; w wielu przypadkach cenne zwi¹zki pochodz¹ z roœlin rzadko wystê-puj¹cych, chronionych lub rosn¹cych tylko w okreœlonych warunkach klimatycznych.W konsekwencji mo¿e to prowadziæ do niedoboru wa¿nych dla lecznictwa zwi¹z-ków, takich jak np. artemizyna – substancja o dzia³aniu przeciwmalarycznym wy-twarzana przez Artemisia annua (bylica roczna) (9). Wszystkie te fakty przemawiaj¹za wykorzystaniem do wytwarzania cennych metabolitów roœlinnych metod bio-technologicznych. Do zalet tych metod nale¿¹ m.in. kontrolowane warunki hodowliumo¿liwiaj¹ce otrzymanie jednorodnego materia³u roœlinnego, uniezale¿nienie siêod stref klimatycznych i pór roku, a tak¿e mo¿liwoœæ znacznego zwiêkszenia pro-duktywnoœci przez selekcjê wysoko wydajnych linii lub elicytacjê. Nie bez znaczeniajest fakt, ¿e kultury in vitro mog¹ syntetyzowaæ nowe zwi¹zki, nie wykryte dotych-czas w œwiecie roœlin. Gräther i Schneider (10) podaj¹, ¿e do po³owy 1999 r. z kulturin vitro wyizolowano i okreœlono strukturê 322 nowych zwi¹zków, z których wiêk-szoœæ nale¿y do terpenoidów (100), alkaloidów (72) i fenoli (144). Siedemnaœcie no-wych taksanów wykryto w wyniku intensywnych badañ fitochemicznych kulturin vitro roœlin z rodzaju Taxus (10). Najwiêcej nowych zwi¹zków znaleziono w kultu-rach zawiesinowych, które s¹ najczêœciej wykorzystywanym modelem badawczym.Oko³o 100 nowych zwi¹zków wykryto w tkankach kalusowych, 30 w kulturach ko-rzeni nietransformowanych, a 20 w kulturach korzeni transformowanych.
Dotychczas niewielki jest udzia³ roœlinnych kultur in vitro w produkcji metaboli-tów wtórnych na skalê przemys³ow¹. Przyk³adem s¹ podjête pod koniec lat 80.XX w. przez japoñsk¹ firmê Nitto Denko Corporation, próby otrzymywania ginseno-zydów z kultur zawiesinowych Panax ginseng, które doprowadzi³y do opracowaniatechnologii realizowanej w bioreaktorach o pojemnoœci 2 m3, a nastêpnie 20 m3
(11). Innym przyk³adem jest przemys³owa produkcja paklitakselu w kulturach Taxusspp. W niemieckiej firmie Phyton Gesellschaft für Biotechnik mbH prowadzone by³ypróby w bioreaktorach o pojemnoœci 75 m3 (12). Brak jest potwierdzenia realizacjitej technologii. Tymczasem od kilku lat koreañska firma SamYang Genex Corp. ofe-
Produkcja roœlinnych metabolitów wtórnych w kulturach organów transformowanych
BIOTECHNOLOGIA 4 (75) 124-135 2006 125

ruje jako produkt handlowy lek Genexol, zawieraj¹cy paklitaksel z bioreaktorowychkultur zawiesinowych Taxus sp. (13).
Niewielkie wykorzystanie kultur in vitro do produkcji metabolitów wtórnych wy-nika m.in. z nie zawsze jeszcze wystarczaj¹cej ich wydajnoœci, ma³ej stabilnoœci ge-netycznej i biochemicznej, co wymaga ci¹g³ej selekcji i zwiêksza koszty wytwarza-nia produktu. Na koszty procesu niekorzystnie wp³ywa równie¿ fakt, ¿e komórkizwykle nie wydzielaj¹ po¿¹danych substancji do pod³o¿a, lecz gromadz¹ je w waku-olach czy innych organellach, w wyniku czego potrzebne jest poddanie uzyskanejbiomasy procesowi ekstrakcji (14). Niska wydajnoœæ lub brak zdolnoœci do biosynte-zy niektórych metabolitów w kulturach zawiesinowych ma czêsto zwi¹zek z bra-kiem zró¿nicowanych struktur. Ten problem mo¿na wyeliminowaæ stosuj¹c kulturyorganów.
Znaczny postêp w wykorzystaniu metod biotechnologicznych do wytwarzaniametabolitów sta³ siê mo¿liwy pod koniec lat 80. XX w., kiedy otrzymano kultury ge-netycznie transformowanych organów. Kultury takie rosn¹ szybciej i pozwalaj¹ osi¹-gaæ znacznie wy¿sze przyrosty biomasy ni¿ kultury organów nietransformowanych.Kultury organów transformowanych obejmuj¹ kultury pêdów tzw. teratomy pêdowei kultury korzeni transformowanych, zwanych równie¿ w³oœnikowatymi lub transge-nicznymi.
2. Kultury transformowanych pêdów
Kultury transformowanych pêdów otrzymuje siê w wyniku transformacji tkanekroœlinnych niektórymi szczepami Agrobecterium tumefaciens. Najczêœciej wykorzysty-wane s¹ oktopinowe i nopalinowe szczepy z genem izopentylotransferazy (gen ipt)(15). Produkt tego genu zmienia w komórkach stosunek auksyn do cytokinin, w wy-niku czego na eksplantatach powstaje tkanka tumorowa, z której regeneruj¹ siêpêdy. Ich morfologia jest zmieniona w porównaniu z pêdami normalnymi; okreœlasiê je czêsto mianem teratom pêdowych. Dotychczas teratomy pêdowe otrzymanodla niewielkiej liczby roœlin; nale¿¹ do nich m.in. przedstawiciele rodziny Solanaceae:Atropa belladonna i Duboisia leichnardhi x D. myoporoides. Obie kultury maj¹ zdolnoœæbiokonwersji hioscyjaminy w skopolaminê (16). Teratomy pêdowe otrzymano rów-nie¿ z Pimpinella anisum (17), Mentha citrata, Coleus forskohlii, Nicotiana tabacum (18)i Artemisia annua (19). Biosynteza metabolitów wtórnych w tych kulturach jest zró¿-nicowana. Teratomy pêdowe Solanum dulcamara syntetyzuj¹ glikoalkaloidy w iloœci1% suchej masy, tj. 5 razy wiêcej ni¿ roœlina macierzysta (20).
W kulturze transformowanych pêdów A. annua (19) wykryto ponad 0,06% artemi-zyny w przeliczeniu na such¹ masê, podczas gdy hodowane in vitro pêdy nietransfor-mowane wytwarzaj¹ zaledwie 0,02% tego izoprenoidu. Jednak¿e obie te iloœci s¹znacznie ni¿sze ni¿ w liœciach roœliny macierzystej, które zawieraj¹ prawie 1% arte-mizyny (21).
Halina Wysokiñska, Aleksander Chmiel
126 PRACE PRZEGL¥DOWE

3. Kultury transformowanych korzeni
Znacznie czêœciej ni¿ teratomy pêdowe dla wytwarzania metabolitów wtórnychwykorzystywane s¹ kultury korzeni w³oœnikowatych. Korzenie te otrzymuje siêw wyniku transformacji genetycznej szczepami Agrobacterium rhizogens. Integracjaz genomem komórki roœliny fragmentu plazmidu Ri (T-DNA) z komórki bakteryjneji ekspresja zawartych w tym fragmencie DNA genów (m.in. rol genów a, b, c, d) pro-wadzi do powstania na eksplantacie, w miejscu zaka¿enia, korzeni przybyszowych.Korzenie te po oddzieleniu od eksplantatu i eliminacji bakterii mog¹ rosn¹æ w kol-bach lub ró¿nego typu bioreaktorach, na sta³ym lub w p³ynnym pod³o¿u bez dodat-ku regulatorów wzrostu. W przeciwieñstwie do kultur komórkowych, kultury korze-ni w³oœnikowych charakteryzuj¹ siê znaczn¹ stabilnoœci¹ i mog¹ utrzymywaæ wy-dajn¹ biosyntezê metabolitów podczas d³ugotrwa³ego pasa¿owania (22). Produkcjametabolitów w tych kulturach jest zbli¿ona lub wy¿sza ni¿ w korzeniach roœliny ma-cierzystej. W tabeli 1 przedstawiono zebrane z ostatnich dziesiêciu lat przyk³adykultur korzeni transformowanych produkuj¹cych znaczne iloœci metabolitów wtór-nych.
T a b e l a 1
Przyk³ady wysoko produktywnych kultur korzeni transformowanych
Roœlina Szczep A. rhizogenes Metabolity Literatura
Linum flavum LBA 9402, TR 105 koniferyna, 58 mg/g s.m. (23)
Lupinus mutabilis AR1601 izoflawony, 2× wiêcej ni¿ korzenie nietransformowane (24)
Camptotheca acuminata ATCC 15834,R-100
kamtotecyna, 1 mg/g s.m.hyroksykamptotecyna, 0,15 mg/g s.m.
(25)
Solanum mauritanum LBA 9402 solasodyna, 184 �g/g s.m.6× wiêcej ni¿ korzenie roœliny macierzystej
(26)
Puerearia phasoloides ATCC 15834 pueraryna, 1,19 mg/g s.m. (w kolbach) i 5,57 mg/g s.m.(w bioreaktorze)5× wiêcej ni¿ w korzeniach nietransformowanych
(27)(28)
Tylophora indica A4 tyloforyna2× wiêcej ni¿ w korzeniach roœlin nietransformowa-nych
(29)
Physalis minima ATCC 15834 glukozyd solasodyny,9,11 �g/ g s.m., 20× wiêcej ni¿ korzenie i 3× wiêcejni¿ liœcie
(30)
Ocimum basilicum MAFF 03-01724 kwas rozmarynowy, 14% s.m.3,5× wiêcej ni¿ liœcie
(31)
Valerianella lacustra A4, ATCC 15834 walepotriaty (po elicytacji MeJA), 50× wiêcej ni¿ w roœ-linie
(32)
Ambrosia maritima ATCC 15834 poliacetyleny, 9,6× wiêcej ni¿ w kulturze zawiesinowej (33)
Produkcja roœlinnych metabolitów wtórnych w kulturach organów transformowanych
BIOTECHNOLOGIA 4 (75) 124-135 2006 127

4. Prace w³asne
Wytwarzanie metabolitów wtórnych w kulturach korzeni transformowanych jestrównie¿ przedmiotem badañ w naszej Katedrze. Spoœród otrzymanych dotychczaskultur (tab. 2) omówione zostan¹ kultury badane w ostatnich latach.
T a b e l a 2
Kultury korzeni transformowanych otrzymane w Katedrze Biologii i Biotechnologii Farmaceutycznej
Gatunek Rodzina Szczep A. rhizogenes Metabolity
Catalpa ovata Bignoniaceae LBA 9402 werbaskozyd, izowerbaskozyd
Hyssopus officinalis Lamiaceae LBA 9402 kwas rozmarynowy
Paulownia tomentosa Bignoniaceae LBA 9402 werbaskozyd
Arnica montana Asteraceae LBA 9402 pochodne tymolu, olejek eteryczny, kwas chlorogenowy
Salvia sclarea Lamiaceae LBA 9402 diterpeny, triterpeny, sterole, kwas rozmarynowy, tan-szinonyS. officinalis Lamiaceae A4, ATCC 15834
S. miltiorrhiza Lamiaceae A4
S. przewalskii Lamiaceae A4
Centaurium erythraea Gentianaceae LBA 9402 glikozydy sekoirydoidowe
4.1. Korzenie transformowane S. sclarea
Korzenie transformowane S. sclarea (sza³wia muszkato³owa) otrzymano po zaka-¿eniu wyhodowanych in vitro pêdów szczepem Agrobacterium rhizogenes LBA9402(34). Kultury prowadzono w p³ynnym pod³o¿u ½B5 (pod³o¿e wg Gamborga, zezmniejszon¹ do po³owy zawartoœci¹ makro- i mikroelementów) uzupe³nionym sa-charoz¹ (30 g/l), w kolbach Erlenmeyera, w ciemnoœci i na œwietle (40 �mol m-2s-1).Z tych kultur wyizolowano cztery abitanowe diterpeny, które zidentyfikowano jako:salwipison, ferruginol, etiopinon i 1-ketoetiopinon (rys. 1). Wykazano, ¿e kulturykorzeni prowadzone na œwietle wytwarzaj¹ 2,4 razy wiêcej diterpenów (25,37 mg/gs.m.) ni¿ korzenie hodowane w ciemnoœci (10,9 mg/g s. m.). W tych pierwszych za-wartoœæ diterpenów by³a 11-krotnie wy¿sza ni¿ w kulturach korzeni nietransformo-wanych i 8 razy wy¿sza ni¿ w korzeniach roœliny macierzystej. Dodatkowo korzeniew³oœnikowate hodowane na œwietle osi¹gaj¹ maksymalny poziom diterpenów 30.dnia cyklu wzrostu, czyli o 5 dni szybciej ni¿ korzenie prowadzone w ciemnoœci(34).
Halina Wysokiñska, Aleksander Chmiel
128 PRACE PRZEGL¥DOWE

Wp³yw œwiat³a na produkcjê metabolitów w kulturach korzeni transformowa-nych by³ obserwowany przez wielu badaczy. Œwiat³o stymulowa³o np. wytwarzaniesteroidowych saponin (akuleatyzydu A i B) w kulturach korzeni w³oœnikowatychSolanum aculeatissmum (35) i zwiêksza³o produkcjê solasodyny (aglikon glikoalkalo-idów wykorzystywany do pó³syntezy hormomów p³ciowych i kortykosteroidów)w kulturach korzeni transformowanych Solanum khasianum (36). Jednak¿e Argoloi wsp. (37) stwierdzili, ¿e rosn¹ce w ciemnoœci kultury korzeni w³oœnikowatych Solanumaviculare akumuluj¹ 4,2 razy wiêcej solasodyny w porówaniu z kulturami hodowany-mi na œwietle. Wysoka produkcja diterpenów w prowadzonych na œwietle korze-niach w³oœnikowatych S. sclarea ma prawdopodobnie zwi¹zek z obecnoœci¹ chloro-plastów, które przez wielu autorów proponowane s¹ jako miejsce biosyntezy tegotypu zwi¹zków (38). Dominuj¹cym diterpenem w korzeniach transformowanychS. sclarea jest etiopinon, którego zawartoœæ w kulturach prowadzonych na œwietlei w ciemnoœci stanowi, odpowiednio, 45 i 37% sumy diterpenów. Jest to wa¿nebior¹c pod uwagê wyniki badañ biologicznych, na podstawie których wykazano, ¿eetiopinon charakteryzuje siê znaczn¹ aktywnoœci¹ przeciwbakteryjn¹ (39,40) i udo-wodnion¹ w badaniach in vitro aktywnoœci¹ cytotoksyczn¹ (40) oraz zdolnoœci¹ doindukcji apoptozy (41). Inny diterpen o dzia³aniu bakteriostatycznym i bakteriobój-czym wobec niektórych bakterii gramdodatnich – salwipison (39), wytwarzany jestw kulturach korzeni transformowanych w znacznie ni¿szych iloœciach (3,1 mg/gs. m. w korzeniach hodowanych na œwietle i 2,5 mg/g s. m. w kulturach rosn¹cychw ciemnoœci).
Kultury korzeni transformowanych S. sclarea prowadzono równie¿ w 10-litro-wym bioreaktorze aeroponicznym (fot. 1). Po 30. dniach hodowli obserowano16-krotny przyrost œwie¿ej i i 14-krotny suchej masy w stosunku do inokulatu. Oka-za³o siê jednak, ¿e te warunki nie sprzyjaj¹ wytwarzaniu diterpenów, których zawar-toœæ by³a 2,2 raza ni¿sza ni¿ w korzeniach rosn¹cych w kolbach Erlenmeyera.
Z korzeni transformowanych S. sclarea oprócz diterpenów wyizolowano jeszczedwa pentacykliczne triterpeny, di- i trihydroksypochodne kwasu ursolowego: kwas
Produkcja roœlinnych metabolitów wtórnych w kulturach organów transformowanych
BIOTECHNOLOGIA 4 (75) 124-135 2006 129
Rys. 1. Diterpeny w kulturze korzeni transformowanych S. sclarea: 1) salwipison (12-hydroksy--4,5-seko-5,10-friedo-4(18),5(10),6,8,12-abietapentaen-11,14-dion), 2) ferruginol (12-hydroksyabieta--8,11,13-trien), 3) etiopinon (4,5-seko-5,10-friedo-abieta-4(18),5,6,8,13-pentaen-11,12-dion), 4) 1-keto-etiopinon (1-keto-4,5-seko-5,10-friedo-abieta-4(18),5,6,8,13-pentaen-11,12-dion).

2�, 3�,-dihydroksyursolowy i kwas 2�, 3�, 24-trihydroksyursolowy (rys. 2). Pierw-szy z nich wyizolowano z korzeni hodowanych na œwietle, drugi produkuj¹ korzenieprowadzone w ciemnoœci. W obu kulturach, metod¹ GC-MS, wykryto kwas ursolowyi kwas oleanolowy oraz trzy sterole (�-sitosterol, stigmasterol i kampesterol (34).
Halina Wysokiñska, Aleksander Chmiel
130 PRACE PRZEGL¥DOWE
Fot. 1. Korzenie transformowane S. sclarea w bioreaktorze.

4.2. Korzenie transformowane S. miltiorrhiza i S. przewalskii
Salvia miltiorrhiza (sza³wia czerwonokorzeniowa) jest wa¿n¹ roœlin¹ lecznicz¹ wy-korzystywan¹ g³ównie w Azji. Surowcem s¹ korzenie tej roœliny, a g³ównymi sk³adnika-mi diterpeny, typu abietanu, z ugrupowaniem fantrenochinonowym, zwane tanszino-nami. Diterpeny te stosowane s¹ g³ównie w chorobach uk³adu kr¹¿enia. Stwierdzono,¿e korzenie sza³wii czerwonokorzeniowej transformowane szczepem A. rhizogenes A4,i hodowane w ciemnoœci, w p³ynnym pod³o¿u ½B5 wytwarzaj¹ cztery tanszinony: tan-szinon I, tanszinon IIA, kryptotanszinon i dihydrokryptotanszinon. Suma tych diterpe-nów wynosi³a prawie 18 mg w przeliczeniu na gram suchej masy i by³a zbli¿ona do za-wartoœci tych zwi¹zków w korzeniach 2-letnich roœlin rosn¹cych w gruncie (16,2 mg/gs.m.) i a¿ 5-krotnie wy¿sza ni¿ w kulturze korzeni nietransformowanych (3,7 mg/gs.m.), hodowanych równie¿ w pod³o¿u ½B5, ale z dodatkiem auksyny (IAA 0,1 mg/l).W kulturze korzeni transformowanych S. miltiorrhiza dominowa³ kryptotanszinon, któ-rego zawartoœæ wynosi³a 5,8 mg/g s.m., czyli by³a dwukrotnie wy¿sza od zawartoœcitego diterpenu w korzeniach roœliny macierzystej (2,6 mg/g s.m.).
Tanszinony wytwarzane s¹ równie¿ w korzeniach roœlin z gatunku S. przewalskii(sza³wia przewalskiego). Jest to endemit wystêpuj¹cy w po³udniowo-wschodnichChinach. Jednak¿e otrzymane po transformacji szczepem A. rhizogenes A4, korzeniew³oœnikowate tej roœliny wytwarzaj¹ jedynie niewielkie iloœci diterpenów; suma tan-szinonu IA, tanszinonu IIA, kryptotanszinonu i dihydrotanszinonu wynosi³a ok.
Produkcja roœlinnych metabolitów wtórnych w kulturach organów transformowanych
BIOTECHNOLOGIA 4 (75) 124-135 2006 131
Rys. 2. Metabolity triterpenowe w kulturze korzeni transformowanych S. sclarea: 1) kwas ursolowy,2) kwas oleanolowy, 3) kwas 2�,3�-dihydroksy-urs-12-en-28-owy, 4) kwas 2�,3�,24-trihydrok-sy-urs-12-en-28-owy.
R1 R2 R3 R4 R5
1 H �OH CH3 CH3 H2 H �OH CH3 H CH3
3 �OH �OH CH3 CH3 H4 �OH �OH CH2OH CH3 H

4 mg/g s.m. i by³a prawie 4-krotnie ni¿sza ni¿ w korzeniach 2-letnich roœlin ros-n¹cych w gruncie. Niska zawartoœæ diterpenów mo¿e mieæ zwi¹zek z morfologi¹i stadium rozwojowym hodowanych in vitro korzeni; by³y one cienkie, ¿ó³tawe o bu-dowie pierwotnej. Natomiast na podstawie obserwacji z korzeniami zebranymiz roœlin hodowanych w gruncie wykazano, ¿e miejscem akumulacji tanszinonów jestg³ównie zewnêtrzna skorkowacia³a czêœæ korowa korzenia.
4.3. Korzenie transformowane S. officinalis
Korzenie transformowane S. officinalis (sza³wia lekarska) otrzymano w wyniku za-ka¿enia pêdów bakteriami A. rhizogenes A4 i ATCC15834. Uzyskano ok. 10 klonów,które charakteryzowa³y siê szybkim wzrostem w pod³o¿u Lloyda i McCowna. Stwier-dzono, ¿e korzenie te s¹ dobrym Ÿród³em kwasu rozmarynowego (depsyd kwasu ka-wowego i �-hydroksydihydrokawowego), zwi¹zku o dzia³aniu przeciwutleniaj¹cym,przeciwzapalnym i antywirusowym. Produkcja kwasu rozmarynowego waha³a siê od16 do 33 mg/g suchej masy. Najbardziej produktywny klon wytwarza³ dwa razy wiê-cej tego zwi¹zku ni¿ kultury korzeni nietransformowanych (19 mg/g s.m.). Zawar-toœæ kwasu rozmarynowego zale¿a³a od szczepu Agrobacterium u¿ytego do transfor-macji i analizowanego klonu (42). Stwierdzono, ¿e klony uzyskane w wyniku trans-formacji szczepem ATCC 15834 wytwarzaj¹ wiêcej kwasu rozmarynowego ni¿ klonytransformowane szczepem A4. Rodzaj u¿ytego szczepu wp³ywa³ równie¿ na wzrosti morfologiê korzeni (fot. 2).
Halina Wysokiñska, Aleksander Chmiel
132 PRACE PRZEGL¥DOWE
Fot. 2. Wygl¹d korzeni w³oœnikowatych S. officinalis, otrzymanych po transformacji szczepamiA. rhizogenes ATCC15834 i A4; skala = 1 cm.

Zale¿noœæ miêdzy produkcj¹ metabolitów wtórnych a rodzajem szczepu bakte-ryjnego by³a ju¿ wczeœniej obserwowana w kulturach korzeni transformowanychHyoscyamus albus (43). W naszych badaniach wykazano równie¿ ró¿nice w biosynte-zie kwasu rozmarynowego miêdzy klonami zapocz¹tkowanymi tym samym szcze-pem Agrobacterium. Mog¹ one wynikaæ z ró¿nic w wielkoœci i lokalizacji fragmentówT-DNA zintegrowanych z genomem komórki roœlinnej (44,45). Nie zaobserwowanonatomiast istotnego wp³ywu warunków hodowli (œwiat³o, ciemnoœæ) na wytwarzaniekwasu rozmarynowego w kulturach korzeni transformowanych S. officinalis.
4.4. Korzenie i pêdy transformowane C. erythraea
Centaurium erythraea (tysi¹cznik pospolity) jest znany z biosyntezy m.in. glikozy-dów sekoirydoidowych, g³ównie gentiopikrozydu, swerozydu i swertiamaryny. Zwi¹z-ki te wykazuj¹ dzia³anie przeciwgrzybowe, przeciwbakteryjne, ¿ó³ciopêdne i hepa-tochronne (46). Stosowane s¹ równie¿ w przemyœle spo¿ywczym, jako sk³adniki li-kierów i napojów energetyzuj¹cych. W korzeniach transformowanych C. erythraearosn¹cych w ci¹gu 4 tygodni w p³ynnym pod³o¿u Lloyda i McCowna (z sacharoz¹30g/l) znaleziono 10-14 mg/g s.m. gentiopikrozydu, swerozydu i swertiamaryny (47).Wœród nich dominowa³ gentiopikrozyd, który stanowi³ 80% sumy wykrytych sekoiry-doidów. W porównaniu z korzeniami roœlin rosn¹cych w gruncie zawartoœæ sekoiry-doidów w kulturach korzeni w³oœnikowatych by³a 2-krotnie wy¿sza, ale znacznieni¿sza ni¿ w pêdach roœliny, które s¹ g³ównym miejscem akumulacji tego typuzwi¹zków. W preparacie handlowym Centaurii herba, który zawiera³ kwitn¹ce pêdytysi¹cznika wykryto ok. 37 mg glikozydów sekoirydoidowych w przeliczeniu na 1 gsuchego surowca.
Zdolnoœæ korzeni w³oœnikowatych C. erythraea do organogenezy i regeneracjip¹ków wykorzystano aby otrzymaæ kultury transgenicznych pêdów. Kultury tych pê-dów hodowane na sta³ym lub w p³ynnym pod³o¿u Murashige i Skooga uzupe³nio-nym IAA (0,1 mg/l) i BAP (1 mg/l) wytwarza³y 34-48 mg/g s.m. gentiopikrozydu, swe-rozydu i swertiamaryny. Podobnie jak w korzeniach, przewa¿a³ gentiopikrozyd (65%sumy sekoirydoidów), chocia¿ w pêdach rosn¹cych in vivo zawsze dominuje swertia-maryna. Dalsze zwiêkszenie zawartoœci sekoirydoidów otrzymano po regeneracjica³ych transgenicznych roœlin. Roœliny te ró¿ni³y siê morfologicznie od roœlin nie-transformowanych (syndrom w³oœnikowatoœci). Ró¿nice dotyczy³y liczby rozga³ê-zieñ pêdu, morfologii liœci i systemu korzeniowego oraz budowy kwiatostanów(krótsze, bardziej rozga³êzione, z wiêksz¹ liczb¹ p¹ków kwiatowych). Najwiêcej se-koirydoidów wytwarzaj¹ 10-tygodniowe roœliny rosn¹ce w doniczkach (tab. 3). Wy-kryto w nich 280 mg/g s.m. gentiopikoryzydu, swerozydu i swertiamaryny, czyli po-nad 7-krotnie wiêcej ni¿ w surowcu Centaurii herba.
Produkcja roœlinnych metabolitów wtórnych w kulturach organów transformowanych
BIOTECHNOLOGIA 4 (75) 124-135 2006 133
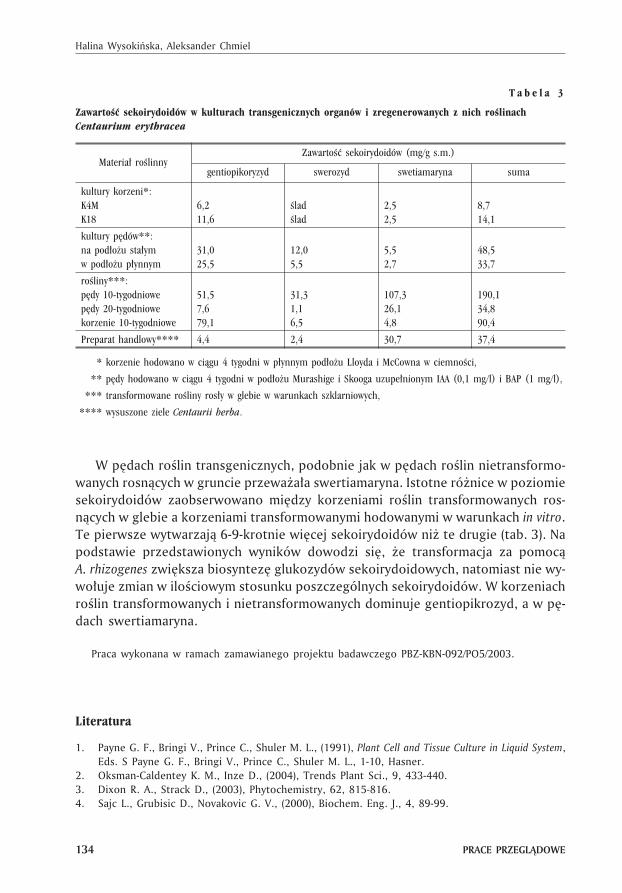
T a b e l a 3
Zawartoœæ sekoirydoidów w kulturach transgenicznych organów i zregenerowanych z nich roœlinachCentaurium erythracea
Materia³ roœlinnyZawartoœæ sekoirydoidów (mg/g s.m.)
gentiopikoryzyd swerozyd swetiamaryna suma
kultury korzeni*:K4MK18
6,211,6
œladœlad
2,52,5
8,714,1
kultury pêdów**:na pod³o¿u sta³ymw pod³o¿u p³ynnym
31,025,5
12,05,5
5,52,7
48,533,7
roœliny***:pêdy 10-tygodniowepêdy 20-tygodniowekorzenie 10-tygodniowe
51,57,679,1
31,31,16,5
107,326,14,8
190,134,890,4
Preparat handlowy**** 4,4 2,4 30,7 37,4
**** korzenie hodowano w ci¹gu 4 tygodni w p³ynnym pod³o¿u Lloyda i McCowna w ciemnoœci,
**** pêdy hodowano w ci¹gu 4 tygodni w pod³o¿u Murashige i Skooga uzupe³nionym IAA (0,1 mg/l) i BAP (1 mg/l),
**** transformowane roœliny ros³y w glebie w warunkach szklarniowych,
**** wysuszone ziele Centaurii herba.
W pêdach roœlin transgenicznych, podobnie jak w pêdach roœlin nietransformo-wanych rosn¹cych w gruncie przewa¿a³a swertiamaryna. Istotne ró¿nice w poziomiesekoirydoidów zaobserwowano miêdzy korzeniami roœlin transformowanych ros-n¹cych w glebie a korzeniami transformowanymi hodowanymi w warunkach in vitro.Te pierwsze wytwarzaj¹ 6-9-krotnie wiêcej sekoirydoidów ni¿ te drugie (tab. 3). Napodstawie przedstawionych wyników dowodzi siê, ¿e transformacja za pomoc¹A. rhizogenes zwiêksza biosyntezê glukozydów sekoirydoidowych, natomiast nie wy-wo³uje zmian w iloœciowym stosunku poszczególnych sekoirydoidów. W korzeniachroœlin transformowanych i nietransformowanych dominuje gentiopikrozyd, a w pê-dach swertiamaryna.
Praca wykonana w ramach zamawianego projektu badawczego PBZ-KBN-092/PO5/2003.
Literatura
1. Payne G. F., Bringi V., Prince C., Shuler M. L., (1991), Plant Cell and Tissue Culture in Liquid System,Eds. S Payne G. F., Bringi V., Prince C., Shuler M. L., 1-10, Hasner.
2. Oksman-Caldentey K. M., Inze D., (2004), Trends Plant Sci., 9, 433-440.3. Dixon R. A., Strack D., (2003), Phytochemistry, 62, 815-816.4. Sajc L., Grubisic D., Novakovic G. V., (2000), Biochem. Eng. J., 4, 89-99.
Halina Wysokiñska, Aleksander Chmiel
134 PRACE PRZEGL¥DOWE

5. von Roepenack-Lahaye E., Degenkolb T., Zerjeski M., Franz M., Roth U., Wessjohann L., Schmidt J.,Schell D., Clemens S., (2004), Plant Physiol., 134, 548-559.
6. Facchini P. J., Bird D. A., St-Pierre B., (2004), Trends Plant Sci., 9,116-122.7. Berdy J., (2005), J. Antibiol., 58, 1-26.8. Oksman–Caldentey K. M., Saito K., (2005), Curr. Opin. Biotechnol., 16, 174-179.9. Cyranoski D., (2004), Nature, 432, 259.
10. Gräther O., Schneider B., (2001), Progress in Botany, 62, 266-304.11. Vansree M., Chen-Yue L., Shu-Fung L., Satiish M., N., Chien Y., L., Hsin-Sheng T., (2004), Bot. Bull.
Acad. Sin., 45, 1-22.12. Doran P. M., (1993), Adv. Biochem. Engin., 98, 115.13. http://www.genexol.com./eng/
14. Bourgaud F., Gravot A., Milesi S., Gonteir E., (2001), Plant Sci., 161, 839-851.15. Floryanowicz-Czekalska K., Wysokiñska H., (2000), Acta Soc. Bot. Polon., 69, 131-136.16. Subroto M. A., Hamill J. D., Doron P. M., (1996), J. Biotechnol., 45, 45-57.17. Salem K. M., Charlwood B. V., (1995), Plant Cell Tissue Organ Cult., 40, 209-215.18. Jha S., (1995), Proc. Indian. Natn. Sci. Acad., 61, 63-72.19. Ghosh B., Mukherjee S., Jha S., (1997), Plant Sci., 122, 193-199.20. Ehmke A., Ohmstede D., Eilert U., (1995), Plant Cell Tissue Organ Cult., 43, 191-197.21. Paniego W. B, Giulietti A. M., (1996), Enzyme Microb. Technol., 18, 526-530.22. Wysokiñska H., Chmiel A., (1997), Acta Biotechnol., 17, 131-159.23. Lin H., Kwok K. H., Doran P. M., (2003), Biotechnology Lett., 25, 521-525.24. Babaoglu M., Davey M. R., Power J. B., Sporer F., Wink M., (2004), Plant Cell Tissue Organ Cult., 78,
29-36.25. Lorence A., Medina-Bolivar F., Nessler C. L., (2004), Plant Cell Rep., 22, 437-441.26. Drews F. E., von Staden J., (1995), Plant Growth Regulation, 17, 27-31.27. Shi H. P., Kintzios S., (2003), Plant Cell Rep., 21, 1103-1107.28. Kintzios S., Makri O., Pistola E., Matakiadis T., Shi H. P., Economou A., (2004), Biotechnology Lett.,
26, 1057-1059.29. Chaudhuri K. N., Ghosh B., Tepfer D., Jha S., (2005), Plant Cell Rep., 24, 25-35.30. Putalin W., Prasarnsiwamai P., Tanaka H., Shoyama Y., (2004), Biotechnology Lett., 26, 545-548.31. Tada H., Murakamii Y., Omoto T., Shimomura K., Ishimaru K., (1996), Phytochemistry, 42, 431-434.32. Kittipongpatana N., Davis D. L., Poter J. R., (2002), Plant Cell Tissue Organ Cult., 71, 65-75.33. Zid S. A., Orihara Y., (2005), Plant Cell Tissue Organ Cult., 81, 65-75.34. KuŸma £., Skrzypek Z., Wysokiñska H., (2006), Plant Cell Tissue Organ Cult., 84, 152-160.35. Ikenaga T., Oyama T., Maranaka T., (1995), Plant Cell Rep., 14, 413-417.36. Jacob A., Malpathak W., (2004), Current Science, 87, 1442-1447.37. Argolo A. C., Charlwood B. V., Pletsch M., (2000), Planta Med., 66, 448-451.38. McGarvey D. J., Croteau R., (1995), Plant Cell, 7, 1015-1026.39. KuŸma £., Ró¿alski M., Walencka E., Ró¿alska B., Wysokiñska H., (2006), Phytomed., (w druku).40. Hernandez-Perez M., Rabanal R. M., Arias A., de la Torre M. C., Rodriguez B., (1997), Pharm. Biol.,
37, 1-6.41. Ró¿alski M., KuŸma £., Krajewska U., Wysokiñska H., (2006), Z. Naturforsch., 61c, 483-488.42. Grzegorczyk I., Królicka A., Wysokiñska H., (2006), Z. Naturforsch., 61c, 351-356.43. Zehra M., Benerjee S., Sharma S., Kumor S., (1999), Planta Med., 65, 60-63.44. Mano Y., Nabeshima S., Matsui C., Ohkawa H., (1986), Agric. Biol. Chem., 50, 2715-2722.45. Batra J., Dutta A., Singh D., Kumar S., Sen J., (2004), Plant Cell Rep., 23, 148-154.46. Kumarasamy Y., Nahar L., Cox P. J., Jaspars M., Saker S. D., (2003), Phytomed., 10, 344-347.47. Pi¹tczak E., Królicka A., Wysokiñska H., (2006), Plant Cell Rep., (w druku).
Produkcja roœlinnych metabolitów wtórnych w kulturach organów transformowanych
BIOTECHNOLOGIA 4 (75) 124-135 2006 135


















