
Mikrobiologia medyczna i środowiskowa wybrane...
243
Mikrobiologia medyczna i środowiskowa – wybrane zagadnienia
Transcript of Mikrobiologia medyczna i środowiskowa wybrane...

Mikrobiologia medyczna
i środowiskowa
– wybrane zagadnienia


Mikrobiologia medyczna
i środowiskowa
– wybrane zagadnienia
Redakcja:
Monika Maciąg
Kamil Maciąg
Lublin 2017

Recenzenci:
prof. dr hab. Jolanta Rzymowska
prof. dr hab. n. farm. Monika Waksmundzka-Hajnos
dr hab. Jolanta Jaroszuk-Ściseł
dr hab. Urszula Kosikowska
dr hab. inż. Zbigniew Suchorab
dr hab. Sylwia Wdowiak-Wróbel
dr Artur Banach
dr inż. Agata Czernigowska
dr inż. Agata Dziubińska
dr Monika Jach
dr Elżbieta Kochanowicz
dr n. med. Małgorzata Kozioł
dr Jolanta Kutkowska
dr Renata Matraszek-Gawron
dr Anna Pytlak
dr Monika Pytka
dr Wojciech Radzki
dr inż. Amelia Staszowska
Wszystkie opublikowane rozdziały otrzymały pozytywne recenzje.
Skład i łamanie:
Monika Maciąg
Projekt okładki:
Marcin Szklarczyk
© Copyright by Wydawnictwo Naukowe TYGIEL sp. z o.o.
ISBN 978-83-65598-77-6
Wydawca:
Wydawnictwo Naukowe TYGIEL sp. z o.o.
ul. Głowackiego 35/341, 20-060 Lublin
www.wydawnictwo-tygiel.pl

Spis treści
Matylda Barbara Mielcarska, Magdalena Bossowska-Nowicka Status badań i rozwoju szczepionek przeciw zakażeniom herpeswirusami typu 1 i 2
(HSV-1, HSV-2) ............................................................................................................................ 7
Marta Kozieł, Karolina Włodarczyk, Dominika Wojton
Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne ............. 14
Andrzej Jurkowski, Ita Szczepańska, Jacek J. Kozioł
Wpływ nanocząstek tlenku żelaza na produkcję kwasu mlekowego w hodowli okresowej
Lactobacillus rhamnosus GG ..................................................................................................... 28
Rafał Kuś, Monika Hałgas, Gary van Lue, Francisco Gonzalez Vizcarra, Małgorzata M.
Kozioł, Alina Olender
Występowanie Staphylococcus aureus w jamie nosowo-gardłowej a czynniki ryzyka
wpływające na kolonizację ......................................................................................................... 36
Dagmara Dobrzyńska, Adrianna Dubisz, Katarzyna Marchwińska
Ocena wrażliwości na wybrane antybiotyki bakterii Staphylococcus spp. wyizolowanych
z powietrza .................................................................................................................................... 45
Agnieszka Ludwiczak
Badania wpływu wybranych związków na aktywność kinazy adenylanowej z Bacillus
stearothermophilus....................................................................................................................... 54
Magdalena Bossowska-Nowicka, Felix N. Toka, Lidia Szulc-Dąbrowska
Rola katepsyn w zakażeniach bakteryjnych .............................................................................. 64
Adriana Pacia, Małgorzata Nabrdalik, Agnieszka Dołhańczuk-Śródka
Ocena aktywności przeciwdrobnoustrojowej olejku geraniowego ....................................... 71
Ewa Kochan, Hanna Zielińska-Bliźniewska, Monika Sienkiewicz
Przeciwdrobnoustrojowe działanie saponin .............................................................................. 80
Monika Sienkiewicz, Hanna Zielińska-Bliźniewska, Jurek Olszewski, Paweł Kwiatkowski,
Ewa Kochan
Aktywność olejków eterycznych przeciwko lekoopornym patogenom odpowiedzialnym
za zakażenia dermatologiczne .................................................................................................... 92
Michał Piotrowski, Hanna Pituch, Piotr Obuch-Woszczatyński
Działanie miodu manuka na wzrost oraz biofilm wybranych bakterii stanowiących
problem terapeutyczny ..............................................................................................................105
Jana Przekwas, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis ..112

Anna Minkiewicz, Kamila Strom, Bartłomiej Iwańczyk, Piotr Wychowański, Sylwia
Jarzynka,, Gabriela Olędzka
Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych ........................................................................................ 126
Julia Szułczyńska
Wykorzystanie preparatów zawierających nanocząsteczki
do usuwania biofilmu bakteryjnego ......................................................................................... 138
Aldona Gajek, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis
izolowane z ran przewlekłych .................................................................................................. 148
Jolanta Małajowicz, Ewa Majewska, Dorota Nowak
Porównanie wydajności syntezy gamma-dekalaktonu w hodowlach ciągłych mutanta
drożdży Yarrowia lipolytica MTLY40-2p .............................................................................. 160
Magdalena Kręcidło, Teresa Krzyśko-Łupicka
Rozprzestrzenianie się bioaerozoli mikrobiologicznych – przegląd badań naukowych .... 171
Ewa Brągoszewska,Monika Rajtor, Józef S. Pastuszka
Porównanie wartości stężeń oraz wyników identyfikacji aerozolu bakteryjnego,
otrzymanych przy użyciu dwóch różnych aspiratorów ......................................................... 182
Sylwia Magdalena Duda, Monika Garbacz
Ocena aktywności mikroorganizmów z wykorzystaniem matrycy czujników gazu ......... 190
Monika Garbacz, Sylwia Magdalena Duda
Syndrom chorego budynku oraz jego wpływ na zdrowie mieszkańców
w kontekście mikrobiologii środowiska wewnętrznego ........................................................ 200
Marta Wyzińska, Jerzy Grabiński, Marzena Mikos-Szymańska
Porażenie kłosów przez grzyby z rodzaju Fusarium ............................................................. 210
Wiktor Pszczółkowski, Zdzisława Romanowska-Duda, Agata Pszczółkowska
Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp. hodowanych
na odciekach po fermentacji metanowej z biogazowni rolniczej ......................................... 220
Agata Pszczółkowska, Zdzisława Romanowska-Duda, Wiktor Pszczółkowski
Potencjał wykorzystania odpadu pofermentacyjnego z biogazowni rolniczej jako substratu
w produkcji podłoża hodowlanego do uprawy cyanobacteria .............................................. 233
Indeks Autorów .......................................................................................................................... 243

7
Matylda Barbara Mielcarska1, Magdalena Bossowska-Nowicka
2
Status badań i rozwoju szczepionek przeciw zakażeniom herpeswirusami typu 1 i 2 (HSV-1, HSV-2)
1. Wstęp
Wirusy opryszczki pospolitej (herpes simplex typu 1 i 2, HSV-1 i HSV-2) należą do podrodziny ludzkich alfaherpeswirusów, szeroko rozpowszechnionych na świecie. Wirusy te po wniknięciu do organizmu przemieszczają się do zakończeń neuronów czuciowych, gdzie przechodzą w stan latencji (utajenia). W sprzyjających warunkach, np. podczas niedoborów odporności, długotrwałego stresu, gorączki, zmian hormonal-nych, po urazie, ulegają one reaktywacji [1]. Zarówno w stanie utajenia, jak i aktywnej opryszczki możliwe jest ich przenoszenie na organizm wrażliwy. Powyższe cechy zapewniają wirusom skuteczne rozprzestrzenianie się w populacji ludzkiej, a także, między innymi, są przyczyną ciągłych zmagań nad konstrukcją skutecznej szczepionki przeciw obu typom wirusa. Już u 40% nastolatków i 90% osób powyżej 70 roku życia można wykryć HSV-1 [2]. Genomy obydwu typów wirusa są homologiczne w 40%, ale już w 83% w regionach kodujących białka [3]. W materiale genetycznym HSV-1 i HSV-2, który stanowi DNA (kwas deoksyrybonukleinowy), zakodowane jest między innymi 10 glikoprotein wirusowych, z których większość umiejscowiona jest w osłon-ce wirusowej: glikoproteiny B, C, D, E, G, H, I, L, M, N [4]. Skład aminokwasowy i budowa tych białek mogą być odmienne u różnych typów herpeswirusów. Należy podkreślić, że właśnie przeciw powyższym glikoproteinom powierzchniowym kierowana jest większość przeciwciał podczas odpowiedzi immunologicznej w czasie zakażenia HSV [3]. HSV-1 i HSV-2 są w 80% homologiczne pod względem polimor-fizmów długości fragmentów restrykcyjnych (restriction fragment length polymor-phism, RFLP) [5], przy czym szczepy HSV-1 są bardziej zróżnicowane niż szczepy HSV-2 [6].
HSV-1 i HSV-2 mogą powodować u człowieka nawracające owrzodzenia i zapa-lenie skóry wokół ust, zapalenie błon śluzowych jamy ustnej i dziąseł, zapalenie gardła lub owrzodzenia i zapalenie skóry i błon śluzowych narządów płciowych. W rzadkich przypadkach mogą stanowić przyczynę zapalenia rogówki lub zapalenia mózgu i opon mózgowych (herpes simplex encephalitis, HSE), które mogą być niezwykle groźne w skutkach, w tym również prowadzić do utraty wzroku, a nawet śmierci. Na dwa ostatnie schorzenia najbardziej narażone są noworodki oraz osoby w podeszłym wieku [7]. Bardzo niebezpieczna jest herpeswirusowa opryszczka noworodków, którą cechuje wysoka zaraźliwość i śmiertelność [8]. Zakażenie HSV-2 jest również istotnym czynnikiem wpływającym na ułatwiającym zakażenie wirusem HIV, uważa się, że nawet 50% zakażeń HIV jest rezultatem transmisji wirusa przez zmiany opryszczkowe spowodowane przez HSV-2 [9, 10]. Stosowanie m. in. acycloviru, valacycloviru czy
1 [email protected], Zakład Immunologii, Katedra Nauk Przedklinicznych, Wydział Medycyny
Weterynaryjnej, Szkoła Główna Gospodarstwa Wiejskiego, www.sggw.pl 2 [email protected], Zakład Immunologii, Katedra Nauk Przedklinicznych, Wydział Medycyny
Weterynaryjnej, Szkoła Główna Gospodarstwa Wiejskiego, www.sggw.pl

Matylda Barbara Mielcarska, Magdalena Bossowska-Nowicka
8
famcycloviru jest tylko częściowo skuteczne w zwalczaniu objawów chorób lub zapo-bieganiu ich nawrotom, gdyż efektywność leków przeciwwirusowych zależy od wielu czynników. Najważniejszymi z nich są: moment zastosowania leku oraz oporność na farmaceutyk, obserwowana u niektórych szczepów wirusów [11]. Zmienność gene-tyczna wśród szczepów HSV-1 i HSV-2 w różnych obszarach geograficznych, złożony cykl replikacyjny, a także zdolność wirusów do dożywotniego zakażenia gospodarza i reaktywacji wciąż stanowią wyzwanie dla naukowców projektujących szczepionki przeciw tym wirusom. W niniejszej pracy opisano etapy opracowania oraz rodzaje szczepionek skonstruowanych w celu zapobiegania zakażeniom i chorobom spowo-dowanym przez HSV-1 i HSV-2.
2. Badania nad szczepionkami przeciw HSV-1 i HSV-2 na modelach zwierzęcych
Już w pierwszej połowie XX wieku naukowcy izolowali herpeswirusy ze spowodo-wanych opryszczką zmian występujących na ciele pacjentów. Zakażali nimi zwierzęta, a następnie ludzi, z nadzieją, że uda im się zapobiec uciążliwym nawracającym objawom choroby [11]. Choć próby te nie zostały uwieńczone sukcesem, dzięki inten-sywnym badaniom skonstruowano szczepionki umożliwiające doprowadzenie do eradykacji zwierzęcych herpeswirusów – bydlęcego herpeswirusa typu 1 i świńskiego herpeswirusa typu 1 [12, 13].
Obecnie intensywne badania prowadzone są zarówno nad szczepionkami prewen-cyjnymi, które mogłyby zapobiegać zakażeniu, jak również terapeutycznymi, które mogłyby łagodzić objawy zakażeń HSV-1 i HSV-2. Podobną strategię przyjmuje się podczas projektowania szczepionek przeciw innym herpeswirusom – szczepionka przeciw wirusowi ospy wietrznej i półpaśca (varicella zoster virus, VZV) o właściwoś-ciach zarówno prewencyjnych, jak i terapeutycznych, znajduje się w III fazie badań klinicznych [14]. VZV jest blisko spokrewniony z HSV-1 i HSV-2, jednak jego szcze-py wykazują dużo mniejszą zmienność genetyczną [15].
Uważa się, że skuteczność szczepionek prewencyjnych powinna być badana wśród młodych dorosłych osób, niezakażonych HSV-1 i/lub HSV-2. Jeśli wybrana szcze-pionka okaże się skuteczna w ochronie przeciw zakażeniu, powinno się kolejno zbadać skuteczność szczepionki u dzieci i niemowląt. Istnieje możliwość, że taka szczepionka mogłaby zapewnić ochronę przed HSE, jak również herpeswirusowym zapaleniem rogówki.
W celu testowania i monitorowania działania szczepionek HSV w badaniach przed-klinicznych wykorzystuje się modele zwierzęce. Należy przy tym brać pod uwagę, że HSV jest patogenem ludzkim, zaś interakcje wirus-gospodarz mogą być odmienne wśród różnych osób. Może to wskazywać na ograniczenia w przewidywaniu, czy skuteczność szczepionek zaobserwowana w modelach zwierzęcych będzie analogiczna u człowieka. Szczególnie popularny jest model mysi, który był powszechnie stoso-wany jako model zakażenia HSV-1 in vivo i odegrał znaczącą rolę w opracowaniu terapii acyclovirem w latach 80-tych XX wieku [16]. HSV-1 i HSV-2 są zdolne do zakażania mysich komórek in vitro, zaś do organizmu gospodarza wnikają przez nabłonek węchowy [17], jednak u myszy opryszczka narządów płciowych występuje tylko podczas pierwotnego zakażenia HSV-2 i nie obserwuje się tej postaci po reakty-wacji wirusa. Podczas zakażenia pierwotnego występuje również wysoka śmiertelność

Status badań i rozwoju szczepionek przeciw zakażeniom herpeswirusami typu 1 i 2 (HSV-1, HSV-2)
9
zwierząt [18]. Innymi gatunkami, u których badana jest patogenność oraz procesy immunologiczne związane z ustaleniem latencji i reaktywacją HSV-1 i HSV-2 są świn-ki morskie, szczury, bawełniaki, króliki, ponocnice i makaki rezusy [19].
Intensywne badania nad szczepionkami przeciw HSV-1 i HSV-2, zapoczątkowane przez instytucje rządowe, akademickie, jak również firmy biotechnologiczne, wciąż trwają i obserwuje się ich znaczący rozwój. Z powodu możliwości reaktywacji herpes-wirusów bardzo trudno jest ustalić punkt końcowy kosztownych badań klinicznych, jak również odpowiednie placebo. Mimo tych przeciwności, wiele szczepionek znajduje się w fazie badań przedklinicznych, a kilka przechodzi obecnie fazę badań klinicznych. Firmy farmaceutyczne również angażują się w testowanie powyższych szczepionek [11, 20].
3. Rodzaje szczepionek przeciw HSV-1 i HSV-2
Obecnie główny nacisk kładziony jest na wynalezienie szczepionki przeciw HSV-2, w celu zwalczenia choroby narządów płciowych. Szczepionka taka mogłaby w przysz-łości przynieść również korzyści terapeutyczne w zakażeniu HSV-1, z uwagi na homo-logię pomiędzy powyższymi typami herpeswirusa [20]. W niniejszym rozdziale przedstawiono najważniejsze rodzaje szczepionek przeciw HSV-1 i HSV-2, które znaj-dują się w fazie badań przedklinicznych lub klinicznych.
3.1. Szczepionki zawierające glikoproteiny wirusowe
Najliczniejszą grupą badanych szczepionek przeciw herpeswirusom są szczepionki zawierające glikoproteiny wirusowe. Jedną ze szczepionek, będącą w fazie badań klini-cznych, jest Herpevac, zawierająca glikoproteinę D2 (gD2) oraz glin i MPL® (mono-phosphoryl lipid A, agonista TLR4 – Toll-like receptor 4) jako adiuwanty [21]. Na podstawie tych badań wykazano, że chroniła ona 58% seronegatywnych kobiet przed zachorowaniem na opryszczkę narządów płciowych spowodowaną HSV-1, a zarazem 32% kobiet przed zakażeniem tym wirusem, przy czym nie miała wpływu na zaka-żenie czy chorobę spowodowaną zakażeniem HSV-2. Jednocześnie podczas powyż-szych badań klinicznych po raz pierwszy zaobserwowano u pacjentów, że miana prze-ciwciał przeciw gD2 korelowały ze stopniem ochrony przed zakażeniem lub chorobą spowodowaną zakażeniem HSV-1. Miana przeciwciał stanowią zatem ważny punkt odniesienia podczas badania poszczepiennej odporności ochronnej.
Kolejna szczepionka przeciw HSV-2 o charakterze terapeutycznym, zawierająca gD2, białko wirusowe ICP4 (infected cell polypeptide 4) i nowatorski adiuwant Matrix M2 znajduje się w I i II fazie badań klinicznych. Wykazuje ona skuteczne działanie przeciwwirusowe, gdyż u pacjentów szczepionych zaobserwowano znaczne obniżenie ilości miana wirusa w obrębie zmian chorobowych, w porównaniu do pacjentów nie-szczepionych. Ilość zmian chorobowych również była zmniejszona u ludzi szcze-pionych [22].
Wśród innych podobnych szczepionek, będących w fazie badań przedklinicznych bądź klinicznych wyróżnić można szczepionki zawierające glikoproteiny D, C2 i E2 (gD, gC2, gE2) [23].
3.2. Szczepionki z wirusem atenuowanym
W fazie badań przedklinicznych znajdują się także szczepionki z wirusem atenu-owanym lub niezdolnym do replikacji. Szczepionki takie zawierają pełen zakres anty-

Matylda Barbara Mielcarska, Magdalena Bossowska-Nowicka
10
genów wirusowych, dzięki czemu mogą pobudzać odpowiedź immunologiczną zwią-zaną zarówno z odpornością wrodzoną, jak i nabytą. Przykładem jest szczepionka HSV529 zawierająca niezdolny do replikacji szczep HSV-2, która wykazuje właści-wości zarówno prewencyjne, jak i terapeutyczne u myszy i świnek morskich [24].
Inna szczepionka z atenuowanym i pozbawionym gD2 wirusem HSV-2 zapobie-gała u myszy wirusowemu zapaleniu mózgu oraz opryszczce skóry. Wspomniana szczepionka była pierwszą, która przyniosła korzyść w postaci eliminacji latentnego wirusa w grzbietowych korzeniach zwojów nerwowych zwierząt [25].
Obiecująca wydaje się również szczepionka zawierająca atenuowany szczep HSV-1 – HF10, która w badaniach przedklinicznych zapewniła u myszy ochronę nie tylko przed zakażeniem HSV-1, lecz również przed opryszczką narządów płciowych spowodowaną przez HSV-2 [26].
3.3. Szczepionki typu „prime-pull”
Inną nowatorską szczepionką jest szczepionka złożona, typu „prime-pull” [27]. Pobudza ona tworzenie („prime”), a następnie napływ limfocytów T pamięci („pull”) do miejsca potencjalnego zakażenia, którym w przypadku HSV-2 są narządy płciowe. Limfocyty T CD8
+ pamięci produkowane są w organizmie po podaniu konwencjo-
nalnej szczepionki w postaci atenuowanego HSV-2 („prime”). Kolejno, dzięki miej-scowemu zastosowaniu chemokin CXCL9/CXCL10 („pull”), limfocyty te napływają do okolic narządów płciowych. Myszy, u których stosowano powyższą szczepionkę, nie wykazywały klinicznych objawów zakażenia HSV-2, a rozprzestrzenianie się wirusa drogą neuronów czuciowych było u zwierząt znacznie ograniczone lub nawet zahamowane.
3.4. Inne szczepionki
Szczepionka zawierająca wektor lentiwirusowy – FIV (feline immunodeficiency virus), eksprymujący glikoproteinę B1 (gB1), również przyniosła obiecujące rezultaty w trakcie badań przedklinicznych na myszach [28]. Inna szczepionka, podawana donosowo w postaci niejonowych pęcherzyków surfaktantowych (non-ionic surfactant vesicles, NISV) zawierających rekombinowaną gB HSV-1 lub peptydy wirusa bogate w lizyny, indukowała u myszy odpowiedź immunologiczną przeciw HSV-1, jak rów-nież chroniła zwierzęta przed zakażeniem HSV-2 [29].
Obiecująca wydaje się być również plazmidowa szczepionka DNA (pDNA), kodu-jąca gD2 oraz białka tegumentu HSV-2 – VP11/12 i VP13/14, która wykazywała działanie profilaktyczne, jak również terapeutyczne podczas badań u świnek morskich [30]. Po podaniu szczepionki osobnikom uprzednio zakażonym wirusem obserwowano także znaczne zmniejszenie ilości DNA latentnego HSV-2 w grzbietowych korzeniach zwojów nerwowych zwierząt.
Inna szczepionka DNA, zawierająca gD HSV-2, jak również specjalnie zmodyf-ikowane polinukleotydy oraz sekwencje kodujące ubikwitynę, po podaniu kierowana była w komórkach mysich do przetwarzania w proteasomach, co pobudzało odpowiedź immunologiczną związaną z aktywnością limfocytów B oraz T CD8
+ i obniżało ilość
latentnego wirusa w zwojach nerwowych zwierząt [31]. Zaprojektowano również szczepionkę terapeutyczną składającą się z 32 syntetycz-
nych peptydów HSV-2, pochodzących z białek wszystkich faz replikacji wirusa,

Status badań i rozwoju szczepionek przeciw zakażeniom herpeswirusami typu 1 i 2 (HSV-1, HSV-2)
11
w połączeniu z różnorodnymi adiuwantami, między innymi ludzkim białkiem szoku cieplnego Hsc70 [32]. W badaniach klinicznych szczepionka ta indukowała silną odpowiedź immunologiczną związaną z limfocytami T CD4
+ i CD8
+ u ludzi zaka-
żonych HSV-2.
4. Podsumowanie
W obecnych czasach w różnych fazach badań znajduje się szereg szczepionek przeciw HSV, przy czym wyzwaniem jest zapewnienie takich składowych szczepion-ki, które zapewnią skuteczną ochronę zarówno przeciw HSV-1, jak i HSV-2. Uważa się, że aby kolejne generacje szczepionek były skuteczne przeciw obu typom wirusa, powinny one pobudzać produkcję przeciwciał neutralizujących. W celu uzyskania pełniejszej ochrony przeciwwirusowej, szczepionki powinny także indukować inne rodzaje odpowiedzi immunologicznej. Nowe adiuwanty, dodawane do wielu szczepio-nek, mogą stymulować różne komórki układu odpornościowego, np. limfocyty T pamięci, których napływ w pobliże miejsca zakażenia ma w ochronie przeciw-wirusowej duże znaczenie [27, 33]. Podczas badań klinicznych dowiedziono, że nowatorskie adiuwanty szczepionkowe, stosowane w szczepionkach terapeutycznych, mogą mieć znaczący wpływ na zmniejszenie miana wirusa w obrębie zmian choro-bowych, co świadczy o zahamowaniu jego replikacji. Nowe metody podawania szcze-pionek przeciw HSV-1 i HSV-2, jak również ekspresji białek wirusowych w wek-torach szczepionkowych są przedmiotem intensywnych badań.
Literatura
1. Grinde B., Herpesviruses: latency and reactivation – viral strategies and host response, Journalof Oral Microbiology, 5 (2013), doi: 10.3402/jom.v5i0.22766.
2. Smith J. S., Robinson N. J. Age-specific prevalence of infection with herpes simplex virus types 2 and 1: a global review, Journal of Infectious Diseases, 186 (2002), s. 3-28.
3. LeGoff J., Péré H., Bélec L. Diagnosis of genital herpes simplex virus infection in the clinical laboratory, Journal of Virology, 11 (2014), doi: 10.1186/1743-422X-11-83.
4. Whitley R. J., Roizman B. Herpes simplex virus infections, Lancet, 357 (2001), s. 1513-8. 5. Szpara M. L., Gatherer D., Ochoa A., Greenbaum B., Dolan A., Bowden R. J., Enquist L.
W., Legendre M., Davison A. J. Evolution and diversity in human herpes simplex virus genomes, Journal of Virology, 88 (2014), s. 1209-27.
6. Norberg P., Kasubi M. J., Haar L., Bergström T., Liljeqvist J. A. Divergence and recombination of clinical herpes simplex virus type 2 isolates, Journal of Virology, 81 (2007), s. 13158-67.
7. Whitley R., Kimberlin D. W., Prober C. G. Human herpesviruses: biology, therapy and immunoprophylaxis; Chapter 32 Pathogenesis and disease, Cambridge University Press (2007), ISBN-13: 978-0-521-82714-1.
8. Cherpes T. L., Matthews D. B., Maryak S. A. Neonatal herpes simplex virus infection, Clinical Obstetrics and Gynecology, 55 (2012), s. 938-44.
9. Freeman E. E., Orroth K. K., White R. G., Glynn J. R., Bakker R., Boily M. C., Habbema D., Buvé A., Hayes R. Proportion of new HIV infections attributable to herpes simplex 2 increases over time: simulations of the new changing role of sexually transmitted infections in Sub-Saharian African HIV epidemics, Sexually Transmitted Infections, 83 (2007), i17-24.
10. Masese L., Baeten J. M., Richardson B. A., Bukusi E., John-Stewart G., Graham S. M., Shafi J., Kiarie J., Overbaugh J., McClelland R. S. Changes in the contribution of genital tract infections to HIV acquisition among Kenyan high-risk women from 1993 to 2012, AIDS, 29 (2015), s. 1077-85.

Matylda Barbara Mielcarska, Magdalena Bossowska-Nowicka
12
11. Kemble G., Spaete R. Human Herpesviruses. Biology, Therapy, and Immunoprophylaxis; Chapter 69 Herpes simpex vaccines, Cambridge University Press (2007), ISBN-13: 978-0-521-82714-0.
12. Raaperi K., Orro T., Viltrop A. Epidemiology and control of bovine herpesvirus 1 infection in Europe, Veterinary Journal 201 (2014), s. 249-56.
13. Pensaert M., Labarque G., Favoreel H., Nauwynck H. Aujeszky’s disease vaccination and differentiation of vaccinated from infected pigs, Developments in Biologicals, 119 (2004), s. 243-54.
14. Lal H., Cunningham A. L., Godeaux O., Chlibek R., Diez-Domingo J., Hwang S. J., Levin M. J., McElhaney J. E., Poder A., Puig-Barberà J., Vesikari T., Watanabe D., Weckx L., Zahaf T., Heineman T. C. Efficacy of an adjuvanted herpes zoster subunit vaccine in older adults, New England Journal of Medicine, 372 (2015), s. 2087-96.
15. Chow V. T., Tipples G. A., Grose C. Bioinformatics of varicella-zoster virus: single nucleotide polymorphisms define clades and attenuated vaccine genotypes, Infection, Genetics and Evolution, 18 (2013), s. 351-6.
16. Field H. J. Development of clinical resistance to acyclovir in herpes simplex virus-infected mice receiving oral therapy, Antimicrobial Agents and Chemotherapy, 21 (1982), s. 744-52.
17. Shivkumar M., Milho R., May J. S., Nicoll M. P., Efstathiou S., Stevenson P. G. Herpes simplex virus 1 targets the murine olfactory neuroepithelium for host entry, Journal of Virology, 87 (2013), s. 10477-88.
18. Dasgupta G., BenMohamed L. Of mice and not humans: how reliable are animal models for evaluation of herpes CD8(+)-T cell-epitopes-based immunotherapeutic vaccine candidates?, Vaccine, 29 (2011), s. 5824-36.
19. Kollias C. M., Huneke R. B., Wigdahl B., Jennings S. R. Animal models of herpes simplex virus immunity and pathogenesis, Journal of Neurovirology, 21 (2015), s. 8-23.
20. Johnston C., Gottlieb S. L., Wald A. Status of vaccine research and development of vaccines for herpes simplex virus, Vaccine, 34 (2016), s. 2948-52.
21. Belshe R. B., Leone P. A., Bernstein D. I., Wald A., Levin M. J., Stapleton J. T., Gorfinkel I., Morrow R. L., Ewell M. G., Stokes-Riner A., Dubin G., Heineman T. C., Schulte J. M., Deal C. D. Efficacy results of a trial of a herpes simplex vaccine, New England Journal of Medicine, 366 (2012), s. 34-43.
22. Wald A., Bernstein D., Fife K., Lee P., Tyring S., Van Wagoner N. Novel therapeutic vaccine for genital herpes reduces genital HSV-2 shedding, 53
rd Interscience Conference on
Antimicrobial Agents and Chemotherapy, September 10-13, Denver, CO, USA, (2013). 23. Awasthi S., Huang J., Shaw C., Friedman H. M. Blocking herpes simplex virus 2
glycoprotein E immune evasion as an approach to enhance efficacy of a trivalent subunit antigen vaccine for genital herpes, Journal of Virology, 88 (2014), s. 8421-32.
24. Bernard M. C., Barban V., Pradezynski F., de Montfort A., Ryall R., Caillet C., Londono-Hayes P. Immunogenicity, protective efficacy, and non-replicative status of the HSV-2 vaccine candidate HSV529 in mice and guinea pigs, PLoS One, 10 (2015), doi: 10.1371/journal.pone.0121518.
25. Petro C., González P. A., Cheshenko N., Jandl T., Khajoueinejad N., Bénard A., Sengupta M., Herold B. C., Jacobs Jr W. R. Herpes simplex type 2 virus deleted in glycoprotein D protects against vaginal, skin and neural disease, eLife, (2015), doi: 10.7554/eLife.06054.
26. Luo C., Goshima F., Kamakura M., Mutoh Y., Iwata S., Kimura H., Nishiyama Y. Immunization with a highly attenuated replication-competent herpes simplex virus type 1 mutant, HF10, protects mice from genital disease caused by herpes simplex virus type 2, Frontiers in Microbiology, 3 (2012), doi: 10.3389/fmicb.2012.00158.
27. Shin H., Iwasaki A. A vaccine strategy protects against genital herpes by establishing local memory T cells, Nature, 491 (2012), s. 463-7.
28. Chiuppesi F., Vannucci L., De Luca A., Lai M., Matteoli B., Freer G., Manservigi R., Ceccherini-Nelli L., Maggi F., Bendinelli M., Pistello M. A lentiviral vector-based, herpes

Status badań i rozwoju szczepionek przeciw zakażeniom herpeswirusami typu 1 i 2 (HSV-1, HSV-2)
13
simplex virus 1 (HSV-1) glycoprotein B vaccine affords cross-protection against HSV-1 and HSV-2 genital infections, Journal of Virology, 86 (2012), s. 6563-74.
29. Cortesi R., Ravani L., Rinaldi F., Marconi P., Drechsler M., Manservigi M., Argnani R., Menegatti E., Esposito E., Manservigi R. Intranasal immunization in mice with non-ionic surfactants vesicles containing HSV immunogens: a preliminary study as possible vaccine against genital herpes, International Journal of Pharmaceutics, 440 (2013), s. 229-37.
30. Veselenak R. L., Shlapobersky M., Pyles R. B., Wei Q., Sullivan S. M., Bourne N. A Vaxfectin®-adjuvanted HSV-2 plasmid DNA vaccine is effective for prophylactic and therapeutic use in the guinea pig model of genital herpes, Vaccine, 30 (2012), doi: 10.1016/j.vaccine.2012.09.057.
31. Dutton J. L., Li B., Woo W., Marshak J. O., Xu Y., Huang M., Dong L., Frazer I. H., Koelle D. M. A novel DNA vaccine technology conveying protection against a lethal herpes simplex viral challenge in mice, PLoS One, 8 (2013), doi: 10.1371/journal.pone.0076407.
32. Wald A., Koelle D. M., Fife K., Warren T., Leclair K., Chicz R. M., Monks S., Levey D. L., Musselli C., Srivastava P. K. Safety and immunogenicity of long HSV-2 peptides complexed with rhHsc70 in HSV-2 seropositive persons, Vaccine, 29 (2011), s. 8520-9.
33. 1. Agenus Vaccine Shows Significant Reduction in Viral Burden after HerpV Generated Immune Activation, (2013), http://www.agenusbio.com/docs/press-releases/2013/herpv-vaccine-for-genital-herpes-meets-primary-endpoint.php.
Status badań i rozwoju szczepionek przeciw zakażeniom herpeswirusami typu 1 i 2 (HSV-1, HSV2)
Streszczenie Wirusy herpes simplex typu 1 i 2 (HSV-1, HSV-2) są szeroko rozpowszechnione wśród ludzi. Zakażony człowiek pozostaje dożywotnim nosicielem wirusa, gdyż HSV-1 i HSV-2 przechodzą w stan latencji w układzie nerwowym, a w późniejszym czasie mogą ulegać reaktywacji. Większość dorosłych na całym świecie nie wykazuje objawów zakażenia lub ograniczają się one do łagodnej opryszczki wargowej. W szczególnych przypadkach HSV-1, jak również HSV-2 mogą powodować groźne choroby, jak herpes-wirusowe zapalenie mózgu (Herpes simpex encephalitis, HSE), zapalenie rogówki, zakażenia noworod-ków. Leczenie pacjentów jest tylko częściowo skuteczne, wielu z nich umiera lub zmaga się z trwałymi na-stępstwami chorób. Do dnia dzisiejszego nie wynaleziono skutecznych szczepionek przeciwko zakażeniom HSV-1 i HSV-2 u ludzi, jednak mogłyby one okazać się dobrym rozwiązaniem w celu zapobieżenia zakażeniom lub efektywnego zmniejszenia ich objawów. Trudności w wynalezieniu wydajnych szczep-ionek związane są z cyklem replikacyjnym wirusów i niepełnym zrozumieniem mechanizmów immunolo-gicznych koniecznych do zwalczania pierwotnego lub nawracającego zakażenia. W niniejszej pracy doko-nano przeglądu szczepionek prewencyjnych oraz terapeutycznych, znajdujących się obecnie w różnych fazach badań. Słowa kluczowe: HSV-1, HSV-2, szczepionki
Status of research and development of vaccines against herpesvirus types 1 and 2 (HSV-1, HSV2)
Abstract Herpes simplex type 1 and 2 viruses (HSV-1, HSV-2) are widespread among humans. The infected person remains a lifelong viral carrier due to HSV-1 or HSV-2 latention in the nervous system. In the later time both types of viruses may undergo reactivation. Most adults around the world have no signs of infection or infection is limited to mild herpes labialis. However, in specific cases, HSV-1 as well as HSV-2 can cause serious illnesses like herpesvirus encephalitis (HSE), keratitis, neonatal infections. Treatment of patients is only partially effective, many of them die or suffer from permanent consequences of the disease. To this day, no effective HSV-1 and HSV-2 vaccines have been developed for humans, although they could constitute good solution to prevent infection or to effectively reduce their symptoms. Difficulties in developing effective vaccines relate to the replication cycle of viruses and incomplete understanding of the immune mechanisms necessary to fight primary or recurrent infections. Preventive and therapeutic vaccines which are at various stages of research were reviewed in the present study. Keywords: HSV-1, HSV-2, vaccines

14
Marta Kozieł1, Karolina Włodarczyk
2, Dominika Wojton
3
Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
1. Wstęp
Ludzki organizm jest zasiedlony przez ogromną liczbę mikroorganizmów, z czego zdecydowaną większość stanowią bakterie. Zespół bakterii, grzybów, wirusów, archeonów, pierwotniaków bytujących w naszym organizmie określamy terminem mikrobiota, a geny przez nie kodowane mikrobiomem. Organizmy te to głównie ko-mensale bądź saprofity. Mikrobiom wykazuje istotny wpływ na prawidłowe funkcjo-nowanie organizmu ludzkiego. Przypisuje mu się przede wszystkim oddziaływanie na układ immunologiczny. Obok jego funkcji ochronnej wymienia się funkcję metabo-liczną i troficzną. Należy podkreślić, że funkcje te zazębiają się ze sobą i nie można ich traktować odrębnie. Z racji istotnego wpływu na ludzki organizm mikrobiom stał się obiektem zainteresowań wielu badaczy. Na podstawie analizy sekwencji nukle-otydowej bakteryjnego genu 16S rRNA wykazano, że w skład mikrobiomu wchodzą głównie cztery typy bakterii tj. Bacteroidetes, Firmicutes, Proteobacteria, Actino-bacteria [3, 6]. Mikrobiom jelitowy noworodka kształtuje się już w chwili narodzin. Wpływ na ten proces ma szereg czynników tj. rodzaj porodu, sposób karmienia, wiek ciążowy, antybiotykoterapia. Ponadto na wykształcenie się prawidłowej flory bakteryjnej wpływ ma stan zdrowotny matki oraz jej własna mikroflora. Uważa się, że okres 3 pierwszych lat jest najbardziej istotny dla prawidłowej kolonizacji organizmu przez bakterie [20-21]. Wszelkie zaburzenia tego procesu prowadzą do tzw. dysbioz jelitowych, które mają udział w patogenezie bądź progresji licznych chorób, takich jak: martwicze zapalenie jelit, otyłość, cukrzyca, alergie, astma, nieswoiste zapalenie jelit i wiele innych [26, 29, 32, 34].
2. Cel pracy
Celem niniejszej pracy jest charakterystyka składu mikrobioty jelitowej, omówienie czynników determinujących wykształcenie prawidłowej flory jelitowej noworodków oraz jej roli w patogenezie i/lub progresji chorób w wieku późniejszym.
3. Mikrobiota jelitowa
3.1. Charakterystyka struktury i funkcji mikrobioty jelitowej
Ludzką mikrobiotę stanowi zespół różnorodnych mikroorganizmów, głównie bakterii oraz występujących w mniejszych ilościach grzybów, wirusów, pierwotnia-
1 [email protected], Studenckie Koło Naukowe Mikrobiologów „Bakcyl”, Zakład Genetyki
i Mikrobiologii, Wydział Biologii i Biotechnologii, Uniwersytet Marii Curie - Skłodowskiej 2 [email protected], Studenckie Koło Naukowe Mikrobiologów „Bakcyl”, Zakład Genetyki
i Mikrobiologii, Wydział Biologii i Biotechnologii, Uniwersytet Marii Curie - Skłodowskiej 3 [email protected], Studenckie Koło Naukowe Mikrobiologów „Bakcyl”, Zakład Wirusologii
i Immunologii, Wydział Biologii i Biotechnologii, Uniwersytet Marii Curie - Skłodowskiej
Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
15
ków, archeonów, kolonizujących błony śluzowe i skórę organizmu ludzkiego [1]. Dawniej przyjęto, że wyżej wymienione drobnoustroje określa się terminem mikro-biota, z kolei geny przez nie kodowane są określane jako mikrobiom. Aktualnie, coraz częściej, terminy te stosuje się zamiennie [2].
Mikroorganizmy należące do mikrobioty człowieka występują na skórze oraz błonach śluzowych układu: moczowo-płciowego, oddechowego, pokarmowego. Szacuje się, że mikrobiom jelitowy zawiera około 10
14 bakterii. Najliczniejsza jest
mikroflora jelita grubego, zawierająca średnio 1011
-1012
komórek bakteryjnych na 1 g kału [3, 4].
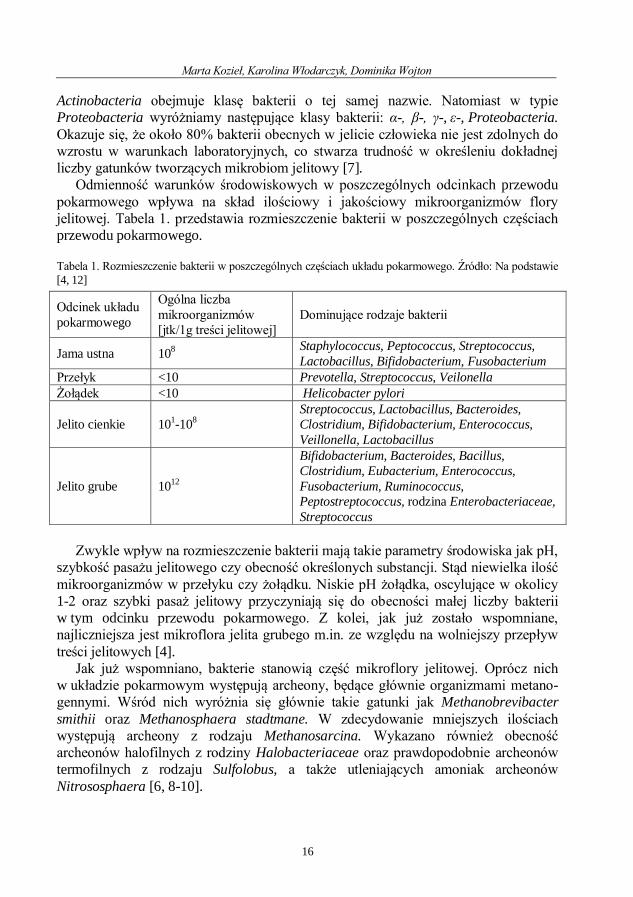
Ze względu na swoją unikatowość, różnorodność oraz wpływ na funkcjonowanie organizmu mikrobiom jelitowy stał się obiektem badań i zainteresowań licznych naukowców. W wyniku czego w 2007 roku powstał Human Microbiome Project, czyli ogólnoświatowy projekt mający na celu scharakteryzowanie genomów mikroorga-nizmów wchodzących w skład mikrobiomu, zwłaszcza jelitowego. Dzięki temu przedsięwzięciu, jak i wielu innym badaniom udało się ustalić typy bakterii tworzące mikrobiotę jelitową. Analiza sekwencji nukleotydowej bakteryjnego genu 16S rRNA wykazała, że 98% bakterii obecnych w przewodzie pokarmowym należy do jednego z czterech następujących typów, tj.: Firmicutes, Bacteroidetes, Proteobacteria, Actino-bacteria, przy czym większość gatunków należy do dwóch pierwszych typów, co dokładnie obrazuje wykres 1. Inne bakterie występujące w jelicie człowieka należą do takich nielicznych typów jak: Verrucomicrobia, Cyanobacteria. Stanowią one około 2% wszystkich mikroorganizmów obecnych w mikrobiocie jelitowej. Przedstawiony powyżej ogólny skład mikrobiomu jelitowego jest charakterystyczny dla flory jelitowej dorosłego człowieka [5, 6].
Wykres 1. Udział poszczególnych typów bakterii w mikrobiocie jelitowej człowieka., [opracowanie własne
na podstawie [6] [5]]. Gatunki bakterii występujące w fizjologicznej mikroflorze jelitowej należą w
większości (98%) do 4 typów tj. Firmicutes, Bacteroidetes, Proteobacteria, Actinobacteria. Pozostałe 2% bakterii należy do innych grup taksonomicznych, których udział w mikrobiomie jest niewielki.
Typ Firmicutes obejmuje głównie bakterie gram dodatnie, wśród których ważną
grupę stanowią klasy: Bacilli, Clostridia, Mollicutes. Do typu Bacteroidetes zaliczamy
następujące klasy bakterii: Bacteroides, Flavobacteria, Sphingobacteria. Gromada
Marta Kozieł, Karolina Włodarczyk, Dominika Wojton
16
Actinobacteria obejmuje klasę bakterii o tej samej nazwie. Natomiast w typie
Proteobacteria wyróżniamy następujące klasy bakterii: α-, β-, γ-, ε-, Proteobacteria.
Okazuje się, że około 80% bakterii obecnych w jelicie człowieka nie jest zdolnych do
wzrostu w warunkach laboratoryjnych, co stwarza trudność w określeniu dokładnej
liczby gatunków tworzących mikrobiom jelitowy [7].
Odmienność warunków środowiskowych w poszczególnych odcinkach przewodu
pokarmowego wpływa na skład ilościowy i jakościowy mikroorganizmów flory
jelitowej. Tabela 1. przedstawia rozmieszczenie bakterii w poszczególnych częściach
przewodu pokarmowego.
Tabela 1. Rozmieszczenie bakterii w poszczególnych częściach układu pokarmowego. Źródło: Na podstawie
[4, 12]
Odcinek układu
pokarmowego
Ogólna liczba
mikroorganizmów
[jtk/1g treści jelitowej]
Dominujące rodzaje bakterii
Jama ustna 108 Staphylococcus, Peptococcus, Streptococcus,
Lactobacillus, Bifidobacterium, Fusobacterium
Przełyk <10 Prevotella, Streptococcus, Veilonella
Żołądek <10 Helicobacter pylori
Jelito cienkie 101-10
8 Streptococcus, Lactobacillus, Bacteroides,
Clostridium, Bifidobacterium, Enterococcus,
Veillonella, Lactobacillus
Jelito grube 1012
Bifidobacterium, Bacteroides, Bacillus,
Clostridium, Eubacterium, Enterococcus,
Fusobacterium, Ruminococcus,
Peptostreptococcus, rodzina Enterobacteriaceae,
Streptococcus
Zwykle wpływ na rozmieszczenie bakterii mają takie parametry środowiska jak pH,
szybkość pasażu jelitowego czy obecność określonych substancji. Stąd niewielka ilość
mikroorganizmów w przełyku czy żołądku. Niskie pH żołądka, oscylujące w okolicy
1-2 oraz szybki pasaż jelitowy przyczyniają się do obecności małej liczby bakterii
w tym odcinku przewodu pokarmowego. Z kolei, jak już zostało wspomniane,
najliczniejsza jest mikroflora jelita grubego m.in. ze względu na wolniejszy przepływ
treści jelitowych [4].
Jak już wspomniano, bakterie stanowią część mikroflory jelitowej. Oprócz nich
w układzie pokarmowym występują archeony, będące głównie organizmami metano-
gennymi. Wśród nich wyróżnia się głównie takie gatunki jak Methanobrevibacter
smithii oraz Methanosphaera stadtmane. W zdecydowanie mniejszych ilościach
występują archeony z rodzaju Methanosarcina. Wykazano również obecność
archeonów halofilnych z rodziny Halobacteriaceae oraz prawdopodobnie archeonów
termofilnych z rodzaju Sulfolobus, a także utleniających amoniak archeonów
Nitrososphaera [6, 8-10].

Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
17
Różnorodność wirusów występujących w mikrobiomie jelitowym jest niewielka. Najczęściej występują fagi charakteryzujące się dużą stabilnością genetyczną. Warto zauważyć, że wirusy obecne w mikrobiomie są charakterystyczne dla danej osoby [6].
Kształtowanie się fizjologicznej flory bakteryjnej jelita noworodków rozpoczyna się już w pierwszych godzinach życia i zależy od wielu czynników. Uważa się, że okres trzech pierwszych lat jest najbardziej istotny dla wykształcenia prawidłowego mikrobiomu. Zmiany mikroflory jelitowej w tym okresie, np. poprzez modyfikowanie składu diety, mają istotny wpływ na prawidłowy rozwój i choroby w wieku póź-niejszym. Mikrobiota jelitowa dziecka zaczyna przypominać, pod względem składu i różnorodności, mikroflorę bakteryjną przewodu pokarmowego dorosłego człowieka w wieku pomiędzy drugim, a piątym rokiem życia [11, 12]. Należy pamiętać o tym, że mikrobiom stanowi indywidualny zespół mikroorganizmów charakterystyczny dla danego człowieka. W ramach Human Microbiome Project wykazano, że zestaw mikroorganizmów każdego człowieka jest unikatowy, zwłaszcza pod względem pochodzenia i udziału w nim poszczególnych gatunków bakteryjnych. Jednocześnie udało się wykazać obecność wspólnych genów u wielu uczestników projektu. Dane te pozwalają nam wysnuć wniosek o istnieniu wspólnego genetycznego mikrobiomu [6].
Wymienia się następujące funkcje mikrobioty jelitowej:
immunologiczną,
troficzną,
metaboliczną. Funkcja metaboliczna jest głównie związana z fermentacją substancji nie rozkła-
danych przez organizm ludzki, np. roślinnych wielocukrów, ksenobiotyków oraz nie-których aminokwasów. Ponadto bakterie jelitowe biorą udział w biosyntezie witamin z grupy B oraz witaminy K. Bakterie obecne w przewodzie pokarmowym przyczyniają się również do zwiększonej absorpcji jonów żelaza, magnezu czy wapnia. Wpływają na metabolizm kwasów tłuszczowych i cholesterolu poprzez wytwarzanie enzymów z grupy hydrolaz. Produkty fermentacji polisacharydów roślinnych to m.in. krótkołań-cuchowe kwasy tłuszczowe (maślan, propionian, octan), które są źródłem substancji odżywczych dla kolonocytów, co pośrednio wiąże się z funkcją troficzną mikrobiomu. Otóż krótkołańcuchowe kwasy tłuszczowe pozytywnie oddziałują na ciągłość nabłoć-ka jelitowego, ponadto mają one działanie przeciwzapalne, co częściowo wiąże się z funkcją immunologiczną. Krótkołańcuchowe kwasy tłuszczowe, zwłaszcza maślan, stymulują proces dojrzewania i różnicowania kolonocytów. Innymi substancjami chemicznymi, do których wytwarzania przyczynia się mikroflora jelitowa, są mucyny, które chronią nabłonek przewodu pokarmowego przed patogennymi mikroorgani-zmami [4]. Mikroflora jelitowa oddziałuje również na rozwój i dojrzewanie układu im-munologicznego jelita (GALT). Dzięki stałej obecności komensalnych bakterii układ immunologiczny jest stale utrzymywany w stanie gotowości, aby zapobiec ewentual-nym infekcjom przez patogeny. Mikrobiom jelitowy pozytywnie oddziałuje na odpo-wiedź komórkową i humoralną [2]. Ponadto komensalne bakterie poprzez interakcje z receptorami na powierzchni nabłonka, konkurencję o składniki pokarmowej, np. kwas sialowy czy wydzielanie substancji przeciwdrobnoustrojowych takich jak: nadtlenek wodoru, kwasy organiczne, bakteriocyny przyczyniają się do eliminacji patogenów [4].

Marta Kozieł, Karolina Włodarczyk, Dominika Wojton
18
3.2. Czynniki determinujące mikroflorę jelitową noworodków
Na kształtowanie się prawidłowej mikrobioty noworodka ma wpływ szereg
istotnych czynników zarówno prenatalnych jak i postnatalnych [13].
Uważa się, że okres niemowlęcy jest najważniejszy dla wykształcenia prawidłowej
flory jelitowej. Należy mieć na uwadze fakt, że raz wykształcona flora bakteryjna nie
jest całkowicie stabilna, ale ulega ciągłym zmianom w trakcie życia pod wpływem
wielu czynników takich jak: dieta, wiek, genotyp, warunki socjalno-bytowe, tryb życia.
3.2.1. Czynniki prenatalne
Do niedawana sądzono, że noworodek swoją przygodę z mikrobiomem rozpoczyna
w chwili narodzin. Ostatnie doniesienia sugerują, że zarówno flora bakteryjna matki
jak i jej stan zdrowia mają wpływ na kształtowanie się flory bakteryjnej dziecka już
w okresie ciąży.
Przez dłuższy czas uważano, że środowisko wewnątrzmaciczne jest jałowe. Jednak
ostatnie badania zaprzeczają tym faktom. Przede wszystkim wykazano obecność
bakterii w smółce czyli pierwszym stolcu noworodka. Badania przeprowadzone przez
grupę badawczą Juan M. Rodriguez na 21 próbkach smółki pobranych od nowo-
rodków, urodzonych w sposób naturalny bądź przez cięcie cesarskie, dowiodły
obecności w nich przede wszystkim bakterii z rodzaju Enterococcus, głównie gatunku
E. fecalis. Był obecny w 17 z 21 próbek. Drugi, występujący w znacznej ilości, rodzaj
bakterii to Staphylococcus, z dominacją S. epidermidis. Został on wykryty w 10
badanych próbkach. Trzecia, dość licznie, występująca grupa bakterii to rodzaj
Escherichia i Enterobacter, występujący odpowiednio w 5 i 6 próbkach. Ponadto
zidentyfikowano takie rodzaje bakterii jak: Streptococcus, Leuconostoc, Rothia,
Klebsiella, Bacteroides, Parabacteroides. Jednak ich obecność wykazano jedynie
w pojedynczych próbach [14]. Inne badania udowodniły obecność bakterii w łożysku,
płynie owodniowym, krwi pępowinowej. Były to głównie bakterie należące do nastę-
pujących rodzajów: Enterococcus, Streptococcus, Staphylococcus i Propionibacterium.
Mikroorganizmy te wykryto w próbkach pobranych od zdrowych noworodków
urodzonych w wyniku cięcia cesarskiego [15]. Badania wykazały, że populacje bakterii
obecne zarówno w płynie owodniowym jak i łożysku są do siebie podobne. Cechują
się one niską różnorodnością. Dominującym typem bakterii w łożysku i płynie
owodniowym jest typ Proteobacteria, zwłaszcza należące do niego gatunki Entero-
bacter, Escherichia i Shigella. Bakterie te były obecne również w smółce, ale
w mniejszych ilościach. Ponadto wykazano obecność, w łożysku, płynie owodniowym
oraz smółce noworodków, następujących rodzajów mikroorganizmów: Propioni-
bacterium, Streptococcus, Staphylococcus, Lactobacillus. Należy podkreślić, że
próbki, w których wykryto te bakterie tj. łożysko, płyn owodniowy oraz smółka,
pochodziły od par matka-dziecko [16]. Obecność bakterii w smółce może być konsek-
wencją ich przemieszczania się drogą krwiopochodną od matki. Potwierdzeniem tego
może być ich obecność we krwi pępowinowej [14, 15]. Ich obecność może się wiązać
z połykaniem płynu owodniowego przez płód. Bakterie pochodzące z płynu

Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
19
owodniowego oraz łożyska mogą odgrywać istotną rolę w początkowym etapie
kolonizacji jelita [16].
Okazuje się, że istotny wpływ na kształtowanie się mikrobiomu jelitowego nowo-
rodka ma stan zdrowotny matki. Potwierdzeniem tego, mogą być badanie przeprowa-
dzone przez zespół badawczy Hu i współpracowników [17]. Charakteryzowali oni
skład gatunkowy mikroorganizmów obecnych w smółce noworodków urodzonych
przez matki zdrowe oraz chorujące na cukrzycę typu 2. Wykazali, że skład mikro-
organizmów obecnych w smółce niemowląt urodzonych przez matki z cukrzycą typu 2
istotnie różni się od mikrobiomu smółki noworodków urodzonych przez zdrowe
kobiety. Ponadto mikrobiom występujący w smółce noworodków urodzonych przez
matki z cukrzycą typu 2 był podobny, pod względem składu, do mikrobioty ziden-
tyfikowanej w kale osób chorujących na wyżej wymienioną jednostkę chorobową [17].
3.2.2. Czynniki postnatalne
Do czynników postnatalnych mających istotny wpływ na kształtowanie się mikro-
biomu jelitowego noworodka zaliczamy przede wszystkim: rodzaj porodu, sposób
karmienia, wiek ciążowy. Coraz większą rolę przypisuję się również zastosowanemu
leczeniu w pierwszych dniach życia, zwłaszcza antybiotykoterapii.
3.2.2.1. Rodzaj porodu
Zarówno ilość jak i rodzaj bakterii nabytych podczas porodu ma istotne znaczenie
dla wykształcenia się fizjologicznej mikroflory jelitowej, co wpływa na rozwój prawi-
dłowej odpowiedzi immunologicznej. Korzystniejszy jest poród naturalny. W tym
przypadku noworodek kolonizowany jest przez bakterie pochodzące z mikroflory
pochwy i przewodu pokarmowego matki. Podczas porodu fizjologicznego fakulta-
tywne anaeroby takie jak Escherichia coli, Staphylococcus, Streptococcus, Lacto-
bacillus, Prevotella kolonizują jelito stwarzając dogodne warunki dla pojawienia się
takich rodzajów bakterii jak Bacteroides, Bifidobacterium [18, 19]. Natomiast podczas
cięcia cesarskiego noworodek kolonizowany jest głównie przez bakterie obecne na
skórze matki oraz pochodzące ze środowiska szpitalnego. W tym przypadku mikro-
biom jelitowy noworodka zawiera głównie bakterie rodzaju: Staphylococcus, Propioni-
bacterium, Corynebacterium, Clostridium. Okazuje się, że mikroflora dzieci urodzo-
nych poprzez cięcie cesarskie charakteryzuje się obniżoną ilością bakterii z rodzaju
Bifidobacterium oraz Bacteroides fragilis. Wykazano również, że kolonizacja jelita
noworodka przez Bacteroides i Bifidobacterium jest opóźniona. Należy zwrócić uwagę
na rolę bakterii z rodzaju Bifidobacterium w utrzymaniu prawidłowego stanu zdrowia.
Ponadto flora jelitowa niemowląt urodzonych poprzez cięcie cesarskie cechuje się
liczniejszym występowaniem gatunku Clostridium difficile, który jest przyczyną
przewlekłych biegunek u noworodków [13].
Liczne badania wykazały korelację pomiędzy cięciem cesarskim a zwiększonym
występowaniem takich chorób jak: alergie, astma, cukrzyca typu I , celiakia, otyłość [13].

Marta Kozieł, Karolina Włodarczyk, Dominika Wojton
20
3.2.2.2. Sposób karmienia
Mleko matki jest najlepszym pokarmem dla noworodka. Zawiera ono prebiotyki,
które pozytywnie wpływają na wzrost, niezbędnych do zachowania właściwego stanu
zdrowia, bakterii z rodzaju Bifidobacterium, Lactobacillus. Bakterie z rodzaju Bifido-
bacterium hamują wzrost patogennych mikroorganizmów oraz stymulują odpowiedź
immunologiczną. Mikrobiom niemowląt karmionych mlekiem sztucznym bądź wcześ-
nie wprowadzonymi pokarmami stałymi charakteryzuje się występowaniem dużej licz-
by bakterii: Clostridium difficile, Bacteroides fragilis, Escherichia coli oraz niską liczeb-
nością niezbędnych do zachowania zdrowia, bakterii z rodzaju Bifidobacterium [13].
3.2.2.3. Wiek ciążowy
Duży wpływ na prawidłową mikroflorę jelita ma też wiek ciążowy. Mikrobiom
dzieci urodzonych wcześniej, przed planowanym terminem rozwiązania ciąży, różni
się od mikrobioty donoszonych noworodków. Charakteryzuje się dominacją bakterii
z rodzaju Clostridium (zwłaszcza C. difficile), Enterococcus, Staphylococcus, rodziny
Enterobacteriaceae oraz obniżeniem liczby Lactobacillus, Bifidobacterium. Ponadto
wykazano, że kolonizacja przewodu pokarmowego wcześniaków jest opóźniona co
może mieć związek z występowaniem m.in.: martwiczego zapalenia jelita (NEC) [20, 21].
3.2.2.4. Antybiotykoterapia
Antybiotykoterapia silnie oddziałuje na mikroflorę jelitową noworodka. Stosowanie
antybiotyków u noworodków jest przyczyną spadku liczby bakterii z rodzaju Bifido-
bacterium oraz gatunku B. fragilis. Ponadto długotrwałe stosowanie antybiotyków
powoduje spadek ilości bakterii typu Bacteroides i wzrost liczebności Firmicutes.
Obserwuje się również znaczny wzrost organizmów patogennych, głównie z rodzaju
Clostridium oraz Klebsiella, które często bywają przyczyną stanów chorobowych,
w tym sepsy noworodkowej [20, 22]. Okazuje się, że stosowanie antybiotyków
w czasie ciąży w znaczący sposób wpływa na mikroflorę jelitową noworodków.
W tym przypadku obserwuje się opóźnioną kolonizację lub zmniejszenie ilości ko-
rzystnych bakterii tj. Lactobacillus, Bifidobacterium w mikrobiomie noworodków, co
może przyczynić się do rozwoju takich chorób jak: alergie, zespół jelita drażliwego czy
nieswoiste zapalenia jelit [23].
4. Wybrane dysbiozy jelitowy a stany chorobowe
Dysbiozy jelitowe czyli zaburzenia ilościowo-jakościowe mikroflory jelitowej
odgrywają istotną rolę w patogenezie i/lub progresji wielu jednostek chorobowych,
takich jak: martwicze zapalenie jelit (NEC), otyłość, astma, alergie, cukrzyca typu 1
oraz 2, zespół jelita nadwrażliwego, nieswoiste zapalenie jelit, nowotwory jelita
grubego [13].

Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
21
4.1. Martwicze zapalenie jelit (NEC)
Martwicze zapalenie jelit jest jedną z chorób najbardziej wyniszczającą przewód
pokarmowy noworodków, zwłaszcza wcześniaków. Jest to stan zapalny przebiegający
z martwicą krwotoczną. Najczęściej występuję w końcowym odcinku jelita krętego
oraz bliższym odcinku jelita grubego [24]. Jednym z potencjalnych mechanizmów tego
schorzenia jest zaburzenie mikroflory jelitowej noworodków, co sprzyja stanom
prozapalnym i pozwala na translokację patogenów przez nabłonek jelitowy [25]. Scho-
rzenie to występuje bardzo często u wcześniaków. Sugeruje się, że brak w pełni
wykształconej odpowiedzi immunologicznej oraz towarzysząca temu czynnościowa
niedojrzałość nabłonka jelitowego jest sprzyjającym czynnikiem występowania NEC.
Jak już wcześniej wykazano, kolonizacja przewodu pokarmowego noworodków przez
bakterie korzystne dla zdrowia, np. Bifidobacterium jest znacznie opóźniona. Sprzyja
to pojawieniu organizmów patogennych, a tym samym większej aktywności receptora
Toll-podobnego 4 (Toll like-receptor 4, TLR4). To z kolei aktywuje jądrowy czyn-nik
NF-κ-B i geny, których aktywność związana jest z powstaniem procesu zapalnego.
Nadaktywność TLR4 wpływa także na wzrost stężenia syntazy tlenku azotu, która
również odgrywa istotną rolę w rozwoju stanu zapalnego [24].
Komensalne mikroorganizmy przyczyniają się do tolerancji patogennych organiz-
mów poprzez zapobieganie ich rozpoznawania przez receptory TLR4 bądź zmniej-
szenie aktywności prozapalnego szlaku NF-κ-B [26]. Mimo, że wykazano związek
pomiędzy rozwojem NEC a obecnością takich patogennych gatunków jak Klebsiella
czy Clostridium, to jednak nie przypisuje im się konkretnej roli w patogenezie tego
schorzenia. Przyczynę tego schorzenia upatruje się raczej w ogólnej dysbiozie jelito-
wej. Wykazano, że noworodki u których wystąpił NEC mają zmienioną florę bakte-
ryjną w porównaniu ze zdrowymi niemowlętami. Zmiany dotyczyły głównie zwięk-
szonej liczebności Enterobacter cloacae, Escherichia coli przy jednoczesnym
zmniejszeniu liczby Streptococcus faecalis oraz gatunku Staphylococcus. Zmiany
mikroflory zostały zaobserwowane 72 h przed wystąpieniem objawów NEC [26].
4.2. Otyłość i cukrzyca
Otyłość zaliczana jest do chorób cywilizacyjnych. Jest to schorzenie o złożonej
etiologii, które może być warunkowane zarówno poprzez czynniki genetyczne jak
i środowiskowe. Przyjmuje się, że często przyczyną otyłości są nieprawidłowe nawyki
żywieniowej, powodujące wysoki dodatni bilans energetyczny oraz brak aktywności
fizycznej. W ostatnich latach wykazano, że mikrobiom jelitowy i jego zaburzenia
ilościowo-jakościowe mogą odgrywać istotną rolę w patogenezie otyłości.
Wykazano, że mikroflora jelitowa osób otyłych różni się od mikroflory osób
o prawidłowej wadze. Na podstawie analizy sekwencji nukleotydowej genu 16S
rRNA, przeprowadzonej przez Ley i współpracowników, wykazano, że mikroflora
jelitowa osób otyłych cechuje się niską liczbą bakterii typu Bacteroidetes i wzrostem
ilości Firmicutes [27-29]. Stosunek Firmicutes do Bacteroidetes pozytywnie koreluje
z występowaniem otyłości niezależnie od diety [28]. Kolonizacja tzw. myszy „germ-
free” czyli wolnych od mikroorganizmów mikroflorą jelita grubego pochodzącą od

Marta Kozieł, Karolina Włodarczyk, Dominika Wojton
22
naturalnie wyhodowanych myszy przyczyniła się do zwiększenie zawartości tłuszczu
w organizmie w ciągu 10-14 dni pomimo zmniejszenia ilości spożywanego pokarmu
[30]. Okazuje się, że mikrobiota jelitowa może być kluczowym czynnikiem w patoge-
nezie otyłości oraz często jej towarzyszących innych zaburzeń metabolicznych, np.
cukrzyca. Mikroflora jelitowa oddziałuje na homeostazę energetyczną oraz magazy-
nowanie spożytych kalorii. Wymienia się następujące mechanizmy oddziaływania
mikrobiomu na gospodarkę energetyczną organizmu:
Jelitowy mikrobiom w wyniku wydzielania licznych związków chemicznych
przyczynia się do zwiększenia gęstości małych naczyń włosowatych w jelicie
cienkim, co sprzyja zwiększonemu wchłanianiu substancji odżywczych m.in.
monosacharydów pochodzących z diety [29],
Liczne roślinne polisacharydy nie są trawione przez ludzki organizm. Obecne
w jelicie bakterie wytwarzają hydrolazy glikozydowe, które umożliwiają fermen-
tację tych polisacharydów do takich związków jak: krótkołańcuchowe kwasy
tłuszczowe (short chain fatty acids, SCFAs) tj. głównie octan, propionian, maślan.
Mają one istotne znaczenie w procesie dojrzewania i ochrony kolonocytów [29]
[28]. SCAFs są ligandami dla receptorów sprzężonych z białkiem G (G protein-
coupled receptors, GPCR) tj. GPR41, GPR43, GPR109A, obecnych na endokryn-
nych komórkach jelitowych. Aktywacja tych receptorów stymuluje wydzielanie
hormonu – peptydu YY (PYY), który spowalnia motorykę jelit oraz ułatwia
wchłanianie substancji odżywczych, co może wskazywać na jego rolę w pato-
genezie otyłości. GPR43 reguluje metabolizm energetyczny przez supresję wrażli-
wości na insulinę adipocytów oraz zwiększenie wrażliwości na ten hormon takich
narządów jak wątroba czy mięśnie [29] [28].
Mikrobiom jelitowy hamuje aktywność tkankowego czynnika indukowanego
głodzeniem (fasting-induced adipose factor, FIAF), który z kolei inaktywuje
lipazę lipoproteinową (lipoprotein lipase, LPL). LPL jest odpowiedzialna za
rozkład trigliceroli zawierających lipoproteiny do kwasów tłuszczowych. Niska
ekspresja FIAF stymuluje magazynowanie triglicerydów w adipocytach [28].
Bakterie jelitowe hamują aktywność kinazy białkowej aktywowanej przez AMP
(adenosine monophosphate-activated protein kinase, AMPK). Prowadzi to do
zmniejszenia intensywności mitochondrialnej oksydacji kwasów tłuszczowych,
wychwytu glukozy, ketogenezy, wydzielania insuliny oraz zwiększenia lipo-
genezy i syntezy cholesterolu, triglicerydów, co ma istotny wpływ na metabolizm
lipidów [28] [29].
Mikrobiom jelitowy przyczynia się do otyłości indukowanej dietą wysokotłusz-
czową przez oddziaływanie na receptor farnezoidowy X (farnesoid X receptor,
FXR), który wpływa na akumulację triglicerydów wątrobowych [28].
Wszystkie powyższe mechanizmy są zależne od mikrobiomu jelitowego i modulują
homeostazę jelitową oraz wpływają na magazynowanie tłuszczów, co ma istotne
znaczenie w patogenezie otyłości.

Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
23
Bardzo często otyłości towarzyszy szereg chorób np. cukrzyca czy choroby
sercowo-naczyniowe. Wykazano związek pomiędzy otyłością a cukrzycą tj. insulino-
opornością. Pomostem łączącym te dwa schorzenia jest stan zapalny wywołany przez
aktywność mikroflory jelitowej [28]. Cani i współpracownicy [31] postawili hipotezę,
że lipopolisacharyd (LPS), będący endotoksyną występującą u bakterii gram
ujemnych, jest czynnikiem przyczyniającym się do rozwoju chorób metabolicznych
indukowanych wysokotłuszczową dietą. Wykazali zwiększone stężenie LPS-u u osób
otyłych. Sugerują, że dieta bogata w tłuszcze przyczynia się do zmiany mikrobiomu
jelitowego, co prowadzi do zmiany przepuszczalności jelitowych barier nabłonkowych
i zwiększonego stężenia LPS, który wędruje m.in. do wątroby, co przyczynia się do
powstania stanu zapalnego [31]. LPS jest jednym z czynników inicjujących proces
zapalny. Może być uwalniany w wyniku śmierci bakterii gram- ujemnych. LPS prze-
chodzi przez błonę śluzową przewodu pokarmowego poprzez mechanizm zależny od
TLR4. Ponadto może być transportowany z jelita do tkanki docelowej poprzez chylo-
mikrony, które są syntetyzowane w komórkach nabłonkowych jelita w odpowiedzi na
dietę bogatą w tłuszcze. Gdy LPS dostanie się np. do wątroby, wiąże się on do białka
wiążącego LPS (LPS-binding protein, LPB). Dochodzi do aktywacji CD14 obecnego
w błonie komórkowej makrofagów, kompleks ten następnie wiąże się z receptorem
TLR4 komórek odpornościowych. W konsekwencji dochodzi do wydzielenia cytokin
prozapalnych, m.in.: interleukiny-1, interleukiny-6, czynnika martwicy nowotworów α
(tumor necrosis factor α, TNF-α) czy inhibitora aktywatora plazminogeny typu 1
(plasminogen activator inhibitor-1, PAI-1). Cytokiny te przyczyniają się do wzrostu
odporności na insulinę, np. TNF-α hamuje aktywność kinaz przez nadmierną
fosforylację seryny/treoniny w IRS-1. Konsekwencją oporności na insulinę może być
cukrzyca, hiperinsulinemia oraz nadmierne magazynowanie lipidów w wątrobie
i tkance tłuszczowej [28, 31].
4.3. Alergia
Mikroflora jelitowa wpływa na wykształcenie się prawidłowej odpowiedzi immu-
nologicznej, która charakteryzuje się m.in. równowagą pomiędzy limfocytami
Th1//Th2. Istotną rolę w oddziaływaniu na limfocyty Th1 i Th2 wykazują limfocyty
regulatorowe T. Płód wykazuje zwiększoną ekspresję Th2, natomiast tuż po porodzie
prawidłowa mikroflora bakteryjna aktywuje Th1. W przypadku zaburzeń mikrobiomu
dochodzi do spadku aktywności Th1. Wówczas Th2 stymulują wytwarzanie IL-4,
IL-5, a te indukują produkcję IgE i eozynofilów, co jest przyczyną m.in. alergii pokar-
mowych. U dzieci cierpiących na alergię obserwuje się spadek liczebności bakterii
z rodzaju Lactobacillus, Bacteroides, Bifidobacterium, a wzrost bakterii E. coli, S.
aureus, Clostridium [32]. Wskazuje to, że wykształcenie się prawidłowej flory bakte-
ryjnej już od momentu narodzin jest niezmiernie ważne, gdyż to ona w istotny sposób
moduluje układ immunologiczny. Wykazano, że dzieci, które przyszły na świat po-
przez cięcie cesarskie były kolonizowane przez mniejszą liczbę bakterii typu Bactero-
idetes, jak również cechowały się one mniejszą różnorodnością własnego mikrobiomu,
i tym samym wykazywały większe ryzyko rozwoju alergii [33].

Marta Kozieł, Karolina Włodarczyk, Dominika Wojton
24
4.4. Nieswoiste zapalenie jelit
Wśród nieswoistych zapaleń jelit (IBD) wyróżniamy chorobę Leśniowskiego-
Crohna (CD) oraz wrzodziejące zapalenie jelita grubego. Osoby chorujące na IBD
wykazują, zwiększone o 10-15%, ryzyko zachorowania na raka jelita grubego [34].
Uważa się, że rozwój IBD jest związany z występowaniem zaburzeń jakościowo-
ilościowych mikrobiomu jelitowego. Badania wykazały wzrost ilości bakterii typu
Proteobacteria oraz zmniejszenie liczebności bakterii należących do typu Firmicutes
w próbkach kału pacjentów z IBD w porównaniu do grupy kontrolnej [34].
Badania przeprowadzone przez Gevers`a i współpracowników [36] wykazały
w mikrobiomie chorych na CD zwiększoną ilość bakterii należących do Enterobac-
teriaceae, Veillonellaceae, Pasteurellacaea, Fusobacteriaceae [36] Neisseriaceae [35]
oraz zmniejszenie liczby Bifidobacteriaceae [35], Erysipelotrichales, Bacteroidales,
Clostridales [36]. Okazuje się, że stosowanie antybiotyków w znaczny sposób przy-
czynia się do dysbiozy jelitowej charakterystycznej dla pacjentów z chorobą
Leśniowskiego-Crohna. [35] [36]. Mikroflora chorych na CD zawiera niewielką liczbę
bakterii o właściwościach ochronnych, przeciwzapalnych, np. Faecalibacterium
prausnitzii, Clostridium leptum, Clostridium coccoides. Wykazano, że bakterie nale-
żące do rodzaju Clostridium wytwarzają maślan należący do krótkołańcuchowych
kwasów tłuszczowych (Short Chain Fatty Acids, SCFAs). Jest on istotnym źródłem
substancji odżywczych dla kolonocytów, wykazuje działanie antybakteryjne oraz
przeciwzapalne poprzez obniżenie poziomu cytokin prozapalnych. Natomiast zwięk-
szona liczba bakterii gram ujemnych może przyczyniać się, poprzez zwiększone
stężenie endotoksyny LPS, do wystąpienia stanu zapalnego charakterystycznego dla
omawianych chorób [37].
5. Podsumowanie
Mikrobiota to zespół organizmów zasiedlający organizm gospodarza warunkujący
homeostazę. Wyróżniamy mikrobiotę układu oddechowego, skóry, układu moczowo-
płciowego oraz pokarmowego. Największe znaczenie obecnie przypisuje się mikro-
biocie przewodu pokarmowego. Do najważniejszych funkcji mikrobioty zaliczamy
modulację układu immunologicznego oraz jego funkcje troficzne i metaboliczne.
Okazuje się, że już okres prenatalny, tj. stan zdrowotny matki oraz jej własna flora
bakteryjna, odgrywa istotną rolę w kształtowaniu się prawidłowej mikroflory jelitowej.
Ponadto wpływ na powstanie fizjologicznej flory jelitowej ma rodzaj karmienia,
sposób porodu, wiek ciążowy czy zastosowana antybiotykoterapia. Okazuje się, że
mikrobiom dzieci które przyszły na świat poprzez cesarskie cięcie, były karmione
sztucznym mlekiem czy są wcześniakami wykazuje istotne zaburzenia ilościowo-
jakościowe tzw. dysbiozy jelitowe. Zaburzenia te odgrywają rolę w patogenezie bądź
progresji chorób takich jak: otyłość, cukrzyca, alergie, martwicze zapalenie jelit,
nieswoiste zapalenie jelit oraz wielu innych.

Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
25
Literatura
1. Costello E.K., Stagaman K., Dethlefsen L., Bohannan B.J., Relman D.A. The application of
ecological theory toward an understanding of the human microbiome, Science., 336
(2012), s. 1255-1262.
2. Strzępa A., Szczepanik M. Wpływ naturalnej flory jelitowej na odpowiedź immunologiczną,
Postępy higieny i medycyny doświadczalnej., 67 (2013), s. 908-920.
3. Hattori M., Taylor T.D. The Human Intestinal Microbiome: A New Frontier of Human
Biology, DNA Research., 16 (1) (2008), s. 1-12.
4. Mroczyńska M., Libudzisz Z., Gałęcka M., Szachta P. Mikroorganizmy jelitowe człowieka
i ich aktywność metaboliczna, Przegląd Gastroenterologiczny., 6(4) (2011), s. 1-7
5. Turnbaugh P.J., Ley R.E., Hamady M., Fraser-Ligget C., Knight R., Gordon J.I. The human
microbiome project: exploring the microbial part of ourselves in a changing world,
Nature., 449 (7164) (2007), s. 804-810.
6. Olszewska J., Jagusztyn-Krynicka E.K. Human Microbiome Project – mikroflora jelit oraz
jej wpływ na fizjologię i zdrowie człowieka, Postępy Mikrobiologii., 4 (2012), s. 243-256.
7. Gill S.R., Pop M., DeBoy R.T., Eckburg P.B., Turnbaugh P.J., Samuel B.S., Gordon J.I.,
Relman D.A., Fraser-Liggett C.M., Nelson K.E. Metagenomic Analysis of the Human
Distal Gut Microbiome., Science, 308 (5778) (2006), s. 1355-1359.
8. Eckurbg P.B., Bik E.M., Bernstein C.N., Purdom E., Dethlefsen L., Sargent M., Gill S.R.,
Nelson K.E., Relman D.A. Diversity of the human intestinal microbial flora, Science., 308
(5728) (2005), s. 1635-1638.
9. Nam Y.D., Chang H.W., Kim K.H., Roh S.W., Kim M.S., Jung M.J., Lee S.W., Kim J.Y.,
Yoon J.H., Bae J.W. Bacterial, archaeal, and eukaryal diversity in the intestines of Korean
people, The Journal of Microbiology., 46(5) (2008), s. 491-501.
10. Oxley A.P., Lanfranconi M.P., Würdemann D., Ott S., Schreiber S., McGenity T.J.,
Timmis K.N., Nogales B. Halophilic archaea in the human intestinal mucosa,
Environmental microbiology., 12(9) (2010), s. 2398-2410.
11. Rodríguez J.M., Murphy K., Stanton C., Ross R.P., Kober O.I., Juge N., Avershina E., Rudi
K., Narbad A., Jenmalm M.C., Marchesi J.R., Collado M.C. The composition of the gut
microbiota throughout life, with an emphasis on early life, Microbial Ecology in Health and
Disease., 2(26) (2015), https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4315782/
12. Górska S., Jarząb A., Gamian A. Bakterie probiotyczne w przewodzie pokarmowym
człowieka jako czynnik stymulujący układ odpornościowy, Postępy Higieny i Medycyny
Doświadczalnej., 63 (2009), s. 653-667.
13. Mueller N.T., Bakacs E., Combellick J., Grigoryan Z., Dominguez-Bello M.G. The infant
microbiome development: mom matters, Trends in Molecular Medicine., 21(2) (2015),
s. 109-117.
14. Jiménez E., Marín M.L., Martín R., Odriozola J.M., Olivares M., Xaus J., Fernández L.,
Rodríguez J.M. Is meconium from healthy newborns actually sterile?, Research in
Microbiology., 159(3) (2008), s. 187-193.
15. Jiménez E., Fernández L., Marín M.L., Martín R., Odriozola J.M., Nueno-Palop C., Narbad
A., Olivares M., Xaus J., Rodríguez J.M. Isolation of commensal bacteria from umbilical
cord blood of healthy neonates born by cesarean section, Current Microbiology., 51(4)
(2005), s. 270-274.
16. Collado M.C., Rautava S., Aakko J., Isolauri E., Salminen S. Human gut colonisation may
be initiated in utero by distinct microbial communities in the placenta and amniotic fluid,
Scientific Reports, 6 (2016), s. 1-13.

Marta Kozieł, Karolina Włodarczyk, Dominika Wojton
26
17. Hu J., Nomura Y., Bashir A., Fernandez-Hernandez H., Itzkowitz S., Pei Z., Stone J.,
Loudon H., Peter I. Diversified microbiota of meconium is affected by maternal diabetes
status, PLOS ONE., 8(11) (2013), s. e78257.
18. Dominguez-Bello M.G., Costello E.K., Contreras M., Magris M., Hidalgo G., Fierer N.,
Knight R. Delivery mode shapes the acquisition and structure of the initial microbiota
across multiple body habitats in newborns, Proceedings of the National Academy of
Sciences of the United States of America., 107(26) (2010), s. 11971-11975.
19. Pantoja-Feliciano I.G., Clemente J.C., Costello E.K., Perez M.E., Blaser M.J., Knight R.,
Dominguez-Bello M.G. Biphasic assembly of the murine intestinal microbiota during early
development, Multidisciplinary Journal of Microbial Ecology., 7(6) (2013), s. 1112-1115.
20. Penders J., Thijs C., Vink C., Stelma F.F., Snijders B., Kummeling I., van den Brandt P.A.,
Stobberingh E.E. Factors influencing the composition of the intestinal microbiota in early
infancy, Pediatrics, 118(2) (2006), s. 511-521.
21. Bartnicka A., Gałęcka M., Mazela J., Wpływ czynników prenatalnych i postnatalnych na
mikrobiotę jelitową noworodków, Standardy medyczne/pediatria., 13 (2016), s. 165-172.
22. Faa G., Gerosa C., Fanni D., Nemolato S., van Eyken P., Fanos V. Factors influencing the
development of a personal tailored microbiota in the neonate, with particular emphasis on
antibiotic therapy, The Journal of Maternal-Fetal & Neonatal Medicine., 26 (S2) (2013),
s. 35-43.
23. Munyaka P.M., Khafipour E., Ghia J.E. External influence of early childhood
establishment of gut microbiota and subsequent health implications, Frontiers in
Pediatrics., 2 (2014), s. 1-9.
24. Sharma R., Young C., Mshvildadze M., Neu J. Mikroflora jelit: czy ma znaczenie
w chorobach u noworodków?, Pediatria po dyplomie., 14(1) (2010), s. 70-87.
25. Elgin T.G., Kern S.L., McElroy S.J. Development of the Neonatal Intestinal Microbiome
and Its Association With Necrotizing Enterocolitis, Clinical Therapeutics., 38(4) (2016), s.
706 -715.
26. Patel R.M., Denning P.W. Intestinal microbiota and its relationship with necrotizing
enterocolitis, Pediatric Research., 78(3) (2016), s. 232-238.
27. Ley R.E., Bäckhed F., Turnbaugh P., Lozupone C.A., Knight R.D., Gordon J.I. Obesity
alters gut microbial ecology, Proceedings of the National Academy of Sciences of the
United States of America., 102(31) (2005), s.11070 -11075.
28. Boulangé C.L., Neves A.L., Chilloux J., Nicholson J.K., Dumas M.E. Impact of the gut
microbiota on inflammation, obesity, and metabolic disease, Genome Medicine., 8(1)
(2016), DOI: 10.1186/s13073-016-0303-2
29. Stachowicz N., Kirsztan A. Rola mikroflory jelitowej w patogenezie otyłości i cukrzycy,
Postępy Higieny i Medycyny Doświadczalnej., 67 (2013), s. 288-303.
30. Turnbaugh P.J., Ley R.E., Mahowald M.A., Magrini V.,2, Mardis E.R., Gordon J.I. An
obesity-associated gut microbiome with increased capacity for energy harvest, Nature., 444
(2006), s. 1027-1031.
31. Cani P.D., Amar J., Iglesias M.A., Poggi M., Knauf C., Bastelica D., Neyrinck A.M., Fava
F., Tuohy K.M., Chabo C., Waget A., Delmée E., Cousin B., Sulpice T., Chamontin B.,
Ferrières J., Tanti JF., Gibson G.R., Casteilla L., Delzenne N.M., Alessi M.C., Burcelin R.
Metabolic endotoxemia initiates obesity and insulin resistance, Diabetes., 56(7) (2007), s.
1761-1772.
32. Riiser A. The human microbiome, asthma, and allergy, Allergy, Asthma & Clinical
Immunology., 11 (2015), DOI: 10.1186/s13223-015-0102-0

Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
27
33. Jakobsson H.E., Abrahamsson T.R., Jenmalm M.C,, Harris K., Quince C., Jernberg C.,
Björkstén B., Engstrand L., Andersson A.F. Decreased gut microbiota diversity, delayed
Bacteroidetes colonisation and reduced Th1 responses in infants delivered by caesarean
section, Gut., 63(4) (2014), s. 559-566.
34. Tomasello G. Tralongo P., Damiani P., Sinagra E., Di Trapani B., Zeenny M.N., Hussein
I.H., Jurjus A., Leone A. Dismicrobism in inflammatory bowel disease and colorectal
cancer: changes in response of colocytes, World journal of gastroenterology., 20(48)
(2014), s. 18121-18130.
35. Hansen J.J., Sartor B.R. Therapeutic Manipulation of the Microbiome in IBD: Current
Results and Future Approaches, Current Treatment Options in Gastroenterology., 13(1)
(2015), S. 105-120.
36. Gevers D., Kugathasan S., Denson L.A., Vázquez-Baeza Y., Van Treuren W., Ren B.,
Schwager E., Knights D., Song S.J., Yassour M., Morgan X.C., Kostic A.D., Luo C.,
González A., McDonald D., Haberman Y., Walters T., Baker S., Rosh J., Stephens M.,
Heyman M., Markowitz J., Baldassano R., Griffiths A., Sylvester F., Mack D., Kim S.,
Crandall W., Hyams J., Huttenhower C., Knight R., Xavier R.J. The treatment-naive
microbiome in new-onset Crohn's disease, Cell host & microbe., 15(3) (2014), s. 382-392.
37. Franczuk A., Jagusztyn-Krynicka E. K. Rola mikroflory jelit w indukcji choroby
Leśniowskiego-Crohna w świetle programu badań Human Microbiome Project, Postępy
Mikrobiologii., 51(4) (2012), s. 257-264.
38. Dethlefsen L., Eckburg P.B., Bik E.M., Relman D.A. Assembly of the human intestinal
microbiota, Trends in ecology & evolution., 21(9) (2006), s. 517-523.
39. Bearfield C., Davenport E.S., Sivapathasundaram V., Allaker R.P. Possible association
between amniotic fluid micro-organism infection and microflora in the mouth, BJOG: An
International Journal of Obstetrics & Gynaecology., 109(5) (2002), S. 527-533.
Kształtowanie się mikrobioty jelitowej noworodków a konsekwencje zdrowotne
Streszczenie
Celem pracy była charakterystyka struktury i funkcji mikrobiomu jelitowego oraz czynników prenatalnych
i postnatalnych mających wpływ na kształtowanie się prawidłowej flory jelitowej. Mikrobiom wcześ-
niaków, bądź dzieci urodzonych poprzez cesarskie cięcie, karmionych sztucznym mlekiem, wykazuje po-
ważne zaburzenia ilościowo-jakościowe, określane jako dysbiozy jelitowe. Dysbiozy jelitowe mają udział
w patogenezie i/lub progresji licznych chorób, np. otyłości, cukrzycy, alergii, martwiczego zapalenia jelita, nieswoistym zapaleniu jelit.
Słowa kluczowe: mikrobiom, otyłość, cukrzyca, mikroorganizmy, noworodek
Forming of intestinal microbiote infants and health implications
Abstract
The aim of the study is to characterize the structure and function of intestinal microbiome and prenatal and
postnatal factors which have a significant influence on the development of normal intestinal flora. Micro-biome of premature babies or babies born via caesarean section who are fed with artificial milk exhibits
qualitative quantitive disorders which are called intestinal dysbiosis. Intestinal dysbiosis has an impact on
pathogenesis and/or progression of numerous diseases, such as obesity, diabetes, allergies, necrotising
enterocolitis, and inflammatory bowel disease. Keywords: microbiome, obesity, diabetes,microorganisms, infant

28
Andrzej Jurkowski1, Ita Szczepańska
2, Jacek J. Kozioł
3
Wpływ nanocząstek tlenku żelaza
na produkcję kwasu mlekowego
w hodowli okresowej Lactobacillus rhamnosus GG
1. Wstęp
1.1. Nanocząstki
Dotychczas w obszarze mikrobiologii nanocząstki metali i ich tlenków stosowano
głównie w celach bakteriobójczych i bakteriostatycznych. Badania w tym zakresie
skupiają się głównie na poszukiwaniu nowych metod syntezy nanocząstek srebra,
a następnie wykazaniu ich właściwości antybakteryjnych i antystatycznych [1-3].
Nanocząstki srebra dzięki swoim właściwościom są także wykorzystywane do ograni-
czania ilości insektów [4]. W sprzedaży dostępne są różnorakie produkty zawierające
lub pokryte nanocząstkami srebra np. bielizna, kosmetyki, sprzęt AGD.
Pierwsze próby zastosowania nanocząstek w celach pozytywnie oddziałujących na
mikroorganizmy wykonał zespół A. Jurkowskiego [5].
W badaniach zastosowano nanocząstki tlenku żelaza (II) diżelaza (III) (Fe3O4), które
wykazują właściwości magnetyczne oraz są uznawane jako nieszkodliwe dla organiz-
mu człowieka. Są one przez organizm ludzki najlepiej tolerowanymi materiałami
magnetycznymi. Obecnie są jedynym zaakceptowanym materiałem magnetycznym do
zastosowań w diagnostyce i terapii medycznej. Stosowane są głównie jako kontrast do
rezonansu magnetycznego (MRI – Magnetic Resonanse Imagining) [6]. Dzięki swoim
właściwościom magnetycznym wykorzystuje się je także jako transportery/przenośniki
leków, gdzie po przyłożeniu zewnętrznie pola magnetycznego możemy zwiększyć
koncentrację danego leku w danym obszarze ciała (w miejscu zmienionym choro-
bowo). Kolejnym przykładem są terapie leczenia chorób nowotworowych podczas
których próbuje się stosować hipertermię, czyli przegrzanie komórek nowotworowych.
Nanocząstki Fe3O4 wykazują również właściwości nanobuforujące opisane przez
zespół B. Zapotocznego [7]. Zdolność do nanobuforowania polega na podwyższeniu
lub obniżeniu wartości pH roztworu, w którym znajdują się nanocząstki tlenku
żelaza(II) diżelaza(III) w zależności od odczynu tego roztworu.
W roztworach o odczynie kwasowym obecność nanocząstek Fe3O4 podwyższa
wartość pH roztworu, natomiast w roztworach o odczynie zasadowym ich obecność
wpływa na obniżenie wartości pH roztworu. Niezależnie więc od odczynu nanocząstki 1 [email protected], Katedra Biotechnologii, Wydział Nauk Biologicznych, Uniwersytet
Zielonogórski, uczelnia, http://wnb.uz.zgora.pl/pl/ 2 [email protected], Katedra Biotechnologii, Wydział Nauk Biologicznych, Uniwersytet
Zielonogórski, uczelnia, http://wnb.uz.zgora.pl/pl/ 3 [email protected], Katedra Biotechnologii, Wydział Nauk Biologicznych, Uniwersytet
Zielonogórski, uczelnia, http://wnb.uz.zgora.pl/pl/

Wpływ nanocząstek tlenku żelaza na produkcję kwasu mlekowego
w hodowli okresowej Lactobacillus rhamnosus GG
29
neutralizują pH zbliżając jego wartości do pH=7. Maksymalna wartość zmian – ∆pH ≈
0,5.
Tę własność nanocząstek tlenku żelaza wykorzystał zespół A. Jurkowskiego.
W warunkach obniżonego pH wykazano pozytywny wpływ nanocząstek tlenku
żelaza(II) diżelaza(III) na przeżywalność bakterii probiotycznych Lactobacillus
acidophilus. W badaniach tych wskazano konieczność zastosowania nanocząstek
Fe3O4 w hodowlach bakterii mlekowych celem sprawdzenia wpływu nanocząstek
tlenku żelaza na produktywność tych bakterii [5].
Badania opisane w niniejszej pracy stanowią kontynuację powyższych badań.
Możliwość zastosowania nanocząstek tlenku żelaza(II) diżelaza(III) w celu zwięk-
szenia produkcji kwasu mlekowego jest ekonomicznie „pożądana”. Zastosowanie
nanocząstek tlenku żelaza w trakcie hodowli bakterii mlekowych neutralizowałoby
wartość pH bulionu hodowlanego, które jest obniżane produkowanym przez bakterie
kwasem mlekowym. Dodatek nanocząstek Fe3O4 do bioreaktora mógłby zmniejszyć
lub wyeliminować konieczność dodawania wodorotlenku sodu lub wapnia stan-
dardowo dodawanych do neutralizacji pH.
1.2. Bakterie mlekowe
Bakterie mlekowe stanowią bardzo różnorodną grupę mikroorganizmów. Zasiedlają
wiele różnych stanowisk, jak również bytują na i wewnątrz innych organizmów. Mają
duże zapotrzebowanie na składniki pokarmowe, najczęściej spotykane są w środo-
wisku bogatym w węglowodany, aminokwasy i pochodne nukleotydów. Ponadto
wykazują one duże przystosowania do trudnych warunków, dzięki czemu spektrum
zamieszkiwanych przez nie nisz znacząco się zwiększa. Wiele z nich bytuje w układzie
pokarmowym zwierząt, w tym człowieka i stanowi jego niezbędną mikroflorę [8].
Bakterie kwasu mlekowego są wykorzystywane w przemyśle spożywczym,
głównie w procesach przetwarzania mleka i surowców roślinnych. Są również uży-
wane w procesach fermentacji zakwasu chlebowego w piekarnictwie, podczas przepro-
wadzania fermentacji mięsa w produkcji wędlin fermentowanych. W przemyśle
mleczarskim przygotowuje się odpowiednio zestawione szczepy bakterii mlekowych
tzw. Startery mleczarskie. Są one następnie wykorzystywane do produkcji serów
twarogowych, śmietany, jogurtów, serów podpuszczkowych i wielu innych rodzajów
produktów mleczarskich [9].
W Polsce przeprowadza się na wielką skalę procesy fermentacji (mlekowej) kapu-
sty, ogórków, sałatek wielowarzywnych i soków warzywnych. Procesy fermentacji
mlekowej są stosowane w krajach Azji i Afryki jako jedne z najważniejszych metod
utrwalania żywności. Kolejnym ważnym zastosowaniem fermentacji mlekowej
i bakterii mlekowych jest kiszenie pasz dla zwierząt. Do zakiszana stosuje się odpady
z przemysłu spożywczego, zieloną masę roślinną, warzywa i inne surowce [9].
Odgrywają również ważną rolę w przemyśle farmaceutycznym, gdzie są wykorzys-
tywane do produkcji wielu związków chemicznych m.in. dekstranu, do produkcji
żywności probiotycznej jak również preparatów probiotycznych [10].

Andrzej Jurkowski, Ita Szczepańska, Jacek J. Kozioł
30
1.3. Produkcja kwasu mlekowego
Znaczna część (95%) światowej produkcji kwasu mlekowego produkowana jest
z wykorzystaniem surowców roślinnych i bakterii mlekowych [11].
W zależności od tego jakie cechy produktu końcowego są pożądane, dobiera się
odpowiednie szczepy produkcyjne. Najczęściej preferowane są szczepy termofilne
bakterii mlekowych, które produkują kwas mlekowy w krótkim czasie. Ich zaletą jest
to, że wytwarzają niewiele produktów ubocznych. Dodatkowo możliwość prowadzenia
procesu fermentacji w podwyższonej temperaturze przeciwdziała rozwojowi niepo-
żądanych drobnoustrojów, które mogłyby zakłócić przebieg procesu fermentacji
mlekowej.
Zaletą stosowania fermentacji mlekowej w przeciwieństwie do metod syntezy che-
micznej jest możliwość otrzymania odpowiedniej formy kwasu mlekowego (D lub L,
ewentualnie mieszaniny racemicznej).
Ponieważ bakterie mlekowe potrafią syntetyzować kwas mlekowy w jednej z trzech
form:
forma prawoskrętna (L+),
forma lewoskrętna (D-),
mieszanina racemiczna DL.
W syntezie chemicznej uzyskujemy jedynie mieszaninę racemiczną kwasu
mlekowego.
Zwiększenie ilości uzyskiwania kwasu mlekowego w procesie fermentacji mle-
kowej wpłynęłoby pozytywnie na światową produkcję tego kwasu. Zmniejszyłoby
koszty uzyskiwania, a tym samym cenę gotowego produktu.
2. Cel badań
Celem przeprowadzonych badań było sprawdzenie czy i w jaki sposób nanocząstki
tlenku żelaza(II) diżelaza(III) wpływają na produkcję kwasu mlekowego przez bakterie
probiotyczne Lactobacillus rhamnosus GG oraz jak obecność nanocząstek w bulionie
hodowlanym wpływa na liczebność bakterii w trakcie trwania hodowli.
3. Materiał i metody
Materiałem wykorzystywanym do badań były bakterie probiotyczne Lactoba-
cillus rhamnosus GG oraz zsyntetyzowane nanocząstki tlenku żelaza(II) diżelaza(III)
o średniej wielkości 20 nm. Nanocząstki Fe3O4 były syntetyzowane i mierzone zgodnie
z opisem w pracy A. Jurkowskiego i in. (2015). Lactobacillus rhamnosus GG należy
do bakterii mlekowych o właściwościach probiotycznych.
W celu określenia wpływu nanocząstek Fe3O4 na produktywność i liczebność
bakterii założono hodowle okresowe L. rhamnosus GG.
Hodowlę wyjściową Lactobacillus rhamnosus GG prowadzono w bulionie MRS
(Merck).
Hodowle okresowe zawierały 1,2 g nanocząstek Fe3O4, 190 ml mleka odtłucz-
czonego (BTL), 10 ml 24 godzinnej płynnej hodowli wyjściowej L. rhamnosus GG.

Wpływ nanocząstek tlenku żelaza na produkcję kwasu mlekowego
w hodowli okresowej Lactobacillus rhamnosus GG
31
Równolegle prowadzono kontrolę składającą się z 190 ml mleka odtłuszczonego
(BTL) i 10 ml 24 godzinnej płynnej hodowli wyjściowej Lactobacillus rhamnosus GG.
Hodowle prowadzono w warunkach beztlenowych, w temp. 37oC, w trzech
powtórzeniach.
W określonych odstępach czasu mierzono:
ilość bakterii jako jednostki tworzące kolonie w mililitrze bulionu hodowlanego
(jtk/ml),
wartość pH,
stężenie kwasu mlekowego (mg/ml).
Liczebność bakterii określano przez posiew w rozcieńczeniach na podłożu stałym
MRS (Merck) w trzech powtórzeniach. Próbki hodowli pobierano po 24, 48 i 72
godzinach hodowli. Ilość jtk/ml określano jako średnią z trzech płytek o tym samym
rozcieńczeniu.
Pomiar pH przeprowadzano w momencie rozpoczęcia hodowli a następnie po 24,
48, 72 godzinach hodowli za pomocą pehametru Multifunction Meter CX-501
(Elmetron). Dla każdej próby wykonano trzy pomiary.
Stężenie kwasu mlekowego określano na podstawie ilości zużytego NaOH przy
miareczkowaniu w obecności fenoloftaleiny. Z każdej próby pobierano 25 ml płynu
hodowlanego, które miareczkowano w obecności 1 ml 2% fenoloftaleiny 0,25 M
roztworem NaOH. Dla każdej próby wykonano dwa pomiary. Pomiary wykonywano
po 24, 48 i 72 godzinach hodowli.
Wszystkie wyniki poddano analizom statystycznym za pomocą testu T-studenta.
4. Wyniki
Wyniki uzyskane podczas badań obejmują zmiany wartości pH, liczebności bakterii
oraz stężenia kwasu mlekowego w próbie badawczej z nanoczastkami i próbie
kontrolnej. Uzyskane wyniki opracowano statystycznie i przedstawiono w poniższych
tabelach (tabele 1-3).
4.1. Wartość pH
Wartość pH hodowli wyjściowej wynosiła 6,6. Natomiast średnie wartości pH dla
badanych prób mierzone po 24, 48 i 72 godzinach przedstawiono w tabeli 1.
Tabela 1. Wartości pH badanych prób z nanocząstkami Fe3O4 i dla kontroli
Czas
[h]
Średnia wartość dla prób
z nanocząstkami Fe3O4
Średnia wartość dla
kontroli
p
0 6,613 6,613 -
24 5,788 6,085 0,20
48 5,559 5,542 0,84
72 5,703 5,537 0,24
Źródło: Opracowanie własne

Andrzej Jurkowski, Ita Szczepańska, Jacek J. Kozioł
32
4.2. Liczebność bakterii
Dla prób zarówno z nanocząstkami tlenku żelaza(II) diżelaza(III) jak i bez nano-
cząstek obserwowano wzrost liczebności bakterii w tracie trwania hodowli. Zauwa-
żono wolniejszy wzrost w próbach z nanocząstkami tlenku żelaza(II) diżelaza(III) (po
24 i 48 godzinach). Dla wyników otrzymanych po 72 godzinach nie stwierdzono
istotnych różnic. Wyniki obliczeń liczby bakterii w trakcie trwania hodowli okresowej
przedstawia tabela 2.
Tabela 2. Liczebność biomasy L. rhamnosus GG
Czas
[h]
Średnia z prób z
nanocząstkami Fe3O4
[jtk/ml]
Średnia z prób
kontrolnych [jtk/ml]
p
0 9,27*107
9,27*107 -
24 3,8*108 12,40*10
8 0,32
48 6,27*108 15,35*10
8 0,87
72 12,58*108 16,4*10
8 0,022
Źródło: Opracowanie własne
4.3. Stężenie kwasu mlekowego
W trakcie trwania hodowli dla prób zarówno z nanocząstkami tlenku żelaza(II)
diżelaza(III) jak i bez nanocząstek Fe3O4 obserwowano wzrost stężenia kwasu
mlekowego w bulionie hodowlanym. Zaobserwowano większy wzrost stężenia kwasu
mlekowego dla prób z nanocząstkami tlenku żelaza(II) diżelaza(III). Wyniki obliczeń
stężenia kwasu mlekowego w trakcie trwania hodowli okresowej przedstawia tabela 3.
Tabela 3. Stężenie kwasu mlekowego w trakcie hodowli okresowej L. rhamnosus GG
Czas
[h]
Średnia z prób z
nanocząstkami Fe3O4 [mg/ml]
Średnia z prób
kontrolnych [mg/ml]
p
0 0 0 -
24 3,93 3,20 0,22
48 5,59 6,14 0,34
72 7,56 6,86 0,57
Źródło: Opracowanie własne
5. Analiza wyników
Dotychczas nanocząstek tlenku żelaza(II) diżelaza(III) nie stosowano w celach
poprawy produktywności bakterii. Tylko w pracy zespołu Jurkowskiego wykorzysty-
wano nanocząstki tlenku żelaza(II) diżelaza(III) w celach poprawiających przeżywalność
bakterii mlekowych Lactobacillus acidophilus w warunkach obniżonego pH [5].
W przeprowadzonych badaniach wartości pH w momencie rozpoczęcia hodowli
(w hodowli wyjściowej) była bliska odczynowi obojętnemu. Wartość pH wynosiła

Wpływ nanocząstek tlenku żelaza na produkcję kwasu mlekowego
w hodowli okresowej Lactobacillus rhamnosus GG
33
wówczas 6,613. W trakcie trwania hodowli we wszystkich próbach obserwowano
obniżenie wartości pH, odczyn stawał się kwasowy. Odczyn pH zależał od warunków
hodowli. Po 24 godzinach średnia wartość pH znacznie obniżyła się w próbach z nano-
cząstkami tlenku żelaza w porównaniu do próby kontrolnej. Dla prób kontrolnych
średnia wartość pH wynosiła 6,085, podczas gdy dla prób z Fe3O4 wartość pH wyno-
siła 5,736. Różnica ta jest istotna statystycznie. Różnice w wartości pH dla kolejnego
pomiaru również były istotne statystycznie i wynosiły 5,559 i 5,542, odpowiednio dla
prób z nanocząstkami i próby kontrolnej.
Analizując wyniki pomiarów stężenia kwasu mlekowego w próbie z nanocząstkami
tlenku żelaza(II) diżelaza(III) oraz w próbie kontrolnej również zauważono różnice
w ilości kwasu mlekowego w obu próbach. Zarówno pomiar po 24 godzinach jak i po
48 i 72 godzinach wskazywał na wyniki wyższe dla prób z nanocząstkami niż dla
próby kontrolnej. Otrzymane w tych badaniach wyniki również okazały się istotne
statystycznie.
Badając liczebność bakterii w hodowlach z nanoczastkami oraz w hodowli kontrol-
nej również zauważono znaczne różnice w obu próbach. Liczebność bakterii w hodow-
lach z nanocząstkami tlenku żelaza(II) diżelaza(III) była mniejsza niż w hodowli
kontrolnej. Największą różnicę w liczebności bakterii zanotowano podczas pomiaru
uzyskanego po 24 godzinach, gdzie liczebność bakterii w hodowli z nanocząstkami
była prawie czterokrotnie mniejsza niż w kontroli. Jednakże znaczną różnicę w liczbie
bakterii zanotowano również po 48 godzinach. Dla wyników analizowanych po 24 i 48
godzinach różnice były istotne statystycznie. Różnice w liczbie bakterii po 72
godzinach nie były istotne statystycznie.
6. Podsumowanie
Obecność nanocząstek tlenku żelaza(II) diżelaza(III) spowodowała w początkowej
fazie hodowli obniżenie zdolności bakterii Lactobacillus rhamnosus GG do podziałów,
co skutkowało mniejszą liczebnością bakterii w próbie badawczej niż w kontroli.
Mimo mniejszej liczebności bakterii (w początkowych godzinach hodowli)
w próbach z nanocząstkami tlenku żelaza(II) diżelaza(III) w rezultacie uzyskano
większe ilości kwasu mlekowego. Obecność nanocząstek tlenku żelaza zwiększa więc
produktywność kwasu mlekowego w hodowli okresowej bakterii probiotycznych
Lactobacillus rhamnosus GG.
Otrzymane wyniki wzrostu uzyskanego produktu fermentacji mlekowej w obec-
ności nanocząstek Fe3O4 są obiecujące. Możliwość zastosowania tego czynnika
w przemysłowej produkcji kwasu mlekowego skutkowałoby korzyściami ekono-
micznymi.
Użycie nanocząstek tlenku żelaza(II) diżelaza(III) zwiększyłoby ilość uzyski-
wanego produktu końcowego reakcji fermentacji mlekowej. Raz dodane do biore-
aktora nanocząstki mogłyby być wykorzystywane stale, ponieważ nie ulegają zużyciu
(pomijając niewielki procent nanocząstek, które mogłyby przenikać przez ściany
komórkowe bakterii mlekowych, i być usuwane z bioreaktora razem z usuwaniem
biomasy bakterii).

Andrzej Jurkowski, Ita Szczepańska, Jacek J. Kozioł
34
Planowane są kolejne badania z użyciem szczepów bakterii mlekowych stoso-
wanych w przemyśle celem sprawdzenia dla szczepów przemysłowych wykazanego
w tej pracy efektu (bowiem każdy szczep może różnie „reagować” na zastosowane
nanocząstki). Jeśli szczepy przemysłowe w hodowlach razem z nanocząstkami tlenku
żelaza(II) diżelaza(III) będą wykazywały większą produkcję kwasu mlekowego niż bez
nanocząstek Fe3O4, istnieje możliwość zastosowania tej metody w przemyśle. Zastoso-
wanie nanocząstek tlenku żelaza(II) diżelaza(III) będzie skutkowało obniżeniem
kosztów produkcji kwasu mlekowego.
Literatura
1. dos Santos Junior V. E., Targino A. G. R., Flores M. A. P., Rodrigez-Diaz J. M., Teixeira J.
A., Heimer M. V., de Luna Freire Pessoa H., Galembeck L., Rosenblatt A. Antimicrobial
activity of silver nanoparticle colloids of different sizes and shapes against Streptococcus
mutants. Research of Chemical Intermediates., (2017), doi:10.1007/s11164-017-2969-5
2. Ocsoy I., Demirbas A., McLamore E. S., Altinsoy B., Ildiz N., Baldemir A. Green
synthesis with incorporated hydrothermal approaches for silver nanoparticles formation
and enhanced antimicrobial activity against bacterial and fungal pathogens. Journal of
Molecular Liquids., 238 (2017), s. 263–269.
3. Erjaee H., Rajaian H., Nazifi S. Synthesis and characterization of novel silver
nanoparticles using Chamaemelum nobile extract for antibacterial application. Advances
in Natural Sciences: Nanoscience and Nanotechnology., 8 (2017) 025004 (9s)
4. Rawani. L.A. Mosquito larvicidal activity of green silver nanoparticle synthesized from
extract of bud of Polianthus tuberose. International Journal of Nanotechnology and
Applications., 11 (2017), s. 17-28.
5. Jurkowski A., Zapotoczny B., Kozioł J. J, Dudek M. R. The effect of Fe3O4 Nanoparticles
on Survival of Probiotic Bacteria Lactobacillus acidophilus PCM2499 at lower pH. Polish
Journal of Microbiology., 64 (2015), 307-310.
6. Arsalani N., Fattahi H., Nazarpoor M. Synthesis and characterization of PVP-
functionalized superparamagnetic Fe3O4 nanoparticles as an MRI contrast agent, Polymer
letters., 4 (2010), s. 329-338.
7. Zapotoczny B., Dudek M. R., Kozioł J. J., Mleczko J. Nanobuffering property of Fe3O4
magnetic nanoparticles in aqueous solution, Physica A. Statistical Mechanics and Its
Applications. 392 (2013), s. 1493-1499.
8. Collins. J. K., Thornton G., Sullivan G. O. Selection of Probiotic Strains for Human
Applications, International Dairy Journal., 8 (1998), s. 487-490.
9. Libudzisz Z., Kowal K., Żakowska Z. Mikrobiologia techniczna: Mikroorganizmy
w biotechnologii, ochronie środowiska i produkcji żywności. Wydawnictwo Naukowe
PWN., 2008.
10. Bednarski W. Biotechnologia żywności, Wydawnictwo Naukowo-Techniczne., 2015.
11. Jem K. J., Pol J. F., van der Vos S., Microbial Lactic Acid, Its Polymer Poly(lactic acid),
and Their Industrial Applications, W: Chen G. G-Q. (red.), Plastics from Bacteria,
Microbiology Monographs. Springer Berlin Heidelberg, 2010, s. 323-346.

Wpływ nanocząstek tlenku żelaza na produkcję kwasu mlekowego
w hodowli okresowej Lactobacillus rhamnosus GG
35
Wpływ nanocząstek tlenku żelaza na produkcję kwasu mlekowego w hodowli
okresowej Lactobacillus rhamnosus GG
Streszczenie Wykorzystanie nanocząstek metali i ich tlenków w obszarze mikrobiologii najczęściej koncentruje się
wokół ich zastosowań bakteriobójczych i bakteriostatycznych. Nowością przedstawianych badań jest
zastosowanie nanocząstek w celu poprawy przeżywalności i produktywności bakterii. Pierwsze badania
wykonane przez Jurkowskiego i in. (2015) wykazały wzrost przeżywalności bakterii probiotycznych Lactobacillus acidophilus w warunkach obniżonego pH, dzięki zastosowaniu nanocząstek Fe3O4. Nano-
czastki Fe3O4 charakteryzują się właściwościami magnetycznymi oraz wykazują właściwości nano-
buforujące, są uważane za nieszkodliwe dla organizmu ludzkiego.
Prezentowane wyniki są efektem kontynuacji badań nad wpływem nanocząstek tlenku żelaza na przeżywalność bakterii probiotycznych Lactobacillus rhamnosus GG. Wykonano hodowle okresowe tej
bakterii mierząc w określonych odstępach czasu liczebność bakterii (jtk/ml), zmiany wartości pH oraz
ilości kwasu mlekowego w bulionie hodowlanym. W początkowych etapach hodowli zaobserwowano
wolniejszy wzrost liczebności bakterii w obecności nanocząstek (w porównaniu z kontrolą) oraz stwierdzono większą ich produktywność. Planowane są kolejne badania wykorzystujące szczepy bakterii
mlekowych wykorzystywanych w przemyśle do produkcji kwasu mlekowego.
Słowa kluczowe: bakterie mlekowe, kwas mlekowy, Lactobacillus rhamnosus GG, nanocząski, Fe3O4
Effect of iron oxide nanoparticles on lactic acid production in the bath culture
of Bactobacillus rhamnosus GG
Abstract
The use of nanoparticles of metals and their oxides in microbiology ist most often centered around their
bactericidal and bacteriostatic applications. The novelty of the presented research is the use of nanoparticles to improve the survival and productivity of bacteria. The first study by Jurkowski et al.
(2015) showed an increase survival of the probiotic bacteria Lactobacillus acidophilus in lower pH by the
use of Fe3O4 nanoparticles in medium. Fe3O4 nanoparticles are characterized by magnetic properties and
have nanobufforing properties, that are considered harmless to the human body. These results are the effect of a follow-up study on the impact of oxide nanoparticles on the viability of the probiotic bacteria
Lactobacillus rhamnosus GG.
Periodic cultures of this bacteria were measured at specified intervals, the number of bacteria (CFU/ml),
changes in pH and amount of lactic acid. In the early stages of the culture, slower growth of bacterial counts were observed in the presence of nanoparticles (as compared to controls) and their productivity
increased. Further studies are planned with lactic acid bacteria strains used in the lactic acid industry.
Keywords: lactic acid bacteria, lactic acid, Lactobacillus rhamnosus GG, nanoparticles, Fe3O4

36
Rafał Kuś1, Monika Hałgas
1, Gary van Lue
1, Francisco Gonzalez Vizcarra
1,
Małgorzata M. Kozioł2, Alina Olender
2
Występowanie Staphylococcus aureus w jamie nosowo-
gardłowej a czynniki ryzyka wpływające na kolonizację
1. Wstęp
W mikrobiologii nazwa Staphylococcus wywodzi się od obrazu oglądanego
w mikroskopie świetlnym, w preparacie barwionym metodą Grama, gdy komórki bak-
terii układają się w charakterystyczne skupiska – grona, przypominając kiść winogron.
Organizmy te zostały po raz pierwszy opisane i sklasyfikowane w 1882 roku przez
szkockiego chirurga Sir Alexander Ogston, a dwa lata później niemiecki lekarz Frie-
drich J. Rosenbach, opisał 2 typy kolonii gronkowców i zaproponował nomenklaturę
Staphylococcus albus (łac. „biały”) i Staphylococcus aureus (z łacińskiego aurum
„złoto”). Od tego czasu S. aureus nazywany gronkowcem złocistym niepokoi naukow-
ców i lekarzy wywołując zakażenia o różnej ciężkości u milionów osób [1].
Gronkowiec złocisty obok innych gatunków rodzaju Staphylococcus, które wcho-
dzą w skład naturalnej mikroflory człowieka, może kolonizować nasz organizm i za-
siedlać bardzo specyficzne dla siebie nisze, jak jama nosowo-gardłowa. Będąc najbar-
dziej zjadliwym gatunkiem rodzaju, powodującym choroby u ludzi, począwszy od
łagodnych zakażeń skóry po ciężkie i zagrażające życiu zakażenia, takie jak zapalenie
wsierdzia, zapalenie płuc i sepsę. Jeśli bytuje w organizmie człowieka nie wywołując
objawów choroby mówi się o tak zwanym stanie nosicielstwa [2]. Nosicielstwo S.
aureus jest poważnym problemem zdrowotnym szczególnie u osób poddawanych
różnym procedurom inwazyjnym w środowisku szpitalnym. Jeśli dochodzi do infekcji
u pacjentów hospitalizowanych bardzo często zakażenia wywołują szczepy oporne na
metycylinę – MRSA (ang. methicillin-resistant Staphylococcus aureus), choć ostatnio
coraz częściej obserwuje się szczepy metycylinooporne jako czynnik etiologiczny
infekcji pozaszpitalnych. To właśnie z rozwojem oporności, zwłaszcza na wspomnianą
metycylinę, a także inne antybiotyki β-laktamowe (z wyjątkiem ceftaroliny i cefto-
biprolu) wiąże się narastający problem zakażeń gronkowcem złocistym, związany
z ograniczonymi możliwościami terapeutycznymi. Do 2003 roku infekcje szczepami
MRSA były stosunkowo rzadkie u ludzi zdrowych w środowisku pozaszpitalnym.
Później pojawiły się doniesienia o nowych szczepach MRSA odpowiedzialnych za
ogniska epidemiczne w populacjach pozaszpitalnych i wywołujących infekcje skóry
oraz zapalenia płuc [3].
1Studenckie Koło Naukowe przy Katedrze i Zakładzie Mikrobiologii Lekarskiej, Uniwersytet Medyczny
w Lublinie, [email protected] 2Katedra i Zakład Mikrobiologii Lekarskiej, Uniwersytet Medyczny w Lublinie,

Występowanie Staphylococcus aureus w jamie nosowo-gardłowej
a czynniki ryzyka wpływające na kolonizację
37
Gronkowiec złocisty przejściowo może kolonizować wilgotne zagłębienia w skórze.
Powszechna jest kolonizacja kikuta pępkowego u noworodków, okolicy krocza, a także
przewodu pokarmowego, dróg moczowych i narządów płciowych oraz przedniej części
gardła. Krótkotrwałe lub stałe nosicielstwo S. aureus u starszych dzieci i dorosłych jest
częstsze w przedniej części nosogardła niż w części ustnej gardła [2, 4]. Około 30%
zdrowych dorosłych osób jest nosicielami S. aureus w nosogardzieli, przy czym
występowanie gronkowca złocistego w tym rejonie ciała jest wyższe u hospitalizo-
wanych pacjentów, personelu medycznego, osób z wypryskowymi chorobami skóry oraz
osób regularnie używających iniekcji. W związku z tym, że gronkowce występują na
skórze i w nosogardzieli, transmisja tych bakterii jest częsta i jest przyczyną wielu
infekcji szpitalnych oraz pozaszpitalnych. Bakterie choć wrażliwe na wysokie
temperatury oraz środki dezynfekcyjne są w stanie przetrwać na suchych powierzchniach
przez dłuższy czas, co stwarza możliwości transmisji. Gronkowiec złocisty może zostać
przekazany drugiej osobie poprzez bezpośredni kontakt albo przez kontakt z przed-
miotami zanieczyszczonymi bakteriami. Dlatego też personel medyczny i przyszły
personel medyczny jako ten najbardziej narażony na stan nosicielstwa i transmisję
bakterii powinien w szczególny sposób, używając odpowiednich technik mycia rąk,
zapobiegać transferowi gronkowców w miejscu pracy i jednocześnie zminimalizować
czynniki ryzyka, które mogą wpłynąć na potencjalną kolonizację, a tym samym na stan
nosicielstwa [5].
2. Cel
Celem niniejszych badań była ocena stanu nosicielstwa Staphylococcus aureus
w jamie nosowo-gardłowej i ocena zależności nosicielstwa od czynników ryzyka
potencjalnie wpływających na kolonizację, w grupie studentów medycyny Uniwer-
sytetu Medycznego w Lublinie. Przeprowadzone badania miały charakter badań
wstępnych.
3. Materiały i metody
Grupę badaną stanowiło 46 studentów medycyny Uniwersytetu Medycznego
w Lublinie. Pochodzili oni z różnych rejonów: Stanów Zjednoczonych, Tajwanu,
Wielkiej Brytanii, Arabii Saudyjskiej, Tajlandii, Nowej Zelandii, Japonii oraz Brazylii.
W badanej grupie przeważali mężczyźni – 60% studentów, a kobiety stanowiły 40%.
Średnia wieku w grupie wynosiła 28 lat (20-51 lat). Wśród tych osób 65% to studenci
II roku, 29% – IV roku, 4% – V roku i 2% – I roku. Wszyscy studenci podpisali for-
mularz świadomej zgody na udział w badaniu. Na wykonanie badań uzyskano zgodę
Komisji Bioetycznej UM w Lublinie.
Materiałem klinicznym do badań były wymazy z gardła i obustronnie przedsionka
nosa. Wymazy pobrano z zachowaniem zasad aseptyki jałowymi wymazówkami
zwilżonymi 0,9% NaCl. Po pobraniu próbki poddano następującemu tokowi diagno-
stycznemu: posiew na odpowiednie podłoża mikrobiologiczne metodą redukcyjną
(podłoże Columbia Agar z 5% odwłóknioną krwią baranią, podłoże Chapmana),
inkubacja w temperaturze 37°C przez 24-48 godzin w warunkach tlenowych; charak-

Rafał Kuś, Monika Hałgas, Gary van Lue, Francisco Gonzalez Vizcarra,
Małgorzata M. Kozioł, Alina Olender
38
terystyka wzrostu wyrosłych bakterii; identyfikacja biochemiczna (test na katalazę,
próba na koagulazę związaną) i oznaczenie lekowrażliwości gronkowców na anty-
biotyki β-laktamowe (metoda dyfuzyjno-krążkowa na podłożu Mueller-Hinntona
z użyciem krążków z cefoksytyną 30 μg).
W celu analizy czynników ryzyka stanu nosicielstwa gronkowca złocistego wśród
przyszłego personelu medycznego badani poproszeni zostali o wypełnienie przygo-
towanej autorskiej ankiety.
4. Wyniki
Wyniki badań mikrobiologicznych w badanej grupie wykazały obecność S. aureus
w jamie nosowo-gardłowej u 27 osób, co stanowi 58,7% badanych studentów.
U pozostałych 19 osób (41,3%) nie stwierdzono wzrostu patogenu. Pozwoliło to
podzielić grupę badaną na tak zwaną grupę nosicieli – GN i grupę nienosicieli – GNN.
Wśród GN przeważali mężczyźni (70% vs 30%). Rozpatrując grupę samych męż-
czyzn, wykazano że nosiciele gronkowca to 68% mężczyzn, natomiast wśród grupy
kobiet 45% to kobiety z nosicielstwem.
Badanie lekowrażliwości izolowanych szczepów S. aureus wykazało, że w 85%
były to gronkowce metycylinowrażliwe MSSA (ang. methicillin-sensitive Staphylo-
coccus aureus ), natomiast 15% stanowiły szczepy metycylinooporne (MRSA). U 78%
GN gronkowca złocistego drobnoustrój izolowano z gardła, u 37% wykryto obecność
S. aureus jednocześnie w gardle oraz w przedsionku nosa, u 3 osób były to różne
szczepy gronkowca złocistego o tym samym wzorze oporności na metycylinę, a u 1
różne szczepy, o różnym wzorze oporności na metycylinę (MSSA i MRSA).
Analiza ankietowa
W przeprowadzonej ankiecie pytano, o dane demograficzne, wiek, płeć, masę,
przebyte w przeszłości infekcje (głównie dróg oddechowych), hospitalizacje, zabiegi
operacyjne, kontakt z dziećmi, zwierzętami, stosowanie antybiotyków, palenie tytoniu,
stosowanie środków czystości, aseptykę i antyseptykę, szczepienie przeciwko grypie,
czy choroby współistniejące (cukrzyca, choroby nerek).
Wyniki przeprowadzonej ankiety przedstawiono poniżej.
W GN, aż 97% badanych zadeklarowało, że myje ręce ≥4 razy w ciągu doby,
a żelu antyseptycznego do rąk używa 19%. Wśród GNN 95% myje ręce ≥4
w ciągu doby, a 32% używa żelu antyseptycznego,
Spośród GN 15% czyści ekran urządzeń elektronicznych, dopiero kiedy zauważy
brud lub wcale tego nie robi, natomiast w GNN jest to 37%,
11% spośród całej grupy badanej jest właścicielem zwierzęcia domowego,
z czego 7% to GN, a 16% GNN.
Kontakt z dziećmi w wieku przedszkolnym był znikomy w obu grupach i nie miał
wpływu na wyniki badań.
W momencie pobierania wymazów 1 osoba nie będąca nosicielem miała stan
podgorączkowy, a 11% GN oraz 11% GNN stosowało doustną antybio-
tykoterapię.

Występowanie Staphylococcus aureus w jamie nosowo-gardłowej
a czynniki ryzyka wpływające na kolonizację
39
Wśród całej grupy badanej 2 osoby przebyły w swoim życiu paciorkowcowe
zapalenie gardła, a 1 osoba (GNN) została poddana zabiegowi usunięcia mig-
dałków podniebiennych.
Na zapalenie oskrzeli lub płuc chorowało GN 11% vs GNN 26%.
Infekcje górnych dróg oddechowych 1-2 razy dominują w obydwu grupach GN
93% vs GNN 84%. Częstsze infekcje, czyli ≥ 3 razy w ciągu roku GN 7% vs GNN
16%. Najczęstszym okresem zapadalności był okres zimowy. Ciężkie zakażenie
dróg oddechowych, przebyły tylko osoby z GN 7%. Stosowanie antybio-
tykoterapii w leczeniu infekcji górnych dróg oddechowych było rzadkie
w obydwu grupach,
Rozpatrując szczepienia przeciwko grypie, odsetek profilaktyki był porów-
nywalny i na średnim poziomie (GN 30% vs GNN 26%).
W całej grupie badanej hospitalizowanych było 46% wszystkich studentów.
W GN odsetek hospitalizowanych wynosił 41% a GNN 53%.
Ponad połowa hospitalizowanych nosicieli przeszła zabieg operacyjny, natomiast
nieco większy odsetek GNN. Żadna osoba z grupy badanej nie zauważyła u siebie
zmian skórnych, które mogłyby świadczyć o zakażeniu gronkowcowym po
zakończonym pobycie w oddziale szpitalnym.
Palenie tytoniu zadeklarował niewielki odsetek osób GN 7% vs GNN 11%, co
stanowi 9% całej grupy badanej.
BMI badanych było prawidłowe lub wskazywało na nadwagę. Nie odnotowano
współistnienia chorób towarzyszących. Żaden z respondentów nie zadeklarował
zakażenia wirusem HIV.
5. Dyskusja
Nosicielstwo S. aureus jest dość powszechnym i szeroko dyskutowanym w piśmie-
nictwie problem, przede wszystkim ze względu na możliwe powikłania infekcyjne dla
nosiciela i osób z jego otoczenia. Jak wspomniano bakteria może być czynnikiem
etiologicznym różnorakich zakażeń: skóry, tkanek miękkich, przyczyną inwazyjnych
infekcji w układzie krążenia czy w ośrodkowym układzie nerwowym oraz przyczyną
zakażeń szpitalnych. Najważniejszym miejscem jego bytowania jest jama nosowo-
gardłowa, pomimo że kolonizuje wiele rejonów naszego organizmu. Związane jest to
ze zdolnością adhezji bakterii do nabłonka śluzowego – glikoprotein pomierzch-
niowych i proteoglikanów błon śluzowych. Dodatkowo węglowodany mucyny
gospodarza i inne substancje reagują z białkami bakterii stanowiąc swoiste miejsca
receptorowe [4].
Badania statystyczne wskazują na stan nosicielstwa w społeczeństwie w zakresie
20-40%. Szczególną grupą ryzyka jest natomiast personel medyczny, jak wynika
z danych epidemiologicznych, odsetek nosicieli jest tu jeszcze wyższy [4, 6]. W wielu
publikacjach poddaje się dyskusji wskaźnik nosicielstwa S. aureus wśród studentów
medycyny. Autorzy niejednokrotnie zastanawiają się czy przypadkiem nie wzrasta on
w momencie rozpoczęcia zajęć klinicznych w oddziałach medycznych. Sugerowałoby
to, że osoby takie zostają szybko skolonizowanie i stają się potencjalnym źródłem

Rafał Kuś, Monika Hałgas, Gary van Lue, Francisco Gonzalez Vizcarra,
Małgorzata M. Kozioł, Alina Olender
40
rozprzestrzeniania bakterii. Badacze nie są zgodni co do tej kwestii. Badania między
innymi Ciftic i wsp. wskazują, że powyższa teoria nie ma wpływu na wzrost liczby
nosicieli [6]. Inni jak Subri i wsp. także to potwierdzają, a nawet wskazują, że w grupie
studentów nie mających zajęć klinicznych wskaźnik nosicielstwa był wyższy [2].
Podobnie w badaniach własnych również nie zaobserwowano takiej istotności. Choć
grupa była mała to należy podkreślić, że większość osób była na wczesnym etapie
edukacji medycznej a odsetek nosicieli (59%) był wysoki, nieco wyższy od tego
obserwowanego w grupie pracowników opieki medycznej. W innych studenckich
grupach stan nosicielstwa był na różnym poziomie – 39,3%, lub jak u tureckich
studentów medycyny nawet o połowę mniejszy (27,9%) [7, 8].
Obecnie kwestią kluczową pozostaje wrażliwość gronkowców na antybiotyki β-
laktamowe. Co istotne, w badaniach własnych, izolowane szczepy były w większości
metycylinowrażliwe (85%), co dobrze rokuje i odzwierciedla stan nosicielstwa
w społeczeństwie. Niemniej jednak, choć niewielka liczba była nosicielem MRSA, to
powinno dążyć się by w obrębie grupy stosunek liczby szczepów MRSA vs MSSA był
niższy. Należy podkreślić, że zakażenia MRSA pozostają swoistego rodzaju terapeu-
tycznym wyzwaniem. Pomimo wprowadzenia w ciągu ostatniego dziesięciolecia kilku
nowych środków przeciwbakteryjnych jest to jeden z trudnych do usunięcia
patogenów, zaliczanych do tak zwanej grupy ESKAPE (Enterococcus faecium, S.
aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa
i Enterobacter). Zmienność epidemiologii zakażeń MRSA, różna oporność na
powszechnie stosowane antybiotyki oraz zaangażowanie MRSA w zakażenia związane
ze szpitalem wpływają na stosowanie i wyniki kliniczne obecnie dostępnych środków
przeciwbakteryjnych. Obecnie opracowywana jest znaczna liczba badanych środków
mających aktywność przeciwko MRSA [9].
Wymienia się wiele czynników ryzyka nabycia gronkowca złocistego, poczynając
od predyspozycji związanej z płcią, przez młody wiek, kontakt z pracownikami opieki
medycznej, wkłucia dożylne, aż po astmę [10]. W badaniach własnych, w analizo-
wanej grupie wykazano, że częściej kolonizacji ulegają mężczyźni. W grupie nosicieli
stosunek ten wynosił 70% vs 30%. Biorąc pod uwagę samych mężczyzn, aż 68% to
właśnie nosiciele, gdzie w porównaniu do kobiet – skolonizowanych było 45%, zatem
jest to znacząca różnica w grupach. Choć badania miały charakter badań wstępnych
i należałoby powiększyć grupę, to jednak potwierdzają one inne dane, z których
wynika, że płeć męska faktycznie dominuje wśród osób skolonizowanych [11].
Wśród czynników potencjalnie mogących wpływać na stan nosicielstwa jest stoso-
wana wcześniej antybiotykoterapia. W szeroko zakrojonych badaniach Yanga i wsp.
przeanalizowano szereg czynników ryzyka, w tym właśnie stosowanie antybiotyków.
Jak wykazano, zarówno w grupie nosicieli MRSA i nieskolonizowanych odsetek osób,
które zażywały antybiotyki był identyczny i zarazem wysoki (42%), nie miało to
jednak wpływu na stan nosicielstwa. Również dożylne przyjmowanie leków było na
porównywalnym poziomie [11]. Podobnie w badaniach własnych nie wykazano
związku ze stosowaniem leków przeciwbakteryjnych, ale osoby w badanej grupie

Występowanie Staphylococcus aureus w jamie nosowo-gardłowej
a czynniki ryzyka wpływające na kolonizację
41
bardzo rzadko lub w ogóle ich nie stosowały, nawet jeśli dochodziło do infekcji
górnych dróg oddechowych.
Większość zakażeń górnych dróg oddechowych (GDO) zaczyna się jako zakażenie
wirusowe, stąd też nie ma potrzeby stosowania antybiotyków, niemniej jednak przy
długotrwałej infekcji dochodzi do powikłań bakteryjnych. Infekcje wirusowe zaburzają
funkcję aparatu rzęskowego, powodują martwicę i złuszczenie komórek nabłonka
urzęsionego czym ułatwiają przyleganie bakterii do błon śluzowych [4]. Jeśli spoj-
rzymy na uzyskane wyniki częstość infekcji była mało zróżnicowana, najczęściej
dochodziło do zakażenia 1-2 razy w roku. Natomiast ≥ 3 razy w ciągu roku na
zakażenie GDO zapadała mała grupa – dwa razy częściej GNN, jednocześnie ciężkie
infekcje pojawiły się tylko w GN.
Nosicielstwo gronkowca to ważny problem dla zdrowia publicznego, powiązywany
wcześniej jedynie z opieką zdrowotną, ograniczany przede wszystkim do oddziałów
intensywnej opieki, a także innych placówek opieki medycznej. Kluczową rolę
w kolonizacji odgrywać powinny zatem przebyte hospitalizacje, operacje oraz długo-
terminowe pobyty w szpitalu (30,3%). Hospitalizacja niewątpliwie może wpływać na
stan nosicielstwa, wszystko zależne jest od polityki antybiotykowej szpitala, ilości
stosowanych leków przeciwbakteryjnych, ponieważ przekłada się to na obecność
szczepów wielolekoopornych w środowisku szpitalnym. Z danych ze Stanów Zjedno-
czonych wynika, że 76,6% nosicieli było kiedykolwiek hospitalizowanych
w porównaniu do 57,7% hospitalizowanych nienosicieli [12]. W badaniach Yanga
i wsp. również częściej nosiciele przebywali w szpitalu (29% vs 17%), choć ryzyko
kolonizacji uplasowało się już na niższym poziomie [11]. W badaniach własnych
hospitalizacja nie miała związku z nabyciem patogenu, a wręcz przeciwnie osoby
z GNN częściej przebywały w szpitalu w celach leczniczo-diagnostycznych (GN 41%
vs GNN 53%). One też częściej były poddawane zabiegom w trybie elektywnym.
Należy przy tym pamiętać, że przed planowaną procedurą inwazyjną pacjenci mogą
profilaktycznie mieć podaną donosowo maść z mupirocyną, którą stosuje się
w procesie eradykacji S. aureus [4]. Operacje niekoniecznie muszą mieć jakikolwiek
wpływ na stan nosicielstwa, na co wskazują autorzy jednej z prac (37% vs 37,6%), ale
już samo przebywanie w szpitalu jako „pacjent” ma newralgiczne znaczenie.
Dodatkowo długoterminowa hospitalizacja niemal dwukrotnie zwiększa ryzyko
kolonizacji florą szpitalną i nabycia drobnoustroju [12].
Co ciekawe, Yang i wsp. nie wykazali związku między kolonizacją szczepami
MRSA, a kontaktem z osobami mającymi różnego rodzaju infekcje skórne. Osoby
nieskolonizowane miały wręcz znacząco częstszy kontakt z osobą będącą potencjal-
nym źródłem patogenu (4% vs 17%). Dopiero jednak przebyta infekcja gronkowcowa
(do 12 miesięcy wcześniej) czy stan nosicielstwa wirusa HIV wpływał w badanej
grupie na kolonizację jamy nosowo-gardłowej i innych obszarów ciała. Stan nosi-
cielstwa po przebytej infekcji określamy jako nosicielstwo ozdrowieńców, więc
wysoce prawdopodobne było osiągnięcie stanu równowagi patogenu w organizmie
chorego po przebyciu zakażenia [11].

Rafał Kuś, Monika Hałgas, Gary van Lue, Francisco Gonzalez Vizcarra,
Małgorzata M. Kozioł, Alina Olender
42
W środowisku szpitalnym dekolonizacja skóry może być osiągnięta przy
zastosowaniu odpowiednich mydeł. I to właśnie w restrykcyjnym przestrzeganiu zasad
aseptyki i antyseptyki w środowisku szpitalnym oraz w higienie rąk i stosowaniu
różnego rodzaju środków antyseptycznych na co dzień upatruje się związku z niskim
odsetkiem nosicielstwa u studentów [13]. Badania własne pokazują, że częstość mycia
rąk pomiędzy grupami jest taka sama i jedynie w stosowaniu dodatkowych środków
antyseptycznych do rąk w ciągu dnia można upatrywać się jakiejś korelacji (GN 19%
vs GNN 32%).Paradoksalnie, grupa nienosicieli w mniejszym stopniu dbała o czystość
swoich urządzeń elektronicznych. Jak pokazują badacze z różnych ośrodków, jeśli
występuje niski odsetek nosicielstwa w grupie studentów medycyny to właśnie
wysokie standardy higieny i przestrzeganie protokołu antyseptyki w placówkach
służby zdrowia odgrywają w tym najważniejszą rolę.
Warto dodać, że w piśmiennictwie przy stanie nosicielstwa ocenie poddaje się
również czynniki takie jak choroby współistniejące – cukrzyca, przewlekła choroba
nerek oraz nosicielstwo wirusa HIV, otyłość i palenie tytoniu. Z danych van Belkum
i wsp. wynika, że takie jak wiek, cukrzyca, palenie były niezależnymi determinantami
[14]. Podobnie praca własna wykluczyła jakoby palenie tytoniu miało związek ze
stanem nosicielstwa, a także żadna z powyższych determinant.
6. Wnioski
Otrzymane wyniki badań wśród przyszłego personelu medycznego, potwierdzają
wysoki odsetek nosicielstwa S. aureus jaki obserwuje się u pracujących w zawodach
medycznych. Pomimo znaczącej przewagi nosicielstwa szczepami MSSA, obecność
nosicieli MRSA, a więc tych szczepów które stwarzają największe ryzyko zakażeń,
jest zbyt wysoka. Eliminacja patogenu jest wówczas bardzo ważnym działaniem
profilaktycznym. Analizując czynniki ryzyka wpływające na stan nosicielstwa,
większość z nich okazała się niezależnymi determinantami. Niemniej jednak potrzebne
są dalsze badania na większej grupie, w celu potwierdzenia wyników badań wstępnych
i pogłębionej analizy zależności między czynnikami ryzyka a kolonizacją gronkowcem
złocistym.
Literatura
1. Stryjewski M.E., Corey G.R.: Methicillin-Resistant Staphylococcus aureus: An Evolving
Pathogen. Clin Infect Dis; 58, 1 (2014), s. 10-19.
2. Subri N.I.B.M., Hlaing S.S., Myint T., Emran N.A., Lin Z., Thein T.T., Masandid H., Aung
T.S.: Nasal Carriage of Staphylococcus aureus and Its Antibiotic Susceptibility Pattern
among Medical and Nursing Students. AJP; 10, 4 (2016), s. 736-740.
3. Paterson G.K., Harrison E.M., Holmes M.A.: The emergence of mecC methicillin-resistant
Staphylococcus aureus. Trends Microbiol.; 22, 1 (2014), s. 42-47.
4. Kozioł M.M., Sikora A., Targońska S., Sikora A.: Nosicielstwo Staphylococcus aureus jako
czynnik ryzyka zakażeń szpitalnych. Forum Zakażeń; 5, 4 (2014), s. 205-209.
5. Wertheim H.F., Melles D.C., Vos M.C., van Leeuwen W., van Belkum A., Verbrugh H.A.,
Nouwen J.L.: The role of nasalcarriage in Staphylococcusaureusinfections. Lancet Infect
Dis; 5 (2005), s. 751–762

Występowanie Staphylococcus aureus w jamie nosowo-gardłowej
a czynniki ryzyka wpływające na kolonizację
43
6. Ciftci Z., Deniz M., Gunes H., Gumus A., Gultekin E., Topkaya A.: Does Attending
Clinical Wards Increase Nasal Carriage of Staphylococcus Aureus Among Medical
Students? Medical Science and Discovery; 2, 5 (2015); s. 288-291.
7. Güçlü E., Yavuz T., Tokmak A., Behçet M., Karali E., Oztürk O., Egeli E. Nasal carriage
of pathogenic bacteria in medical students: effects of clinic exposure on prevalence and
antibiotic susceptibility. Eur Arch Otorhinolaryngol.; 264 (2007), s. 85-88.
8. López-Aguilera S., Goñi-Yeste Mdel M., Barrado L., González-Rodríguez-Salinas M.C.,
Otero J.R., Chaves F. Staphylococcus aureus nasal colonization in medical students:
imprtance in nosocomial transmission. Enferm Infecc Microbiol Clin.; 31 (2013); s. 500-505.
9. Rodvold K.A., McConeghy K.W.: Methicillin-resistant Staphylococcus aureus therapy:
past, present, and future.Clin Infect Dis.; 58, 1 (2014), s. 20-27.
10. Halablab M.A., Hijazi S.M., Fawzi M.A., Araj G.F.: Staphylococcus
aureus nasalcarriagerate and associated risk factors in individuals in the community.; 138, 5
(2010), s. 702-706.
11. Yang E.S., Tan J., Eells S., Rieg G., Tagudar G., Miller L.G.: Body site colonization in
patients with community-associated methicillin-resistant Staphylococcus aureus and other
types of S. aureus skin infections. Clin Microbiol Infect.; 16, 5 (2010), s. 425-431.
12. Klevens R.M., Morrison M.A., Nadle J., Petit S., Gershman K., Ray S., Harrison
L.H., Lynfield R., Dumyati G., Townes J.M., Craig A.S., Zell E.R., Fosheim
G.E., McDougal L.K., Carey R.B., Fridkin S.K.; Active Bacterial Core surveillance (ABCs)
MRSA Investigators: Invasive methicillin-resistant Staphylococcus aureus infections in the
United States. JAMA.; 298, 15 (2007), s. 1763-1771.
13. Bode L.G.M, Kluytmans A.J.W., Wertheim HFL., Bogaers D., Vandenbroucke-Grauls
C.M.J.E., Roosendaal R., Troelstra a., Box A.T.A., Voss A., van der Tweel I., van Belkum
A., Verbrugh HA.,. Vos MC.: Preventing Surgical-Site Infections in Nasal Carriers
of Staphylococcus aureus. N Engl J Med; 362 (2010), s. 9-17.
14. van Belkum A., Verkaik N.J., de Vogel C.P., Boelens H.A., Verveer J., Nouwen
JL., Verbrugh HA., Wertheim HF.: Reclassification of Staphylococcus aureus nasal
carriage types. J Infect Dis.; 15,199,12 (2009), s. 1820-1826.
Występowanie Staphylococcus aureus w jamie nosowo-gardłowej
a czynniki ryzyka wpływające na kolonizację
Streszczenie
Staphylococcus aureus jest zarówno drobnoustrojem komensalnym, jak i bezwzględnym patogenem
chorobotwórczym będącym częstą przyczyną zakażeń klinicznych o różnej lokalizacji i ciężkości prze-biegu. Uważa się, że jeśli bytuje w organizmie to najczęściej w jamie nosowo-gardłowej. Tam namnaża się
i jest źródłem przeniesienia bakterii na osoby z otoczenia nosiciela, co jest szczególnie ważne w środo-
wisku szpitalnym. Obecnie dane szacują, że nosicielstwo u zdrowych dorosłych sięga 20-40%, a znacząco
wzrasta u personelu medycznego. Wiele czynników może wpływać na stan nosicielstwa takich jak płeć, wiek, otyłość, hospitalizacja, przebyte operacje, choroby skóry, stosowanie środków antyseptycznych, palenie,
przebyte infekcje górnych dróg oddechowych czy choroby współistniejące (cukrzyca, choroby nerek).
Cel:. Celem pracy była ocena stanu nosicielstwa Staphylococcus aureus w jamie nosowo-gardłowej
i ocena zależności nosicielstwa od czynników ryzyka, potencjalnie wpływających na kolonizację, w grupie studentów medycyny Uniwersytetu Medycznego w Lublinie. Przeprowadzone badania miały charakter
badań wstępnych.
Materiały i metody: Grupę badaną stanowiło 46 osób, z którymi przeprowadzono wywiad w celu zbadania
potencjalnych czynników ryzyka mających wpływ na kolonizację S. aureus. Pobrano od nich materiał kliniczny do badań mikrobiologicznych: wymazy z nosa i gardła. Uzyskany materiał posiano na odpo-
https://www.ncbi.nlm.nih.gov/pubmed/?term=Klevens%20RM%5BAuthor%5D&cauthor=true&cauthor_uid=17940231
https://www.ncbi.nlm.nih.gov/pubmed/?term=Gershman%20K%5BAuthor%5D&cauthor=true&cauthor_uid=17940231
https://www.ncbi.nlm.nih.gov/pubmed/?term=Lynfield%20R%5BAuthor%5D&cauthor=true&cauthor_uid=17940231
https://www.ncbi.nlm.nih.gov/pubmed/?term=Fosheim%20GE%5BAuthor%5D&cauthor=true&cauthor_uid=17940231
https://www.ncbi.nlm.nih.gov/pubmed/?term=Fosheim%20GE%5BAuthor%5D&cauthor=true&cauthor_uid=17940231
https://www.ncbi.nlm.nih.gov/pubmed/?term=Fridkin%20SK%5BAuthor%5D&cauthor=true&cauthor_uid=17940231
https://www.ncbi.nlm.nih.gov/pubmed/?term=Verkaik%20NJ%5BAuthor%5D&cauthor=true&cauthor_uid=19419332

Rafał Kuś, Monika Hałgas, Gary van Lue, Francisco Gonzalez Vizcarra,
Małgorzata M. Kozioł, Alina Olender
44
wiednie podłoża mikrobiologiczne i przeprowadzono diagnostykę z oznaczeniem mechanizmu leko-
wrażliwości.
Wyniki: Nosicielstwo S. aureus rozpoznano u 27 badanych (58,7%), z czego u 4 osób zdiagnozowano szczepy metycylinooporne – MRSA. Gronkowiec u większości izolowano z gardła. U 3 osób rozpoznano
nosicielstwo różnych szczepów w gardle i nosie. W większości analiza wybranych czynników ryzyka nie
wykazała istotnych różnic pomiędzy grupą nosicieli i nie nosicieli.
Wnioski: Otrzymane wyniki badań wśród przyszłego personelu medycznego, potwierdzają wysoki odsetek nosicielstwa S. aureus jaki obserwuje się u pracujących w zawodach medycznych. W większości nosi-
cielstwo dotyczyło szczepów MSSA. Obecność szczepów MRSA można uznać za stosunkowo wysoką, co
stwarza ryzyko inwazyjnych zakażeń, a eliminacja patogenu jest wówczas bardzo ważnym działaniem
profilaktycznym. Analizując czynniki ryzyka wpływające na stan nosicielstwa, większość z nich okazała się niezależnymi determinantami. Niemniej jednak badania przeprowadzono na małej grupie i potrzebne
jest ich kontynuacja w celu weryfikacji wyników wstępnych.
Słowa kluczowe: Staphylococcus aureus, nosicielstwo, metycylinoporność
Nosopharyngeal carriage of Staphylococcus aureus and risk factors influencing
colonization Abstract
Staphylococcus aureus is a commensal microorganism and an absolute pathogen that is a common cause of
clinical infections of varied locations and severity. It is believed that if found in human body, it is usually in the nasopharynx. There it grows and is a source of transmission of bacteria to people from the environment of
the carrier, which is particularly important in the hospital environment. Currently, the data estimates that
carriers in healthy adults reach 20-40% and increase significantly among medical staff. Many factors like
gender, age, obesity, hospitalization, undergone surgeries, skin infections, usage of antiseptics, smoking, upper respiratory tract infections and coexisting diseases (diabetes mellitus, kidney diseases) can influence carrier
state.
Aim: The aim of the study was to assess the status of Staphylococcus aureus carriage in nasopharynx and to
asses dependency of carrier state from risk factors potentially influencing colonization among a group of medical students of Medical University of Lublin.
Materials and Methods: The study group consisted of 46 people with whom the interview was conducted to
investigate potential risk factors that can influence colonization of S. aureus. The nasal and throat swabs were
collected to carry microbiological tests. The obtained clinical material was sown on appropriate microbial media and diagnostics were performed with the designation of the drug susceptibility mechanism.
Results: Carriers of S. aureus was identified in 27 people (58.7%), 4 of whom were diagnosed with methicillin
resistant MRSA. Staphylococcus was mostly isolated from the throat. Three people were diagnosed with
various strain of bacteria in the throat and nose. In the majority, the analysis of chosen risk factors shown no significant difference between the group of carriers and non-carriers.
Conclusions: The results of the research among future medical staff confirm the high rate of S. aureus carriage
observed in the medical profession. In the majority, carriage concerned was MSSA. The presence of MRSA
carriers was considerably too high, which pose a risk of invasive infections. Elimination of the pathogen is then a very important preventive action. Nevertheless, the research was carried among small group and further
studies are needed in to confirm the initial results.
Key words: Staphylococcus aureus, carriage, methicillin-resistance

45
Dagmara Dobrzyńska1 2, Adrianna Dubisz
2, Katarzyna Marchwińska
2
Ocena wrażliwości na wybrane antybiotyki bakterii Staphylococcus spp. wyizolowanych z powietrza
1. Wprowadzenie
Powietrze atmosferyczne jest siedliskiem zróżnicowanej mikroflory, pośród której najczęściej występują bakterie z rodzaju Micrococcus, Staphylococcus, Bacillus oraz grzyby z rodzaju Alternaria, Penicillium, Aspergillus, Cladosporium czy też Mucor. Jako mieszanina gazów, powietrze nie stwarza odpowiednich warunków do wzrostu drobnoustrojów, jest to jednak środowisko w którym można zaobserwować obecność zróżnicowanych mikroorganizmów, w tym także chorobotwórczych. Wszystkie drobnoustroje obecne w powietrzu atmosferycznym występują w postaci bioaerozolu. Jest to układ dwu- lub trójfazowy, który składa się z fazy rozpraszającej – stanowi ją powietrze i rozproszonej czyli stałej lub ciekłej. Powstający bioaerozol to zbiór rozproszonych cząsteczek takich jak pyłki kwiatów, nasiona, kurz, komórki wegetatywne i przetrwalne bakterii, fragmenty strzępek grzybów i zarodniki jak również komórki drożdży i wirusy. Wysoka liczba drobnoustrojów w powietrzu w zamkniętych pomieszczeniach, w których występuje duża rotacja ludzi warunkuje skład ilościowy i jakościowy bioaerozolu i przyczynia się do pogorszenia stanu higie-nicznego, może tym samym prowadzić do powstawania infekcji [1-3].
Człowiek w miejscach o dużym zaludnieniu może być narażony na działania zróżnicowanych czynników szkodliwych m.in. psychospołecznych, fizycznych oraz biologicznych. Jakościowe i ilościowe zanieczyszczenie środowiska drobnoustrojami istotnie wpływa na występowanie chorób. Bakterie i grzyby mogą powodować m.in. katar sienny, astmę, zapalenie oskrzeli, zapalenie płuc, reakcje alergiczne, gruźlicę a także zapalenie zatok i spojówek [4-6]. Dotychczas do zwalczania infekcji pochodze-nia bakteryjnego najczęściej stosowano antybiotyki. Niestety bakterie na skutek mutacji i horyzontalnego transferu genów coraz szybciej dostosowują się do nieko-rzystnego dla nich środowiska. W wyniku tych mechanizmów obronnych wzrasta ich odporność na antybiotyki. Jest to przyczyną pogłębiającego się w ostatnim czasie problemu antybiotykooporności. Obecnie większość z drobnoustrojów wykazuje naturalną oporność na znaczną liczbę antybiotyków oraz zdolność nabywania nowych mechanizmów oporności. Geny oporności są przekazywane przez bakterie w proce-sach koniugacji, transformacji i transdukcji. Zjawisko antybiotykooporności narasta z taką szybkością że obecnie niektóre bakterie jak np. Klebsiella pneumoniae NDM, zwana potocznie New Delhi, wykazują oporność na wszystkie dostępne antybiotyki. Ich zwalczanie jest niemożliwe tradycyjnymi metodami, a przy użyciu eksperyment-talnych technik często stosowane są potencjalnie toksyczne leki [7, 8].
1 [email protected]; 2 Studenckie Koło Naukowe INVENTUM, Katedra Przyrodniczych Podstaw Jakości, Wydział
Towaroznawstwa, Uniwersytet Ekonomiczny w Poznaniu

Dagmara Dobrzyńska, Adrianna Dubisz, Katarzyna Marchwińska
46
Antybiotykooporność stała się poważnym zagrożeniem dla zdrowia publicznego na całym świecie, dlatego też nie można bagatelizować stanu higienicznego pomieszczeń, w których następuje duża wymiana ludzi, jak np. w urzędach, dworcach czy na uczelniach. W miejscach tych powinno się przeprowadzać okresowe badania czystości powierzchni oraz powietrza [2, 9].
Niewątpliwie na wszystkich uczelniach sesja jest okresem, kiedy w stosunkowo krótkim czasie w uczelnianych pomieszczeniach przebywa wielu studentów i nauczy-cieli akademickich. Jest to doskonała okazja do rozprzestrzeniania się bakterii. Dlatego w celu izolacji bakterii Staphylococcus wybrano okres sesji zimowej oraz pomiesz-czenia Uniwersytetu Ekonomicznego w Poznaniu, w których rotacja studentów była bardzo duża: hol główny, w którym mieszczą się szatnia, toaleta damska oraz winda.
2. Cel badania
Celem badań było oznaczenie wrażliwości na wybrane antybiotyki bakterii z rodzaju Staphylococcus wyizolowanych z powietrza pomieszczeń Uniwersytetu Ekonomicznego w Poznaniu.
3. Metodyka badań
3.1. Materiał badawczy
Przedmiotem badań było 15 izolatów bakterii z rodzaju Staphylococcus, pocho-dzących z powietrza pobranego z holu, windy oraz jednej z toalet usytuowanych na Uniwersytecie Ekonomicznym w Poznaniu.
3.2. Metody badań
3.2.1. Izolacja mikroorganizmów z powietrza
Z określonego miejsca pobrano 1 m³ powietrza metodą zderzeniową, za pomocą aparatu do pomiaru czystości powietrza atmosferycznego BC 100 BIOCOLLECTOR na płytki Petriego z zestalonym podłożem ilościowym Plate Count LAB-AGAR (BIOCORP) oraz selektywnym Baird Parker LAB-AGAR (BIOCORP). Następnie
płytki z pobranym materiałem inkubowano przez 24 godziny w temperaturze 37C. W celu izolacji czystych kultur, pojedyncze kolonie zostały posiane na podłoże Baird Parker LAB AGAR z wykorzystaniem metody posiewów redukcyjnych [10].
3.2.2. Określenie właściwości hemolitycznych mikroorganizmów
Wyizolowane kultury drobnoustrojów przeszczepiono metodą powierzchniową na podłoże selektywne Columbia Agar z 5% dodatkiem krwi baraniej (BIOCORP) w celu określenia ich właściwości hemolitycznych. Próbki inkubowano przez 24 godziny w temperaturze 37°C.
3.2.3. Określenie morfologii mikroorganizmów
Określenie morfologii komórek mikroorganizmów wykonano na podstawie analizy preparatów mikroskopowych. Na odtłuszczone szkiełko podstawowe naniesiono kroplę 0,85% roztworu sterylnej soli fizjologicznej. Wyjałowioną ezą pobrano kolonię z hodowli na podłożu stałym i wykonano rozmaz. Preparaty utrwalono w płomieniu

Ocena wrażliwości na wybrane antybiotyki bakterii Staphylococcus spp. wyizolowanych z powietrza
47
palnika. W celu potwierdzenia przynależności bakterii do grupy Gram-dodatnich ziarniaków wykonano barwienie preparatów metodą Grama [11].
3.2.4. Oznaczanie aktywności katalazy bakterii
Wytwarzanie enzymu katalazy zbadano poprzez zawieszenie badanych bakterii, pobranych z hodowli na pożywce stałej, w kropli 3% wody utlenionej na szkiełku podstawowym. Wynik testu jest dodatni, jeżeli w ciągu 30 sekund pojawią się pęche-rzyki gazu.
3.2.5. Identyfikacja wyizolowanych bakterii na podstawie ich cech biochemicznych
Identyfikację wykonano z wykorzystaniem biochemicznego testu STAPHY test 24 (Erba Lachema). Zestaw ten przeznaczony jest do ostatecznej kierunkowej identy-fikacji gatunków rodzaju Staphylococcus oraz ich różnicowania wobec pozostałych gram dodatnich, katalazo-dodatnich ziarniaków.
Z czystej 24 godzinnej hodowli bakterii na podłożu Agar odżywczy (bez glukozy) sporządzono zawiesinę o gęstości 2 w skali McFarlanda. Następnie wprowadzano po 100 µl do każdej ze studzienek, odpowiednie studzienki zabezpieczono 2 kroplami
oleju parafinowego (ponieważ te reakcje biochemiczne zachodzą w warunkach beztlenowych). Inkubację próbek przeprowadzano w woreczkach z polietylenu w temperaturze 37°C przez 24 godziny.
Na podstawie wyników 24 barwnych reakcji wykonano identyfikację badanych bakterii poprzez porównanie zabarwienia próbek z wzorcową kartą barw dołączoną do testu przez producenta oraz interpretację uzyskanych wyników wykorzystując opro-gramowanie identyfikacyjne dostarczone przez producenta.
3.2.6. Oznaczenie wrażliwości na antybiotyki metodą dyfuzji krążkowej
Ocenę wrażliwości na wybrane antybiotyki przeprowadzono zgodnie z zaleceniami Krajowego Ośrodka Referencyjnego ds. Lekowrażliwości Drobnoustrojów (KORLD) [12]. Oznaczenie wrażliwości na antybiotyki zostało przeprowadzone przy użyciu
podstawowego antybiogramu, który składa się z: cefoksytyny (30 g), erytromycyny
(15 g) oraz klindamycyny (2 g) (OXOID) (Tabela 1).
Tabela 1. Wykaz antybiotyków zastosowanych w doświadczeniach
L.p. Antybiotyk Podział ze względu na budowę chemiczną
Stężenie [µl]
Skrót
Oddziaływanie bakteriobójcze
1. cefoksytyna beta-laktamy (cefemy) 30 FOX
Oddziaływanie bakteriostatyczne
2. erytromycyna makrolidy 15 E
3. klindamycyna linkozamidy 2 DA
Źródło: Opracowanie własne

Dagmara Dobrzyńska, Adrianna Dubisz, Katarzyna Marchwińska
48
Z 24 godzinnych hodowli badanych bakterii, prowadzonych na agarowym podłożu Mueller Hinton, przygotowano zawiesiny bakteryjne o gęstości 0,5 w skali McFarlanda i wykonano posiew metodą zalewową z wykorzystaniem podłoża Mueller Hinton Agar. Następnie naniesiono krążki antybiotykowe na zestaloną powierzchnię pożywki. Próbki inkubowano w temperaturze 37°C przez 24 godziny. Badania przeprowadzono w trzech powtórzeniach. Wrażliwość badanych bakterii na zasto-sowane antybiotyki odczytywano w formie średnic stref zahamowanego wzrostu wokół krążków. Interpretację wyników przeprowadzonych oznaczeń wykonano w oparciu o wytyczne zawarte w rekomendacjach KORLD [12].
4. Wyniki i ich analiza
Analiza mikrobiologiczna powietrza wykazała niewielką liczbę bakterii z rodzaju Staphylococcus w ilości 12 jtk/m³ w holu, 23 jtk/m³ w toalecie oraz 17 jtk/m³ w windzie, co przedstawione jest na wykresie 1.
Wykres 1. Liczba jednostek tworzących kolonie wyizolowanych z powietrza [opracowanie własne]
Na podstawie oceny makroskopowej kolonii na podłożu agarowym Baird Parker, zaobserwowano występowanie wyłącznie czarnych kolonii. Na podłożu Plate Count LAB-AGAR odnotowano natomiast zróżnicowany wzrost mikroorganizmów (Foto-grafie 1-3). Kolonie charakteryzowały się średnicą od 1 do 4 mm, zabarwieniem od mlecznego do żółtego oraz różnym stopniem przejrzystości.
Fotografia 1. Fotografia 2. Fotografia 3.
Fotografie 1-3. Posiewy powietrza wykonane metodą zderzeniową na podłoże Plate Count LAB-AGAR [opracowanie własne]
0
20
40
Hol Toaleta winda
12
23 17
Licz
ba
kolo
nii
[jtk
/m³]
Miejsce

Ocena wrażliwości na wybrane antybiotyki bakterii Staphylococcus spp. wyizolowanych z powietrza
49
Wyizolowane szczepy wykazywały wszystkie typy hemolizy (Wykres 2), co wskazuje na różnorodność pozyskanych izolatów. Bakterie z rodzaju Staphylococcus wywołują trzy rodzaje lizy erytrocytów. Hemoliza alfa stanowi częściowe zniszczenie krwinek czerwonych przez badane bakterie, objawia się zazielenieniem wokół strefy wzrostu, która jest mało przejrzysta i słabo odgraniczona. Hemoliza beta charaktery-zuje się całkowitym rozpadem erytrocytów owczych, obserwuje się strefy wyraźnego przejaśnienia wokół wzrostu bakterii. Natomiast hemoliza gamma nie powoduje lizy krwinek, w ocenie makroskopowej nie obserwuje się zmian wokół kolonii bakterii [11]. Hemolizę alfa zaobserwowano w przypadku wzrostu 3 wyizolowanych bakterii. Hemolizę beta i strefę wyraźnego przejaśnienia wokół kolonii odczytano w przypadku 1 spośród badanych izolatów. Natomiast brak lizy erytrocytów zaobserwowano w przypadku wzrostu 11 badanych bakterii na podłożu agarowym uzupełnionym krwią baranią.
Wykres 2. Liczba izolatów wykazująca dany rodzaj hemolizy [opracowanie własne]
Przynależność wyizolowanych bakterii do grupy Gram-dodatnich ziarniaków została potwierdzona na postawie oceny morfologii (Fotografie 4-6). W preparatach mikroskopowych występowały zarówno pojedyncze komórki bakterii jak również charakterystyczne skupiska, które przypominały grona.
Fotografia 4. Fotografia 5. Fotografia 6.
Fotografie 4-6. Obraz mikroskopowy ziarniaków Gram-dodatnich wyizolowanych z holu (Fotografia 1), toalety (Fotografia 2) oraz windy (Fotografia 3) [opracowanie własne]
3
1
11
Hemoliza alfa
Hemoliza beta
Hemoliza gamma
Dagmara Dobrzyńska, Adrianna Dubisz, Katarzyna Marchwińska
50
Badanie aktywności katalazy wykazało, że wszystkie spośród badanych mikroorga-nizmów były katalazo-dodatnie. Enzym ten katalizuje rozkład nadtlenku wodoru na wodę i tlen. Na szkiełku podstawowym, po dodaniu wody utlenionej do zawiesiny komórek bakterii, obserwowano obfite wydzielanie się pęcherzyków gazu [11]. Taki wynik badania w toku identyfikacyjnym, po raz kolejny potwierdza przynależność wyizolowanych mikroorganizmów do grupy bakterii z rodzaju Staphylococcus, ponieważ wytwarzanie enzymu katalazy jest cecha charakterystyczną gronkowców.
Następnie na podstawie reakcji barwnych przyporządkowano 15 badanych izolatów za pomocą biochemicznego testu do identyfikacji STAPHY test 24 do gatunków: Staphylococcus aureus (2), Staphylococcus chromogenes (3), Staphylococcus epidermidis (4), Staphylococcus haemolyticus (5) oraz Staphylococcus intermedius (1).
W celu potwierdzenia wyników identyfikacji na podstawie testów biochemicznych planowana jest dalsza diagnostyka.
Badanie wrażliwości na antybiotyki wykazało, że wyizolowane gronkowce charakteryzowały się różną opornością na użyte antybiotyki (Tabela 2). Wszystkie poddane doświadczeniom izolaty wrażliwe były na działanie cefoksytyny w zastoso-wanym stężeniu.
Fotografia 7. Fotografia 8.
Fotografia 9. Fotografia 10.
Fotografie 7-10 Wrażliwość badanych izolatów bakterii na działanie cefoksytyny, erytromycyny oraz klindamycyny (Fotografia 7. – S. haemolyticus, Fotografia 8. – S. haemolyticus, Fotografia 9. – S. aureus,
Fotografia 10. – S. epidermidis) [opracowanie własne]
Ocena wrażliwości na wybrane antybiotyki bakterii Staphylococcus spp. wyizolowanych z powietrza
51
Sześć badanych izolatów: S. chromogenes (1), S. haemolyticus (4), S. aureus (1), nie wykazywało oporności na żaden z badanych antybiotyków. Rodzaj i występowanie fenotypów oporności było uzależnione od wrażliwości poszczególnych badanych bakterii na klindamycynę i erytromycynę. Izolaty oporne na erytromycynę i wrażliwe na klindamycynę oznaczono jako MSB. Osiem z pośród badanych izolatów bakterii wykazywało ten rodzaj fenotypu oporności: S. chromogenes (1), S. haemolyticus (1), S. intermedius (1), S. aureus (1), S. epidermidis (4). W przypadku infekcji spowo-dowanych przez bakterie z rodzaju Staphylococcus wykazujące ten rodzaj fenotypu oporności, nie zaleca się stosowania makrolidów 14- i 15-węglowych oraz stretto-gramin z grupy B.
Tabela 2. Wyniki wrażliwości na badane antybiotyki oraz testu identyfikacji STAPHYtest24
Identyfikacja STAPHYtest 24
Wrażliwość izolatów na wybrane antybiotyki Rodzaj fenotypu oporności
cefoksytyna klindamycyna erytromycyna
A B A B A B
S. chromogenes
33,67 ± 0,47 S 24,00 ± 0,82 S 26,67 ± 0,47 S -
36,00 ± 0,82 S 23,67 ± 1,25 S 9,33 ± 0,47 R MSB
34,00 ± 0,82 S 6,00 ± 0,82 R 0,00 ± 0,00 R MLSB
S. haemolyticus
28,67 ± 1,25 S 25,33 ± 1,25 S 21,67 ± 0,94 S -
33,00 ± 0,82 S 28,00 ± 0,82 S 10,00 ± 0,82 R MSB
34,00 ± 1,63 S 17,67 ± 0,47 S 36,00 ± 1,41 S -
33,33 ± 0,47 S 29,67 ± 0,47 S 31,00 ± 0,82 S -
24,33 ± 0,47 S 25,67 ± 0,47 S 21,33 ± 0,47 S -
S. intermedius 35,33 ± 1,25 S 23,00 ± 0,82 S 9,33 ± 0,47 R MSB
S. aureus 36,67 ± 0,94 S 25,67 ± 0,47 S 0,00 ± 0,00 R MSB
23,00 ± 0,82 S 25,33 ± 1,25 S 25,00 ± 0,82 S -
S. epidermidis
40,33 ± 1,89 S 26,67 ± 0,47 S 9,33 ± 0,47 R MSB
28,33 ± 1,25 S 21,67 ± 0,47 S 10,00 ± 0,82 R MSB
30,00 ± 0,00 S 25,33 ± 0,94 S 9,33 ± 0,47 R MSB
32,00 ± 0,82 S 24,33 ± 0,47 S 0,00 ± 0,00 R MSB
Legenda: A – strefa zahamowania wzrostu (mm), B – wrażliwość na badane antybiotyki: S – wrażliwy (sensitive), R – oporny (resistant), MSB – izolat oporny na erytromycynę i wrażliwy na klindamycynę, MLSB – izolat oporny na erytromycynę i na klindamycynę. Źródło: Opracowanie własne
Izolat oporny zarówno na erytromycynę jak i klindamycynę oznaczono jako MLSB. U bakterii z rodzaju Staphylococcus może występować bowiem mechanizm oporności typu MLSB, który oznacza oporność na makrolidy, linkozamidy i streptograminy B. Oporność ta może występować jako konstytutywna lub indukcyjna. Ponieważ nie

Dagmara Dobrzyńska, Adrianna Dubisz, Katarzyna Marchwińska
52
zaobserwowano spłaszczenia stref zahamowanego wzrostu bakterii w przypadku antybiogramów izolatu S. chromogenes wykazującego oporność na erytromycynę i na klindamycynę, jest to oporność MLSB konstytutywna. Infekcje spowodowane przez bakterie o tym fenotypie oporności nie powinny być leczone z wykorzystaniem makrolidów, linkozamidów i streptogramin z grupy B [12].
5. Wnioski
Przeprowadzone doświadczenia pozwoliły na sformułowanie następujących wniosków: Wykonane badania umożliwiły wyizolowanie z powietrza 15 izolatów należących
prawdopodobnie do 5 gatunków z rodzaju Staphylococcus: S. aureus, S. chromo-genes, S. epidermidis, S. haemolyticus oraz S. intermedius.
Wszystkie izolaty bakterii charakteryzowały się wrażliwością na działanie cefoksytyny, zróżnicowane wyniki uzyskano w przypadku klindamycyny, i erytromycyny. Analiza uzyskanych wyników prowadzi do stwierdzenia, że izolaty pozyskane w badaniu nie stanowią problemu w potencjalnej anty-biotykoterapii.
Stwierdzono, że zanieczyszczenie powietrza gronkowcami w pomieszczeniach Uniwersytetu Ekonomicznego w Poznaniu nie stwarza bezpośredniego niebezpie-czeństwa dla osób tam przebywających, ponieważ stanowią one głównie mikro-florę skóry człowieka. Ruch powietrza oraz różny poziom czystości pomieszczeń ma istotny wpływ na występowanie aerozolu bakteryjnego, ale ilościowe zanie-czyszczenie powietrza w badanych miejscach nie było wysokie.
Literatura
1. Chmiel M., Frączek K., Grzyb J. Problemy monitoringu zanieczyszczeń mikrobiolo-gicznych powietrza, Woda-Środowisko-Obszary Wiejskie, 15(1), (2015), s. 17-27.
2. Hospodsky D., Qian J., Nazaroff W. W., Yamamoto N., Bibby K., Rismani-Yazdi H., Peccia J. Human occupancy as a source of indoor airborne bacteria. Public Library of Science One, 7(4), (2012), e34867. doi:10.1371/journal.pone.0034867
3. Yassin M. F., Almouqatea S. Assessment of airborne bacteria and fungi in an indoor and outdoor environment. International Journal of Environmental Science & Technology, 7(3), (2010), s. 535-544.
4. Lee J., Jo W. Characteristics of indoor and outdoor bioaerosols at Korean high-rise apartment buildings, Environmental Research 101, (2006), s. 11-17.
5. Jo W., Kang J., Workplace exposure to bioaerosols in pet shop, pet clinics and flower garden, Chemosphere 65, (2006), s. 1755-1761.
6. Maus R., Goppelsroder A., Umhauer H. Survival of bacterial and mold spores in air filter media, Atmospheric Environment 35, (2001), s. 105-113.
7. Todar K. Bacterial Resistance to Antibiotics. Lectures in Microbiology, (2009). http://textbookofbacteriology.net/resantimicrobial.html
8. Kumarasamy, K. K., Toleman, M. A., Walsh, T. R., Bagaria, J., Butt, F., Balakrishnan, R., Chaudhary U., Doumith M., Giske C., Irfan S., Krishnan P., Kumar A., Maharjan S., Mushtaq S., Noorie T., Paterson D., Pearson A., Perry C., Rachel Pike, Bhargavi Rao, Ujjwayini Ray, Jayanta B Sarma, Madhu Sharma, Sheridan E., Thirunarayan M., Turton J., Upadhyay S., Warner M., Welfare W., Livermore D., Woodford N. Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. The Lancet infectious diseases, 10(9), (2010), s. 597-602.

Ocena wrażliwości na wybrane antybiotyki bakterii Staphylococcus spp. wyizolowanych z powietrza
53
9. Antybiotykooporność: Zagrożenie dla zdrowia publicznego, Materiał prasowy Europejskiego Dnia Wiedzy o Antybiotykach i Światowego Tygodnia Wiedzy o Antybiotykach, (2015). http://antybiotyki.edu.pl/edwa/pdf/EDWA_inf%20prasowa_2015.pdf
10. Kaiser K., Wolski A., Kontrola czystości mikrobiologicznej powietrza, Technika chłodnicza i klimatyzacyjna, 4 (2007), s. 158-162.
11. Szewczyk E. Ćwiczenia z mikrobiologii i immunologii dla studentów farmacji, (2014), s. 10-13, 58-59.
12. Żabicka D., Hryniewicz W., Rekomendacje doboru testów do oznaczania wrażliwości bakterii na antybiotyki i chemioterapeutyki. Oznaczanie wrażliwości ziarniaków Gram-dodatnich z rodzaju Staphylococcus spp., Narodowy Instytut Leków Centralny Ośrodek Badań Jakości w Diagnostyce Mikrobiologicznej, (2010), s. 1-10.
Ocena wrażliwości na wybrane antybiotyki bakterii Staphylococcus spp. wyizolowanych z powietrza Streszczenie W powietrzu atmosferycznym występują zróżnicowane mikroorganizmy, jakimi są między innymi bak-terie z rodzaju Staphylococcus czy Bacillus oraz grzyby z rodzaju Penicillium, Aspergillus czy też Mucor. W zamkniętych pomieszczeniach, o zwiększonym zagęszczeniu liczby osób, mikroflora ta występuje w większym stężeniu, co może wpływać negatywnie na stan zdrowia człowieka, w tym wywoływać wiele infekcji i chorób. Celem badań było wyizolowanie, identyfikacja oraz ocena antybiotykooporności bakterii z rodzaju Staphylococcus. Bakterie te wyizolowano z powietrza w holu, windzie oraz jednej z toalet znajdujących się w budynku głównym Uniwersytetu Ekonomicznego w Poznaniu. Materiał badawczy pobrano metodą zderzeniową za pomocą aparatu do pomiaru czystości powietrza atmosferycznego BC 100 BIOCOLLECTOR. Następnie bakterie przeszczepiono na podłoże selektywne Baird Parker LAB-AGAR w celu izolacji czystych kultur, oznaczono występowanie katalazy oraz spo-rządzono preparaty mikroskopowe. Następnie wykonano badanie lekooporności izolatów przeprowadzono przy użyciu podstawowego antybiogramu, który składa się z następujących antybiotyków: cefoksytyna, erytromycyna, klindamycyna. Wykonano również kierunkową identyfikację bakterii zestawem STAPHYtest 24 dedykowanym do gramdodatnich, katalazo-dodatnich ziarenkowców. Dokonana analiza pozwoliła na wyizolowanie z powietrza 15 izolatów należących do 5 gatunków z rodzaju Staphylococcus, z czego wszystkie z nich charakteryzowały się wrażliwością na cefoksytynę. Na podstawie otrzymanych wyników, można stwierdzić że izolaty stanowiły głównie mikroflorę skóry czło-wieka, a więc nie wpływają negatywne na stan zdrowa osób przebywających na Uniwersytecie Ekono-miczny w Poznaniu. Słowa kluczowe: powietrze, gronkowce, hemoliza, antybiotykooporność, identyfikacja
Evaluation of sensitivity to selected antibiotics of Staphylococcus spp. isolated from air Abstract In atmospheric air are different microorganisms, which are for example bacteria of the genus Staphylococcus and Bacillus, and fungi of the genus Penicillium, Aspergillus and Mucor. In closed rooms with increased density of people, this microflora occur in higher concentrations, which can negative affect health, cause many infections and diseases. Research purpose was isolation, identification and evaluation of antibiotic resistance of Staphylococcus bacteria. These bacteria were isolated from the air in the lobby, elevator and one of the toilets located in the main building of Poznan University of Economics and Business. Test material was collected by crash method using BC 100 BIOCOLLECTOR atmospheric air purifier. Then the bacteria were transplanted onto selective substrate Baird Parker LAB-AGAR to isolate pure cultures, catalase were determined and microscopic preparations were prepared. Then made a research of antibiotic resistance of isolates using the basic antibiogram, which consist of the following antibiotics: cefoxitin, erythromycin, clindamycin. Also done was the directional identification of the bacteria with the STAPHYtest24 dedicated to gram positive, catalse-positive grains. The analysis allowed for isolation from the air 15 isolates belonging to 5 species of the genus Staphylococcus, all of which were sensitive to cefoxitin. Based on the obtained results, it can be stated that the isolates were mainly microflora of the human skin, so they do not affect the healthy state of people staying at Poznań University of Economics and Business. Keywords: air, Staphylococcus, haemolysis, antibiotic resistance, identification

54
Agnieszka Ludwiczak1
Badania wpływu wybranych związków na aktywność kinazy adenylanowej z Bacillus stearothermophilus
1. Wstęp
Kinazy adenylanowe (AK) (ATP-AMP fosfotransferazy; EC 2.7.4.3) należą do rodziny białek wiążących nukleozydotrójfosforany (z ang. NTP-binding proteins) [1]. Enzymy te katalizują reakcję dwukierunkowego fosfotransferu AMP i ATP zgodnie z poniższą reakcją:
Mg2+:ATP + AMP ↔ Mg2+:ADP + ADP
Obok regulacji energetyki i metabolizmu komórkowego, AK uczestniczą w sygnalizacji międzykomórkowej i reakcji na stres. W komórkach eukariotycznych zidentyfikowano kilka izoform AK, różniących się wielkością, lokalizacją komórkową i właściwościami kinetycznymi zgodnymi z wymaganiami metabolicznymi różnych typów komórek [1,2]. Izoformy kinazy adenylanowej mogą tworzyć dimery i większe struktury molekularne [3].
Zmiany stężenia nukleotydów spotyka się w etiologii wielu chorób. Między innymi główną przyczyną chorób układu krążenia jest niekontrolowana i postępująca agregacja płytek krwi. Tworzenie skrzeplin w naczyniach wieńcowych serca jest inicjowane aktywacją płytek krwi [4]. Wzrost stężenia nukleotydów we krwi pacjen-tów po zawale serca wynika ze zwiększenia preferencji kinaz adenylanowych do ATP jako substratu [5]. Uniwersalnym sposobem obniżania stężenia nukleotydów we krwi wydaje się być selektywna i ukierunkowana regulacja aktywności AK. Możliwość regulacji aktywności AK wydaje się więc bardzo atrakcyjna. Najsku-teczniejszym znanym inhibitorem kinaz adenylanowych jest Ap5A [6], zaś aktywatory tych enzymów nie są opisywane. Znajomość budowy centrum katalitycznego kinazy adenylanowej pozwala na postawienie hipotez o możliwej aktywności wybranych związków niskocząsteczkowych w stosunku do enzymu.
Celem pracy jest opracowanie metody nadekspresji i oczyszczania bakteryjnej
kinazy adenylanowej z Bacillus stearothermophilus oraz analiza aktywności enzymu w obecności małocząsteczkowych regulatorów z grupy chiralnych zasad Schiffa.
2. Materiały i metody
2.1. Materiały
2.1.1. cDNA kodujące bakteryjną kinazę adenylanową
cDNA odpowiadające sekwencji aminokwasowej kinazy adenylanowej z Bacillus stearothermophilus zdeponowanej w bazie PDB (PDB ID białka B. stearother-mophilus – 1ZIP) zostało zsyntetyzowane przez serwis GeneArt, Life Technologies Polska Sp. z o.o.
1 [email protected], Wydział Chemii, Uniwersytet Mikołaja Kopernika w Toruniu

Badania wpływu wybranych związków na aktywność kinazy adenylanowej z Bacillus stearothermophilus
55
2.1.2. Wektor wykorzystany do ekspresji kinazy adenylanowej
Wektorem wykorzystywanym w kolejnych etapach badań był bakteryjny wektor
ekspresyjny pET3a. W sekwencji tego wektora odnaleźć można silny promotor
bakteriofaga T7, specyficzny dla bakteriofagowej polimerazy T7. Istotnym jest to, że
promotor T7 nie jest skutecznie rozpoznawany przez polimerazę RNA E. coli. Zatem
system ekspresyjny pET musi obejmować również genetycznie zmodyfikowane
komórki bakteryjne. Typowymi bakteriami komplementarnymi do wektora pET są E.
coli szczep BL(DE3).
2.1.3. Szczep ekspresyjny E. coli CodonPlus BL21(DE3)pLysS
Komórki szczepu BL21(DE3)pLysS umożliwiają ekspresję białka z wysoką
wydajnością z genu, który znajduje się pod kontrolą promotora T7. Genotyp użytego
szczepu: F–, ompT, hsdSB (rB–, mB–), dcm, gal, λ(DE3), pLysS, Cmr. Szczep E.
coli BL21(DE3)pLysS jest oporny na chloramfenikol, zaś oporność na ampicylinę
nadaje wektor.
2.2. Metody
2.2.1. Klonowanie
W badaniach do klonowania użyto komórek bakteryjnych E. coli NovaBlue szczep
BL21(DE3)pLysS.
2.2.2. Transformacja komórek szczepu ekspresyjnego E. coli NovaBlue
szczep BL21(DE3)pLysS
Zamrożone w -80°C kompetentne komórki szczepu ekspresyjnego E. coli
NovaBlue szczep BL21(DE3)pLysS umieszczono na 30 minut w lodzie. Po tym czasie
do probówki z bakteriami dodano 1 μl plazmidu zligowanego z insertem, wymieszano
i inkubowano w lodzie 20 minut. Następnie eppendorfy z bakteriami umieszczono
w łaźni wodnej o temperaturze 42°C i po 35 s przeniesiono na 2 minuty do lodu. Po
tym czasie dodano 250 μl płynnej pożywki SOC (2%trypton, 0,5% ekstrakt
drożdżowy, 0,05% NaCl, 2,5 mM KCl, 10 mM MgCl2, 10 mM MgSO4, 20 mM
glukoza) ogrzanej do temperatury pokojowej i zamieszano. Probówki umieszczono na
1 godzinę w wytrząsarce (37°C, 100 rpm). Całą procedurę prowadzono w warunkach
sterylnych.
2.2.3. Hodowla stansformowanych bakterii
Na dwie szalki ze stałą pożywką LB (0,5% (w/o) ekstrakt drożdżowy, 1% (w/o)
trypton, 1% (w/o) NaCl, 1,5% agaroza) dodano odpowiednio 50 μl lub 100 μl trans-
formowanych bakterii. Bakterie rozprowadzono równomiernie na szalce. Płytki
inkubowano 16 h w 37ºC, bez wytrząsania. Pojedynczą kolonię przeniesiono sterylną
ezą z szalki do kolby z płynną pożywką LB (10 ml) z antybiotykami i glukozą.
Pozostawiono na 12 h w temperaturze 37ºC z wytrząsaniem (250 rpm). W celu
zamrożenia komórek bakteryjnych, bakterie hodowano 12 h w temperaturze 37ºC

Agnieszka Ludwiczak
56
z wytrząsaniem (250 rpm) do gęstości optycznej OD600= 1,5. Następnie do hodowli
dodano pożywkę LB z 60% glicerolem w stosunku 1:1 i wymieszano. Zawiesinę
komórek podzielono na 100 μl porcje, zamrażano w ciekłym azocie i przechowywano
w temperaturze -80ºC.
W celu wykonania hodowli właściwej do 500 ml pożywki LB dodano 5 ml hodowli
startowej. Kolbę z hodowlą właściwą umieszczono w 37ºC z wytrząsaniem (250 rpm).
Po 2 godzinach zmierzono spektrofotometrycznie gęstość optyczną hodowli i porów-
nano z gęstością próby odniesienia (pożywka LB). Po osiągnięciu przez hodowlę
bakteryjną gęstości (OD600= 0,6-0,8) bakterie indukowano dodając IPTG do
końcowego stężenia 1mM. Hodowlę prowadzono przez 3h w 37ºC z wytrząsaniem
(250 rpm). Następnie temperaturę obniżono do 22°C i hodowlę prowadzono jeszcze
przez 12-16 godzin z wytrząsaniem (250 rpm). Po tym czasie bakterie chłodzono
w lodzie i wirowano 30 minut przy 3000 rpm w 4°C. Osad komórek bakteryjnych
zawieszano w 10 ml buforu startowego (50 mM HEPES, pH 7,6) i przechowywano
w temperaturze -20°C.
2.2.4. Liza komórek bakteryjnych
Osad komórek bakteryjnych zawieszony w 10 ml 50 mM buforu HEPES, pH 7,6
rozmrożono i dodano mieszaniny inhibitorów proteaz (firmy Sigma). Tak przygoto-
waną zawiesinę bakterii sonikowano z udziałem dezintegratora UD-20 firmy Techpan
w lodzie 3 x 15s z 30-sekundowymi przerwami pomiędzy każdym z cyklów sonikacji.
Lizat bakteryjny po sonikacji wirowano przez 10 minut w 4°C przy 20 000 x g.
Supernatant zawierający kinazę adenylanową przenoszono do czystych probówek,
a osad zawierający frakcję białek nierozpuszczalnych zawieszano w 50 mM HEPES,
pH 7,6. Otrzymane preparaty przechowywano w temperaturze -20ºC.
2.2.5. Denaturacja termiczna białek balastowych obecnych w preparacie
kinazy adenylanowej z B. stearothermophilus
Denaturacja termiczna białek kontaminujących preparat kinazy adenylanowej z B.
stearothermophilus była dwuetapowa. Pierwszy z etapów obejmował denaturację
białek kontaminujących w 60°C przez 15 minut, zaś kolejny denaturację w 70°C przez
15 minut.
2.2.6. Chromatografia powinowactwa na Blue Sepharose z użyciem
rosnącego gradientu stężenia roztworu NaCl
Chromatografię powinowactwa na Blue Sepharose prowadzono na kolumnie
o wymiarach 1,5 cm x 8 cm wypełnionej 14 ml złoża. Kolumnę zintegrowano
z zestawem EconoPump firmy Biorad i równoważono 100 ml buforu startowego (50
mM HEPES, pH 7,6). Na kolumnę naniesiono roztwór białka o stężeniu 250 mg/ml
(V= 10 ml). Następnie kolumnę przemywano buforem HEPES pH 7,6. Białko
z kolumny eluowano stosując gradient stężenia roztworu NaCl (0-2M). Rozdział
chromatograficzny prowadzono z szybkością 0,5 ml/min, objętość zbieranych frakcji
wynosiła 3 ml. W zebranych frakcjach dokonano spektrofotometrycznego pomiaru

Badania wpływu wybranych związków na aktywność kinazy adenylanowej z Bacillus stearothermophilus
57
zawartości białka przy λ= 280nm oraz metodą HPLC analizowano aktywność
transferazową kinazy adenylanowej.
2.2.7. Dializa i zagęszczanie preparatu enzymatycznego
W celu usunięcia NaCl z preparatu białkowego zastosowano kilkukrotne przemy-
wanie roztworu białka 50 mM buforem HEPES pH 7,6. Preparat przeniesiono do filtra
Amicon Ultra 10K o objętości 50 ml i uzupełniono do 30 ml 50 mM buforem HEPES
pH 7,6. Wirowano 3000 x g przez 20 minut w 4°C, aż do momentu, kiedy w przesączu
nie było już jonów NaCl. Pozbawiony soli roztwór białka, wstępnie zagęszczono do
objętości 2 ml. Zagęszczanie białka do końcowego stężenia przeprowadzono
z użyciem filtrów Amicon Ultra 10K o objętości 0,5 ml. Roztwór białka przeniesiono
do filtra i wirowano przez 15 minut (14,000 x g) oraz 1 minutę (1,000 x g) dla odzysku
roztworu białka z filtra.
2.2.8. Elektroforetyczny rozdział białek w żelu poliakrylamidowym z SDS
(SDS-PAGE)
Do rozdziału białek stosowano 4% żel zagęszczający. Składniki mieszaniny
akrylamid/metyleno-bis-akrylamid, buforu żelowego I i II prezentuje poniższa tabela
(Tabela 1).
Tabela 1. Składniki mieszanin użytych do elektroforezy (SDS-PAGE).
Rodzaj odczynnika Skład
akrylamid/metyleno-bis-akrylamid 28% akrylamid + 3%
bisakrylamid
bufor żelowy I 1,5 M Tris-Cl pH 8,8 z 0,4% SDS
bufor żelowy II 0,5 M Tris-Cl pH 6,8 z 0,4% SDS
Źródło: [7]
Przed nałożeniem na żel roztwór białka mieszano w stosunku 1:1 z buforem
redukującym (100 mM Tris-HCl, 4% SDS, 10% β-merkaptoetanol, 20% glicerol
i 0,2% błękit bromofenolowy, pH 6,5). Tak przygotowane próby denaturowano
termicznie (15 min w 95°C) i krótko wirowano (1 minuta, 1000 x g). Rozdział białek
prowadzono w temperaturze pokojowej w buforze elektrodowym o następującym
składzie: 25 mM Tris-HCl, 60 mM glicyna, 1% SDS, pH 8,3. Elektroforezę prowa-
dzono w aparacie Bio-Rad Mini-Protean 3 Cell. Próby o odpowiedniej zawartości
białka nanoszono do poszczególnych studzienek metodą podwarstwiania. Elektrofo-
rezę prowadzono przy napięciu 90 V, aż do wniknięcia prób do żelu, następnie
napięcie zwiększono do 120 V i elektroforezę prowadzono do momentu, aż barwnik
markerowy znalazł się kilka milimetrów od dolnego brzegu żelu (1,5 godziny).
Stosowano następujące markery mas cząsteczkowych: Unstained Protein Molecular
Wright Marker 116-11,4 kDa i Spectra Multicolor Broad Range Protein Ladder.

Agnieszka Ludwiczak
58
2.2.9. Analiza aktywności kinazy adenylanowej
Kinaza adenylanowa katalizuje dwukierunkową reakcję, zatem konieczne było
zbadanie przebiegu katalizy w kierunku tworzenia ADP oraz AMP i ATP.
ATP + AMP ↔ 2 ADP
2 ADP ↔ AMP + ATP
Reakcję enzymatyczną inicjowano dodając 25 μl enzymu (1 μg kinazy adenyla-
nowej B. stearothermophilus) do 25 μl mieszaniny reakcyjnej. Stosunek objętości
roztworu białka do objętości mieszaniny reakcyjnej wynosił 1:1.
Skład mieszaniny reakcyjnej:
50 mM bufor HEPES, pH 7,5,
1 mM MgCl2,
nukleotyd adenozynowy (ADP= 12 mM oraz ATP i AMP= 5 mM),
badany regulator w stężeniu 50 μM i 100 μM.
Badane związki regulatorowe rozpuszczono w mieszaninie DMSO-woda 1:1. Czas inkubacji enzymu w mieszaninie reakcyjnej wynosił 10 minut w temperaturze
37°C. Reakcję hamowano dodając do mieszaniny 50 μl 1M zimnego HClO4. Następnie
próby wytrząsano, zobojętniano 25 μl 1M KOH, ponownie wytrząsano i wirowano
w 4°C 18000 rpm przez 15 minut. Do 80 μl supernatantu dodano 15 μl n-heptanu,
w celu usunięcia białka z prób. Następnie wirowano w 4°C 18000 rpm przez 10 minut.
2.2.10. Analiza nukleotydowych produktów reakcji enzymatycznej
Analizę ilościową i jakościową nukleotydowych produktów reakcji przeprowa-
dzono z zastosowaniem chromatografii cieczowej w odwróconej fazie (RP-HPLC),
przy użyciu zestawu HPLC firmy Waters: In-Line Degasser AF, Waters 515 HPLC
Pump, Waters Pump Control Module, Waters 2487 Dual λ Absorbance Detector,
Waters 717 Plus Autosampler. Rozdziału produktów reakcji enzymatycznej dokonano
na kolumnie Merck Chromolith Performance 100-4,6 mm z prekolumną Merck
Chromolith Performance. Kolumnę równoważono 0,1 M buforem KH2PO4/K2HPO4,
pH 7,0, zawierającym 5 mM EDTA, 25 mM TBA (przeciwjon) i 2% MetOH. Rozdział
prowadzono z szybkością 1 ml/min. Pomiaru absorbancji dokonywano przy długości
fali λ= 260 nm odpowiadającej maksimum absorbancji większości puryn i pirymidyn.
3. Wyniki
3.1. Oczyszczanie kinazy adenylanowej przez denaturację termiczną białek
balastowych
Po uzyskaniu wydajnej nadekspresji białek w szczepach bakteryjnych, preparat
kinazy adenylanowej oczyszczono wykorzystując termostabilność AK. W tym celu
zastosowano dwukrotną 15-minutową denaturację termiczną w temperaturach 60°C
i 70°C. W tych warunkach kinaza adenylanowa pozostaje enzymatycznie aktywna.
Dwustopniowa denaturacja termiczna usunęła znaczną część białek towarzyszących.

Badania wpływu wybranych związków na aktywność kinazy adenylanowej z Bacillus stearothermophilus
59
Preparaty enzymatyczne wymagały jednak kolejnych etapów oczyszczania w celu
pozyskania homogennego enzymu (Rys. 1).
Rysunek 1. Elektroforogram białek supernatantu uzyskanego na kolejnych etapach denaturacji termicznych.
M) marker mas cząsteczkowych, 1) frakcja białek supernatantu hodowli B. stearothermophilus przed
denaturacją termiczną, 2) frakcja po denaturacji białek kontaminujących w 60°C, 3) frakcja po denaturacji
białek kontaminujących w 70°C. Źródło: opracowanie własne
3.2. Oczyszczanie kinazy z użyciem chromatografii powinowactwa
na Blue Sepharose
W celu uzyskania homogennego preparatu termofilnej kinazy adenylanowej
przeprowadzono oczyszczanie techniką chromatografii powinowactwa na złożu Blue-
Sepharose z zastosowaniem elucji gradientowej (0-2 M roztwór NaCl). Enzym ulegał
elucji z kolumny przy 0,5-0,6 M roztworze NaCl. Stopień oczyszczenia enzymu
prezentuje poniższy elektroforogram (Rys. 2). Chromatografia na Blue Sepharose CL-
4B pozwoliła na uzyskanie jednorodnego preparatu enzymatycznego.
Rysunek 2. Elektroforogram enzymu uzyskanego w wyniku chromatografii preparatów Blue Sepharose.
1) białka po denaturacji termicznej naniesione na kolumnę, 2, M – Marker, 3) białka uzyskane po elucji 0,6 M roztworem NaCl. Źródło: opracowanie własne
3.3. Analiza wpływu związków z grupy chiralnych zasad Schiffa na
aktywność kinazy adenylanowej z B. stearothermophilus
Chiralne zasady Schiffa jako potencjalne regulatory aktywności AK wybrano
z uwagi na ich podobieństwo strukturalne do istniejących inhibitorów nukleotydaz. Do
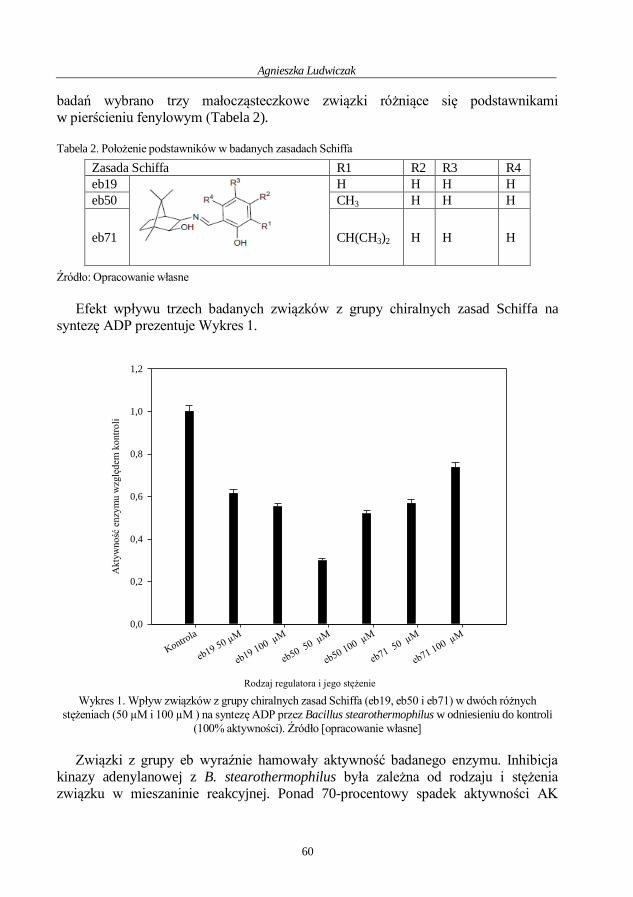
Agnieszka Ludwiczak
60
badań wybrano trzy małocząsteczkowe związki różniące się podstawnikami
w pierścieniu fenylowym (Tabela 2).
Tabela 2. Położenie podstawników w badanych zasadach Schiffa
Zasada Schiffa R1 R2 R3 R4
eb19
H H H H
eb50 CH3 H H H
eb71 CH(CH3)2 H H H
Źródło: Opracowanie własne
Efekt wpływu trzech badanych związków z grupy chiralnych zasad Schiffa na
syntezę ADP prezentuje Wykres 1.
Rodzaj regulatora i jego stężenie
Kontrola
eb19 50 µM
eb19 100 µM
eb50 50 µ
M
eb50 100 µM
eb71 50 µ
M
eb71 100 µM
Akty
wno
ść e
nzy
mu w
zglę
dem
ko
ntr
oli
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Wykres 1. Wpływ związków z grupy chiralnych zasad Schiffa (eb19, eb50 i eb71) w dwóch różnych
stężeniach (50 µM i 100 µM ) na syntezę ADP przez Bacillus stearothermophilus w odniesieniu do kontroli
(100% aktywności). Źródło [opracowanie własne]
Związki z grupy eb wyraźnie hamowały aktywność badanego enzymu. Inhibicja
kinazy adenylanowej z B. stearothermophilus była zależna od rodzaju i stężenia
związku w mieszaninie reakcyjnej. Ponad 70-procentowy spadek aktywności AK
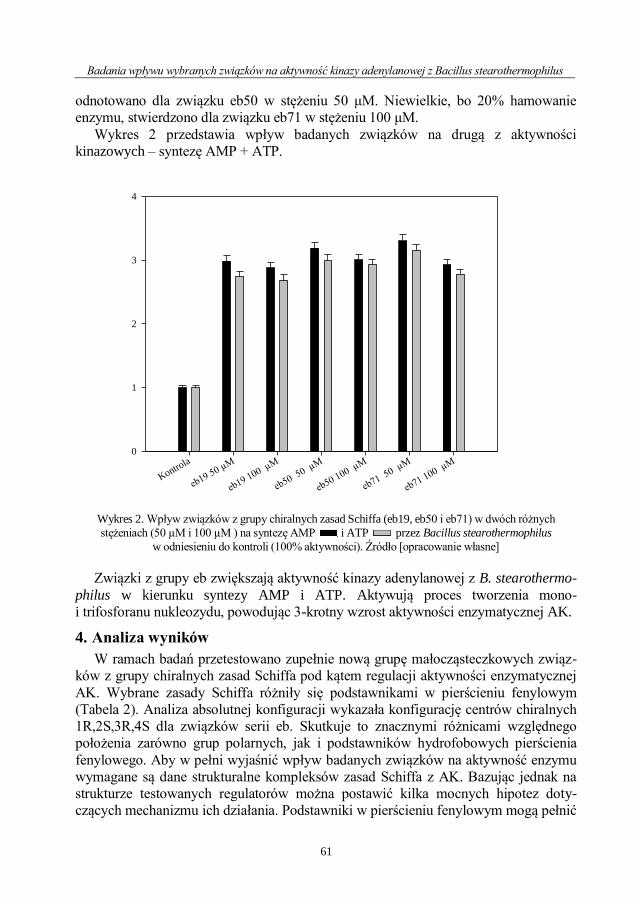
Badania wpływu wybranych związków na aktywność kinazy adenylanowej z Bacillus stearothermophilus
61
odnotowano dla związku eb50 w stężeniu 50 μM. Niewielkie, bo 20% hamowanie
enzymu, stwierdzono dla związku eb71 w stężeniu 100 μM.
Wykres 2 przedstawia wpływ badanych związków na drugą z aktywności
kinazowych – syntezę AMP + ATP.
Kontrola
eb19 50 µM
eb19 100 µM
eb50 50 µ
M
eb50 100 µM
eb71 50 µ
M
eb71 100 µM
0
1
2
3
4
Wykres 2. Wpływ związków z grupy chiralnych zasad Schiffa (eb19, eb50 i eb71) w dwóch różnych
stężeniach (50 µM i 100 µM ) na syntezę AMP i ATP przez Bacillus stearothermophilus
w odniesieniu do kontroli (100% aktywności). Źródło [opracowanie własne]
Związki z grupy eb zwiększają aktywność kinazy adenylanowej z B. stearothermo-
philus w kierunku syntezy AMP i ATP. Aktywują proces tworzenia mono-
i trifosforanu nukleozydu, powodując 3-krotny wzrost aktywności enzymatycznej AK.
4. Analiza wyników
W ramach badań przetestowano zupełnie nową grupę małocząsteczkowych związ-
ków z grupy chiralnych zasad Schiffa pod kątem regulacji aktywności enzymatycznej
AK. Wybrane zasady Schiffa różniły się podstawnikami w pierścieniu fenylowym
(Tabela 2). Analiza absolutnej konfiguracji wykazała konfigurację centrów chiralnych
1R,2S,3R,4S dla związków serii eb. Skutkuje to znacznymi różnicami względnego
położenia zarówno grup polarnych, jak i podstawników hydrofobowych pierścienia
fenylowego. Aby w pełni wyjaśnić wpływ badanych związków na aktywność enzymu
wymagane są dane strukturalne kompleksów zasad Schiffa z AK. Bazując jednak na
strukturze testowanych regulatorów można postawić kilka mocnych hipotez doty-
czących mechanizmu ich działania. Podstawniki w pierścieniu fenylowym mogą pełnić

Agnieszka Ludwiczak
62
istotną rolę w wiązaniu i pozycjonowaniu badanych związków w AK. Aktywacja AK
z B. stearothermophilus może być związana z odpowiednim położeniem grupy
hydroksylowej fragmentu kamforowego sprzyjającym utworzeniu dodatkowego
mostka pomiędzy terminalnymi fosforanami substratów. Aktywacja syntezy ATP +
AMP i hamowanie syntezy ADP potwierdza tę hipotezę (Wykres 1, Wykres 2).
Oddziaływania wiążące zasad Schiffa mogą pozycjonować grupę hydroksylową
kamfory w położeniu odpowiednim dla substratów o jednakowej długości (ADP +
ADP) stąd kierunkowa aktywacja syntezy ATP + AMP. Hamowanie aktywności AK
w kierunku syntezy ADP może wynikać z kompetycyjnego wiązania regulatora
w centrum aktywnym enzymu (Wykres 1). Jeśli inhibicja nie jest typu kompe-
tycyjnego, inhibitory mogłyby wiązać się z aminokwasami poza centrum aktywnym,
blokując osiągnięcie właściwej konformacji przez aminokwasy centrum aktywnego
AK. Aby potwierdzić powyższe hipotezy, planowane jest określenie typu inhibicji
badanych zasad Schiffa oraz wykonanie badań strukturalnych kompleksów AK
z zasadami Schiffa.
5. Podsumowanie
W niniejszej pracy poddano analizie chiralne zasady Schiffa, które aktywowały AK
z B. stearothermophilus w kierunku syntezy AMP + ATP (Wykres 2). Dotychczas
w literaturze brak jest jakichkolwiek doniesień o aktywacji AK przez małocząsteczkowe
ligandy. Dzięki takiej selektywnej aktywacji możliwe jest „włączenie” aktywności
kinazy adenylanowej danego szczepu bakterii i regulacja poziomu nukleotydów
adeninowych w komórce. Zasady Schiffa mogą stać się innowacyjnym lekiem
w chorobach, w których etiologii występują zmiany stężeń nukleotydów.
6. Podziękowania
Dziękuję zespołowi z Katedry Chemii Organicznej WCh UMK pod kierownictwem
Prof. dra hab. M. Wełniaka za syntezę chiralnych zasad Schiffa.
Praca powstała w wyniku realizacji projektu badawczego PRELUDIUM 7 (UMO-
2014/13/N/ST4/04119) finansowanego ze środków Narodowego Centrum Nauki.
Literatura
1. Dzeja P., Terzic A. Adenylate kinase and AMP signaling networks: metabolic monitoring,
signal communication and body energy sensing, International Journal of Molecular
Sciences, 10 (2009), s. 1729-1772.
2. Thach T.T, Luong T.T, Lee S. , Rhee D.K. Adenylate kinase from Streptococcus
pneumoniae is essential for growth through its catalytic activity, FEBS Open Bio, 4 (2014),
s. 672-682.
3. Henzler-Wildman K.A., Vu T., Lei M., Ott M., Wolf-Watz M., Fenn T., Pozharski E.,
Wilson M.A., Petsko G.A., Karplus M., Hübner C.G., Kern D. Intrinsic motions along an
enzymatic reaction trajectory, Nature, 450 (2007), s. 838-844.
4. Broos K., De Meyer S.F., Feys H.B., Vanhoorelbeke K., Deckmyn H. Blood platelet
biochemistry, Thrombosis Research, 129(3) (2012), s. 245-9.

Badania wpływu wybranych związków na aktywność kinazy adenylanowej z Bacillus stearothermophilus
63
5. Studzińska B., Seroka A., Lepicka M., Roszek K., Komoszyński M. The increase of
adenylate kinase activity in the blood can control aggregation of platelets in coronary or
peripheral arterial ischemia, Health 2(3) (2010), s. 246-252.
6. Feldhau P., Fröhlich T., Goody R.S., Isakov M., Schirmer R.H., Synthetic inhibitors of
adenylate kinases in the assays for ATPases and phosphokinases, European Journal of
Biochemistry 57 (1975), s. 197-204.
7. Gniot-Szulżycka J., Leźnicki A., Komoszyński M., Wojczuk B., Materiały do ćwiczeń
z biochemii. Białka. Metody ilościowego oznaczania, rozdziału i oczyszczania (2005),
Wydawnictwo UMK Toruń.
Badania wpływu wybranych związków na aktywność kinazy adenylanowej
z Bacillus stearothermophilus
Streszczenie
Kinazy adenylanowe (AK) (ATP-AMP fosfotransferazy; EC 2.7.4.3) to enzymy uczestniczące w metabo-
lizmie energetycznym i syntezie nukleotydów, które leżą u podstawy wielu chorób. Selektywna i ukierun-kowana regulacja aktywności AK może okazać się doskonałym celem terapeutycznym. Najskutecz-
niejszym znanym inhibitorem kinaz adenylanowych jest Ap5A, zaś aktywatory tych enzymów nie są poz-
nane. W ramach badań wykazano wpływ małocząsteczkowych ligandów na aktywność kinazy adenyla-
nowej. Wybrana grupa zasad Schiffa okazała się w innowacyjny i jak dotąd niespotykany sposób regulo-wać aktywność badanego enzymu. Dzięki selektywnej aktywacji możliwym stanie się „włączenie” aktyw-
ności kinazy adenylanowej danego szczepu bakterii i regulacja poziomu nukleotydów adeninowych
w komórce. Przebadane zasady Schiffa mogą stać się innowacyjnym lekiem w chorobach, w których
etiologii występują zmiany stężeń nukleotydów. Słowa kluczowe: kinaza adenylanowa, zasady Schiffa, nukleotydy.
Study of the effect of selected compounds on the activity of the adenylate kinase
from Bacillus stearothermophilus
Abstract Adenylate kinases (AK) (ATP-AMP-phosphotransferase, EC 2.7.4.3) are enzymes that are involved in
energy metabolism and synthesis of nucleotides, which are responsible of many diseases. Selective and
targeted regulation of activity of AK may be an excellent therapeutic target. The most effective known
inhibitor of the adenylate kinase is Ap5A although activators of this enzyme are not known. In this study has been shown the ability to affect on adenylate kinase activity by a small molecule ligands with a similar
arrangement of heteroatoms as in the substrate. A selected group of Schiff bases proved to be an innovative
and unparalleled way to regulate the activity of the enzyme. Thanks to the selective activation of enzyme it
will be possible to “turn on” the activity of adenylate kinase in the bacterial strain and control the adenine nucleotides in the cell. Thus tested Schiff bases may be an innovative drugs for diseases with changes of
nucleotide concentrations in etiology.
Keywords: adenylate kinase, Schiff bases, nucleotides.

64
Magdalena Bossowska-Nowicka1, Felix N. Toka
2, Lidia Szulc-Dąbrowska
3
Rola katepsyn w zakażeniach bakteryjnych
1. Wprowadzenie
Katepsyny cysteinowe są enzymami proteolitycznymi, znajdującymi się głównie
w lizosomach, gdzie bezpośrednio przyczyniają się do zabijania patogenów lub
pośrednio prowadzą do jego zwalczenia poprzez udział w procesach przetwarzania
i prezentacji antygenów. Do tej pory zidentyfikowane zostały następujące katepsyny: A,
B, C, D, E, F, G, H, L, K, O, S, V, W i X. Enzymy te są szeroko rozpowszechnione
u ludzi oraz u większości zwierząt, a poziom ich ekspresji oraz swoistość jest
zróżnicowana i zależy głównie od lokalizacji w komórce. Większość katepsyn
występuje powszechnie w różnych komórkach i tkankach, natomiast niektóre z nich,
takie jak katepsyna K, S lub W odgrywają bardzo specyficzną rolę w konkretnych
typach komórek. Ekspresja katepsyny K jest specyficzną cechą komórek biorących
udział w przebudowie kości, ale występuje również w makrofagach i komórkach
nabłonkowych. Katepsyna S znajduje się przede wszystkim w komórkach
prezentujących antygen, natomiast ekspresja katepsyny W zachodzi głównie
w komórkach NK.
Do tej pory opisano udział poszczególnych katepsyn cysteinowych w wielu zaka-
żeniach bakteryjnych, gdyż regulują one zarówno wrodzoną jak i nabytą odpowiedź
immunologiczną. Jednak dokładny mechanizm ich działania pozostaje nadal nie
wyjaśniony i wymaga dalszych badań.
2. Rola katepsyny L w zakażeniach wywołanych przez bakterie z rodzaju
Mycoplasma oraz Mycobacterium
Katepsyna L należy do endopeptydaz, czyli enzymów, które hydrolizują wiązania
wewnątrz łańcucha peptydowego. Wytwarzana jest jako preprokatepsyna L,
transportowana przez aparat Golgiego jako prokatepsyna L w pęcherzykach
wydzielniczych, a następnietrafia do lizosomów, gdzie jest przechowywana jako
dojrzała katepsyna L [1]. Wewnątrzkomórkowa degradacja białek przeprowadzana
przez katepsynę L odgrywa istotną rolę w licznych procesach komórkowych oraz
regulacji cyklu komórkowego, ze względu na zdolność do rozkładu czynników
transkrypcyjnych [2]. Jedną z istotnych funkcji tego enzymu jest również degradacja
łańcucha niezmiennego Ii podczas przetwarzania cząsteczki MHC klasy II, co jest
1 [email protected], Zakład Immunologii, Katedra Nauk Przedklinicznych, Wydział Medycyny
Weterynaryjnej, Szkoła Główna Gospodarstwa Wiejskiego w Warszawie 2 [email protected], Department of Biomedical Sciences School, Ross University of Veterinary Medicine,
St. Kitts, West Indies 3 [email protected], Zakład Immunologii, Katedra Nauk Przedklinicznych, Wydział Medycyny
Weterynaryjnej, Szkoła Główna Gospodarstwa Wiejskiego w Warszawie

Rola katepsyn w zakażeniach bakteryjnych
65
krytycznym etapem podczas prezentacji antygenu limfocytom T oraz warunkuje
rozwinięcie swoistej odpowiedzi immunologicznej [3, 4].
Rolę katepsyny L scharakteryzowano niedawno w procesach autofagii i apoptozy,
co potencjalnie może stanowić terapeutyczną strategię w leczeniu stanów patolo-
gicznych związanych z neurodegeneracją, np. wywołanych chorobą Parkinsona [5].
Ponadto, udział tej katepsyny został jednoznacznie udokumentowany podczas
wnikania takich wirusów jak Ebola, Hendra, HIV, wirusa grypy typu A czy rota-
wirusów do komórek [6-10]. Niewiele jednak wiadomo na temat znaczenia katepsyny
L w zapobieganiu zakażeń bakteryjnych.
Wpływ niedoboru katepsyny L na rozwój odpowiedzi przeciwbakteryjnej
w przebiegu zakażenia Mycoplasma pulmonis zbadano myszy z deficytem genu tej
katepsyny (Ctsl -/-) oraz myszy, u których ekspresja tego enzymu była obecna (Ctsl +/+
).
Na podstawie uzyskany wyników zaobserwowano, że myszy z wyłączonym genem
katepsyny L wykazywały upośledzenie mechanizmów odpornościowych oraz osła-
bioną produkcję TNF-α i chemokin, co skutkowało zwiększona wrażliwością na zaka-
żenie M. pulmonis. Z badań tych wynika zatem, że katepsyna L jest niezbędna do zai-
nicjowania prawidłowej odpowiedzi immunologicznej gospodarza, a odpowiedź ta jest
zaburzona podczas nieobecności tego enzymu w komórkach odpornościowych [11].
Z badań przeprowadzonych przez Nepala i współautorów [12] wynika, że aktywność
katepsyny L podczas zakażenia Mycobacterium avium i Mycobacterium tuberculosis
w makrofagach mysich pochodzących ze szpiku kostnego jest zmieniona. Zmiana ta
dotyczy powstawania aktywnej formy katepsyny L o wielkości 24 kDa. Autorzy
zaobserwowali, że w makrofagach niezakażonych przeważała forma aktywna enzymu,
natomiast komórki zakażone wykazywały zwiększone poziomy aktywności form
niedojrzałych o wielkości 30 kDa oraz 25 kDa, co świadczy o zaburzenia w powstawaniu
dojrzałej i jednocześnie aktywnej enzymatycznie formy katepsyny L. Badania te
wskazują, iż pomimo wykształconych przez makrofagi mechanizmów obrony przed
patogenami, zakażenie M. avium i M. tuberculosis powoduje znaczne nieprawidłowości
w aktywności istotnego enzymu, co prowadzi do rozwoju zakażenia [12]. Ponadto
dowiedziono, że M. tuberculosis kontroluje ekspresję katepsyn B, L i S na poziomie
mRNA, białka, a także reguluje aktywność tych enzymów w makrofagach ludzkich,
hamując ich powstawanie w komórkach, co zwiększa przeżywalność bakterii [13].
3. Udział katepsyn B i D w zakażeniach wywołanych przez Streptococcus
pneumoniae
Katepsyna B jest proteazą cysteinową, znajdującą się przede wszystkim w lizoso-
mach i wykazuje właściwości zarówno endopetydaz, jak i egzopeptydaz [14]. Zloka-
lizowana jest ona przede wszystkim w hepatocytach, kanalikach nerkowych, trzustce,
fibroblastach i gruczołach ślinowych. Główną rolą tej katepsyny jest wewnątrz-
komórkowa proteoliza białek, lecz może również uczestniczyć w takich procesach
fizjologicznych, jak przetwarzanie antygenów w odpowiedzi immunologicznej oraz
resorpcja kości.

Magdalena Bossowska-Nowicka, Felix N. Toka, Lidia Szulc-Dąbrowska
66
Natomiast katepsyna D należy do proteaz asparaginianowych. Wyróżniono trzy
formy tej katepsynyobecne w komórkach: prekursorową – prokatepsynę D, pośrednią
– pojedynczy łańcuch oraz dojrzałą – podwójny łańcuch. Prokatepsyna D, zloka-
lizowana przede wszystkim w aparacie Golgiego jest enzymatycznie nieaktywna,
natomiast formy pośrednia oraz dojrzała są aktywne enzymatycznie i znajdują się
odpowiednio w endosomach i lizosomach. Do głównych funkcji katepsyny D należy
udział w procesach autofagii i apoptozy, przez co odgrywa kluczową rolę w home-
ostazie tkankowej [15].
Zakażenia wywołane przez Streptococcus pneumoniae, wiążą się ze zwiększoną
zachorowalnością oraz śmiertelnością przede wszystkim dzieci i osób starszych.
Bakterie te są odpowiedzialne za infekcje błon śluzowych, m.in. zapalenie płuc, czy
zapalenie ucha środkowego oraz są głównym czynnikiem etiologicznym chorób
inwazyjnych takich jak zapalenie opon mózgowo rdzeniowych czy sepsa [16].
Badania przeprowadzone przez Lawrence’a i współautorów [17] dotyczyły analizy
potencjału immunosupresyjnego inhibitora katepsyny B – z-FA-FMK. Autorzy wyka-
zali, że inhibitor tej katepsyny skutecznie blokuje proliferację ludzkich limfocytów T
indukowaną przez miogeny oraz wytwarzanie IL-2 in vitro. Ponadto z-FA-FMK
obniża aktywację czynnika transkrypcyjnego NF-κB oraz dalszą produkcję cytokin
przez limfocyty T. Udowodniono także, że podawanie in vivo znacząco nasila objawy
zakażenia myszy wywołane przez S. pneumoniae w porównaniu do myszy niejaka-
żonych. Ponieważ limfocyty T odgrywają istotną rolę w uruchomieniu odpowiedzi
odpornościowej gospodarza, a ich funkcja zostaje zablokowana po podaniu inhibitora
katepsyny B wynika, że inhibitor z-FA-FMK może być lekiem o działaniu immuno-
supresyjnym.
W początkowych etapach zakażenia pneumokokami niezwykle istotną rolę odgry-
wają makrofagi, które jako pierwsze docierają do miejsca zakażenia i rozpoczynają
reakcję zapalną w organizmie. Indukowanie apoptozy przez makrofagi odgrywa istotną
rolę w obronie gospodarza przed zakażeniem [18]. Proces apoptozy wymaga interna-
lizacji bakterii do fagolizosomu, gdzie obecne są katepsyny. Aktywna katepsyna D jest
uwalniana do cytozolu i odpowiada przede wszystkim za indukcję programowanej
śmierci komórki. Badania przeprowadzone przez Bewley’a i współautorów [19] wska-
zują, że myszy z delecją genu katepsyny D wykazują znaczne zaburzenia w zdolności
do zwalczania zakażenia wywołanego przez S. pneumoniae, ze względu na zahamo-
wany proces apoptozy. Zatem prawidłowe działanie katepsyny D w komórkach
odporności wrodzonej jest niezbędne do efektywnej eliminacji pneumokoków
w organizmie.
4. Udział katepsyny B w zakażeniach wywołanych przez Francisella novicida
Francisella novicida należy do Gram-ujemnych bakterii z rodziny Francisellaceae.
Dokładna klasyfikacja tego gatunku pozostaje kontrowersyjna, gdyż F. novicida
umieszczona została na zatwierdzonej liście nazw bakterii, natomiast w publikacjach
pojawia się również jako podgatunek F. tularensis. Dlatego też podawane są dwie
różne nazwy i nadal nie ma jednoznacznej decyzji, dotyczącej prawidłowej no-

Rola katepsyn w zakażeniach bakteryjnych
67
menklatury [20]. Zakażenie F. novicida u ludzi występuje rzadko oraz jest trudne do
zdiagnozowania, natomiast bakteria ta jest wykorzystywana podczas badań in vitro lub
in vivo na organizmach modelowych takich jak myszy czy króliki jako odniesienie do
badania patomechanizmu tularemii wywoływanej przez F. tularensis.
Ostatnio przeprowadzone badania u myszy pozbawionych genu katepsyny B (Cstb-/-)
wykazały, że są one bardziej oporne na zakażenie F. novicida w porównaniu
z osobnikami o prawidłowej ekspresji tego białka. Ponadto, w trzeciej dobie po zaka-
żeniu u myszy z wyłączonym genem katepsyny B zaobserwowano obniżoną produkcję
cytokin prozapalnych oraz chemokin w wątrobie. Rolę katepsyny B zbadano także
w makrofagach, ponieważ odgrywają kluczową rolę podczas wewnątrzkomórkowej
replikacji F. novicida. Na podstawie otrzymanych wyników stwierdzono, że makro-
fagi, pochodzące od myszy pozbawionych katepsyny B, charakteryzowały się zwięk-
szoną aktywnością bakteriobójczą. Co ciekawe, podczas analizy struktur komór-
kowych w transmisyjnym mikroskopie elektronowym zaobserwowano, że niezakażone
makrofagi, pozbawione katepsyny B, wykazywały powiększoną pojedynczą błonę
lizosomalną oraz podwójną błonę autofagosomów w porównaniu do komórek typu
dzikiego. Autorzy badań twierdzą, że zwiększona aktywność bakteriobójcza w komór-
kach bez ekspresji katepsyny B wobec F. novicida może wynikać ze zwiększonej
biogenezy lizosomów, oraz tym samym, zwiększonej skuteczności transportu bakterii
do lizosomu w celu zwalczenia patogenu. Jednocześnie autorzy widzą możliwość
modulowania aktywności katepsyny w komórkach, co niesie za sobą potencjał
terapeutyczny i opcję leczenia osób z chorobą lizosomalną [21].
5. Rola katepsyny S w zakażeniach Mycobacterium bovis
Bakterie należące do rodzaju Mycobacterium są szeroko rozpowszechnione
w środowisku naturalnym. Niektóre gatunki tych bakterii, m.in. M. tuberculosis jest
najczęstszą przyczyną gruźlicy płucnej występującej u ludzi oraz u małp. M. bovis
natomiast wywołuje gruźlicę głównie u bydła, rzadziej u świń, psów, kotów czy koni.
Patogen może również być transmitowany na człowieka drogą bezpośredniego kon-
taktu z chorym zwierzęciem lub pośrednio po spożyciu mleka bądź jego przetworów
zawierających prątki [22, 23].
Katepsyna S należy do proteaz cysteinowych i znajduje się w lizosomach wielu
rodzajów komórek. Odgrywa zasadnicze znaczenie podczas dojrzewania cząsteczek
MHC klasy II, tzn. degradacji łańcucha Ii w endosomach, dzięki czemu dochodzi do
związania antygenu, transportu na powierzchnię komórki oraz prezentacji limfocytom
T CD4+ i uruchomieniu swoistej odpowiedzi immunologicznej [24, 25].
Dane literaturowe wskazują, ze zahamowanie ekspresji katepsyny S w makrofagach
zakażonych M. bovis było ściśle związane ze znaczną redukcją ekspresji dojrzałych
cząsteczek MHC klasy II, a więc jednocześnie zmniejszeniem efektywności wiązania
antygenów bakteryjnych. Ponadto, zaobserwowano zwiększoną pulę niedojrzałych
cząsteczek MHC klasy II na powierzchni tych komórek. Zaburzenie w ekspresji
prawidłowych, dojrzałych cząsteczek MHC klasy II na powierzchni makrofagów może
zatem tłumaczyć niepowodzenie stosowania szczepionek w celu wywołania skutecznej

Magdalena Bossowska-Nowicka, Felix N. Toka, Lidia Szulc-Dąbrowska
68
odporności przeciwko M. bovis [26]. Inne badania wykazały, że przywrócenie aktyw-
ności katepsyny S w makrofagach wpływa na zwiększenie ekspresji dojrzałych
cząsteczek MHC klasy II oraz późniejszą efektywną prezentację antygenów limfo-
cytom T, co stanowi istotny etap w tworzeniu nowego rodzaju szczepionek przeciwko
omawianemu patogenowi [27].
6. Podsumowanie
Rola poszczególnych katepsyn jest aktualnie intensywnie badana, zarówno
w zakażeniach bakteryjnych, jak i wirusowych. Bez wątpienia białka te pełnią istotne
funkcje w regulacji mechanizmów odpowiedzi immunologicznej. Ze względu na fakt,
iż jedną spośród wielu strategii mikroorganizmów do unikania odpowiedzi immuno-
logicznej i efektywnego namnażania się w organizmie gospodarza jest wykorzys-
tywanie katepsyn, mogą one stanowić ważny element w opracowaniu skutecznych
terapii bądź szczepionek w celu eliminacji patogenów.
Literatura
1. Collette J., Bocock J.P., Ahn K., Chapman R.L., Godbold G., Yeyeodu S., Erickson A.H. Biosynthesis and alternate targetingof the lysosomal cysteine protease cathepsin L. International Review of Cytology., 241 (2004), s. 1-51
2. Morin V., Sanchez A., Quiñones K., Huidobro J.G., Iribarren C., Bustos P., Puchi M., Genevière A.M., Imschenetzky M. Cathepsin L inhibitor I blocks mitotic chromosomes decondensation during cleavage cell cycles of sea urchin embryos. Journal of Cell Physiology., 216 (2008), s. 790-795.
3. Hsing L.C., Rudensky A.Y. The lysosomal cysteine proteases in MHC class II antigen presentation. Immunological Reviews., 207 (2005), s. 229-241.
4. Hsieh C.S., deRoos P., Honey K., Beers C., Rudensky A.Y. A role for cathepsin L and cathepsin S in peptide generation for MHC class II presentation. Journal of Immunology., 168 (2002), s. 2618-2625.
5. Li L., Gao L., Song Y., Qin Z.H., Liang Z. Activated cathepsin L is associated with the switch from autophagy to apoptotic death of SH-SY5Y cells exposed to 6-hydroxydopamine. Biochemical and Biophysical Research Communications., 470 (2016), s. 579-585.
6. Schornberg K., Matsuyama S., Kabsch K., Delos S., Bouton A., White J. Role of endosomal cathepsins in entry mediated by the Ebola virus glycoprotein. Journal of Virology., 80 (2006), s. 4174-4178.
7. Pager C.T., Dutch R.E. Cathepsin L is involved in proteolytic processing of the Hendra virus fusion protein. Journal of Virology., 79 (2005), s. 12714-12720.
8. Harman A.N., Kraus M., Bye C.R., Byth K., Turville S.G., Tang O., Mercier S.K., Nasr N., Stern J.L., Slobedman B., Driessen C., Cunningham A.L. HIV-1-infected dendritic cells show 2 phases of gene expression changes, with lysosomal enzyme activity decreased during the second phase. Blood., 114 (2009), s. 85-94.
9. Xu X., Greenland J.R., Gotts J.E., Matthay M.A , Caughey G.H. Cathepsin L Helps to Defend Mice from Infection with Influenza A. PLoS One., 11 (2016), e0164501.
10. Johnson E.M., Doyle J.D., Wetzel J.D., McClung R.P., Katunuma N., Chappell J.D., Washington M.K., Dermody T.S. Genetic and pharmacologic alteration of cathepsin expression influences reovirus pathogenesis. Journal of Virology., 83 (2009), s. 9630-9640.
11. Xiang X., Greenland J., Baluk P., Adams A., Bose O., McDonald D. M., Caughey G.H.. Cathepsin L Protects Mice from Mycoplasmal Infection and Is Essential for Airway Lymphangiogenesis. American Journal of Respiratory Cell and Molecular Biology., 49 (2013), s. 437-444.

Rola katepsyn w zakażeniach bakteryjnych
69
12. Nepal R.M., Mampe S., Shaffer B., Erickson A.H., Bryant P. Cathepsin L maturation and activity is impaired in macrophages harboring M. avium and M. tuberculosis. International Immunology., 18 (2006), s. 931-939.
13. Pires D., Marques J., Pombo J.P., Carmo N., Bettencourt P., Neyrolles O., Lugo-Villarino G., Anes E. Role of Cathepsins in Mycobacterium tuberculosis Survival in Human Macrophages. Scientific Reports., 6 (2016) s. 32247.
14. Tanaka Y., Tanaka R., Kawabata T., Noguchi Y., Himeno M. Lysosomal cysteine protease, cathepsin B, is targeted to lysosomes by the mannose 6-phosphate-independent pathway in rat hepatocytes: site-specific phosphorylation in oligosaccharides of the proregion. Journal of Biochemistry., 128 (2000), s. 39-48.
15. Tan G.J., Peng Z.K., Lu J.P., Tang F.Q. Cathepsins mediate tumor metastasis. World Journal of Biological Chemistry., 4 (2013), s. 91-101.
16. Korona-Głowniak I., Niedzielski A. Nosicielstwo Streptococcus pneumoniae a rozwój choroby pneumokokowej, Otorynolaryngologia., 12 (2013), s. 1-7.
17. Lawrence C.P., Kadioglu A., Yang A.L., Coward W.R., Chow S.C. The cathepsin B inhibitor, z-FA-FMK, inhibits human T cell proliferation in vitro and modulates host response to pneumococcal infection in vivo. Journal of Immunology., 177 (2006), 3827-3836.
18. Kubina R., Smycz M., Wojtyczka R.D., Kabała-Dzik A., Pacha J. Proces apoptozy w komorkach zainfekowanych bakteriami z rodzaju Chlamydia. Farmaceutyczny Przegląd Naukowy., 7-8 (2008), s. 9-11.
19. Bewley M.A., Marriott H.M., Tulone C., Francis S.E., Mitchell T.J., Read R.C., Chain B., Kroemer G., Whyte M.K., Dockrell D.H. A Cardinal Role for Cathepsin D in Co-Ordinating the Host-Mediated Apoptosis of Macrophages and Killing of Pneumococci. PLoS Pathogenes., 7 (2011), e1001262.
20. Huber B., Escudero R., Busse H.J., Seibold E., Scholz H.C., Anda P., Kämpfer P., Splettstoesser W.D. Description of Francisella hispaniensis sp. nov., isolated from human blood, reclassification of Francisella novicida (Larson et al. 1955) Olsufiev et al. 1959 as Francisella tularensis subsp. novicida comb. nov. and emended description of the genus Francisella. International Journal of Systematic and Evolutionary Microbiology., 60 (2010), s. 1887-1896.
21. Qi X., Man S.M., Malireddi R.K., Karki R., Lupfer C., Gurung P., Neale G., Guy C.S., Lamkanfi M., Kanneganti T.D. Cathepsin B modulates lysosomal biogenesis and host defense against Francisella novicida infection. The Journal of Experimental Medicine., 213 (2016), s. 2081-2097.
22. Bagieńska M., Rzewuska M. Występowanie prątków z kompleksu Mycobacterium tuberculosis u zwierząt – transmisja wybranych gatunków między ludźmi a zwierzętami. Życie Weterynaryjne., 85 (2010), s. 742-746.
23. Sweetline A.N., Ronald B.S., Kumar T.M., Kannan P., Thangavelu A. Molecular identification of Mycobacterium tuberculosis in cattle. Veterinary Microbiology., 198 (2017), s. 81-87.
24. Shi G.P., Villadangos J.A., Dranoff G., Small C., Gu L., Haley K.J., Riese R., Ploegh H.L., Chapman H.A. Cathepsin S required for normal MHC class II peptide loading and germinal center development. Immunity., 10 (1999), s. 197-206.
25. Bania, J., Gatti E., Lelouard H., David A., Cappello F., Weber E., Camosseto V., Pierre P. Human cathepsin S, but not cathepsin L, degrades efficiently MHC class II-associated invariant chain in nonprofessional APCs. Proceedings of the National Academy of Science of the United States of America., 100 (2003), s. 6664–6669.
26. Sendide K., Deghmane A. E., Pechkovsky D., Av-Gay Y., Talal A., Hmama Z. Mycobacterium bovis BCG attenuates surface expression of mature class II molecules through IL-10-dependent inhibition of cathepsin S. Journal of Immunology., 175 (2005), s. 5324–5332.

Magdalena Bossowska-Nowicka, Felix N. Toka, Lidia Szulc-Dąbrowska
70
27. Soualhine H., Deghmane A.E., Sun J., Mak K., Talal A., Av-Gay Y., Hmama Z. Mycobacterium bovis bacillus Calmette-Guérin secreting active cathepsin S stimulates expression of mature MHC class II molecules and antigen presentation in human macrophages. Journal of Immunology., 179 (2007), s. 5137-5145.
Rola katepsyn w zakażeniach bakteryjnych
Streszczenie Katepsyny odgrywają istotną rolę w funkcjonowaniu układu odpornościowego. Enzymy te biorą udział w przetwarzaniu i prezentacji antygenu, adhezji i migracji komórek oraz degradacji wewnątrzkomórko-wych bakterii w lizosomach, warunkując wrodzoną i nabytą odpowiedź immunologiczną na wiele zakażeń bakteryjnych. Wykazano, że myszy z niedoborem katepsyny L wykazują większą wrażliwość na zakażenie Mycoplasma pulmonis oraz osłabioną produkcję TNF-α i chemokin. Podobnie myszy z genomową delecją katepsyny E są bardziej wrażliwe na zakażenie Staphylococcus aureus i Porphyromonas gingivalis. W mysich komór-kach dendrytycznych, zakażonych Chlamydia psittaci obserwuje się kolokalizację patogenu z przedziałami komórkowymi, zawierającymi katepsynę D – krytyczny enzym w procesie autofagii. Ponadto makrofagi myszy pozbawione katepsyny D wykazują zmniejszoną zdolność do zabijania bakterii podczas zakażenia pneumokokami. Natomiast myszy pozbawione aktywności katepsyny B są bardziej oporne na zakażenie Francisella novicida. Co więcej Mycobacterium bovis wykorzystuje indukcję wytwarzania IL-10 do zahamowania przetwarzania łańcucha niezmiennego (Ii) cząsteczek MHC klasy II, zależnego od katepsyny S w makrofagach ludzkich, co skutkuje zmniejszoną prezentacją peptydów bakteryjnych limfocytom T CD4+. Taka zdolność może stanowić skuteczną strategię patogenu do unikania odpowiedzi immunolo-gicznej i efektywnego namnażania się w organizmie gospodarza. Mimo licznych doniesień naukowych, udział katepsyn w zakażeniach bakteryjnych nadal jest słabo pozna-ny i wymaga dalszych badań. Praca sfinansowana ze środków dotacji KNOW Konsorcjum „Zdrowe Zwierzę – Bezpieczna Żywność” nr 05-1/KNOW2/2015 MNiSW i grantu nr UMO-2012/05/D/NZ6/02916 NCN. Słowa kluczowe: katepsyny, zakażenia bakteryjne, strategie ucieczki immunologicznej
Role of cathepsins in bacterial infections
Abstract Cathepsins play an important role in the functioning of the immune system. They are involved in antigen processing and presentation, cell adhesion and migration or degradation of intracellular bacteria in lysoso-mes, thereby determine an innate and adaptive immune response to many bacterial infections. It has been shown that cathepsin L-deficient mice exhibit greater susceptibility to Mycoplasma pulmonis infection, and decreased production of TNF-α and chemokines. Similarly, cathepsin E-deficient mice show increased susceptibility to infection with Staphylococcus aureus and Porphyromonas gingivalis. In murine dendritic cells infected with Chlamydia psittaci, colocalization between this pathogen and compartments containing cathepsin D – a critical enzyme in autophagy, is observed. Moreover, cathepsin D-deficient mu-rine macrophages show reduced ability to kill bacteria during pneumococcal infection. In contrast, mice lacking cathepsin B activity exhibited increased resistance to Francisella novicida. Furthermore, Myco-bacterium bovis exploits IL-10 induction to inhibit cathepsin S-dependent processing of MHC class II degradation of the invariant chain (Ii) in human macrophages, which result in reduced presentation of bac-terial peptides to CD4+ T cells. Such ability may represent an effective pathogen strategy to avoid immune response and persist in the host. Despite numerous scientific reports, the participation of cathepsins in antibacterial host defense is still largely unknown and requires further investigation. Work funded by KNOW Scientific Consortium "Healthy Animal - Safe Food", No. 05-1/KNOW2/2015" from Ministry of Science and Higher Education and grant No. UMO-2012/05/D/NZ6/02916 from National Science Center. Keywords: cathepsins, bacterial infections, immune evasion strategies

71
Adriana Pacia1, Małgorzata Nabrdalik
2, Agnieszka Dołhańczuk-Śródka
3
Ocena aktywności przeciwdrobnoustrojowej olejku geraniowego
1. Wprowadzenie
Antyseptyczne właściwości roślin leczniczych jak i ich wyciągów zauważone i wykorzystywane były już w czasach antycznych, natomiast próby ich zbadania w laboratoriach datowane są na początek XX wieku [1]. Lotne olejki eteryczne są pozyskiwane głównie z niezdrewniałych części roślin, takich jak: liście, kwiaty, pąki czy łodygi, najczęściej za pomocą metody destylacji z parą wodną. W związku z nadu-żywaniem przez medycynę nowożytną substancji o działaniu mikrobójczym, w ostatnich latach odnotowuje się wzrastającą liczbą bakteryjnych i grzybiczych szczepów opornych na stosowane dotąd antybiotyki. Spowodowało to zainteresowanie naturalnymi substancjami antymikrobowymi i antyoksydacyjnymi, pomagającymi kontrolować choroby zarówno ludzi, jak i roślin. Olejki eteryczne, dzięki swoim bogatym składom chemicznym, mają wiele biologicznych właściwości. Mogą być użyte prewencyjnie, jak i leczniczo w terapiach różnych chorób układowych. W niektórych takich przypadkach możliwe jest ustalenie terapii dodatkowej w postaci podawania mieszaniny kilku olejków eterycznych lub ich połączenia z antybiotykiem. W przemyśle spożywczym przeciwutleniające substancje zawarte w olejkach hamują szkodliwą działalność oksydacyjną produktów spożywczych, a także zapobiegają ich zakażeniom [2, 3].
Przykładem roślin wykazujących powyższe właściwości są należące do rodziny Geraniaceae (Bodziszkowate) rośliny rodzaju Pelargonium (Pelargonia). Rodzaj Pelargonium, liczący około 250 gatunków, wywodzi się głównie z Południowej Afryki, gdzie zajmuje dużą część naturalnej flory. Pelargonie występują również między innymi w: północnej i wschodniej Afryce, Australii, Nowej Zelandii, Madaga-skarze, wyspach Oceanu Atlantyckiego czy na Bliskim Wschodzie. Największym producentem olejku geraniowego są natomiast Chiny, zaraz za nimi Egipt, Algieria, Maroko i Reunion (Ocean Indyjski). Najbardziej znanymi przedstawicielami rodzaju Pelargonium są: P. graveolens, P. odoranissimum, P. roseum, P. reniforme, P. sidoides czy P. zonale [2, 4-7].
Olejek geraniowy działa wielokierunkowo: przeciwgrzybiczo, przeciwbakteryjnie, przeciwwirusowo, przeciwbólowo, przeciwzapalnie, a podany doustnie wykazuje właściwości uspokajające, wykrztuśne, żółciopędne oraz moczopędne. Ponadto reguluje ciśnienie krwi i łagodzi także zaburzenia związane z menopauzą. Wykazano,
1 [email protected], Samodzielna Katedra Biotechnologii i Biologii Molekularnej, Wydział Przyrodniczo-
Techniczny, Uniwersytet Opolski, www.uni.opole.pl 2 [email protected], Samodzielna Katedra Biotechnologii i Biologii Molekularnej, Wydział Przyrod-
niczo-Techniczny, Uniwersytet Opolski, www.uni.opole.pl 3 [email protected], Samodzielna Katedra Biotechnologii i Biologii Molekularnej, Wydział Przyrodniczo-
Techniczny, Uniwersytet Opolski, www.uni.opole.pl

Adriana Pacia, Małgorzata Nabrdalik, Agnieszka Dołhańczuk-Śródka
72
że pomocny jest w leczeniu: zakażeń błony śluzowej jamy ustnej, stanu zapalnego języka czy związanego z użytkowaniem protez zębowych, a także czerwonki, bie-gunki, wrzodów żołądka, cukrzycy czy nowotworów. Może być stosowany pomoc-niczo w terapii chorób i zaburzeń skórnych, takich jak: owrzodzenia, egzemy, opa-rzenia, trudno gojące się rany, trądzik, cellulit, a także łupież. Co więcej, dzięki zawar-tości głównie cytronelolu i geraniolu olejek geraniowy skutecznie odstrasza owady, w tym także komary [2, 3, 8-12].
Pelargonie są cenione ze względu na pachnące różą liście, z cytrusową bądź mięto-wą nutą. Wykorzystywane jest to w przemyśle kosmetycznym, gdzie najwyższej jako-ści olejek eteryczny – olejek Bourbon (z Reunion), używany jest w perfumerii jako tańszy zamiennik olejku różanego, a pozostałe olejki geraniowe znajdują zastosowanie w różnego rodzaju kosmetykach: mydłach, kremach, wodach toaletowych, a także w aromaterapii [6, 13-14].
Głównymi składnikami, którym przypisuje się powyższe właściwości lecznicze i pielęgnacyjne olejku, są: cytronelol, geraniol, linalol, izomenton, nerol oraz mrów-czan cytronelolu. Zawartość poszczególnych substancji różni się w roślinach w zależ-ności od gatunku, pochodzenia i warunków, w jakich rosły. Ilości poszczególnych sub-stancji aktywnych w pelargoniach wahają się w granicach: 12,4-38,2% cytronelol, 5,6-36,0% geraniol, 2,8-21,8% linalol, 0,3-8,4% izomenton, 1,2-8,7% nerol i 0,1-16,9% mrówczanu cytronelolu [2, 10, 15, 16].
Celem przeprowadzonych badań była ocena właściwości przeciwdrobnoustro-jowych olejku geraniowego i możliwość użycia go jako naturalnej substancji konser-wującej w preparatach kosmetycznych.
2. Materiały i metody
Do badań użyty został komercyjny geraniowy olejek eteryczny firmy Avicenna-Oil, pozyskany z Pelargonium odoranissimum. Aktywność przeciwdrobnoustrojową olejku określono wobec referencyjnych szczepów bakterii: Escherichia coli (ATCC 10536), Staphylococcus aureus (ATCC 25923), Pseudomonas aeruginosa (ATCC 27853) oraz grzybów Candida albicans (ATCC 10231). Obecność powyższych gatunków jest zabroniona we wszystkich produktach kosmetycznych [17, 18], dlatego działanie przeciwdrobnoustrojowe badanego olejku eterycznego świadczyłaby o potencjalnej możliwości wykorzystania go jako naturalnego środka konserwującego w kosme-tykach.
W przeprowadzonych badaniach wykorzystano metody: seryjnych rozcieńczeń w bulionie, seryjnych rozcieńczeń w agarze oraz metodę dyfuzyjno-krążkową. Posłużyły one do wyznaczenia wartości minimalnego stężenia hamującego MIC (ang. Minimal Inhibitory Concentration) oraz minimalnego stężenia bakterio- i grzybobój-czego MBC/MFC (ang. Minimal Bactericidal/Fungicidal Concentration).
Wszystkie hodowle prowadzono na komercyjnych podłożach: bakteryjne na podłożu wzbogaconym – Mueller-Hinton (producent BioMaxima), grzybicze na podłożu YPD – Yeast extract Peptone Dextrose (producent BioMaxima). Inkubację bakterii prowadzono w temperaturze 37°C przez 24 h, a drożdży w 30°C/48 h.

Ocena aktywności przeciwdrobnoustrojowej olejku geraniowego
73
Przygotowano inokulum badanych szczepów o gęstości 0,5 w skali McFarlanda poprzez zawieszenie kolonii w płynie fizjologicznym z dodatkiem Tweenu 80. Tak przygotowane zawiesiny wykorzystano w kolejnych etapach badań.
2.1. Metoda seryjnych rozcieńczeń w bulionie
Przygotowano roztwór olejku geraniowego w etanolu w stężeniach: 0,25; 0,5; 1,0; 2,5; 5,0 oraz 10,0 µl/ml. Dla bakterii użyto bulionu Mueller-Hinton, a dla szczepu drożdży bulionu YPD. Do tak przygotowanych podłoży dodano po 100 µl inokulum badanych mikroorganizmów. Kontrole wzrostu szczepów stanowiły podłoża bulio-nowe bez dodatku olejku (K) oraz podłoża bulionowe z dodatkiem alkoholu (K-). W kontrolach pozytywnych oceniających skuteczność olejku (K+) zastosowano gentamycynę dla szczepów bakterii oraz nystatynę dla drożdży. Próby badane i kontrolne poddano inkubacji. Po 24 h wykonano serię rozcieńczeń i posiano na odpo-wiednie podłoża mikrobiologiczne. Obliczono ilość jednostek tworzących kolonie w 1 ml hodowli [jtk/ml]. Za wartość MBC/MFC przyjęto stężenie olejku, przy którym następuje zmniejszenie liczby żywych komórek drobnoustroju o 99,9% w stosunku do próby kontrolnej K-.
2.2. Metoda seryjnych rozcieńczeń w agarze
Do upłynnionego podłoża Mueller-Hinton oraz YPD o temperaturze 45˚C wpro-wadzono roztwór olejku geraniowego w etanolu, w ilości odpowiadającej wyzna-czonym stężeniom (0,25-10,0 µl/ml) oraz po 100 µl odpowiedniego inokulum z drobnoustrojami. Po wymieszaniu, materiał rozlano na sterylne szalki Petriego, pozostawiono do zastygnięcia i inkubowano w odpowiednich warunkach. Wykonano również próby kontrolne analogicznie jak poprzednio. Po inkubacji określono wzrost na podłożu i wyznaczono wartość MIC.
2.3. Metoda dyfuzyjno-krążkowa
Na stałe podłoże Mueller-Hinton oraz YPD naniesiono i rozprowadzono po 100 µl zawiesiny badanych bakterii i grzybów. Następnie ułożono sterylne krążki bibułowe (średnica Ø 5,0 mm), które nasączono 10 µl alkoholowych roztworów olejku geraniowego w różnych stężeniach (0,25-10,0 µl/ml), sterylnej wody (K) oraz alkoholu (K
-). W przypadku kontroli pozytywnej (K+) użyto gotowych krążków z antybio-
tykiem (gentamycyna/nystatyna). Oznaczenia przeprowadzono w trzech powtórze-niach. Płytki inkubowano przez 24 h, a następnie zmierzono strefy zahamowania wzrostu mikroorganizmów [mm].
3. Wyniki
3.1. Seryjne rozcieńczenia w bulionie
W Tabeli 1 zebrano i przedstawiono wyniki przeprowadzonych badań. Wykazały one, że liczba bakterii i drożdży w próbie kontrolnej była różna, a obecność alkoholu w podłożu nieznacznie ograniczyła ich wzrost. Zastosowane antybiotyki całkowicie zahamowały wzrost badanych szczepów mikroorganizmów. W niższych stężeniach alkoholowych roztworów olejków, w zakresie 0,25-1,0 µl/ml, zaobserwowano nieznaczną stymulację wzrostu szczepów E. coli oraz C. albicans, a w przypadku S.
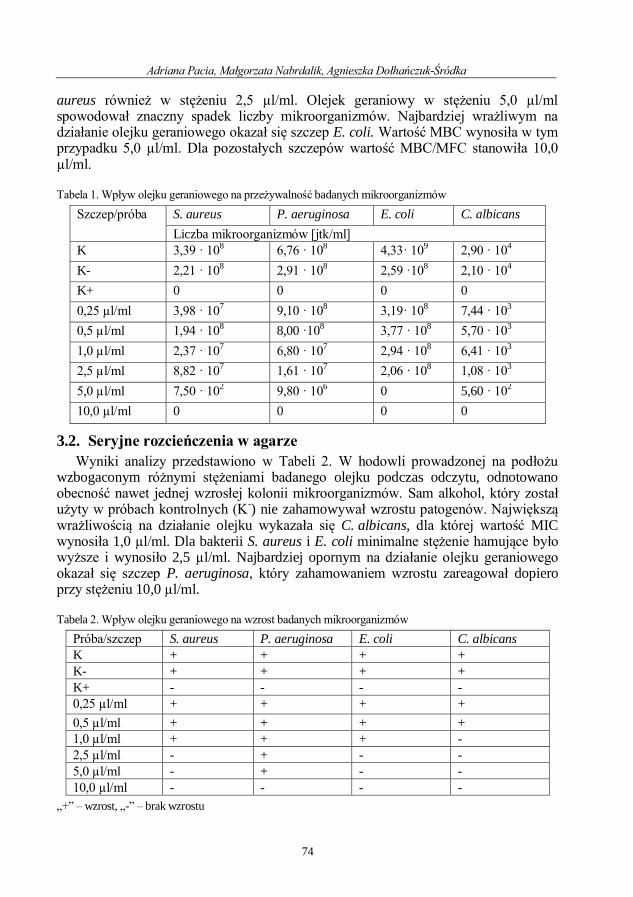
Adriana Pacia, Małgorzata Nabrdalik, Agnieszka Dołhańczuk-Śródka
74
aureus również w stężeniu 2,5 µl/ml. Olejek geraniowy w stężeniu 5,0 µl/ml spowodował znaczny spadek liczby mikroorganizmów. Najbardziej wrażliwym na działanie olejku geraniowego okazał się szczep E. coli. Wartość MBC wynosiła w tym przypadku 5,0 µl/ml. Dla pozostałych szczepów wartość MBC/MFC stanowiła 10,0 µl/ml.
Tabela 1. Wpływ olejku geraniowego na przeżywalność badanych mikroorganizmów
Szczep/próba S. aureus P. aeruginosa E. coli C. albicans
Liczba mikroorganizmów [jtk/ml]
K 3,39 · 108 6,76 · 10
8 4,33· 10
9 2,90 · 10
4
K- 2,21 · 108 2,91 · 10
8 2,59 ·10
8 2,10 · 10
4
K+ 0 0 0 0
0,25 µl/ml 3,98 · 107 9,10 · 10
8 3,19· 10
8 7,44 · 10
3
0,5 µl/ml 1,94 · 108 8,00 ·10
8 3,77 · 10
8 5,70 · 10
3
1,0 µl/ml 2,37 · 107 6,80 · 10
7 2,94 · 10
8 6,41 · 10
3
2,5 µl/ml 8,82 · 107 1,61 · 10
7 2,06 · 10
8 1,08 · 10
3
5,0 µl/ml 7,50 · 102 9,80 · 10
6 0 5,60 · 10
2
10,0 µl/ml 0 0 0 0
3.2. Seryjne rozcieńczenia w agarze
Wyniki analizy przedstawiono w Tabeli 2. W hodowli prowadzonej na podłożu wzbogaconym różnymi stężeniami badanego olejku podczas odczytu, odnotowano obecność nawet jednej wzrosłej kolonii mikroorganizmów. Sam alkohol, który został użyty w próbach kontrolnych (K
-) nie zahamowywał wzrostu patogenów. Największą
wrażliwością na działanie olejku wykazała się C. albicans, dla której wartość MIC wynosiła 1,0 µl/ml. Dla bakterii S. aureus i E. coli minimalne stężenie hamujące było wyższe i wynosiło 2,5 µl/ml. Najbardziej opornym na działanie olejku geraniowego okazał się szczep P. aeruginosa, który zahamowaniem wzrostu zareagował dopiero przy stężeniu 10,0 µl/ml.
Tabela 2. Wpływ olejku geraniowego na wzrost badanych mikroorganizmów
Próba/szczep S. aureus P. aeruginosa E. coli C. albicans
K + + + +
K- + + + +
K+ - - - -
0,25 µl/ml + + + +
0,5 µl/ml + + + +
1,0 µl/ml + + + -
2,5 µl/ml - + - -
5,0 µl/ml - + - -
10,0 µl/ml - - - -
„+” – wzrost, „-” – brak wzrostu
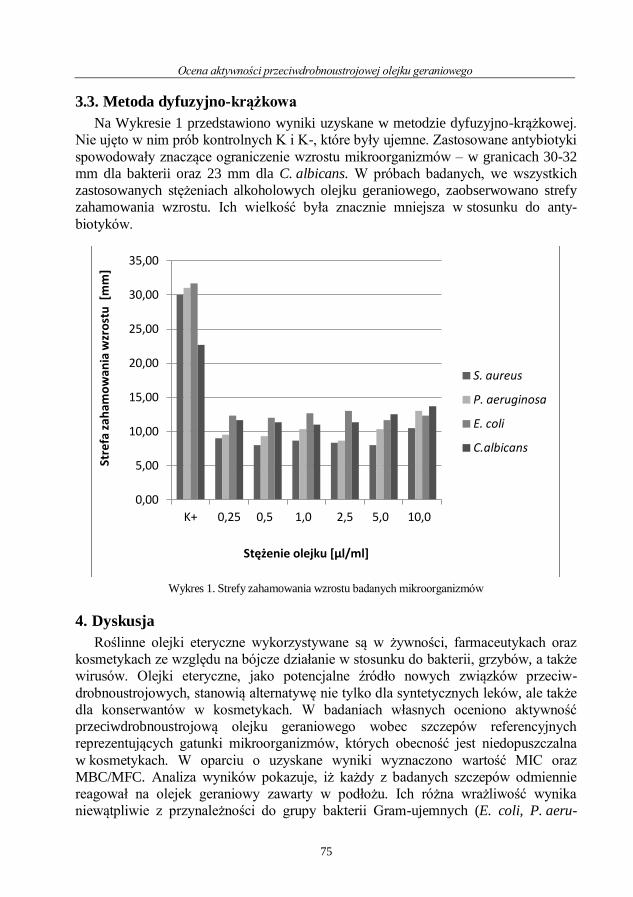
Ocena aktywności przeciwdrobnoustrojowej olejku geraniowego
75
3.3. Metoda dyfuzyjno-krążkowa
Na Wykresie 1 przedstawiono wyniki uzyskane w metodzie dyfuzyjno-krążkowej.
Nie ujęto w nim prób kontrolnych K i K-, które były ujemne. Zastosowane antybiotyki
spowodowały znaczące ograniczenie wzrostu mikroorganizmów – w granicach 30-32
mm dla bakterii oraz 23 mm dla C. albicans. W próbach badanych, we wszystkich
zastosowanych stężeniach alkoholowych olejku geraniowego, zaobserwowano strefy
zahamowania wzrostu. Ich wielkość była znacznie mniejsza w stosunku do anty-
biotyków.
Wykres 1. Strefy zahamowania wzrostu badanych mikroorganizmów
4. Dyskusja
Roślinne olejki eteryczne wykorzystywane są w żywności, farmaceutykach oraz
kosmetykach ze względu na bójcze działanie w stosunku do bakterii, grzybów, a także
wirusów. Olejki eteryczne, jako potencjalne źródło nowych związków przeciw-
drobnoustrojowych, stanowią alternatywę nie tylko dla syntetycznych leków, ale także
dla konserwantów w kosmetykach. W badaniach własnych oceniono aktywność
przeciwdrobnoustrojową olejku geraniowego wobec szczepów referencyjnych
reprezentujących gatunki mikroorganizmów, których obecność jest niedopuszczalna
w kosmetykach. W oparciu o uzyskane wyniki wyznaczono wartość MIC oraz
MBC/MFC. Analiza wyników pokazuje, iż każdy z badanych szczepów odmiennie
reagował na olejek geraniowy zawarty w podłożu. Ich różna wrażliwość wynika
niewątpliwie z przynależności do grupy bakterii Gram-ujemnych (E. coli, P. aeru-
0,00
5,00
10,00
15,00
20,00
25,00
30,00
35,00
K+ 0,25 0,5
1,0
2,5 5,0
10,0
Stre
fa z
ah
amo
wan
ia w
zro
stu
[m
m]
Stężenie olejku [µl/ml]
S. aureus
P. aeruginosa
E. coli
C.albicans

Adriana Pacia, Małgorzata Nabrdalik, Agnieszka Dołhańczuk-Śródka
76
ginosa), Gram-dodatnich (S. aureus) oraz drożdży (C. albicans). Olejek geraniowy,
w zależności od szczepu, wykazywał działanie bakteriostatyczne (w zakresie 2,5-10,0
µl/ml) oraz bakteriobójcze (w zakresie 5,0-10,0 µl/ml). W przypadku C. albicans
działanie grzybostatyczne uzyskano przy stężeniu 1,0 µl/ml, a grzybobójcze 10 µl/ml.
W badaniach własnych, w celu określenia wartości MIC, posłużono się między innymi
metodą dyfuzyjno-krążkową. Jednak uzyskane wielkości stref zahamowania wzrostu
były do siebie zbliżone i znacznie niższe w porównaniu do kontroli (5,0-10,0 mm).
Przypuszcza się, że zastosowany olejek geraniowy charakteryzuje się niską rozpusz-
czalnością w alkoholu lub/oraz niską szybkością dyfuzji do podłoża agarowego.
Dlatego planowane są dalsze badania w zakresie modyfikacji metody dyfuzyjno-
krążkowej, co pozwoli na lepsze uwalnianie badanych substancji do pożywki.
Wysoką skuteczność olejku geraniowego opisuje wielu badaczy. W przeprowadzo-
nej przez Bigos i wsp. analizie wykazano, że wartość MIC alkoholowych roztworów
olejku geraniowego wobec S. aureus była dziesięciokrotnie niższa niż w badaniach
własnych i wynosiła 1,0 µl/ml. Wartość MIC dla większości badanych szczepów była
niższa lub równa 1,0 µl/ml [2]. Niemniej jednak w opozycji znajdują się dane
w Kędzia i wsp., gdzie stężenie olejku geraniowego (roztwór DMSO), które hamowało
wzrost wzorcowego szczepu S. aureus wynosiło aż 100 µg/ml. Badany olejek geranio-
wy nie wykazywał silnego działania wobec drobnoustrojów takich jak chorobotwórcza
E. coli, P. aeruginosa i C. albicans [8]. Z kolei w badaniach Prabuseenivasan i wsp.
olejek geraniowy, rozcieńczony w DMSO, charakteryzował się wysoką skutecznością
wobec E. coli – wartość MIC ˃6,4 mg/ml. Natomiast dla S. aureus oraz P. aeruginosa
wartość MIC była dwukrotnie wyższa i wynosiła ˃12,8 mg/ml [19]. Wyniki badań
własnych są zbliżone. Znacznie wyższe wartości uzyskali Andrade i wsp. W badaniach
tych wartość MIC dla E. coli stanowiła 100 µl/ml, a dla S. aureus 200 µl/ml [20].
Również w próbach Sienkiewicz i wsp., szczep E. coli oraz dodatkowo P. aeruginosa
były najbardziej wrażliwe na działanie olejku geraniowego. Wartość MIC dla klini-
cznych szczepów E. coli była nieznacznie wyższa niż w badaniach własnych i mieściła
się w przedziale 3,0-3,75 µl/ml, a dla P. aeruginosa, podobnie jak w badaniach
własnych, w granicach 9,25-10,5 µl/ml [11]. Odmienne dane prezentują Oulkheir
i wsp., którzy analizując marokański olejek geraniowy nie zaobserwowali hamującego
działania olejku wobec Gram-dodatnich i Gram-ujemnych bakterii [21]. Autorzy
cytowanych prac nie podają wartości MBC/MFC olejku geraniowego wobec badanych
szczepów [2, 8, 11, 19-21].
Analizują wyniki doniesień naukowych można stwierdzić, że wpływ na skutecz-
ność badanych substancji ma wykorzystanie różnych szczepów danego gatunku.
Szczepy różnić się będą opornością na antybiotyki/chemioterapeutyki i inne środki, co
bezpośrednio wynika z ich zjadliwości. Dlatego należy wziąć pod uwagę to, że na
aktywność olejków eterycznych ma wpływ ich pochodzenie, skład chemiczny, a także
zastosowany rozcieńczalnik.

Ocena aktywności przeciwdrobnoustrojowej olejku geraniowego
77
5. Wnioski
Przedstawione wyniki badań potwierdzają naukowe doniesienia na temat wysokiej
aktywności przeciwdrobnoustrojowej geraniowego olejku eterycznego.
Użyta metoda seryjnych rozcieńczeń w agarze dostarczyła informacji, że najwięk-
szą wrażliwość na sporządzony alkoholowy roztwór olejku wykazał szczep C.
albicans, wrażliwe na działanie olejku były też szczepy E. coli i S. aureus, a najmniej
P. aeruginosa. Wyniki metody seryjnych rozcieńczeń w bulionie wykazały, że za
minimalne stężenie bójcze MBC/MFC wobec badanych szczepów należy uznać
stężenie olejku 10,0 µl/ml. Odnosząc się do metody dyfuzyjno-krążkowej, jej przydat-
ność wydaje się ograniczona ze względu na nierównomierne wnikanie w agar olejku.
Brak stref zahamowania wzrostu nie wynika z oporności badanego szczepu na działa-
nie testowanego związku, ale z ograniczeń związanych z dyfuzją.
Ze względu na wysoką skuteczność olejku geraniowego wobec testowanych mikro-
organizmów powinien on sprawdzić się jako skuteczny konserwant, zapobiegający
rozwojowi w preparatach kosmetycznych patogennych szczepów E. coli, S. aureus. P.
aeruginosa oraz C. albicans.
Powyższe wyniki pozostawiają otwartą furtkę do dalszych badań nad właści-
wościami geraniowego olejku eterycznego i pozwalają przypuszczać, że dzięki
aktywności przeciwbakteryjnej i przeciwgrzybowej może on stanowić alternatywę dla
chemicznych konserwantów. W toku dalszych badań należy również sprawdzić jak
omawiany olejek zachowa się w gotowym produkcie kosmetycznym.
Literatura
1. Dorman H.J., Deans S.G. Antimicrobial agents from plants: antibacterial activity of plant
volatile oils, Journal of Applied Microbiology., 88 (2000), s. 308-316.
2. Bigos M., Wasiela M., Kalemba D., Sienkiewicz M. Antimicrobial Activity of Geranium
Oil against Clinical Strains of Staphylococcus aureus, Molecules., 17 (2012), s. 10276-
10291.
3. Hsouna A. B., Hamdi N. Phytochemical composition and antimicrobial activities of the
essential oils and organic extracts from Pelargonium graveolens growing in Tunisia,
Lipids in Health and Disease., 11 (2012).
4. Lalli J.Y.Y., Viljoen A.M., Ba¸ser K. Hüsnü C., Demirci B., Özek T. The Essential Oil
Composition and Chemotaxonomical Appraisal of South African Pelargoniums
(Geraniaceae), Journal of Essential Oil Research., 18 (2006), s. 89-105.
5. Romitelli I., Martins M.B.G. Comparison of leaf morphology and anatomy among Malva
sylvestris (“gerânioaromático”), Pelargonium graveolens (“falsa-malva”) and
Pelargonium odoratissimum (“gerânio-de-cheiro”), Revista Brasileira de Plantas
Medicinais., 15 (2013), s. 91-97.
6. Juliani H.R., Koroch A., Simon J.E., Hitimana N., Daka A., Ranarivelo L., Langenhoven P.
Quality of Geranium Oils (Pelargonium Species): Case Studies in Southern and Eastern
Africa, Journal of Essential Oil Research., 18 (2006), s. 116-121.
7. Lancu, C.E., Cioanca, O., Mircea, C., Hăncianu, M. Contributions regarding leaf histo-
anatomy of some Pelargonium species. Rev. Med. Chir. Soc. Med. Nat. Iași. 117 (2013),
s. 812-818.

Adriana Pacia, Małgorzata Nabrdalik, Agnieszka Dołhańczuk-Śródka
78
8. Kędzia B., Hołdera-Kędzia E. Badanie wpływu olejków eterycznych na bakterie, grzyby
i dermatofity chorobotwórcze dla człowieka, Postępy Fitoterapii., 2 (2007), s. 71-77.
9. Kędzia A. Przeciwgrzybicze działanie olejku geraniowego (Oleum Geranii), Postępy
Fitoterapii., 4 (2009), s. 217-221.
10. Boukhris M., Simmonds M.J., Sayadi S., Bouaziz M. Chemical Composition and
Biological Acitivities of Polar Extracta and Essential Oil of Rose-scented Geranium,
Pelargonium graveolens, Phytotherapy Research., 27 (2013), s. 1206-1213.
11. Sienkiewicz M., Poznańska-Kurowska K., Kaszuba A., Kowalczyk E. The antibacterial
activity of geranium oil against Gram-negative bacteria isolated from difficult-to-heal
wounds, Burns., 40 (2014), s. 1046-1051.
12. Džamić A.M., Soković M.D., Ristić M.S. , Grujić S.M., Mileski K.S. Marin P.D. Chemical
composition, antifungal and antioxidant activity of Pelargonium graveolens essential oil,
Journal of Applied Pharmaceutical Science., 4 (2014), s. 1-5.
13. Ravindra N.S., Kulkarni R.R.N., Gayathri M.C., Ramesh S. Somaclonal variation for some
morphological traits, herb yield, essential oil content and essential oil composition in an
Indian cultivar of rose-scented geranium, Plant Breeding., 2004 (123), s. 84-86.
14. Motsa N.M., Soundy P., Steyn J.M., Robin A., Mojela L. Mojela N. Teubes C. Plant Shoot
Age and Temperature Effects on Essential Oil Yield and Oil Composition of Rose-Scented
Geranium (Pelargonium sp.), Journal of Essential Oil Research., 18 (2006), s. 106-110.
15. Cerempei A., Muresan E.I., Cimpoesu N. Biomaterials with controlled release of geranium
essential oil, Journal of Essential Oil Research., 2014 (4), s. 267-273.
16. Hitimana N., Daka A., Ranarivelo L., Langenhoven P. Quality of Geranium Oils
(Pelargonium Species): Case Studies in Southern and Eastern Africa, Journal of Essential
Oil Research., 2006 (18), s. 116-121.
17. Rozporządzenie Ministra Zdrowia z dnia 23 grudnia 2002 r. w sprawie określenia procedur
pobierania próbek kosmetyków oraz procedur przeprowadzania badań laboratoryjnych
(Dz.U. 2003 nr 9 poz. 107).
18. Rozporządzenie Ministra Zdrowia z dnia 13 listopada 2015 r. w sprawie jakości wody
przeznaczonej do spożycia przez ludzi (Dz.U. 2015 poz. 1989).
19. Prabuseenivasan S, Jayakumar M., Ignacimuthu S. In vitro antibacterial activity of some
plant essential oils, BMC Complementary and Alternative Medicine, 2006 (6).
20. Andrade M.A., Cardoso
M.G., Batista
L.R., Freire
J.M., Nelson D.L. Antimicrobial activity
and chemical composition of essential oil of Pelargonium odoratissimum, Revista
Brasileira de Farmacognosia., 2011 (21).
21. Oulkheir S., Aghrouch M., Mourabit F. El., Dalha F., Graich H., Amouch F., Ouzaid K.,
Moukale A., Chadli S. Antibacterial Activity of Essential Oils Extracts from Cinnamon,
Thyme, Clove and Geranium Against a Gram Negative and Gram Positive Pathogenic
Bacteria, Journal of Diseases and Medicinal Plants., 2017 (3), s. 1-5.

Ocena aktywności przeciwdrobnoustrojowej olejku geraniowego
79
Ocena aktywności przeciwdrobnoustrojowej olejku geraniowego
Streszczenie
Olejki eteryczne od lat poddaje się badaniom poszukując w nich alternatywy dla syntetycznych środków
konserwujących w kosmetykach. Olejek geraniowy dzięki zawartości głównie geraniolu i cytronelolu
stanowi potencjalnie właśnie taką substancję. Celem przeprowadzonych badań było określenie właściwości przeciwdrobnoustrojowych olejku geraniowego. W pracy wykorzystano komercyjny olejek (Avicenna-
Oil). Oceniono aktywność przeciwdrobnoustrojową etanolowego roztworu olejku w zakresie stężeń od
0,25 do 10,0 µl/ml. Materiał biologiczny stanowiły szczepy referencyjne z kolekcji ATCC: Escherichia
coli, Staphylococcus aureus, Pseudomonas aeruginosa oraz Candida albicans. Do badań wrażliwości szczepów na olejek wykorzystano metodę seryjnych rozcieńczeń w agarze oraz zmodyfikowaną metodę
seryjnych rozcieńczeń w bulionie. Na ich podstawie wyznaczono watość MIC oraz MBC/MFC.
W przeprowadzonych badaniach olejek geraniowy wykazywał dużą aktywność. Największą wrażliwość na
olejek wykazywał szczep C. albicans, gdzie wartość MIC wynosiła 0,1 µl/ml. Natomiast dla E. coli i S. aureus wartość ta była wyższa (0,25 µl/ml). Najniższą wrażliwością na olejek okazał szczep P. aeruginosa
(MIC 10 µl/ml). Za minimalne stężenie bójcze MBC/MFC wobec badanych szczepów należy uznać
stężenie olejku 10,0 µl/ml. Znacząca aktywnością olejku geraniowego wobec bakterii oraz drożdży poz-
wala uznać go za skuteczny preparat naturalny o aktywności przeciwdrobnoustrojowej co stwarza możli-wość wykorzystania go jako naturalnego konserwantu w preparatach kosmetycznych.
Słowa kluczowe: Pelargonia, aktywność przeciwdrobnoustrojowa, geraniowy olejek eteryczny
Evaluation of antimicrobial activity of geranium essential oil
Abstract
For many years essential oils have been tested due to their potential to replace the synthetic preservatives in
cosmetics. Because of the geraniol and citronellol content in geranium oil, it is assumed that this essential oil could be one of the natural cosmetic preservative. The aim of this study was to evaluate the
antimicrobial activity of geranium essential oil. A commercial geranium oil (Avicenna-Oil) was used in
this research. The antimicrobial activity of oil ethanol solution in concentration of 0,25-10,00 µl/ml on
reference strains of ATCC collection of Escherichia coli, Staphylococcus aureus, Pseudomonas aeruginosa and Candida albicans was investigated. The microdilution in agar method and the modified
microdilution in broth technique were used to determined the MICs and MBC/MFCs.
This study showed that the most susceptible strain was C. albicans. In this case, the minimal inhibitory
concentration was 0,1 µl/ml, for E. coli and S. aureus strains the MIC was higher (0,25 µl/ml). The most resistant strain was P. aeruginosa (MIC 10 µl/ml). The MBC/MFC for all tested strains was 10,0 µl/ml.
High antibacterial and antifungal activity of geranium essential oil allows researchers to acknowledge it as
a natural effective antimicrobial substance, which can be used as a natural cosmetic preservative.
Key words: Pelargonium, antimicrobial activity, geranium essential oil

80
Ewa Kochan1, Hanna Zielińska-Bliźniewska
2, Monika Sienkiewicz
3
Przeciwdrobnoustrojowe działanie saponin
1. Wprowadzenie
Saponiny (saponozydy) są glikozydami, których nazwa pochodzi od łacińskiego
słowa „sapo” oznaczającego mydło i jest związana ze zdolnością do zmniejszania
napięcia powierzchniowego wodnych roztworów oraz wywoływania efektu pienienia
się. Związki te są bardzo rozpowszechnione w świecie roślin zarówno w klasie Dwu-
liścinnych (Dicotyledones), jak i Jednoliściennych (Monocotyledones). Znaleziono je
dotychczas w roślinach należących do około 80 rodzin, a najliczniej występują
w rodzinach: Agavaceae (Agawowate), Araliaceae (Araliowate), Caryophyllaceae
(Goździkowate), Chenopodiaceae (Komosowate), Dioscoreaceae (Pochrzynowate),
Fabaceae (Bobowate), Hippocastanceae (Kasztanowate), Fabaceae (Bobowate),
Liliaceae (Liliowate), Poaceae (Wiechlinowate), Polygonaceae, (Rdestowate),
Primulaceae (Prymulkowate) i Theaceae (Herbatowate) [1, 2]. Zawartość tych meta-
bolitów w materiale roślinnym zależy od wielu czynników, począwszy od tych biolo-
gicznych jak gatunek rośliny, uwarunkowania genetyczne, czy części rośliny, w której
się akumulują, przez czynniki środowiskowe, agrotechniczne, sposób przechowywania
czy czas zbioru surowca.
Saponiny występują nie tylko w roślinach, ale także w świecie zwierzęcym. Przy-
kładem jest ich obecność w ciele strzykw (rodzina Holothuroidea) zwierząt morskich,
które zamieszkują głównie obszary denne [3].
2. Budowa saponin
Saponiny jako związki glikozydowe o masie cząsteczkowej 600-1500 u składają się
z aglikonu (sapogeniny) i glikonu (cukier). W zależności od typu sapogeniny rozróżnia
się saponiny triterpenowe i steroidowe. Aglikony saponin triterpenowych zawierają 30
atomów węgla i zaliczane są do jednego z 3 typów: β-amaryny (oleananu), α-amaryny
(ursanu) oraz lupeolu. W strukturze saponozydów steroidowych występuje rdzeń
steranu o 27 atomach węgla z bocznym ugrupowaniem cyklicznym w pozycji 17.
Wśród saponin steroidowych wyróżnia się dodatkowo pochodne spirostanu oraz
furostanu [2, 4].
Glikon może zawierać od 1 do 12 cząsteczek cukrów prostych takich jak: D-
glukoza, D-galaktoza, D-ksyloza, D-arabinoza, L-ramnoza, L-fukoza, rzadziej kwas
glukuronowy i D-galakturonowy. Łańcuch cukrowy jest prosty bądź rozgałęziony,
1 [email protected], Zakład Biotechnologii Farmaceutycznej, Wydział Farmaceutyczny,
Uniwersytet Medyczny w Łodzi 2 [email protected], Zakład Alergologii i Rehabilitacji Oddechowej, Wydział
Wojskowo-Lekarski, Uniwersytet Medyczny w Łodzi 3 [email protected], Zakład Alergologii i Rehabilitacji Oddechowej, Wydział Wojskowo-
Lekarski, Uniwersytet Medyczny w Łodzi

Przeciwdrobnoustrojowe działanie saponin
81
mogą też występować jeden, dwa lub trzy oddzielne łańcuchy. W zależności od liczby
łańcuchów cukrowych, wchodzących w skład cząsteczki saponin, dzieli się je na:
monodesmozydy, bidesmozydy oraz tridesmozydy. Niektóre saponozydy zawierają
dodatkowo reszty kwasów organicznych przyłączonych do grupy OH aglikonu lub
cukru (saponiny estrowe) [2, 4].
Szkielet aglikonu, liczba i miejsce występowania podwójnych wiązań, położenie
grup funkcyjnych, jak również liczba i rodzaj cukrów stanowią o bardzo dużym
zróżnicowaniu tej grupy metabolitów. Różnorodność strukturalna tych związków
znajduje odzwierciedlenie w ich właściwościach fizykochemicznych oraz biolo-
gicznych [2, 4].
3. Wybrane cechy saponin i ich wykorzystanie
W większości saponiny są substancjami bezpostaciowymi, dobrze rozpuszczalnymi
w wodzie i alkoholu. Niektóre z nich mają słodki smak (np. kwas glicyryzynowy,
pozyskiwany z lukrecji gładkiej Glycyrrhiza glabra), a inne gorzki (np. saponiny
z nasion komosy ryżowej Chenopodium quinoa). Związki te mają różnorodne właści-
wości. Ze względu na obecność części polarnej (glikon) i niepolarnej (aglikon) są
związkami amfifilowymi co sprawia, że charakteryzują się wysoką zdolnością od
obniżania napięcia powierzchniowego wodnych roztworów. Dzięki temu w kontakcie
z wodą mogą tworzyć stabilną pianę oraz emulgować tłuszcze [4].
Przykładem komercyjnego wykorzystania tych cech jest ich zastosowanie
w przemyśle kosmetycznym, spożywczym, farmaceutycznym i medycynie. W kosme-
tologii saponiny izolowane z Camellia oleifera i Sapindus mukorossi wykorzystuje się
jako składniki detergentów, szamponów, żeli pod prysznic, lakierów do włosów,
płynnych mydeł, produktów do pielęgnacji niemowląt, płynów do płukania ust, czy
past do zębów [5]. Saponozydy okazały się użyteczne również w przemyśle spożyw-
czym. Związki aktywne pozyskiwane z juki (Yukka schidigera) i mydłodrzewa właści-
wego (Quillaja saponaria) są stosowane jako dodatki do żywności. Źródłem cennych
saponozydów są także koncentraty sojowe, stanowiące komponenty żywności
funkcjonalnej i nutraceutyków. Kwas oleanolowy wykorzystywany jest jako środek
zapachowy, maskuje również posmak sztucznego słodzika [6]. Z kolei saponozydy
z Q. saponaria znalazły zastosowanie w naukach medycznych pełniąc funkcję adiu-
wantów w szczepionkach weterynaryjnych [7, 8], czy ludzkich przeciwko wirusowi
ospy wietrznej/półpaśca [9], Mycobacterium tuberculosis [10] oraz tworzonych
eksperymentalnych szczepionkach przeciwko HIV [11].
Związki te stanowią również prekursory w syntezie kortykosteroidów i hormonów
płciowych (np. sarsasapogenina, smilagenina, tigogenina, ruskogenina oraz heko-
genina).
Powszechne zastosowanie w przemyśle farmaceutycznym saponiny zawdzięczają
swoim właściwościom wykrztuśnym. Na przykład, primulasaponiny zawarte
w korzeniu pierwiosnka (Primula sp.), czy saponazyd A z korzeni mydlnicy lekarskiej
(Saponaria officinalis) działają drażniąco na zakończenia nerwowe błony śluzowej
żołądka, powodując w ten sposób pobudzenie ośrodka nerwu błędnego. Prowadzi to do

Ewa Kochan, Hanna Zielińska-Bliźniewska, Monika Sienkiewicz
82
zwiększenia produkcji i wydzielania wodnistego śluzu w drogach oddechowych,
a w rezultacie powoduje odruch kaszlu. Zatem uwzględniając powyższe właściwości
saponiny te stały się obecnie składnikami aktywnymi wielu preparatów dostępnych na
rynku, które mają działanie przeciwkaszlowe i ułatwiające odksztuszanie (np.
Pectosol®, Tussipect
®) [2].
Saponozydy wykazują także właściwości przeciwzapalne (np. fruticesaponina B
z Bupleurum fruticescens, escyna z Aesculus hippocastanum), cytotoksyczne
(np. saponiny soi, ginsenozydy Rh2 i Rh3 z żeńszenia), hepatoochronne (np. saponiny
z Pueraria lobata) i adaptogenne (np. saponiny żeńszenia) [6]. Powodują hemolizę
czerwonych krwinek, przez co stają się bardzo toksyczne, po bezpośrednim podaniu do
krwiobiegu. Inaczej jest po podaniu doustnym, nie są wówczas szkodliwe, ponieważ
ich toksyczność jest minimalizowana ze względu na to, że bardzo słabo resorbują się
z przewodu pokarmowego – ulegają rozpadowi do sapogenin, które wydalane są
z moczem [6]. Hemolityczne właściwości zależą w głównej mierze od struktury
aglikonu. Największą aktywność hemolityczną wykazują saponiny triterpenowe
o aglikonie typu kwasu oleanolowego (np. escyna) lub hederageniny (np. α-hederyna),
natomiast saponiny triterpenowe o aglikonie tetracyklicznym typu dammaranu nie
oddziałują na czerwone krwinki. Do silnie hemolizujących saponin należą również
monodesmozydy [5, 12].
Ponadto wśród innych właściwości saponozydów wymienia się aktywność
pierwotniakobójczą (α- i β-hederyny z bluszczu) oraz mięczakobójczą (saponozydy
z nasion Camellia oleifera) [6, 13].
4. Właściwości przeciwdrobnoustrojowe saponin
Wiele światowych doniesień wskazuje na przydatność saponin w walce z drobno-
ustrojami. Jest to szczególnie ważny aspekt badań zwłaszcza w odniesieniu do narasta-
jącego problemu oporności mikroorganizmów na antybiotyki i chemioterapeutyki.
4.1. Właściwości przeciwbakteryjne saponin
Typowym przykładem ilustrującym rozprzestrzenianie się antybiotykooporności
jest rosnący brak wrażliwości wielu bakterii na stosowane leki, wynikający w dużej
mierze z nazbyt szerokiego stosowania antybiotyków w medycynie, rolnictwie czy
sadownictwie. Wiele danych sugeruje, że saponiny mogłyby być użyteczne zarówno
w prewencji, jak i w walce z chorobami wywoływanymi przez różne gatunki bakterii.
Według doniesień naukowych glicyryzyna i kwas glicyryzynowy – saponiny
triterpenowe, izolowane z lukrecji (Glycyrrhiza glabra), okazały się aktywne wobec
izolatów Streptococcus mutans, odpowiedzialnych między innymi za próchnicę zębów.
W powszechnie wykorzystywanej metodzie badań wrażliwości patogenów na anty-
biotyki/chemioterapeutyki wykazano, że strefa zahamowania wzrostu S. mutans przez
etanolowe wyciągi z tej rośliny była stosunkowo duża i wynosiła 26,7 mm [14].
Autorzy przytoczonych badań wskazali, że wyciągi z lukrecji w związku z ich
właściwościami hamującymi wzrost S. mutans mogą stać się składnikiem past do
zębów i płukanek do jamy ustnej, ograniczając w ten sposób występowanie próchnicy.

Przeciwdrobnoustrojowe działanie saponin
83
W przypadku saponin, wyizolowanych z żeńszenia północnoamerykańskiego
(Panax quinquefolius) okazało się, że wykazywały one przeciwbakteryjne właściwości
wobec trzech gatunków wywołujących halitozę – Fusobacterium nucleatum,
Clostridium perfringens, Porphyromonas gingivalis. Spośród 12 ginzenozydów –
saponozyd Rg5 działał najsilniej, przy czym minimalne stężenie hamujące – MIC (ang.
Minimal Inhibitory Concentration) i minimalne stężenie bakteriobójcze – MBC (ang.
Minimal Bactericidal Concentration) wynosiło odpowiednio: 16 i 62,5 μg/ml. Ponadto
stwierdzono, że mniej polarne saponozydy t.j. Rg2, Rg3, Rg6, F4, Rg5, Rk1
charakteryzują się większą skutecznością niż ginzenozydy Rg1, Re, Rc, Rb2 i Rd.
Uzyskane wyniki wiążą się ze zwiększonym powinowactwem mniej polarnych
związków do elementów błon komórkowych, a tym samym silniejszego ich oddziały-
wania z błoną. Efektem tego są uszkodzenia integralności i permeabilizacja błony
komórkowej bakterii. Wyjaśnienie mechanizmu działania ginsenozydów dowodzi, że
metabolity te mogłyby być potencjalnymi składnikami przeciwbakteryjnych środków
kontrolujących halitozę [15].
Triterpenowe glikozydy żeńszenia hamują również rozwój Propionibacterium
acnes, bakterii będącej czynnikiem etiologicznym trądziku. Badając wpływ
saponinowych ekstraktów z żeńszenia na wzrost szczepów wzorcowych P. acnes
(ATCC 11827 i ATCC 6919), a także Staphylococcus epidermidis (ATCC 12228)
i Staphylococcus aureus (ATCC 25923) wykazano, że największą strefą zahamowania
wzrostu charakteryzował się P. acnes. Strefa ta była porównywalna z tą otrzymaną dla
erytromycyny. Dla tego gatunku wartości MIC i MBC wyniosły 64 μg/ml. Dla
pozostałych badanych bakterii oba parametry osiągnęły wyższe wartości [16]. Biorąc
pod uwagę przeciwbakteryjne działanie wyciągów z żeńszenia sugeruje się ich
zastosowanie w maściach, kremach lub produktach do pielęgnacji skóry trądzikowej.
Lunga i wsp. [17] podają, że saponiny izolowane z liści i łodyg afrykańskiej rośliny
Paullinia pinnata skutecznie hamowały wzrost takich patogenów jak: Enterobacter
aerogenes (ATCC 13048), Enterococcus faecalis (ATCC 10541), Klebsiella
pneumoniae (ATCC 13883), Pseudomonas aeruginosa (ATCC 27853), Providencia
smartii (ATCC 29916) i Staphylococcus aureus (ATCC 25922). Warto podkreślić, dla
S. aureus właściwości bójcze, wybranych saponozydów, były silniejsze niż uzyskane
po zastosowaniu aminoglikozydu – gentamycyny. Według autorów powyższych badań
metabolity pozyskane z P. pinnata mogłyby stanowić uzupełnienie istniejących terapii
bądź w przyszłości stać się alternatywą leczenia zakażeń [17].
Saponozydy izolowane z innych egzotycznych roślin – Allophylus cobbe
i Allophylus serratus wykazały silniejsze działanie hamujące wzrost Bacillus subtilis
(strefa zahamowania wzrostu wynosiła odpowiednio od 17,3 do 20,3 mm w zależności
od rodzaju wyciągu) niż cefalosporyna III generacji – cefotaksym (strefa zahamowania
wzrostu: 14,75 mm). Takie właściwości saponin z Allophylus sp. wskazują, że związki
te można by zastosować np. w produkcji opakowań przeznaczonych do przecho-
wywania pieczywa, celem eliminacji rozwoju B. subtilis, odpowiedzialnego jego
śluzowacenie, szary nalot, mazistość, i nieprzyjemny zapach [18].

Ewa Kochan, Hanna Zielińska-Bliźniewska, Monika Sienkiewicz
84
4.2. Właściwości przeciwgrzybicze saponin
Wśród doniesień dotyczących przeciwdrobnoustrojowej aktywności saponin
znaleźć można badania poświęcone działaniu przeciwgrzybiczemu tych związków.
Szczególnie cenne są dane opisujące wpływ tych metabolitów na rozwój dermatofitów
– grzybów odpowiedzialnych za grzybicę skóry, włosów i paznokci [19].
Według danych Światowej Organizacji Zdrowia, około 25% ludności świata ulega
zakażeniu różnymi gatunkami dermatofitów. Wśród gatunków wyizolowanych od
pacjentów z grzybicą stóp, pachwinową, skóry gładkiej czy dermatofitową grzybicą
paznokci najczęściej występującymi okazały się Epidermophyton occosum, Tricho-
phyton rubrum i Trichophyton mentagrophytes [20].
Xue i wsp. [21] stwierdzili, że triterpenowe saponiny – ginsenozydy – pozyskane
z Panax notoginseng wykazują hamującą aktywność względem wymienionych
powyżej dermatofitów. Badane ginsenozydy podzielono na grupę mniej (ginsenozydy
-RK3, -Rh4, -Rh5) i bardziej polarnych (notoginsenozyd-R1, ginsenozyd-Rg1, -Re,
-Rb2, -Rd). Otrzymane wyniki wykazały, że znacznie wyższą zdolność hamowania
wzrostu grzybiczych patogenów skóry miały mniej polarne saponiny (-RK3, -Rh4,
-Rh5). Było to zbieżne z wcześniej opisanymi obserwacjami, uzyskanymi dla bakterii
wywołujących halitozę. W badaniach dermatofitów dolna wartość MIC wynosiła 16
µg/ml dla T. rubrum oraz 31,25 µg/ml dla E. occosum i T. mentagrophytes. Określono
również wartość minimalnego stężenia grzybobójczego MFC (ang. Minimal Fun-
gicidal Concentration) dla T. rubrum – 125 µg/ml i pozostałych 2 gatunków – 250
µg/ml. Zauważono również prawidłowość, wskazującą, że przeciwgrzybicza aktyw-
ność poszczególnych ginsenozydów rośnie wraz z mniejszą ilością ugrupowań
cukrowych w cząsteczce metabolitu. Ponadto stwierdzono, że działanie przeciwgrzy-
bicze badanych saponin może wynikać z uszkodzeń błony komórkowej i zmniejszenia
potencjału błonowego, do czego w łatwy sposób przyczyniają się mniej polarne
związki. Uzyskane wyniki sugerują również, że mniej polarne ginsenozydy można
zastosować jako środki wspomagające leczenie grzybicy [21]. Warto zaznaczyć, że
znacznie silniejsze działanie wobec dermatofitu T. mentagrophytes wykazał triterpe-
nowy glikozyd holoturin b, pozyskiwany z gatunku strzykw Actinopyga lecanora.
Minimalne stężenie hamujące wzrost grzyba odnotowano przy stężeniu tego związku
wynoszącym 1,56 µg/ml [22].
Inne badania podają, że mieszanina saponin z drzewa Maesa lanceolata w stężeniu
50 µg/ml znacznie ograniczała wzrost gatunków, takich jak: Epidermophyton
floccosum, Microides interdigitalis, Trichophyton rubrum. W przypadku pozostałych
dermatofitów (Microsporum canis, Microsporum Langerowi) i dla drożdżaków
(Candida albicans) ilość aplikowanych saponin musiała być zwiększona do 100 lub
250 µg/ml [4].
Powszechnymi i dobrze poznanymi oportunistycznymi patogenami grzybiczymi
ludzi i zwierząt są grzyby z rodzaju Candida spp., np. C. albicans. Drobnoustrój ten
może bytować w przewodzie pokarmowym i układzie moczowo-płciowym u ponad
70% ludzi. Jednakże, w pewnych okolicznościach, C. albicans może powodować

Przeciwdrobnoustrojowe działanie saponin
85
infekcje skórne i błon śluzowych (np. kandydozy paznokci, jamy ustnej, gardła, jelit).
Bywa także przyczyną ogólnoustrojowych zakażeń zagrażających życiu, takich jak
kandydemia, zapalenie płuc, grzybicze zapalenie wsierdzia, czy zapalenie opon
mózgowych szczególnie u osób z deficytami immunologicznymi, w tym u przewlekle
chorych np. w przypadku cukrzycy [23]. Okazuje się, że saponiny wyizolowane
z części nadziemnych oraz korzeni Medicago sativa i korzeni Saponaria officinalis,
stosowane samodzielnie wykazują aktywność przeciwgrzybiczną, a w skojarzeniu
z lekami przeciwgrzybicznymi, preparatami chemicznymi jest ona wzmożona, co
pozwala na zmniejszenie aktywnych stężeń tych leków. Saponiny te hamują wzrost
strzępek, adhezję i tworzenie biofilmu przez C. albicans (ATCC 10231) już w stężeniu
125-500 µg/ml [24].
Inne triterpenowe saponiny – ginsenozydy Rb1, Rb2, Rc, Re, Rf Rg1, Rg2 Rg3
i Rh2, pozyskane z koreańskiego żeńszenia (Panax ginseng), także działają bójczo
wobec C. albicans (MIC 100 µg/ml). Podobnie jak metabolity z M. sativa i S.
officinalis, ginsenozydy blokują tworzenie micelarnych struktur tego grzyba, które
odpowiedzialne są za patogenność i w tej formie przede wszystkim zasiedlają tkanki
gospodarza [25].
Innym ważnym patogenem, wywołującym chorobę przewlekłą nazywaną sporo-
trychozą jest Sporothrix schenckii. Najczęściej zajmuje on skórę, choć może również
atakować stawy, kości, płuca i mózg. Zakażenie wśród ludzi występuje zwykle przez
ukłucie się kolcami skontaminowanej róży. Patogen ten może także przenosić się przez
glebę, rośliny, drewno czy zainfekowane zwierzęta. Choroba postępuje wolno, pierw-
sze objawy mogą wystąpić od 1 do 12 tygodni po zakażeniu. W leczeniu stosuje się
leki przeciwgrzybicze. Wspomniana wcześniej triterpenowa saponina holoturin b,
wykazała porównywalną z flukonazolem aktywność wobec S. schenckii [26].
Identyczne obserwacje w badaniu saponiny holoturin b odnotowano w stosunku do
grzyba jakim jest Aspergillus fumigatus. U ludzi jest on przyczyną różnych postaci
klinicznych zakażeń takich jak aspergiloza u pacjentów z obniżoną odpornością, ostre
i przewlekłe inhalacyjne zakażenia dróg oddechowych, grzybica uszu, przewodu
słuchowego, jamy sutkowej, oka i zatok przynosowych, czy reakcji alergicznych pod
postacią kataru siennego i astmy oskrzelowej. A. fumigatus to również patogen
zwierząt, w tym ptaków oraz owadów. Stąd też saponina holoturin b byłaby cennym
uzupełnieniem terapii u osób dotkniętych aspergilozą, a także mogłaby mieć
zastosowanie w weterynarii [22].
Analizując rezultaty przytoczonych różnych badań na temat właściwości triterpe-
nowych saponin, wydaje się bardzo obiecującym ich wykorzystanie jako składników
środków dezynfekujących lub nowych przeciwgrzybiczych terapeutyków, wspie-
rających klasyczne leki.
Istotnym problemem jest także walka z chorobami grzybowymi wielu roślin
uprawnych. Chapagain i wsp. [27] wykazali, że bogate w saponiny ekstrakty, pozys-
kane z mezokarpu owoców kolibła egipskiego (Balanites aegyptiaca), kory mydło-
drzewa właściwego (Quillja saponaria) (QE) i juki Schidigera (Yucca schidigera)
mogą hamować wzrost fitopatogennych grzybów takich jak: Pythium ultimum,

Ewa Kochan, Hanna Zielińska-Bliźniewska, Monika Sienkiewicz
86
Fusarium oxyporum, Alternaria solani, Colletotrichum coccodes i Verticillium dahliae.
Inhibicyjny wpływ wyciągów badano w warunkach in vitro określając stężenia
ekstraktów, przy których dochodziło do zmniejszenia średnicy kolonii grzybów do
50% w stosunku do grupy kontrolnej. Dla prób pozyskanych z Y. schidigera okazało
się, że stopień hamowania wzrostu wszystkich badanych grzybów był bardzo wysoki
lub umiarkowany (54,1-100%). Słabsze działanie wykazały wyciągi z B. aegyptiaca,
bowiem wyraźnie ograniczony rozrost grzybni uzyskano tylko dla P. ultimum (81,1%).
W przypadku wyciągów, zawierających triterpenowe saponiny z Q. saponaria
wykazano umiarkowane zahamowanie wzrostu (35.9-59.1%) wszystkich grzybów,
poza C. coccodes [27]. Inne saponiny hemojedemozydy A i B oraz patagonikozyd A,
pozyskiwane ze strzykw (odpowiednio z Hemoiedema spectabilis i Psolus pata-
gonicus) działały silnie przeciwgrzybiczo wobec patogennego dla ogórków Clado-
sporium cucumerinum [22]. Wyniki przytoczonych badań sugerują, że wybrane
saponinowe ekstrakty mogą odgrywać ważną rolę w ekologicznej walce z patogenami
roślin, które przyczyniają się do strat w uprawie takich roślin jak kukurydza, soja,
ziemniaki, ogórki, pszenica, jodła czy gatunki ozdobne.
Wielu autorów podjęło się wyjaśnienia mechanizmu działania saponin na błony
komórkowe grzybów. Są oni zgodni, że związki te tworzą kompleksy ze sterolami błon
komórek grzybowych, zaburzając w ten sposób ich integralność. Uszkodzenia błon
komórkowych doprowadzają także do destabilizacji potencjału błonowego, jak również
mogą powodować specyficzne zakłócenia w funkcjonowaniu kanałów jonowych oraz
doprowadzić do tworzenia w błonach niespecyficznych porów [21, 24, 25].
4.3. Antywirusowe działanie saponiny
Jednym z częściej spotykanych środowisku wirusów zakażających ludzi jest wirus
z rodziny Herpesviridae – wirus opryszczki HSV(ang. Herpes Simplex Virus). Jest on
przyczyną zmian zapalnych na granicy skóry i błony śluzowej. W zależności od
rodzaju wirusa atakuje jamę ustną i okolice warg (HSV-1) lub narządy płciowe (HSV-
2). Wirus opryszczki w rzadkich przypadkach może powodować zapalenie spojówek
i rogówki, skóry lub opryszczkowe zapalenie mózgu. Wykazano, że saikosaponina-D
pozyskiwana z korzeni przewiercienia sierpowatego (Bupleurum falcatum) powoduje
inaktywację obu typów wirusów w stężeniu większym niż 5 µM w warunkach in vitro,
a glicyryzyna – saponina pozyskiwana z lukrecji, hamuje namnażanie wirusa HSV-1
i wywoływany przez niego efekt cytopatyczny [28]. Wykazano również, że triter-
penowe saponozydy izolowane z wielu roślin, należących do rodziny Fabaceae,
skutecznie hamują replikację HSV-1 [6, 29], a saponinowe wyciągi z żeńszenia
zapobiegają penetracji wirusa, silnie aktywują układ immunologiczny gospodarza
i ułatwiają eliminację patogena z miejsca objawów klinicznych [30]. Ze względu na
różnorodne działania przeciwwirusowe wydaje się zatem, że zastosowanie wyciągów
saponin byłoby korzystne w zapobieganiu lub łagodzeniu pierwotnych i nawracających
infekcji HSV w kombinacji z konwencjonalnymi lekami przeciwwirusowymi.
Innym powszechnym w środowisku wirusem jest Infulenza wirus będącym przy-
czyną grypy. Na chorobę tą co roku zapada od 5% do 25% populacji, a śmiertelność

Przeciwdrobnoustrojowe działanie saponin
87
z jej powodu według Światowej Organizacji Zdrowia (WHO, ang. World Health
Organization) wynosi od 0,5 mln do 1 mln osób szczególnie dotyczy to dzieci i osób
w podeszłym wieku. Stwierdzono, że niektóre saponiny, jak kwas gymnemowy wystę-
pujący w Gymnema sylvestre, escyna kasztanowca, a także saponiny izolowane
z prymuli czy herbaty utrudniają lub hamują proces adsorbcji wirusa grypy do ko-
mórek gospodarza. Asterosaponiny uzyskiwane z Asterias torbesi, Acanthaster planci
czy Asterina pectinifera natomiast hamują jego namnażanie [28].
Informacja ta ma szczególne znaczenie jeśli zwrócimy uwagę na wirusa ptasiej
grypy, który może przełamać barierę gatunkową i zagrażać naszemu życiu. Wirus
ptasiej grypy H9N2 jest odpowiedzialny za wysoką śmiertelność komórek śródbłonka
i poważne stany zapalne, a w konsekwencji apoptozę komórek gospodarza. Piśmien-
nictwo donosi, że niektóre triterpenowe saponiny żeńszenia – pochodne protopana-
ksartiolu, mogą działać ochronnie na zainfekowane komórki, łagodząc tym samym
skutki zakażenia [31]. Co więcej podobne obserwacje odnotowano wobec innych
wirusów przenoszonych drogą kropelkową, tzw. rinowirusów, będących główną
przyczyną przeziębień w okresie jesienno-zimowym i zimowo-wiosennym [30].
Jednym z najważniejszych wirusów, nad którym prowadzone są intensywne
badania już od lat 80-tych i którym zakażenie dotyka kilku tysięcy osób dziennie, jest
ludzki wirus niedoboru odporności, HIV (ang. Human Immunodeficiency Virus), AIDS
(ang. Human Immunodeficiency Syndrome). Dotychczas poznano 2 typy wirusa HIV-1
i HIV-2. Szacuje się, że 95% zakażeń na całym świecie jest powodowanych przez
HIV-1, bardziej zjadliwy i zakaźny typ niż HIV-2. Przy znaczącym spadku liczby
komórek CD4 należy wdrożyć wysoce aktywną terapię antyretrowirusową, HAART
(ang. Higly Active Antiretroviral Therapy). Polega ona na podaniu kilku leków
blokujących replikację HIV. Leczenie nie umożliwia całkowitego wyeliminowania
wirusa, ale oferuje redukcję jego replikacji. Wyniki badań wielu autorów pokazują, że
ginsenozydy – saponozydy żeńszenia wykazują silne oddziaływanie przeciwwirusowe
wobec HIV-1, zarówno po podaniu wraz z typowymi przeciwwirusowymi preparatami
jak i bez nich. Wspomniane rezultaty okazują się na tyle obiecujące, że postuluje się
podawanie wyciągów z żeńszenia jako alternatywną formę leczenia. Co ciekawe,
ginsenozydy zwiększają częstotliwość mutacji – delecji w obrębie jednego z kore-
ceptorów dla HIV, biorącego udział w procesie wnikania patogena do komórki. Tym
samym podawanie ich pacjentom z HIV-1 prowadzi do opóźnienia rozwoju choroby
i zwiększenia przeżywalności. Dodatkową zaletą saponin żeńszenia (np. ginsenozydu
Rh) jest zmiana farmakologicznych właściwości leków anty-HIV, poprzez zwięk-
szenie ich akumulacji w komórkach [30].
Innym RNA wirusem wobec, którego badano wpływ saponin jest wirus HAV (ang.
Hepatitis A Virus) odpowiedzialny za wirusowe zapalenie wątroby przenoszone drogą
pokarmową tzw. żółtaczka pokarmowa. Zakażenie następuje przez kontakt z wydzie-
linami chorego lub skontaminowanymi produktami (epidemie mleczne, wodne itp.).
Obecnie nie ma żadnego dostępnego środka przeciwwirusowego dla HAV, a najsku-
teczniejszym sposobem prewencji jest szczepienie oraz zachowywanie zasad higieny
i poprawa warunków sanitarnych. Stąd też poszukuje się substancji, które działałyby

Ewa Kochan, Hanna Zielińska-Bliźniewska, Monika Sienkiewicz
88
przeciwko infekcji wirusem Przykładem tego typu związków są ginsenozydy Rb1
i Rg1 (saponiny izolowane z gatunków należących do rodzaju Panax), które
zmniejszają miano HAV, a regularne ich spożycie mogłoby pomóc w zapobieganiu
rozwojowi zakażenia HAV [30].
Zgodnie z piśmiennictwem kolejny ginsenozyd – Rg3 wykazuje działanie przeciw-
wirusowe na inny typ wirusa zapalenia wątroby – typ B (HBV, ang. Hepatitis B Virus).
Może powodować on ostre lub rzadziej przewlekłe zapalenie wątroby prowadzące
nawet do marskości i raka wątroby [30]. Pomimo powszechnych szczepień ochron-
nych starsza część populacji, która nie przyjęła profilaktyki sztucznej czynnej, wciąż
ulega zakażeniu, zatem informacja ta ma duże znaczenie na przyszłość w ograniczeniu
rozwoju infekcji [30]. Saponinowe wyciągi z żeńszenia (w szczególności ginsenozydy
Rg1 i Rb1), działają hamująco również na rozwój takich wirusów jak: rotawirusy,
mysie norowirusy (MNV, ang. Maurine Noro Virus) i kaliciwirusy (FCV, ang. Feline
Calicivirus) kota czy syncitialny wirus oddechowy (RSV, ang. Respiratory Syncycal
Virus) [30, 31].
Omawiane metabolity zostały również poddane testom na przydatność w walce
z bezosłonkowymi RNA wirusami – enterowirusami. Jak się okazało spośród
ginsenozydów Rb1, Rb2, Rc Rd Re, Rf i Rg2 jedynie saponina Rg2 charakteryzowała
się aktywnością przeciwwirusową wobec większości enterowirusów i wykazywała
działanie hamujące proces adhezji wirusów do komórki gospodarza. Odmienne rezul-
taty uzyskano w odniesieniu do jednego z nich - wirusa Coxsackie B3 (CVB3, ang.
Coxackievirus B3), powodującego zapalenie mięśnia sercowego, aseptyczne zapalenie
opon mózgowych i trzustki. Ginsenozydy Re, Rf, i Rg2 wykazały znaczące działanie
przeciw wirusowi CVB3. Najsilniejszą aktywność, zarówno w warunkach in vitro jak
i in vivo, przejawiał 20 (S) – protopanaksatriol. Aktywność ta była wyższa od ryba-
wiryny – typowego środka przeciwwirusowego. Wśród saponin znalazły się również
takie, które inaktywują wirusa odry. Należy do nich saikosaponina-D wyizolowana
z korzeni przewiercienia sierpowatego (Bupleurum falcatum) [28].
Problem chorób wirusowych dotyczy nie tylko ludzi, ale i zwierząt. Wirusowa
martwica układu nerwowego (VNN, ang. Viral Nervous Necrosis), wywoływana przez
wirusy z grupy betanodawirusów, to choroba ryb morskich. Stała się ona poważnym
utrudnieniem i przyczyną ogromnych strat w hodowlach larw i młodocianych ryb
morskich na całym świecie. Ogromną przeszkodą w efektywnym leczeniu VNN jest
pojawianie się nowych szczepów wirusowych, opornych na istniejące preparaty. Stąd
też poszukuje się alternatywnych źródeł skutecznych leków przeciwwirusowych.
Takim potencjałem charakteryzuje się gymnemagenol wyodrębniony z liści Gymnema
sylvestre. Zgodnie z doniesieniami naukowymi związek ten przy stężeniu 20 µg ml/l
hamuje proliferację nodawirusa GNNV (ang. Grouper Nervous Necrosis Virus) oraz
znacznie zmniejsza jego miano. Uzyskane wyniki pozwalają uznać, że gymnemagenol
może być stosowany jako środek przeciw infekcji GNNV [32].
Formy bezkomórkowe patogenów nie oszczędzają również upraw roślin użytko-
wych powodując straty w rolnictwie. Takim patogenem generującym straty jest wirus
mozaiki ogórka (CMV, ang. Cucumber Mozaic Virus). Szczególnie często poraża on

Przeciwdrobnoustrojowe działanie saponin
89
uprawy ogórków, melonów, kabaczków, pomidorów, fasoli szpinaku, bananów, czy
papryki. Okazuje się, że β-escyna – saponia, w którą szczególnie bogate są nasiona
Aesculus indica, w 94% hamuje aktywność wirusa CMV. Efektywną dawką jest 1000-
2000 ppm. Ponadto β-escyna działa silniej w porównaniu z typowymi przeciw-
wirusowymi preparatami takimi jak acyklowir (ACV), czy zydowudyna (AZT) [28].
Wydaje się, więc, że saponina ta mogłaby znaleźć zastosowanie w ochronie upraw
przed wirusem mozaiki ogórka.
5. Podsumowanie
Analiza dostępnego piśmiennictwa naukowego dotyczącego właściwości
przeciwdrobnoustrojowych saponin wskazuje, że metabolity te mogą być cennym
uzupełnieniem terapii przeciwinfekcyjnej. Są aktywne zarówno wobec patogenów
bakteryjnych, grzybowych i wirusowych. Ponadto badania dowodzą, że związki te
z powodzeniem mogą być stosowane wobec patogenów odpowiedzialnych za
zakażenia ludzi, zwierząt, a także roślin uprawnych.
Literatura
1. Faizal A., Geelen D. Saponins and their role in biological processes in plants,
Phytochemistry Reviews., 21 (2013), s. 877-893.
2. Lamer-Zarawska E., Kowal-Gierczak B. Fitoterapia i leki roślinne, PZWL., 2012
3. Bahrami Y., Zhang W., Franco C. Discovery of novel saponins from the Viscera of the Sea
Cucumber Holothuria lessoni, Marine Drugs., 12(5) (2014), s. 2633-2667.
4. Parus A. Właściwości farmakologiczne saponin, Postępy Fitoterapii., 3, 2013, s. 200-204.
5. Moses T., Papadopoulou K.K., Osbourn A. Metabolic and functional diversity of saponins,
biosynthetic intermediates and semi-synthetic derivatives, Critical Reviews in Biochemistry
and Molecular Biology., 49(6), (2014), s. 439-462. doi:10.3109/10409238.2014.953628
6. Deore S.L., Khadabadi S.S., Chittam K.P., Bhujade P.G., Wane T.P., Nagpurkar Y.R.,
Chanekar P.D., Jain R.G. Properties and pharmacological applications of saponins,
Pharmacognosyonline., 2, (2009), s. 61-84.
7. Moghimipour E., Handali S. Saponin: properties, methods of evaluation and aplications,
Annual Research &Review in Biology., 5(3), (2015), s. 207-220.
8. Plotkin S.A., Orenstein W.A., Offit P.A.: Vaccines. 6th Ed. Philadelphia, PA: Saunders;
(2012).
9. Lal H., Zahaf T., Heineman T.C. Safety and immunogenicity of an AS01-adjuvanted
varicella zoster virus subunit candidate vaccine (HZ/su): a phase-I, open-label study in
Japanese adults, Human Vaccines Immunotherapeutics., 9, (2013), s. 1425-9.
10. Montoya J., Solon J.A., Cunanan S.R., Acosta L., Bollaerts A., Moris P., Janssens M.,
Jongert E., Demoitié M.A., Mettens P., Gatchalian S., Vinals C., Cohen J., Ofori-Anyinam
O. A randomized, controlled dose-finding phase II study of the M72/AS01 candidate
tuberculosis vaccine in healthy PPD-positive adults, Journal of Clinical Immunology., 33,
(2013), s. 1360-75.
11. Leroux-Roels G., Bourguignon P., Willekens J., Janssens M., Clement F., Didierlaurent
A.M., Fissette L., Roman F., Boutrau D. Immunogenicity and safety of a booster dose of an
investigational adjuvanted polyprotein HIV-1 vaccine in healthy adults and effect of
administration of chloroquine, Clinical and Vaccine Immunology., 21, (2014) s. 302-11.

Ewa Kochan, Hanna Zielińska-Bliźniewska, Monika Sienkiewicz
90
12. LimT.K. Edible medicinal and non-medicinal plants: Volume 6, fruits, Springer-Dordrecht
Heidelberg, New York, London ISBN 978-94-007-5628-1 (e-Book) (2013), s.17.DOI: 10.
1007/978-94-007-5628-1
13. Kijprayoona S., Toliengb V., Petsoma A., Chaicharoenpongb Ch. Molluscicidal activity of
Camellia oleifera seed meal, ScienceAsia., 40 (2014) s. 393-399.
14. Ajagannanavar S.L., Battur H., Shamarao S., Sivakumar V., Patol P.U., Shanavas P. Effect
of aqueous and alcoholic licorice (Glycyrrhiza Glabra) root extract against Streptococcus
mutans and Lactobacillus acidophilus in comparison to chlorhexidine: an in vitro study,
Journal of International Oral Health., 6(4) (2014), s. 29-34.
15. Hue P., Yao Y., Yang X., Feng J., Ren G. Improved antimicrobial effect of ginseng extract
by heat transformation, Journal of Ginseng Research., 41 (2), (2017) s. 180-187.
16. Wang L., Yang X., Yu X., Yao Y., Ren G. Evaluation of antibacterial and anti-inflammatory
activities of less polar ginsenosides produced from polar ginsenosides by heat –transfor-
mation, Journal of Agricultural and Food Chemistry., 61, (2013), s. 12274-12282.
17. Lunga P.K., Qin X-J., Yang X.W., Kuiate J-R., Du Z.Z., Gatsing D. Antimicrobial steroidal
saponin and oleanane-type triterpenoid saponins from Paullinia pinnata, BioMedCentral
complementary and Alternative Medicine., 14, (2014), s. 369.
18. Chavan R.B., Gaikwad D.K. Antibacterial activity of medicinally important two species of
allophylus – Allophylus cobbe(L.) Raeusch. and Allophylus serratus (roxb.) Kurz., Journal
of Pharmacognosy and Phytochemistry., 2(1), (2013), s.1-7.
19. Santos M.L., Maqlhaes C.F., da Rosa M.B., de Assis Santos D., Brasileiro B. G., de
Carvalho L.M., da Silva M.B., Zani C. L., de Siqueira E.P., Peres R.L., Andrade A.A.
Antifungal activity of extracts from Piper aduncum leaves prepared by different solvents
and extraction techniques against dermatophytes Trichophyton rubrum and Trichophyton
interdigitale, Brazilian Journal of Microbiology., 4, (2014), s. 1275-1278.
20. de AguiarPeres N.T., Maranhao F.C.A., Rossi A., Martinez-Rossi N.M. Dermatófitos:
interaçao patógeno-hospedeiro e resistência a antifúngicos, Anais Brasileiros de
Dermatologia., 85, (2010), s. 657-667.
21. Xue P., Yang X., Sunab X., Ren G. Antifungal activity and mechanism of heattransformed
ginsenosides from notoginseng against Epidermophyton floccosum, Trichophyton rubrum,
and Trichophyton mentagrophytes, The Royal Society of Chemistry Advances., 7, (2017)
s. 10939.
22. Kim S-K, Himaya S.W.A. Triterpene glycosides from sea cucumbers and their biological
activies w: Advences in food and nutrition research, marine medical foods - implication and
aplication - animals and microbes, rozdział 20, (2012), Vol.65, 297-319 K.
23. Kabir M.A., Ahmad Z. Candida Infections and Their Prevention, International Scholarly
Research Notices Preventive Medicine., 2013, (2013), s 1-13
http://dx.doi.org/10.5402/2013/763628
24. Sadowska B., Budzyńska A., Więckowska-Szakiel M., Paszkiewicz M., Stochmal A.,
Moniuszko-Szajwaj B., Kowalczyk M., Różalska B. New pharmacological properties of
Medicago sativa and Saponaria officinalis saponin-rich fractions addressed to Candida
albicans, Journal of Medical Microbiology., 63, (2014), s. 1076-86.
25. Sung W.S., Lee D.G. In vitro candidacidal action of Korean red ginseng saponins against
Candida albicans, Biological Pharmaceutical Bulletin., 31 (1), 2008, s. 139-142.
26. Téllez M. D., Batista-Duharte A., Portuondo D., Quinello C., Bonne-Hernández R., Carlos
Z.I. Sporothrix schenckii complex biology: environment and fungal pathogenicity,
Microbiology., 160, (2014), s. 2352-2365.

Przeciwdrobnoustrojowe działanie saponin
91
27. Chapagain B.P., Wiesman Z., Tsror (Lahkim) L. In vitro study of the antifungal activity of
saponin-rich extracts against prevalent phytopathogenic fungi, Industrial Crop and
Product., 26(2), (2007), s. 109-115.
28. European patent specification, EP1 489 910 B1, Date of publication and mention of the
grant of the patent: 11.04.2007 Bulletin 2007/15.
29. Bartnik M., Facey P.C. Glycosides w: Pharmacognosy: fundamentals, applications and stra-
tegies, edited Simone Badal, Rupica Delgoda, Elsevier Inc, Rozdział 8, (2017), s. 137-141.
30. Im K., Kim J., Min H. Ginseng, the natural effectual antiviral: Protective effects of Korean
Red Ginseng against viral infection, Journal of Ginseng Research., 40, (2016), s. 309-314.
31. Kang S., Min H. Ginseng, the 'Immunity Boost': The effects of Panax ginseng on immune
system, Journal of Ginseng Research., Vol. 36, (4), (2012), s. 354-368.
32. KhannaV.G., Kannabiran K., Babu V.S., HameedI A.S.S. Inhibition of fish nodavirus by
gymnemagenol extracted from Gymnema sylvestre, Journal of Ocean University of China.,
10 (4), 2011, s. 402–408. doi:10.1007/s11802-011-1841-2
Przeciwdrobnoustrojowe działanie saponin
Streszczenie
Saponiny (saponozydy) pod względem chemicznym należą do glikozydów. W zależności od budowy agli-
konu wyróżnia się saponiny steroidowe i triterpenowe. Są one szeroko rozpowszechnione w świecie roślin-
nym, występują również w ciele strzykw. W niniejszym opracowaniu scharakteryzowano właściwości sa-ponozydów ze szczególnym zwróceniem uwagi na działanie przeciwdrobnoustrojowe tych metabolitów.
Szereg doniesień potwierdza hamujące/bójcze ich działanie wobec patogennych bakterii i grzybów oraz
działanie przeciwwirusowe. Stąd też w przyszłości mogłyby znaleźć zastosowanie w prewencji próchnicy
czy halitozy, jako składnik różnych preparatów, czy byłyby pomocne w uzupełnieniu terapii bakteryjnych chorób skórnych. Saponiny oddziałując na zakażenia wirusowe poprzez hamowanie procesu adsorbcji
niektórych wirusów do komórek gospodarza, blokują wirusy atakujące zarówno ludzi, zwierzęta jak
i rośliny. W dobie rosnącej lekooporności zainteresowanie tymi metabolitami wzrasta w kontekście ich po-
tencjalnego wykorzysta w medycynie czy weterynarii. Ponadto obiecującym wydaje się być zastosowanie saponin w ekologicznej walce z drobnoustrojami zagrażającymi uprawom, czy tymi powodującymi psucie
żywności.
Słowa kluczowe: saponiny, właściwości przeciwbakteryjne, przeciwgrzybicze, przeciwwirusowe.
Antimicrobial activity of saponins
Abstract
Saponins (saponosides) are glycosides. Depending on the structure of the aglycone they are devided into to groups: triterpene and steroidal saponins. They are widely distributed in the vegetable kingdom, but also
occur in the body of sea cucumbers. In this study the properties of saponosides were characterized with
particular emphasis on the antimicrobial activity of these metabolites. A number of literature data, confir-
ming that these compounds inhibit the growth or they are lethal to the pathogenic bacteria, fungi and viruses. Therefore, in the future, some of saponins could be used in the prevention of tooth decay and
halitosis, as the ingredient of preparations for oral hygiene. It seems that the saponosides would be helpful
in the treatment of dermatological diseases. It is known that, saponins affect on the viral infections by
suppressing the adsorption of some viruses to host cells and blocking their proliferation. They can be useful in the fight against viruses that attack human, animals and plants particularly against the growth of
pathogens resistance. The interest of these metabolites is not only connected with their potential use in
medicine or veterinary medicine, but also as effective agents in the fighting against environmental
pathogens, which contributing to losses in the cultivation of crops and food contamination. Keywords: saponis, antibacterial, antifungal, antiviral properties.

92
Monika Sienkiewicz1, Hanna Zielińska-Bliźniewska
1, Jurek Olszewski
2,
Paweł Kwiatkowski3, Ewa Kochan
4
Aktywność olejków eterycznych
przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
1. Wprowadzenie
Stale rosnąca oporność na leki przeciwdrobnoustrojowe niesie za sobą potrzebę
poszukiwań skutecznych i możliwie bezpiecznych środków, mogących mieć zasto-
sowanie zarówno w leczeniu zakażeń, jak również w zapobieganiu wzrostowi leko-
oporności drobnoustrojów [1]. Jednocześnie ze względu na szybkie rozprzestrzenianie
się bakterii dysponujących szeregiem mechanizmów oporności, niezwykle ważna jest
skuteczna eradykacja wielolekoopornych drobnoustrojów obecnych zarówno
w środowiskach szpitalnych, jak i w lecznictwie otwartym oraz wszelkiego rodzaju
placówkach związanych z opieką medyczną.
2. Oporne drobnoustroje
Szczególną pozycję wśród opornych bakterii od wielu lat zajmują gronkowce. Są to
nie tylko, jak często donosi literatura, metycylinooporne gronkowce złociste MRSA,
ale również gronkowce koagulazo-ujemne, jak metycylinooporne Staphylococcus
epidermidis MRSE, które są coraz częściej przyczyną poważnych infekcji, w tym
również zakażeń szpitalnych. Gronkowce te cechuje brak wrażliwości na antybiotyki
beta-laktamowe, w tym penicyliny, cefalosporyny, monobaktamy i karbapenemy [2-6].
Co więcej wśród Gram-dodatnich bakterii, u gronkowców i paciorkowców występuje
mechanizm oporności MLSB, którego obecność wyklucza zastosowanie makrolidów,
linkosamidów i streptogamin B [7-9]. Szczególnych trudności klinicznych nastręczają
też enterokoki oporne na: aminoglikozydy – HLAR jak: gentamycyna, streptomycyna,
amikacyna, tobramycyna; glikopeptydy – VRE, które nie są wrażliwe nawet na
działanie wankomycyny określanej mianem leku „ostatniej szansy”, w tym także
enterokoki należące do klasy Van A oporne na wankomycynę i teioplaninę [10].
Wśród Gram-ujemnych bakterii poważne zagrożenie stanowią pałeczki z rodziny
1 [email protected], [email protected], Zakład Alergologii
i Rehabilitacji Oddechowej, II Katedra Otolaryngologii, Wydział Wojskowo-Lekarski, Uniwersytet Medyczny w Łodzi, www.umed.lodz.pl 2 [email protected], Klinika Otolaryngologii, Onkologii Laryngologicznej, Audiologii i Foniatrii,
II Katedra Otolaryngologii, Wydział Wojskowo-Lekarski, Uniwersytet Medyczny w Łodzi,
www.umed.lodz.pl 3 [email protected], Katedra Mikrobiologii, Immunologii i Medycyny Laboratoryjnej,
Pomorski Uniwersytet Medyczny, www.pum.edu.pl, 4 [email protected], Zakład Biotechnologii Farmaceutycznej, Wydział Farmaceutyczny, Uniwersytet
Medyczny w Łodzi, www.umed.lodz.pl

Aktywność olejków eterycznych przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
93
Enterobacteriaceae wytwarzające betalaktamazy o ciągle rozszerzającym się spektrum
substratowym, w tym ESBL, a także karbapenemazy KPC, MBL (NDM-1) czy OXA-
48, które pojawiły się wraz z nadużywaniem zarówno karbapenemów, jak również
cefalosporyn III generacji i fluorochinolonów. Problem ten dotyczy głównie izolatów
klinicznych Klebsiella pneumoniae i Escherichia coli, a także K. oxytoca, Citrobacter
freundii, Enterobacter cloacae, Proteus mirabilis, Pseudomonas aeruginosa, czy
Acinetobacter spp [11-16]. Rosnące niebezpieczeństwo rozpowszechniania się
antybiotykooporności wiąże się z jednej strony ze zdolnościami bakterii do inaktywo-
wania antybiotyków, a z drugiej wynika z nadużywania antybiotykoterapii. Rozwój
turystyki sprzyja przemieszczaniu się opornych bakterii z odległych regionów geogra-
ficznych. Z kolei pacjenci są źródłem regionalnych epidemii związanych z przeno-
szeniem bakterii między placówkami medycznymi. Transmisja zakażeń związana jest
również z brakiem należytej staranności przy przestrzeganiu obowiązujących stan-
dardów przeciwzakażeniowych w szpitalach i innych placówkach medycznych
i opiekuńczych.
3. Olejki eteryczne – skład, właściwości, zalety
Olejki eteryczne są to mieszaniny związków chemicznych, będących produktami
metabolizmu wtórnego roślin. Rośliny wytwarzają i magazynują je w różnorodnych
strukturach anatomicznych jak: włoski gruczołowe, komórki olejkowe, zbiorniki
wydzielnicze czy kanały olejkowe. Olejki roślinne zawierają od kilkudziesięciu do
kilkuset składników, są to głównie mono- i seskwiterpenowe węglowodory oraz ich
tlenowe pochodne, a także fenylopropanoidy [17]. Skład olejku eterycznego zależny
jest od rodziny, rodzaju, gatunku i chemotypu, do którego należy roślina, a także od
warunków uprawy, pory zbioru i pochodzenia geograficznego. Charakteryzują się
złożonym, trudnym do odtworzenia składem chemicznym, co niestety jest jednym
z głównych powodów sceptycznego podejścia klinicystów do ich stosowania. Jednak
z drugiej strony różnorodność składników powoduje ich szerokie i kompleksowe
działanie, w tym przeciwdrobnoustrojowe, immunostymulujące, a także przeciw-
zapalne i przeciwbólowe, a nawet przeciwnowotworowe [18]. Prawdopodobnie ze
względu na unikatowy i bogaty skład chemiczny drobnoustroje, w tym oporne na
antybiotyki izolaty kliniczne są wrażliwe na działanie olejków eterycznych. Literatura
opisuje, że wielolekooporność nie warunkuje zmniejszonej wrażliwości na olejki
eteryczne. Synergizm działania poszczególnych składników olejku zależy nie tylko od
zawartości związków dominujących, ale także od komponentów, których zawartość
procentowa jest mniejsza. Ma to wpływ na specyficzne właściwości olejków eterycz-
nych jako naturalnych produktów roślinnych. Wiele cennych olejków eterycznych pod
względem właściwości przeciwinfekcyjnych, to surowce farmakopealne umieszczone
w Farmakopei Europejskiej, Farmakopei Polskiej, a także w rodzimych farmakopeach
krajów z różnych regionów Świata. Normy ISO i WHO zawierają również niezbędne
informacje na temat źródeł ich pozyskiwania, wymaganej zawartości składników
aktywnych, a także dawek terapeutycznych. Należy zauważyć, że od kiedy przetwory
roślinne zostały prawnie uznane za środki lecznicze zgodnie z artykułem I Dyrektywy

Monika Sienkiewicz, Hanna Zielińska-Bliźniewska, Jurek Olszewski, Paweł Kwiatkowski, Ewa Kochan
94
Rady European Economic Community oraz Comission for Proprietary Medicinal
Products wiele ośrodków naukowych żywo zajmuje się badaniem aktywności
metabolitów roślinnych, w tym także olejków eterycznych. Jest to w dużej mierze
związane z ciągłą potrzebą poszukiwań związków aktywnych o możliwie niewielkich
działaniach ubocznych, a także ze świadomością niespełnionych nadziei jakie wiązano
z lekiem syntetycznym. Z kolei dzięki nowym możliwościom jakie przyniósł rozkwit
biologii molekularnej, biochemii, fitochemii, czy farmakologii możliwe są szczegół-
łowe badania aktywności, mechanizmu działania i efektów ubocznych stosowania
związków naturalnych. Badania nad mechanizmem działania olejków eterycznych
dowiodły, że ich właściwości przeciwdrobnoustrojowe zależą od lipofilowego charak-
terem szkieletu węglowodorowego i hydrofilowego charakteru grup funkcyjnych.
Stwierdzono, że lipofilowy charakter monoterpenów powoduje rozluźnienie struktury
błon, wzrost płynności i przepuszczalności, zaburzenia funkcjonowania białek
membranowych, procesów oddychania i transportu jonowego [19,20]. Należy zwrócić
uwagę, że w dostępnym piśmiennictwie naukowym można odnaleźć wiele pozycji
literatury opisującej właściwości biologiczne olejków eterycznych, szczególnie
dotyczące aktywności przeciwdrobnoustrojowej, jednak w większości są to badania
wobec szczepów wzorcowych. Aktywność wobec wielolekoopornych bakterii
wyizolowanych z przypadków klinicznych i środowiska szpitalnego pod kątem
zastosowania w preparatach handlowych wymaga udokumentowania. Olejki cieszą się
również dużym zainteresowaniem jako alternatywa dla sztucznych konserwantów
żywności. W publikowanych badaniach dotyczących działania przeciwdrobno-
ustrojowego olejków eterycznych używanych jest szereg metod, nie wszystkie z nich
można uznać za dokładne i możliwe do porównania. Bardzo często w artykułach brak
również informacji o składzie badanego olejku eterycznego. Badania pod kątem
aplikacyjnym powinny obejmować nie tylko aktywność przeciwdrobnoustrojową, ale
także jednoczesną analizę składu chemicznego olejku by można go było zweryfikować
z obowiązującymi normami [21]. Wśród światowych opracowań patentowych pojawia
się coraz więcej patentów dotyczących aktywności przeciwdrobnoustrojowej olejków
eterycznych, możliwości ich zastosowania w medycynie, przemyśle spożywczym
i kosmetycznym [22]. Badania własne nad aktywnością wybranych olejków eterycz-
nych dowodzą, że oprócz powszechnie znanych zastosowań olejków w leczeniu
schorzeń infekcyjnych układu oddechowego (olejek z drzewa herbacianego, tymian-
kowy, eukaliptusowy, olejki eteryczne z drzew iglastych) czy układu pokarmowego
i dróg żółciowych (olejki miętowe, olejek kminkowy, imbirowy, manuka, tatarakowy,
rumiankowy) olejki eteryczne mogą być szczególne przydatne w zakażeniach derma-
tologicznych, głównie w leczeniu nawracających przypadków trudno gojących się ran
i owrzodzeń wywoływanych przez bakterie oporne na rekomendowane antybiotyki.
Bardzo często zakażenia ran pooperacyjnych ze względu na brak skutecznej terapii
zwłaszcza u osób z chorobami współistniejącymi jak np. cukrzyca czy u chorych
z niedoborami odporności niweczą efekty leczenia operacyjnego, a także są przyczyną
groźnych powikłań włącznie z rozwinięciem się zakażeń uogólnionych i zgonami.
Zakażenia pooperacyjne tkanek miękkich są również przyczyną trwałego kalectwa

Aktywność olejków eterycznych przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
95
i niezdolności do pracy. W praktyce klinicznej stosuje się więc nagminnie antybiotyki,
co jest przyczyniają stale rosnącej oporności i prowadzi do „wyczerpania” możliwości
skutecznego leczenia zakażeń. Dodatkowo nadużywanie antybiotyków układowych
może prowadzić do szkodliwych działań ubocznych zwłaszcza u osób o obniżonej
odporności, osób starszych, długo hospitalizowanych. Z kolei antybiotyki stosowane
miejscowo nie są zalecane w większości wytycznych, ponieważ mogą wywołać reak-
cje nadwrażliwości typu późnego i nadkażenia [23]. Środki przeciwbakteryjne takie jak
alkohole, bisfenole, podchloryn sodu, związki srebra i związki jodu stosuje się głównie
do zapobiegania występowaniu infekcji, niektóre wykazują również wysoką cytoto-
ksyczność in vitro [24, 25]. Szukamy więc wciąż nowych skutecznych środków
przeciwinfekcyjnych jako alternatywnych bądź zwiększających aktywność tych już
stosowanych. Olejki eteryczne stosowane są w aromaterapii jednak ich właściwości
biologiczne pod kątem szerszego zastosowania klinicznego wymagają badań klinicz-
nych. Zgodnie z danymi literatury dla większości olejków eterycznych wartości LD50
mieszczą się powyżej 5 g/kg, natomiast dawki terapeutyczne wynoszą zwykle kilka
kropli na dobę [26]. Wiadomo, że olejki mogą być stosowane na skórę w postaci
maści, emulsji, balsamów czy żeli. Zaleta olejków eterycznych, w stosunku do innych
środków przeciwdrobnoustrojowych, polega na tym, że oferują szeroką skuteczność
przeciwbakteryjną nie indukując mechanizmów oporności. Ponadto, badania
wykazały, że ich aktywność farmakologiczna jest bardzo szeroka, szczególnie cenne
jest ich działanie przeciwzapalne, przyśpieszające gojenie, immunostymulujące czy
przeciwnowotworowe [18, 27, 28]. Zgodnie z doniesieniami literatury produkty
roślinne, a wśród nich olejki eteryczne mogą okazać się interesujące również jako
czynniki hamujące powstawanie biofilmu bakteryjnego [29]. Z kolei Edrys [18],
sugeruje, że olejki eteryczne i ich składniki terpenowe przy zastosowaniu miejscowym
mogą stanowić alternatywne, naturalne środki polepszające penetrację skóry. Ich
toksyczność jest dobrze udokumentowana, i stosunkowo mała w porównaniu
z większością syntetycznych środków zwiększających przenikanie [30,31].
4. Badania własne nad aktywnością przeciwbakteryjną wybranych
olejków eterycznych
Celem podjętych badań była ocena aktywności przeciwbakteryjnej wybranych
olejków eterycznych wobec bakterii odpowiedzialnych za zakażenia dermatologiczne
o znacznym stopniu oporności na rekomendowane antybiotyki.
Badaniami objęto olejki eteryczne:
cynamonowy z kory,
geraniowy,
lawendowy,
bazylikowy,
rozmarynowy,
z szałwii muszkatołowej,
tymiankowy,
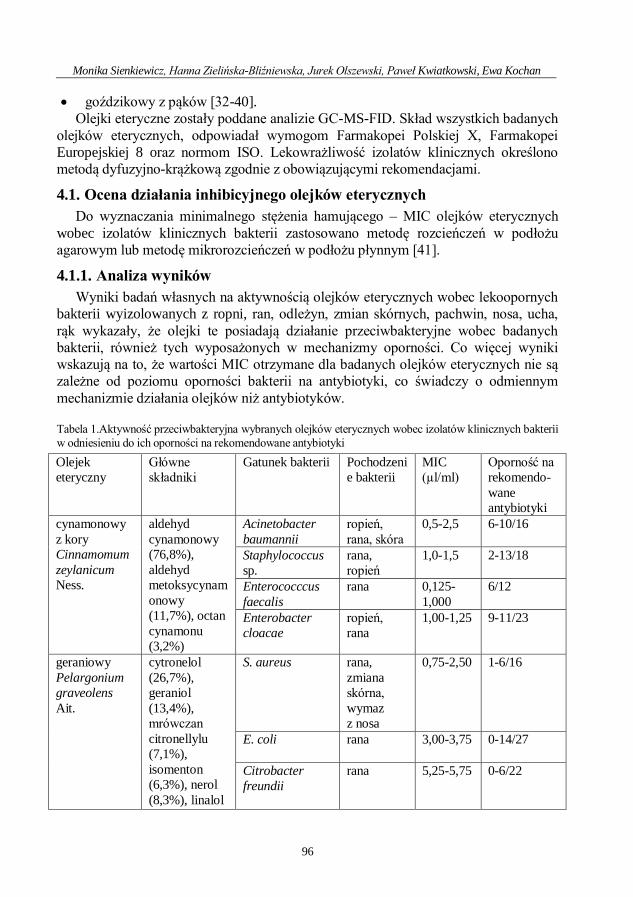
Monika Sienkiewicz, Hanna Zielińska-Bliźniewska, Jurek Olszewski, Paweł Kwiatkowski, Ewa Kochan
96
goździkowy z pąków [32-40].
Olejki eteryczne zostały poddane analizie GC-MS-FID. Skład wszystkich badanych
olejków eterycznych, odpowiadał wymogom Farmakopei Polskiej X, Farmakopei
Europejskiej 8 oraz normom ISO. Lekowrażliwość izolatów klinicznych określono
metodą dyfuzyjno-krążkową zgodnie z obowiązującymi rekomendacjami.
4.1. Ocena działania inhibicyjnego olejków eterycznych
Do wyznaczania minimalnego stężenia hamującego – MIC olejków eterycznych
wobec izolatów klinicznych bakterii zastosowano metodę rozcieńczeń w podłożu
agarowym lub metodę mikrorozcieńczeń w podłożu płynnym [41].
4.1.1. Analiza wyników
Wyniki badań własnych na aktywnością olejków eterycznych wobec lekoopornych
bakterii wyizolowanych z ropni, ran, odleżyn, zmian skórnych, pachwin, nosa, ucha,
rąk wykazały, że olejki te posiadają działanie przeciwbakteryjne wobec badanych
bakterii, również tych wyposażonych w mechanizmy oporności. Co więcej wyniki
wskazują na to, że wartości MIC otrzymane dla badanych olejków eterycznych nie są
zależne od poziomu oporności bakterii na antybiotyki, co świadczy o odmiennym
mechanizmie działania olejków niż antybiotyków.
Tabela 1.Aktywność przeciwbakteryjna wybranych olejków eterycznych wobec izolatów klinicznych bakterii
w odniesieniu do ich oporności na rekomendowane antybiotyki
Olejek
eteryczny
Główne
składniki
Gatunek bakterii Pochodzeni
e bakterii
MIC
(µl/ml)
Oporność na
rekomendo-
wane
antybiotyki
cynamonowy
z kory
Cinnamomum
zeylanicum
Ness.
aldehyd
cynamonowy
(76,8%),
aldehyd
metoksycynam
onowy
(11,7%), octan
cynamonu
(3,2%)
Acinetobacter
baumannii
ropień,
rana, skóra
0,5-2,5 6-10/16
Staphylococcus
sp.
rana,
ropień
1,0-1,5 2-13/18
Enterococccus
faecalis
rana 0,125-
1,000
6/12
Enterobacter
cloacae
ropień,
rana
1,00-1,25 9-11/23
geraniowy
Pelargonium
graveolens
Ait.
cytronelol
(26,7%),
geraniol
(13,4%),
mrówczan
citronellylu
(7,1%),
isomenton
(6,3%), nerol
(8,3%), linalol
S. aureus rana,
zmiana
skórna,
wymaz
z nosa
0,75-2,50 1-6/16
E. coli
rana 3,00-3,75 0-14/27
Citrobacter
freundii
rana 5,25-5,75 0-6/22
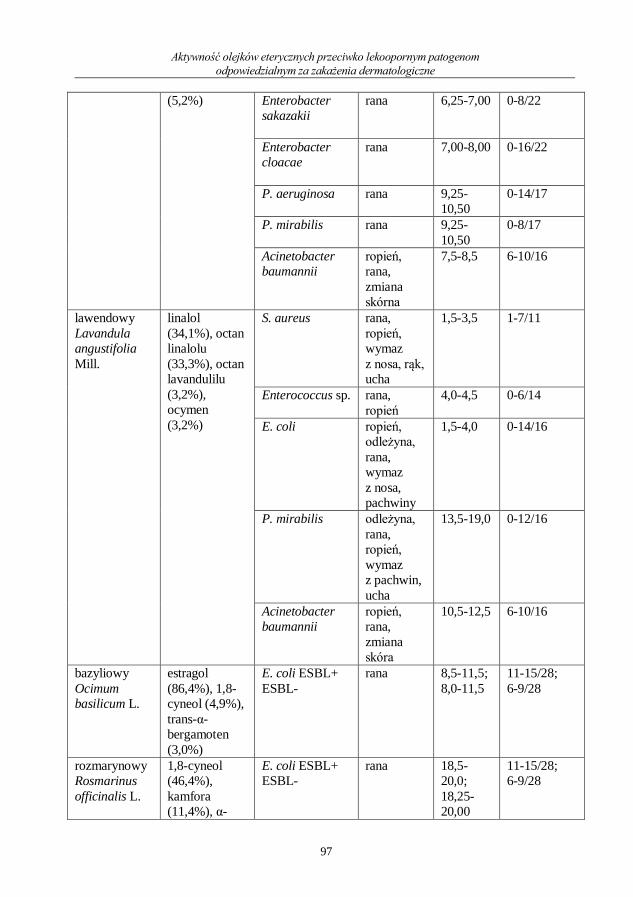
Aktywność olejków eterycznych przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
97
(5,2%) Enterobacter
sakazakii
rana 6,25-7,00 0-8/22
Enterobacter
cloacae
rana 7,00-8,00 0-16/22
P. aeruginosa rana 9,25-
10,50
0-14/17
P. mirabilis rana 9,25-
10,50
0-8/17
Acinetobacter
baumannii
ropień,
rana,
zmiana
skórna
7,5-8,5 6-10/16
lawendowy
Lavandula
angustifolia
Mill.
linalol
(34,1%), octan
linalolu
(33,3%), octan
lavandulilu
(3,2%),
ocymen
(3,2%)
S. aureus rana,
ropień,
wymaz
z nosa, rąk,
ucha
1,5-3,5 1-7/11
Enterococcus sp. rana,
ropień
4,0-4,5 0-6/14
E. coli ropień,
odleżyna,
rana,
wymaz
z nosa,
pachwiny
1,5-4,0 0-14/16
P. mirabilis odleżyna,
rana,
ropień,
wymaz
z pachwin,
ucha
13,5-19,0 0-12/16
Acinetobacter
baumannii
ropień,
rana,
zmiana
skóra
10,5-12,5 6-10/16
bazyliowy
Ocimum
basilicum L.
estragol
(86,4%), 1,8-
cyneol (4,9%),
trans-α-
bergamoten
(3,0%)
E. coli ESBL+
ESBL-
rana 8,5-11,5;
8,0-11,5
11-15/28;
6-9/28
rozmarynowy
Rosmarinus
officinalis L.
1,8-cyneol
(46,4%),
kamfora
(11,4%), α-
E. coli ESBL+
ESBL-
rana 18,5-
20,0;
18,25-
20,00
11-15/28;
6-9/28
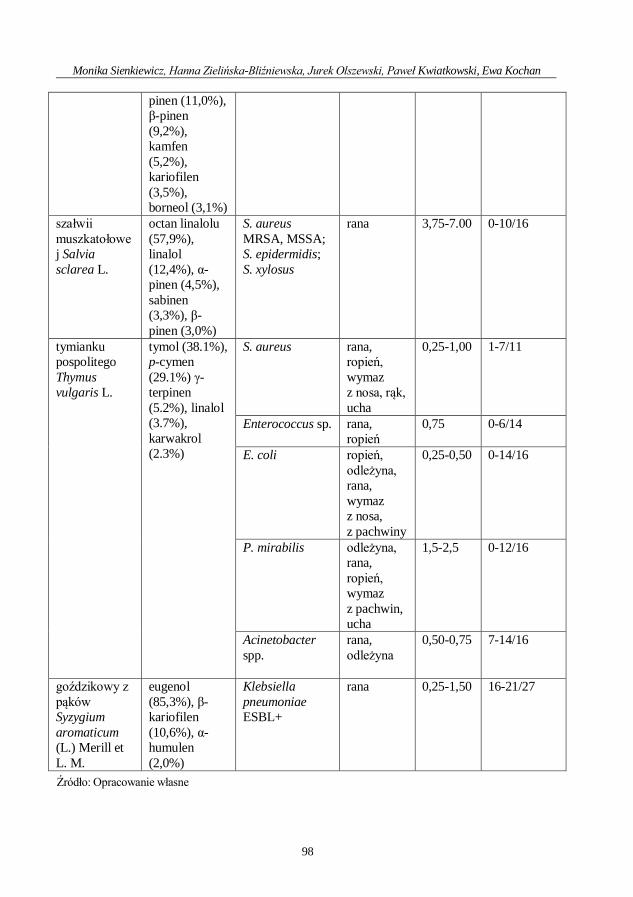
Monika Sienkiewicz, Hanna Zielińska-Bliźniewska, Jurek Olszewski, Paweł Kwiatkowski, Ewa Kochan
98
pinen (11,0%),
β-pinen
(9,2%),
kamfen
(5,2%),
kariofilen
(3,5%),
borneol (3,1%)
szałwii
muszkatołowe
j Salvia
sclarea L.
octan linalolu
(57,9%),
linalol
(12,4%), α-
pinen (4,5%),
sabinen
(3,3%), β-
pinen (3,0%)
S. aureus
MRSA, MSSA;
S. epidermidis;
S. xylosus
rana 3,75-7.00 0-10/16
tymianku
pospolitego
Thymus
vulgaris L.
tymol (38.1%),
p-cymen
(29.1%) γ-
terpinen
(5.2%), linalol
(3.7%),
karwakrol
(2.3%)
S. aureus rana,
ropień,
wymaz
z nosa, rąk,
ucha
0,25-1,00 1-7/11
Enterococcus sp. rana,
ropień
0,75 0-6/14
E. coli ropień,
odleżyna,
rana,
wymaz
z nosa,
z pachwiny
0,25-0,50 0-14/16
P. mirabilis odleżyna,
rana,
ropień,
wymaz
z pachwin,
ucha
1,5-2,5 0-12/16
Acinetobacter
spp.
rana,
odleżyna
0,50-0,75 7-14/16
goździkowy z
pąków
Syzygium
aromaticum
(L.) Merill et
L. M.
eugenol
(85,3%), β-
kariofilen
(10,6%), α-
humulen
(2,0%)
Klebsiella
pneumoniae
ESBL+
rana 0,25-1,50 16-21/27
Źródło: Opracowanie własne

Aktywność olejków eterycznych przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
99
Wyniki badań własnych dowiodły, że najsilniejsze właściwości inhibicyjne posia-dają olejki tymiankowy i goździkowy z pąków – wartości MIC dla izolatów klinicz-nych wszystkich badanych bakterii wahały się w granicach od 0,25 do 2,5 μl/ml. Z pewnością tak wysoka aktywność przeciwbakteryjna tych olejków związana jest z dużą zawartością związków fenolowych jak tymol, eugenol i karwakrol. Wartości MIC dla olejku cynamonowego z kory zwierającego głównie aldehyd cynamonowy oraz metoksycynamonowy kształtowały się na poziomie 0,125-2,5 μl/ml. Dobrymi właściwościami inhibicyjnymi wobec opornych Gram-dodatnich i Gram-ujemnych bakterii charakteryzowały się również olejki eteryczne zawierające alkohole i alde-hydy, w tym linalol, cytronelol, geraniol, jak olejek lawendowy czy geraniowy. Olejek geraniowy hamował wzrost badanych izolatów w stężeniach od 0,75 do 10,50 μl/ml, najwyższe wartości MIC wyznaczono dla izolatów Pseudomonas aeruginosa i Proteus mirabilis. Dobrą aktywność przeciwbakteryjną wykazywał również olejek z szałwii muszkatołowej wobec wyizolowanych z ran trzech gatunków gronkowców, w tym S. aureus MRSA – MIC (3,75-7,00 μl/ml). Nieco słabsze działanie hamujące wykazały olejki bogate w węglowodory jedno i dwupierścieniowe, choć były one aktywne również wobec izolatów Escherichia coli produkujących betalaktamazy o rozsze-rzonym spektrum substratowym – ESBL.
4.2. Ocena działania synergistycznego i addycyjnego połączeń olejków z rekomendowanymi antybiotykami
W badaniach sprawdzono również potencjał aktywności synergistycznej i addy-cyjnej wybranych farmakopealnych, spełniających normy ISO, olejków eterycznych w połączeniu z rekomendowanymi antybiotykami. W tym celu zastosowano metodę dyfuzyjno-krążkową. Badania były prowadzone wobec bakterii wyizolowanych z ran od pacjentów z nawracającymi infekcjami dermatologicznymi. Były to izolaty kli-niczne: Staphylococcus aureus, Enterococccus faecalis, Escherichia coli, Klebsiella pneumoniae, Enterobacter clacae i Acinetobacter baumannii [42].
4.2.1. Analiza wyników
Wyniki prac pozwoliły na stwierdzenie, że olejki eteryczne: cynamonowy, goź-dzikowy, tymiankowy, geraniowy i lawendowy mają silne właściwości inhibicyjne i bakteriobójcze, jednak olejki: bazyliowy, z szałwii muszkatołowej i rozmarynowy wykazują najlepsze działanie addycyjne i synergistyczne w połączeniu z reko-mendowanymi antybiotykami.
Wyniki przeprowadzonych badań wykazały, że olejek rozmarynowy posiadał najsilniejsze właściwości zwiększające aktywność antybiotyków wobec opornych bakterii. Pod wpływem tego olejku stwierdzono 70% zwiększenie aktywności antybiotyków wobec S. aureus oraz 40% polepszenie ich skuteczności wobec E. coli. Z kolei olejek bazyliowy w połączeniu z rekomendowanymi antybiotykami wzmagał ich aktywność o około 50% wobec S. aureus i Acinetobacter baumannii oraz o ponad 60% wobec E. coli. Natomiast olejek eteryczny z szałwii muszkatołowej powodował polepszenie/zwiększenie aktywności antybiotyków o około 40% wobec S. aureus i E. faecalis.
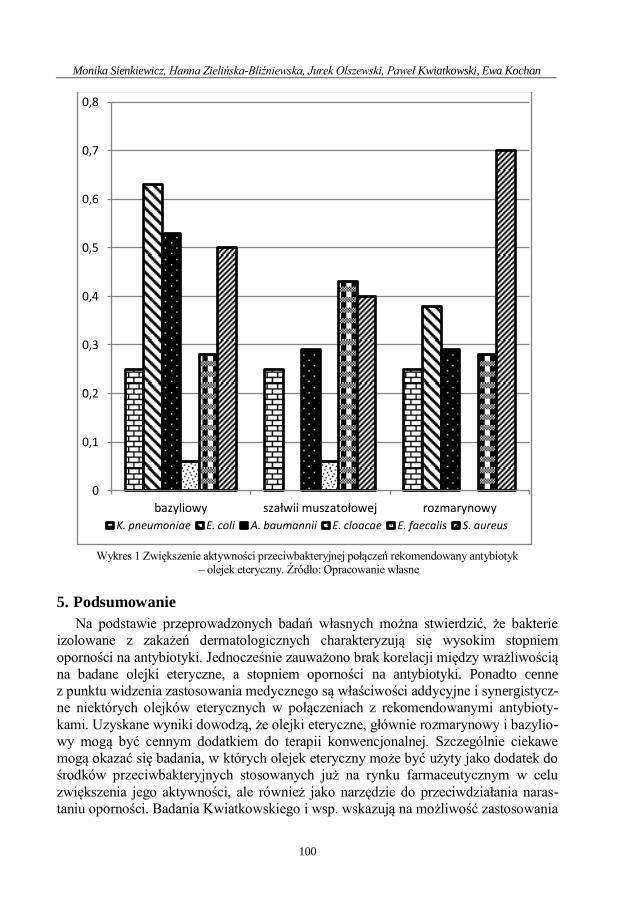
Monika Sienkiewicz, Hanna Zielińska-Bliźniewska, Jurek Olszewski, Paweł Kwiatkowski, Ewa Kochan
100
Wykres 1 Zwiększenie aktywności przeciwbakteryjnej połączeń rekomendowany antybiotyk
– olejek eteryczny. Źródło: Opracowanie własne
5. Podsumowanie
Na podstawie przeprowadzonych badań własnych można stwierdzić, że bakterie
izolowane z zakażeń dermatologicznych charakteryzują się wysokim stopniem
oporności na antybiotyki. Jednocześnie zauważono brak korelacji między wrażliwością
na badane olejki eteryczne, a stopniem oporności na antybiotyki. Ponadto cenne
z punktu widzenia zastosowania medycznego są właściwości addycyjne i synergistycz-
ne niektórych olejków eterycznych w połączeniach z rekomendowanymi antybioty-
kami. Uzyskane wyniki dowodzą, że olejki eteryczne, głównie rozmarynowy i bazylio-
wy mogą być cennym dodatkiem do terapii konwencjonalnej. Szczególnie ciekawe
mogą okazać się badania, w których olejek eteryczny może być użyty jako dodatek do
środków przeciwbakteryjnych stosowanych już na rynku farmaceutycznym w celu
zwiększenia jego aktywności, ale również jako narzędzie do przeciwdziałania naras-
taniu oporności. Badania Kwiatkowskiego i wsp. wskazują na możliwość zastosowania
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
bazyliowy szałwii muszatołowej rozmarynowy
K. pneumoniae E. coli A. baumannii E. cloacae E. faecalis S. aureus

Aktywność olejków eterycznych przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
101
olejku z kopru włoskiego jako dodatku do preparatów zawierających mupirocynę
stosowanych w eradykacji nosicielstwa MRSA [43]. Wskazane są więc dalsze badania
nad aktywnością przeciwdrobnoustrojową oraz nad cytotoksycznością olejków
eterycznych, które częściowo również przeprowadziliśmy w odniesieniu do olejku
cynamonowego, geraniowego i lawendowego. W kierunku aplikacyjnym konieczne są
szerokie badania w warunkach in vivo mające na celu ustalenie dawek, formy i często-
ści podania preparatów zawierających olejki eteryczne. Ze względu na problemy
w leczeniu zakażeń ran, owrzodzeń, a także malejącą skuteczność antybiotyków i che-
micznych preparatów antyseptycznych przeznaczonych do stosowania miejscowego
niniejsze badania wskazują na możliwość użycia olejków jako cennych dodatków do
leczenia konwencjonalnego, pozwalających zmniejszyć skuteczne dawki antybio-
tyków, a także jako dodatek do środków antyseptycznych i dezynfekcyjnych.
Podziękowania
Autorzy serdecznie dziękują Pani prof. dr hab. inż. Danucie Kalembie za przepro-
wadzenie analizy chemicznej badanych olejków eterycznych i pomoc naukową przy
pracach dotyczących badań własnych opisanych w niniejszej monografii.
Literatura
1. Lee Ventola C. The antibiotic resistance crisis Part 1. Causes and threats, Pharmacy and
Therapeutics., 40 (2015), s. 277-283.
2. Chastre J. Evolving problems with resistant pathogens, Clinical Microbiology and
Infections., 14 (2008), s. 3-14.
3. DeLeo F.R., Otto M., Kreiswirth B.N., Chambers H.N. Community-associated
methicillinresistant Staphylococcus aureus, Lancet., 375 (2010), s. 1557-1568.
4. Boers S.A., van Ess I., Euser S.M., Jansen R., Tempelman F.R., Diederen B.M. An
outbreak of a multiresistant methicillin-susceptible Staphylococcus aureus (MR-MSSA)
strain in a burn centre: The importance of routine molecular typing, Burns., 37 (2011), s.
808-813.
5. Wang J.T., Liao C.H., Fang C.T., Chie W.C., Lai M.S., Lauderdale T.L., Chang S.C.
Incidence of and risk factors for community-associated methicillin-resistant Staphylococcus
aureus acquired infection or colonization in intensive-care-unit patients, Journal of Clinical
Microbiology., 48 (2010), s. 4439-4444.
6. von Eiff C., Proctor R.A., Peters G. Coagulase-negative staphylococci. Pathogens have
major role in nosocomial infections, Prostgrad Medical Journal., 11 (2001), s. 63-76.
7. Nikam A.P., Bhise P.R., Deshmukh M.M. Phenotypic detection of inducible clindamycin
resistance among Staphylococcus aureus isolates, International Journal of Research in
Medical Science., 5 (2017), s. 543-547.
8. Shoji K., Shinjoh M., Horikoshi Y., Tang J., Watanabe Y., Sugita K., Tame T., Iwata S.,
Miyairi I., Saitoh A. High rate of inducible clindamycin resistance in Staphylococcus
aureus isolates–A multicenter study in Tokyo, Japan, Journal of Infection Chemotherapy.,
21 (2015), s. 81-3.
9. Ozansy F.A., Cevahir N., Kaleli I. Investigation of macrolide, lincosamide and
streptogramin B resistance in Staphylococcus aureus strains isolated from clinical samples
by phenotypic and genotypic methods, Mikrobyologji Bulteni., 49 (2015), s. 1-14.

Monika Sienkiewicz, Hanna Zielińska-Bliźniewska, Jurek Olszewski, Paweł Kwiatkowski, Ewa Kochan
102
10. Baldır G., Engin D.Ö., Küçükercan M., İnan A., Akçay S., Özyürek S., Aksaray S. High-
level resistance to aminoglycoside, vancomycin, and linezolid in enterococci strains,
Journal of Microbiology and Infectious Diseases., 3 (2013), s. 100-103.
11. Doi Y., Potoski B.A., Adams-Haduch J.M., Sidjabat H.E., Pasculle A.W., Paterson D.L.
Simple disk-based method for detection of Klebsiella pneumoniae carbapenemase-type β-
lactamase by use of a boronic acid compound, Journal of Clinical Microbiology., 46
(2008), s. 4083-4086.
12. Meyer E., Schwab F., Schroeren-Boersch B., Gastmeier P. Dramatic increase of thirdgene-
ration cephalosporin-resistant E. coli in German intensive care units: secular trends in
antibiotic drug use and bacterial resistance, 2001to 2008, Critic Care., 14 (2010), R113.
13. Thapa B., Adhikari P., Mahat K., Chhetri M.R., Joshi L.N. Multidrug-resistant nosocomial
Citrobacter in a Hospital in Kathmandu, Nepal Medical College Journal., 11 (2009),
s. 195-199.
14. Canton R., Novais A., Valverde A., Machado E., Peixe L., Baquero F., Coque T.M.
Prevalence and spread of extended-spectrum β-lactamase-producing Enterobacteriaceae
in Europe, Clinical Microbiology and Infection., 14 (2008), s. 144-153.
15. Cornaglia G., Garau J., Livermore D.M. Living with ESBLs. Clinical Microbiology and
Infection., 14 (2008) s. 1-2.
16. Hryniewicz W. Narodowy Program Ochrony Antybiotyków. Enterobacteriaceae jako
drobnoustroje alarmowe. www.antybiotyki.edu.pl
17. Kalemba D., Kunicka A. Essential Oils – the Powerful Weapon against Microorganisms,
Leopold Irovetz and Gerhard Buchbauer (Editors): Processing, Analysis and Aplication of
Essential Oils, Har Krishan Bhalla & Sons, Dehradum, India 2005.
18. Edris A.E. Pharmaceutical and Therapeutic Potentials of Essential Oils and Their
Individual Volatile Constituents: A Review, Phytotherapy Research., 21 (2007), s. 308-323.
19. Trombetta D., Castelli F., Sarpietro M.G., Venuti V., Cristani M., Daniele C., Saija A.,
Mazzanti G., Bisignano G. Mechanisms of antibacterial action of three monoterpenes,
Antimicrobial Agents and Chemotherapy., 49 (2005), s. 2474–2478.
20. Cristani M., D’arrigo M., Mandalari G., Castelli F., Sarpietro M.G., Micieli D., Venuti V.,
Bisignano G., Saija A., Trombetta D. Interaction of four monoterpenes contained in
essential oils with model membranes: implications for their antibacterial activity, Journal
of Agricultural and Food Chemistry., 55 (2007), s. 6300−6308.
21. van Vuuren S.F. Antimicrobial activity of South African medicinal plants, Journal of
Ethnopharmacology., 119 (2008), s. 462-472.
22. Sienkiewicz M., Kowalczyk E., Wasiela M. Recent Patents Regarding Essential Oils and
the Significance of their Constituents in Human Health and Treatment. Recent Patents on
Anti-Infective Drug Discovery., 7 (2012), s. 133-140.
23. Siddiqui A.R., Bernstein J.M. Chronic wound infection: Facts and controversies. Clinical
Dermatology., 28 (2010), s. 519-526.
24. Percival S.L., Bowler P.G., Russell D. Bacterial resistance to silver in wound care. Journal
of Hospital Infections., 60 (2005), s. 1-7.
25. Lipsky B.A., Hoey K. Topical antimicrobial therapy for treating chronic wounds, Clinical
infectious Diseases., 49 (2009), s.1541-1549.
26. Cal K. Olejki eteryczne w preparatach leczniczych – mechanizm działania i zastosowanie,
Aromaterapia., 33/34 (2003), s. 20-29.

Aktywność olejków eterycznych przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
103
27. Serafino A., Vallebona P.S., Andreola F., Zonfrillo M., Mercuri L., Federici M., Rasi G.,
Garaci E., Pierimarchi P. Stimulatory effect of Eucalyptus essential oil on innate cell
mediated immune response, BMC Immunology., 9 (2008), s. 17.
28. Cavanagh H.M.A., Wilkinson J.M. Lavender essential oil: review, Australian Infection
Control., 10 (2005), s.35-37.
29. Łysakowska M., Sienkiewicz M., Banaszek K., Sokołowski J. The sensitivity of endodontic
Enterococcus spp. strains to geranium essential oil, Molecules., 20 (2015), s. 22881-22889.
30. Reichling J., Schnitzler P., Suschke U., Saller R. Essential oils of aromatic plants with
antibacterial, antifungal, antiviral and cytotoxic properties - an oview. Forsch
Komplementment., 16 (2009), s. 79-90.
31. Bakkali F., Averbeck S., Averbeck D., Idaomar M. Biological effect of essential oils –
A review, Food Chemistry and Toxicology., 46 (2008), s. 446-475.
32. Sienkiewicz M., Głowacka A., Kowalczyk E., Wiktorowska-Owczarek A., Jóźwiak-
Bębenista M., Łysakowska M. The biological activities of cinnamon, geranium and
lavender essential oils, Molecules., 19 (2014), s. 20929-20940.
33. Sienkiewicz M., Łysakowska M., Pastuszka M., Bienias W, Kowalczyk E, The potential of
use basil and rosemary essential oils as effective antibacterial agents, Molecules., 18
(2013), s. 9334-9351.
34. Sienkiewicz M., Głowacka A., Poznańska-Kurowska K., Kaszuba K., Urbaniak A.,
Kowalczyk K. The effect of clary sage oil on staphylococci responsible of wounds
infections, Postępy Dermatologii i Alergologii., XXXII (2015), s. 21-25.
35. Łysakowska M., Denys A., Sienkiewicz M. Antimicrobial activity of thyme essential oil
against clinical strains of Acinetobacter genus, Central European Journal of Biology., 6
(2012), s. 405-413.
36. Sienkiewicz M., Kalemba D., Wasiela M. The susceptibility of Klebsiella pneumoniae
ESBL+ strains to clove (Syzygium aromaticum) essential oil, Anti-Infective Agents., 11
(2012), s. 159-167.
37. Urbaniak A., Głowacka A., Kowalczyk E., Łysakowska M., Sienkiewicz M.
Przeciwbakteryjne działanie olejku cynamonowego na wybrane bakterie Gram-dodatnie i
Gram-ujemne, Medycyna Doświadczalna i Mikrobiologia., 66 (2014), s. 131-141.
38. Sienkiewicz M., Łysakowska M., Ciećwierz J., Denys P., Kowalczyk E. Antibacterial
activity of thyme and lavender essential oils, Medicinal Chemistry., 7 (2011), s. ,674-689.
39. Bigos M., Wasiela M., Kalemba D., Sienkiewicz M. Antimicrobial activity of geranium oil
against Staphylococcus aureus clinical strains, Molecules., 17 (2012), s. 10276-10291.
40. Sienkiewicz M., Poznańska-Kurowska K., Kaszuba A., Kowalczyk E. The antibacterial
activity of geranium oil against Gram-negative bacteria isolated from difficult-to-heal
wounds, Burns., 40 (2013), s. 1046-1051.
41. Kalemba D., Kunicka A. Antibacterial and antifungal properties of essential oils, Current
Medicinal Chemistry., 10 (2003), s. 813-829.
42. Sienkiewicz M., Łysakowska M., Kowalczyk E., Szymańska G., Kochan E., Krukowska J.,
Olszewski J., Zielińska-Bliźniewska H. The ability of selected plant essential oils to
enhance the action of recommended antibiotics against pathogenic wound bacteria, Burns.,
43 (2017), s. 310-317.
43. Kwiatkowski P., Mnichowska-Polanowska M., Pruss A., Masiuk H., Dzięcioł M., Giedryś-
Kalemba S., Sienkiewicz M. Evaluation of the effect of the combination between choice
antibiotics and fennel essential oil on Staphylococcus aureus strains isolated from carriers,
Burns., (in press).

Monika Sienkiewicz, Hanna Zielińska-Bliźniewska, Jurek Olszewski, Paweł Kwiatkowski, Ewa Kochan
104
Aktywność olejków eterycznych przeciwko lekoopornym patogenom
odpowiedzialnym za zakażenia dermatologiczne
Streszczenie Problem szerzenia się lekooporności zmusza do poszukiwań nowych, efektywnych i bezpiecznych
środków mogących mieć zastosowanie w zwalczaniu uporczywych, trudnych w terapii infekcji. Zgodnie
z danymi literatury produkty roślinne, w tym olejki eteryczne są interesujące nie tylko ze względu na ich
działanie przeciwmikrobowe i zdolność do hamowania tworzenia się biofilmu bakteryjnego, ale także ze względu na ich właściwości przeciwzapalne i immunostymulujące. W badaniach własnych sprawdziliśmy
działanie inhibicyjne, a także synergistyczne i addycyjne w połączeniu z rekomendowanymi antybiotykami
olejków: tymiankowego, goździkowego, cynamonowego, geraniowego, lawendowego, szałwii muszka-
tołowej, bazylikowego i rozmarynowego. W badaniach aktywności inhibicyjnej używaliśmy metody roz-cieńczeń w podłożu agarowym i metody mikrorozcieńczeń w podłożu płynnym. Właściwości syner-
gistyczne i addycyjne były określane za pomocą metody dyfuzyjno-krążkowej. Lekooporność izolatów
klinicznych określano metodą dyfuzyjno-krążkową zgodnie z rekomendacjami. Testowane olejki eteryczne
były analizowane metodą GC-MS-FID, ich skład odpowiadał wymogom Farmakopei Polskiej X, Farma-kopei Europejskiej 8 oraz normom ISO. Olejki eteryczne: tymiankowy, goździkowy, cynamonowy, gera-
niowy, i lawendowy wykazywały najsilniejsze działanie przeciwbakteryjne, również wobec bakterii dys-
ponujących mechanizmami oporności. Najlepszą zdolność do aktywności synergistycznej i addycyjnej
posiadały olejki: rozmarynowy, bazyliowy i szałwii muszkatołowej wobec badanych bakterii odpo-wiedzialnych za zakażenia dermatologiczne. Biorąc pod uwagę znaczny wzrost oporności bakterii na
powszechnie stosowane środki przeciwbakteryjne, nasze prace wskazują na potencjału olejków ete-
rycznych, jako związków naturalnych mogących mieć powszechne zastosowanie medyczne w zapo-
bieganiu i w terapii schorzeń infekcyjnych, w tym w zwalczaniu opornych bakterii odpowiedzialnych za
zakażenia dermatologiczne.
Słowa kluczowe: aktywność przeciwbakteryjna; lekooporność; olejki eteryczne; działanie hamujące;
synergistyczne, addycyjne.
Activity of essential oils against multidrug resistant pathogens caused
by dermatological infections
Abstract
The problem of increasing multidrug resistance forces us to look for safe and effective agents that could be
used to treat persistent bacterial infections. According to items of literature, plant products, including essential oils seem to be interesting not only due to their significant antimicrobial activity, and ability to
inhibit the formation of bacterial biofilm, but also because of their anti-inflammatory and immune-
stimulatory effects. In own research we checked the inhibiting activity and also synergistic and additive
properties with recommended antibiotics of thyme, clove, cinnamon, geranium, lavender, clary sage, basil and rosemary essential oils against bacteria isolated from dermatological infections. In investigations of
inhibiting activity we used of dilution method in agar medium and microdilution method in liquid medium.
Synergistic and additive properties were assessed using the disc-diffusion method. In the studies of drug
resistance of clinical isolates were applied the disc-diffusion method in accordance with the recommendations. The tested essential oils were analysed using GC-MS-FID and their composition was
consistent with the requirements of the Polish Pharmacopoeia X, the European Pharmacopoeia and ISO
standards. Thyme, clove, cinnamon, geranium and lavender oils showed the greatest antibacterial activity
against bacteria including those with resistance mechanisms. The best ability to synergistic and additive action with recommended antibiotics possessed rosemary, basil and clary sage oils against bacteria isolated
from dermatological infections. Considering the significant increase in bacterial resistance to commonly
used antibacterial agents, our works presented the potential of essential oils as natural compounds that may
have widespread medical use in the prevention and treatment of infectious diseases including in fighting against resistant bacteria caused by dermatological infections.
Keywords: antibacterial activity; drug resistance, essential oils, inhibiting activity; synergistic, additive
activity.

105
Michał Piotrowski1, Hanna Pituch
2, Piotr Obuch-Woszczatyński
3
Działanie miodu manuka na wzrost oraz biofilm
wybranych bakterii stanowiących problem
terapeutyczny
1. Wprowadzenie
W ostatnich latach notuje się zwiększoną zachorowalność na choroby infekcyjne
powodowane przez drobnoustroje oporne na antybiotyki. Nadmierne stosowanie
antybiotyków prowadzi do powstawania szczepów bakterii wielolekoopornych. To
zjawisko ogranicza możliwości terapeutyczne i wymusza potrzebę poszukiwania
alternatywnych środków o działaniu przeciwbakteryjnym Popularna staje się apiterapia
– wykorzystywanie produktów pszczelich, przede wszystkim miodu w leczeniu róż-
nych schorzeń [1]. Miody posiadają właściwości antyoksydacyjne oraz są naturalnymi
środkami biologicznie czynnymi, aktywnie działającymi na niektóre drobnoustroje.
Zastosowanie miodu w leczeniu różnych schorzeń o podłożu infekcyjnym sięga
czasów starożytnych. W ostatnich latach bardzo intensywnie prowadzi się badania nad
miodem manuka, pochodzącym z Nowej Zelandii. Swoje właściwości antybiotyczne
zawdzięcza on zawartości metyloglioksalu (MGO) [1].
2. Charakterystyka oraz właściwości antybakteryjne miodu manuka
Miód składa się z substancji organicznych i nieorganicznych. W zależności od
odmiany można wyróżnić ponad 300 składników. Około 70-80% masy miodu stano-
wią naturalne cukry: glukoza i fruktoza. Stosunek tych dwóch cukrów waha się
w zależności od odmiany miodu, większa ilość fruktozy powoduje wolniejszą krysta-
lizację miodu. Duża zawartość cukrów w miodzie warunkuje wysokie ciśnienie osmo-
tyczne co jest jednym z czynników bakteriobójczych. Kwaśne pH miodu (3,8-4,9)
spowodowane obecnością kwasów organicznych hamuje rozwój wielu mikro-
organizmów [2].
Szczególnie silne właściwości przeciwdrobnoustrojowe posiadają miody zawie-
rające związki flawonoidowe. Natomiast miody z zawartością garbników katechu-
nowych odznaczają się działaniem przeciwgrzybicznym. Ważną grupą substancji
warunkujących właściwości antybiotyczne miodów są terpeny i seskwiterpeny.
Związki te występują w miodach produkowanych ze spadzi drzew iglastych oraz
1 [email protected], Katedra i Zakład Mikrobiologii Lekarskiej Warszawskiego Uniwersytetu
Medycznego, https://mikrobiologia.wum.edu.pl/ 2 [email protected], Katedra i Zakład Mikrobiologii Lekarskiej Warszawskiego Uniwersytetu
Medycznego, https://mikrobiologia.wum.edu.pl/ 3 [email protected], Katedra i Zakład Mikrobiologii Lekarskiej Warszawskiego
Uniwersytetu Medycznego, https://mikrobiologia.wum.edu.pl/

Michał Piotrowski, Hanna Pituch, Piotr Obuch-Woszczatyński
106
nektaru roślin olejkowych. Przykładowe substancje to: tymol i karwakrol (miód
tymiankowy), tujon i kamfora (miód szałwiowy), β- i α-pinen (miód sosnowy) [2].
Antybiotyczne właściwości miodu wynikają również z aktywności enzymów
wytwarzanych przez gruczoły pszczół. Przykładem jest oksydaza glukozowa, katali-
zująca utlenianie β-D-glukozy do glukonolaktonu, który reagując z wodą przechodzi
w kwas glukonowy. Dodatkowym produktem tej reakcji jest H2O2 o właściwościach
bakteriobójczych. Rozcieńczenie miodu wodą przed spożyciem powoduje nawet 220-
krotny wzrost działania antybiotycznego. Nadtlenek wodoru jest wrażliwy na działanie
temperatury oraz promieni słonecznych, dlatego miody należy przechowywać
w ciemnym, chłodnym miejscu. Kolejnym enzymem wchodzącym w skład miodu jest
lizozym. Działanie bakteriobójcze tej substancji wynika ze zdolności do hydrolizy
wiązań między resztami N-acetyloglukozaminy (NAG) i kwasu N-acetylomura-
minowego (NAM), które są składnikami ściany komórkowej bakterii. W wyniku
wysokiego ciśnienia osmotycznego wewnątrz komórki, rozcięcie niewielkiego frag-
mentu polisacharydu prowadzi do rozerwania ściany komórkowej bakterii i pęknięcia
komórki [2]. Wielu naukowców zwraca uwagę na obecność pszczelej defensyny-1
w miodzie. Jest to białko o właściwościach bakteriobójczych wydzielane przez
gruczoły pszczół [3].
Miód manuka jest wytwarzany przez pszczoły miodne (Apis mellifera) z kwiatów
krzewu Manuka (Leptospermum scoparium) rosnącego w Nowej Zelandii. Produkt ten
jest środkiem dietetycznym, którego popularność rośnie na całym świecie. Według
doniesień naukowych miód manuka wykazuje szerokie właściwości przeciwdrob-
noustrojowe, przeciwzapalne oraz korzystnie wpływa na układ pokarmowy i skórę.
Właściwości antybakteryjne uwarunkowane są zawartością UMF (ang. unique manuka
factor), specyficznych związków biologicznie czynnych. Głównym składnikiem jest
metyloglioksal (MGO). Związek ten jest produktem nieenzymatycznej przemiany
dihydroksyacetonu (DHA), która następuje podczas przechowywania miodu. Ilość
MGO w miodzie manuka jest ok 100 razy większa w porównaniu do innych miodów,
co warunkuje wysokie właściwości antybakteryjne tego miodu [4].
Wilczyńska (2013) [5] określiła skład oraz potencjał antyoksydacyjny miodów
manuka dostępnych na polskim rynku, różniących się zawartością metyloglioksalu (od
100 do 550 mg/kg). Potencjał antyoksydacyjny zdefiniowano jako całkowitą zawartość
polifenoli oraz zdolność zmiatania kationorodników DPPH+ (Tab. 2)
Tabela 1. Aktywność antyoksydacyjna miodów manuka w zależności od stężenia metylglioksalu (MGO). TP
– ogólna zawartość polifenoli, GAE – ekwiwalent kwasu galusowego, AADPHH – zdolność zmiatania
kationorodników. Na podstawie [5].
Typ miodu TP (mgGAE/100g) AADPHH (%)
MGO 100 133,8 74,25
MGO 250 117,0 78,84
MGO 400 132,4 77,78
MGO 550 145,9 74,07

Działanie miodu manuka na wzrost oraz biofilm wybranych bakterii stanowiących problem terapeutyczny
107
Największą ogólną zawartością polifenoli odznaczał się miód MGO 550 (145,9
mgGAE/100 g). W porównaniu z polskimi miodami miody manuka charakteryzowały
się zbliżoną zawartością polifenoli do ciemnych miodów odmianowych takich jak
gryczany i wrzosowy (ok. 150 mgGAE/100 g). Zdecydowanie mniej polifenoli za-
wierały miody jasne, rzepakowe (ok. 30 mgGAE/100 g). Aktywność przeciwutle-
niająca badanych miodów manuka była zbliżona (ok. 75%) [5, 6]. Spośród polskich
miodów najwyższą aktywność przeciwutleniająca charakteryzowała miody wrzosowe,
gryczane i spadziowe (ok. 70%), natomiast najniższa wartość charakteryzowała miody
akacjowe i rzepakowe (ok. 30%) [5].
Miód manuka jest wykorzystywany m.in. do produkcji preparatów oraz opatrunków
przeznaczonych do leczenia przewlekłych ran i oparzeń, a także past do zębów.
3. Rola biofilmu bakteryjnego w patogenezie infekcji bakteryjnych
Biofilm bakteryjny jest złożoną strukturą biologiczną w postaci zagregowanych
komórek bakteryjnych przylegających do siebie i/lub do podłoża. Biofilmy charak-
teryzują się znacznie wyższą tolerancją na wysokie stężenia środków bakteriobójczych
oraz dezynfekcyjnych w porównaniu do bakterii w formie planktonowej. Oporność
bakterii wchodzących w skład biofilmu na leki jest 10-1000 razy większa w porów-
naniu do formy pływającej tych samych szczepów, natomiast niskie stężenia antybio-
tyków mogą pobudzać wzrost biofilmu jako odpowiedź na stres [7]. Biofilmy są w sta-
nie przemieszczać się w obrębie organizmu gospodarza i rozsiewać komórki pochodne,
najczęściej drogą układu krwionośnego. Bakterie tworzące biofilm wytworzyły mecha-
nizmy komunikacji oraz wymiany materiału genetycznego w systemie quorum sen-
sing. Komórki drobnoustrojów w biofilmie otaczają się macierzą pozakomórkową EPS
(ang. extracellular polymeric substance) składającej się z białek, polisacharydów oraz
DNA. Struktura macierzy komórkowej ma za zadanie stabilizować strukturę przes-
trzenną biofilmu, ochronę komórek przed wysychaniem oraz przed komórkami
żernymi organizmu. Biofilmy mogą składać się z jednego lub wielu gatunków bakterii.
Biofilmy wielogatunkowe charakteryzują się złożonymi relacjami międzygatun-
kowymi, takimi jak komensalizm, mutualizm lub antagonizm. Najczęstszym miejscem
występowania tego typu biofilmów jest powierzchnia zębów, pochwa oraz środowisko
naturalne [8]. Biofilmy w warunkach in vivo osiągają grubość od 4 do 200 µm. Zdecy-
dowanie większe agregaty powstają na powierzchni wszczepów od 5 do 1200 µm. [9].
Pojęcie biofilmu zostało wprowadzone do nauk medycznych przez prof. Williama
Costertona. Jako jedno z kryteriów, wskazujących na udział biofilmu w zakażeniu,
uznał on trudność wyleczenia infekcji przy pomocy antybiotyków lub chemio tera-
peutyków pomimo określenia lekowrażliwości izolatów na stosowany lek. Biofilmy są
powszechne w przyrodzie, przemyśle a także w środowisku szpitalnym. Do najczęst-
szych zakażeń przebiegających z udziałem biofilmów zalicza się zakażenia otolaryn-
gologiczne, oskrzelowo-płucne, ran, zapalenie wsierdzia, ozębnej i martwicze zapa-
lenie powięzi [10].

Michał Piotrowski, Hanna Pituch, Piotr Obuch-Woszczatyński
108
4. Działanie miodu manuka na wybrane gatunki bakterii
Potwierdzono skuteczność działania środków na bazie miodu manuka wobec takich
bakterii jak Staphylococcus aureus, Psudomonas aeruginosa, Eschericha coli i wiele
innych. Działanie miodu zapobiega wiązaniu się bakterii z ludzką fibronektyną oraz
powoduje dysocjację komórek bakteryjnych z biofilmu. Główną korzyścią stosowania
preparatów zawierających miód manuka jest niska cena oraz łagodny wpływ na ludzką
tkankę [11].
4.1. Clostridium difficile
Clostridium difficile jest bakterią Gram dodatnią, rosnącą w warunkach bezwzględ-
nie beztlenowych. Zakażenie tym drobnoustrojem prowadzi do wystąpienia objawów
określanych anglojęzycznym skrótem CDI (Clostridium difficile infection). Rozwój
zakażenia może przebiegać pod postacią lekkiej, samoograniczającej się biegunki,
infekcji o średnim przebiegu, ale może także dojść do ciężkiej biegunki i w konsek-
wencji rozwoju rzekomobłoniastego zapalenia okrężnicy. Na podstawie badań
własnych [12] potwierdzono, że szczepy C. difficile są wrażliwe na działanie miodu
manuka. Wartość MBC była taka sama jak wartość MIC dla szczepów należących do
wszystkich badanych szczepów i wyniosła 6,25% (v/v). Uzyskany wynik jest iden-
tyczny z wynikiem jedynego dostępnego badania na temat wpływu miodu manuka na
wzrost C. difficile, gdzie także uzyskano wartości MIC oraz MBC na poziomie 6,25%
(v/v) [13]. W najnowszych badaniach opisano zdolność C. difficile do wytwarzania
biofilmu in vitro. Istnieją nieliczne badania na temat wpływu miodu manuka na biofilm
C. difficile. Hammond i wsp. określili stężenie 20-50% (v/v) jako takie któro istotnie
zmniejsza ilość biofilmu [14]. Wyniki badań własnych były zbliżone, miód manuka
w badanym zakresie stężeń 10-50% (v/v) istotnie redukował ilość wytworzonego
biofilmu. Jednak w obu przypadkach niemożliwa była całkowita eradykacja biofilmu
przy użyciu miodu manuka. Dodatkowo zaobserwowano, że niskie stężenie miodu (1-
2% v/v) zwiększa wytwarzanie biofilmu, może to być spowodowane odpowiedzią na
stres, w tym wypadku niskie stężenie substancji przeciwdrobnoustrojowej [12].
Opis przypadku Na oddział gastroenterologii i hepatologii szpitala klinicznego (Tilburg, Holandia)
została przyjęta 71 letnia pacjentka. Objawy takie jak wodnista biegunka i niemożność utrzymywania stolca oraz badania laboratoryjne wskazały na infekcję Clostridium difficile. Wdrożono terapię wankomycyną, jednak biegunka nie ustąpiła po zastoso-wanym leczeniu. W konsekwencji zastosowano fidaksomycynę przez kolejne 10 dni, podczas kuracji objawy ustąpiły, natomiast następnego dnia po zakończonej terapii wodniste stolce powróciły. Nawrotowy charakter infekcji skłonił do rozważenia zabie-gu przeszczepu flory kałowej, jednak pacjentka nie wyraziła na to zgody. Jako alter-natywny sposób leczenia zaproponowano lewatywę do okrężnicy z 8% roztworu miodu manuka, po pięciu dniach od zabiegu objawy powróciły. Zabieg powtórzono z 15% roztworem miodu, który zakończył się powodzeniem. U pacjentki nie odno-

Działanie miodu manuka na wzrost oraz biofilm wybranych bakterii stanowiących problem terapeutyczny
109
towano objawów infekcji C. difficile przez kolejny rok od zabiegu, a także nie stwier-dzono obecności bakterii w kale.
Przytoczony opis przypadku ilustruje możliwości terapeutycznego wykorzystania miodu manuka w leczeniu biegunki o etiologii C. difficile opornej na konwencjonalne leczenie [15].
4.2. Staphylococcus aureus
Bakterie należące do gatunku Staphylococcus aureus są bardzo rozpowszechnione i nosicielstwo występuje u ok. 30% populacji. U większości osób skolonizowanych przez te bakterie nie rozwija się infekcja. Jeśli jednak bakterie są w stanie pokonać naturalne bariery, mogą wywołać infekcję, a jej objawy zależą od rodzaju wywołanego zakażenia. Większość zakażeń bakteriami to infekcje skórne, w tym czyraki, ropnie, zapalenie tkanki łącznej i liszajce. Największe zagrożenie stwarza coraz większy odsetek szczepów MRSA (ang. methicyllin-resistant Staphylococcus aureus). Fakt oporności gronkowca złocistego na najważniejszą grupę terapeutyczną jaką są β-laktamy dodatkowo potęguje często towarzysząca wielolekooporność. Zastosowanie w leczeniu zakażeń wywoływanych przez S. aureus znajduje miód manuka. Przeba-dano 58 szczepów gronkowca złosistego wrażliwych na metycylinę izolowanych z zakażonych ran. Średnia wartość MIC miodu manuka dla badanych szczepów wyniosła 2,88% (v/v) [16]. Miód manuka wykazuje skuteczne działanie bakteriobójcze w stosunku do szczepów MRSA, średnia wartość MIC wynosi 4% (v/v) [17]. Jednym z czynników wirulencji S. aureus jest wytwarzanie biofilmu. Jing i współpr. sprawdzili wrażliwość in vitro biofilmu tworzonego przez gronkowca złocistego, zaobserwowano zmniejszenie ilości biofilmu o 90% przy stężeniu miodu manuka 8% (v/v) [18].
4.3. Pseudomonas aeruginosa
Pałeczka ropy błękitnej Pseudomonas aeruginosa jest patogenem oportunistycznym mogącym wywołać niebezpieczne zakażenia takie jak zapalenie płuc, skóry i tkanek miękkich. Szczególne zagrożenie stanowią zakażenia szpitalne, często o skomplikowa-nym przebiegu. Jednym z pierwszych etapów patogenezy zapalenia płuc jest koloni-zacja, która stopniowo prowadzi do nasilenia procesu zapalnego oraz upośledzenia funkcji oddechowych. Niektóre szczepy P. aeruginosa wytwarzają pozakomórkowy polisacharyd – alginian, który umożliwia tworzenie biofilmu przez kolonie bakterii. To zjawisko w połączeniu z naturalną opronością P. aeruginosa na szeroką gamę anty-biotyków czyni ten patogen szczególnie trudnym do eradykacji. Dane literaturowe dostarczają informacji na temat wpływu miodu manuka na rozwój pałeczek ropy błękitnej. Cooper i współpr. [19] określili wartość MIC miodu manuka w stosunku do tych bakterii i wyniosła ona od 5.5 do 8.7% (v/v). Bakterie w biofilmie wykazują zdecydowanie większą oporność na miód manuka. Okhiria i współpr. [20] zaobserwo-wali istotne zmniejszenie ilości wytworzonego biofilmu przez P. aeruginosa przy zastosowanym stężenia 40% (v/v), natomiast stężenie 20% (v/v) nie przyniosło żadnych efektów. Doświadczenie przeprowadzone przez Camplin (2014) dało podobne wynik, MIC miodu manuka wobec izolowanych szczepów wyniosło od 5 do 7% (v/v), natomiast w stosunku do kolonii tworzących biofilm 40-45% (v/v) [21].

Michał Piotrowski, Hanna Pituch, Piotr Obuch-Woszczatyński
110
5. Podsumowanie
Zjawisko lekooporności bakterii coraz częściej utrudnia terapię, a nadmierna antybiotykoterapia dodatkowo nasila selekcję szczepów wielolekopornych. Innym problemem terapeutycznym jest wytwarzanie biofilmu przez bakterie. Bakterie wcho-dzące w skład biofilmu są bardziej odporne na szkodliwe warunki środowiska oraz charakteryzują się zwiększoną opornością na antybiotyki. W ostatnich latach bardzo intensywnie bada się miód manuka, pochodzący z Nowej Zelandii. Przedstawione dane literaturowe wskazują na szerokie możliwości zastosowania preparatów składających się z miodu manuka w leczeniu zakażeń bakteryjnych, również tych przebiegających przy udziale biofilmu. Warto rozważyć terapię miodem manuka ze względu na jego szeroką dostępność, stosunkowo niską cenę, a także brak toksycznego wpływu na tkanki człowieka.
Literatura
1. Lutomski J. Szczególna rola miodów w ziołolecznictwie. Postępy Fitoterapii, 4 (2001), s. 2-5. 2. Gałuszka H. Miód pszczeli, powstawanie, wartość odżywcza, zastosowanie. Sądecki
Bartnik; Nowy Sącz, (1998). 3. Kwakman P. How honey kills bacteria. The FASEB Journal, 24 (2010), s. 2576-82. 4. Schneider M., Coyle S., Warnock M. Anti-microbial activity and composition of manuka
and portobello honey. Phytotherapy Research, 27 (2013), s. 1162-68. 5. Wilczyńska A. Skład chemiczny i właściwości antyoksydacyjne miodu manuka. Problemy
Higieny Epidemiologicznej, 94 (2013), s. 873-75. 6. Stephens J.M. Phenolic compound and methyloglyoxal in some New Zealand kanuka and
manuka honeys. Food Chemistry, 120 (2010), s. 78-86. 7. Dapa T., Leuzzi R., Yen K. Multiple factors modulate biofilm formation by the anaerobic
pathogen Clostridium difficile. Journal of Bacteriology, 195 (2013), s. 545-55. 8. Flemming H.C. Biofilms: an emergent form of bacterial life. Nature Reviews
Microbiology, 49 (2016), s. 563-75. 9. Hoiby N. ESCMID guideline for the diagnosis and treatment biofilm infections. Clinical
Microbiology and Infections, 1 (2015), s. 1-25. 10. Costerton J.W. Microbial biofilms. Annual Review of Microbiology, 49 (1995), s. 711-45. 11. Maddocks S.E., Lopez M.S., Rowlands R.S. Manuka honey inhibits the development of
Streptococcus pyogenes biofilms and causes reduced expression of two fibronectin binding proteins. Microbiology, 158 (2012), s. 781-90.
12. Piotrowski M., Karpiński P., Pituch H., Obuch-Woszczatynski P. Antimicrobial effects of Manuka honey on in vitro biofilm formation by Clostridium difficile. European Journal of Clinical Microbiology and Infectious Diseases, (2017).
13. Hammond E., Donkor E. Antibacterial effect of manuka honey on Clostridium difficile. BMC Research Notes, 6 (2013), s. 188.
14. Hammond E., Donkor E. Biofilm formation of Clostridium difficile and susceptibility to manuka honey. BMC Complementary and Alternative Medicine, 14 (2014), s. 329.
15. Giles L.S, Laheij R. Successful treatment of persistent Clostridium difficile infection with manuka honey. International Journal of Antimicrobial Agents, 49 (2017), s. 522-23.
16. Cooper R.A., Molan P.C., Harding K.G. Antibacterial activity of honey against strains of Staphylococcus aureus from infected wounds. Journal of the Royal Society of Medicine, 92 (1999), s. 283-5.

Działanie miodu manuka na wzrost oraz biofilm wybranych bakterii stanowiących problem terapeutyczny
111
17. Allen K.L., Hutchinson G., Molan P.C. The potential for using honey to treat wounds infected with MRSA and VRE. First World Wound Healing Congress, 10-13 September (2000), Melbourne, Australia.
18. Lu J., Turnbull L., Burke C.M. Manuka-type honeys can eradicate biofilms produced by Staphylococcus aureus strains with different biofilm-forming abilities. Peer Journal, 2 (2014), s. 326.
19. Cooper R., Halas E., Molan P. The efficacy of honey in inhibiting strains of Pseudomonas aeruginosa from infected burns. Journal of Burn Care and Rehabilitation, 23 (2002), s. 366-70.
20. Okhiria O.A., Henriques A.F.M., Burton N.F. Honey modulates biofilms of Pseudomonas aeruginosa in a time and dose dependent manner. Journal of Api Product and Api Medical Science. 1 (2009) s. 6-10.
21. Camplin A.L., Maddocks S.E. Manuka honey treatment of biofilms of Pseudomonas aeruginosa results in the emergence of isolates with increased honey resistance. Annals of Clinical Microbiology and Antimicrobials, (2014), 13:19.
Działanie miodu manuka na wzrost oraz biofilm wybranych bakterii istotnych klinicznie
Streszczenie Miód manuka jest wytwarzany przez pszczoły z kwiatów krzewu Manuka (Leptospermum scoparium). Jest szczególnie ceniony za wysokie właściwości antybakteryjne pochodzące z zawartości naturalnego czyn-nika o działaniu antybiotycznym – metylgioksalu. W dzisiejszych czasach poszukiwanie nowych alterna-tywnych środków o działaniu przeciwbakteryjnym jest szczególnie ważne. Nadmierne stosowanie anty-biotyków prowadzi do powstawania szczepów bakterii wielolekoopornych. Udowodniono wpływ przeciw-drobnoustrojowy miodu manuka na wiele bakterii patogennych między innymi: Clostridium difficile, Staphylococcus aureus, Pseudomonas aeruginosa, również tych opornych na antybiotyki. Bakterie te są zdolne do wytwarzania biofilmu – złożonej struktury biologicznej w postaci zagregowanych komórek bak-teryjnych przylegających do siebie i/lub do podłoża. Biofilmy charakteryzują się znacznie wyższą to-lerancją na wysokie stężenia środków bakteriobójczych oraz dezynfekcyjnych w porównaniu do bakterii w formie planktonowej. Miód manuka wykazuje silne działanie antybakteryjne, zarówno wobec bakterii w formie planktonowej oraz tych wchodzących w skład biofilmu. Warto zatem rozważyć ewentualne zastosowanie roztworów miodu manuka o odpowiednim stężeniu jako alternatywnego środka antysep-tycznego na powierzchnie, rany oraz jak wskazują najnowsze doniesienia w formie wlewek dood-bytniczych. Słowa kluczowe: miód manuka, biofilm, Clostridium difficile, MRSA, Pseudomonas aeruginosa
Effect of manuka honey on planktonic growth and biofilm of selected clinically significant bacteria
Abstract Manuka honey, derived from the Manuka tree (Leptospermum scoparium), is known to exhibit antimicrobial properties that are associated with its significant content of methylglyoxal. Finding new antibacterial agents is very important today. Excessive usage of antibiotics leads to selection multi-drug resistant strains. Antimicrobial effects of manuka honey have been demonstrated for many pathogenic bacteria including Clostridium difficile, Staphylococcus aureus, Pseudomonas aeruginosa, also those resistant to antibiotics. These bacteria are able to producing biofilm – a complex biological structure in the form of aggregated bacterial cells adhering to each other and/or to the surface. Bacteria in biofilms are characterized by a much higher tolerance for high concentrations of bactericides. Manuka honey demonstrate antimicrobial activity relative to biofilms. Manuka honey is, therefore, a novel, promising agent and can be used as an alternative antiseptic agent to surfaces, wounds and for colon lavage. Keywords: manuka honey, biofilm, Clostridium difficile, MRSA, Pseudomonas aeruginosa

112
Jana Przekwas1, Joanna Kwiecińska-Piróg
2, Eugenia Gospodarek-Komkowska
3
Wpływ miodu Manuka na tworzenie biofilmu i wzrost
mgławicowy Proteus mirabilis
1. Wstęp
Rozprzestrzeniające się mechanizmy oporności bakterii powodują, że używane
dotychczas antybiotyki mają coraz węższe spektrum działania. Dodatkowym prob-
lemem jest tworzący się w ranach biofilm, utrudniający penetrację leków. W związku
z tym obserwuje się tendencję powrotu do metod alternatywnych leczenia ran
i oparzeń. Na rynku dostępne są już od kilku lat opatrunki i maści zawierające miód
Manuka (MEDIHONEY®) oraz inne miody. Prowadzone są także badania wery-
fikujące efektywność działania opatrunków wprowadzonych na rynek.
Proteus mirabilis (pałeczka odmieńca) jest trzecim pod względem częstości izolacji
gatunkiem bakterii po Staphylococcus aureus i Pseudomonas aeruginosa, koloni-
zującym rany przewlekłe, szczególnie owrzodzenia kończyn dolnych. Procentowo jego
udział jako czynnika etiologicznego zakażeń ran przewlekłych wynosi od 10 do 17%
[1-4].
Pałeczki P. mirabilis występują w glebie, nawozie i skażonej wodzie. Mogą stano-
wić mikroflorę układu pokarmowego człowieka i zwierząt. Najczęściej wywołują
zakażenia układu moczowego i wtórne zakażenia ran. Ich czynnikiem zjadliwości jest
m.in. ureaza, enzym rozkładający mocznik do amoniaku. Amoniak może prowadzić do
uszkadzania komórek nabłonkowych, alkalizacji moczu i, w rezultacie, tworzenia się
kamieni moczowych. Wewnątrz tych kamieni mogą znajdować się żywe komórki
pałeczek Proteus spp.
Bakterie te wytwarzają też proteazy, rozkładające ludzkie przeciwciała odporno-
ściowe. Innym czynnikiem jest zdolność tych drobnoustrojów do bardzo szybkiego
namnażania się. Charakterystyczny jest dla nich wzrost mgławicowy. Posianie punk-
towe na agarze powoduje zarośnięcie całej powierzchni płytki w ciągu 24 godzin.
Przyczyną tego zjawiska jest przemiana krótkich wegetatywnych form w wydłużone
formy [5, 6].
Celem niniejszej pracy jest ocena działania miodu Manuka na tworzenie biofilmu
przez P. mirabilis na płytkach polistyrenowych oraz na wzrost mgławicowy tych
pałeczek na podłożach agarowych. Hipoteza badawcza zakłada, że roztwór miodu
Manuka ogranicza zarówno formowanie biofilmu na płytkach jak i wzrost mgła-
wicowy P. mirabilis na podłożach stałych.
1 [email protected]; Studenckie Koło Naukowe przy Katedrze i Zakładzie Mikrobiologii; Wydział
Farmaceutyczny; Collegium Medicum im. Ludwika Rydygiera w Bydgoszczy, Uniwersytet Mikołaja
Kopernika w Toruniu 2 [email protected]; Katedra i Zakład Mikrobiologii; Wydział Farmaceutyczny; Collegium Medicum
im. Ludwika Rydygiera w Bydgoszczy, Uniwersytet Mikołaja Kopernika w Toruniu 3 [email protected]; Katedra i Zakład Mikrobiologii; Wydział Farmaceutyczny; Collegium Medicum
im. Ludwika Rydygiera w Bydgoszczy, Uniwersytet Mikołaja Kopernika w Toruniu

Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis
113
1.1. Dimorfizm Proteus mirabilis
Ze względu na rodzaj środowiska, w jakim się znajdują, bakterie P. mirabilis
przyjmują jedną z form. W środowisku płynnym są krótkimi pałeczkami z kilkoma
rzęskami. Na powierzchniach stałych zachodzi różnicowanie – pałeczki wydłużają się,
wielokrotnie wzrasta liczba rzęsek oraz nukleoidów wewnątrz komórki nieprze-
dzielonych w żaden sposób. Populacja tych komórek w sposób skoordynowany prze-
mieszcza się. Wykorzystuje w tym celu swoisty sposób komunikacji między poje dyn-
czymi komórkami tj. quorum sensing. Zachodzi on z udziałem specyficznych czynni-
ków chemicznych, między innymi putrescyny, glutaminy czy cyklicznych peptydów.
Po rozejściu się bakterii następuje konsolidacja, czyli rozpad pałeczek długich do form
krótkich. W ten sposób zachodzi wzrost rozpełzły P. mirabilis o charakterze cyk-
licznym [5].
W procesie różnicowania form krótkich do długich można wyróżnić kluczowe
etapy. Pierwszy z nich zachodzi po dotarciu komórki do powierzchni stałej. Kontakt
z powierzchnią powoduje hamowanie ruchu rzęsek, co inicjuje inne przemiany meta-
boliczne, prowadzące do różnicowania. Następuje zwiększona aktywacja czynnika
FIhD2C2, który reguluje syntezę białek budujących fimbrie niezbędne do migracji.
Kolejnymi sygnałami są pozakomórkowa akumulacja putrescyny, będącej jednym
z sygnałów quorum sensing oraz interakcja antygenu O, występującego w ścianie
komórek, z powierzchnią adhezji [7].
Krytycznym czynnikiem środowiskowym wpływającym na wzrost rozpełzły P.
mirabilis jest obecność aminokwasu glutaminy w podłożu. Jej brak powoduje zaha-
mowanie wzrostu rozpełzłego. Inne czynniki wpływające na wzrost mgławicowy to
między innymi: dostęp tlenu, gęstość komórek na podłożu, stężenie elektrolitów w śro-
dowisku [8].
Formy długie nie są potrzebne do utworzenia biofilmu. Były jednak obserwowane
jako komórki opuszczające biofilm, zwłaszcza w niesprzyjającym, ubogim w substan-
cje odżywcze środowisku [9].
1.2. Tworzenie się biofilmu bakteryjnego
Biofilm tworzy się zazwyczaj na styku dwóch faz (ciecz-gaz, ciało stałe-ciecz, ciało
stałe-gaz itd.) na materiałach zarówno pochodzenia biologicznego, jak i tworzywach
sztucznych. Jest to wielokomórkowy twór, w którego skład mogą wchodzić drobno-
ustroje jednego lub więcej gatunków. Z zewnątrz otoczony jest glikokaliksem, a jego
wewnętrzną strukturę stabilizują substancje polimeryczne wydzielane przez komórki –
EPS (extracellular polymeric substances). Jego grubość oraz kształt zależą od panu-
jących warunków [10].
Powstawanie biofilmu można podzielić na kilka etapów, w których można
wyszczególnić fazy. Pierwszym etapem jest adhezja komórek. Adhezja odwracalna
komórek bakterii do powierzchni jest spowodowana przede wszystkim oddziały-
waniami fizycznymi, takimi jak siła grawitacji, siły hydrodynamiczne, ruchy Browna,
siły van der Waalsa. Odległość między komórkami a powierzchnią adhezyjną pozos-
taje większa niż 1,5 nm. Na tym etapie łatwo jest usunąć tworzący się biofilm.

Jana Przekwas, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
114
Adhezja nieodwracalna jest zależna od obecności na powierzchni bakterii fimbrii
i rzęsek. Fimbrie pozwalają na pokonanie sił odpychania między ujemnie naładowa-
nymi ścianą komórkową bakterii i powierzchnią adhezji. Rzęski umożliwiają prze-
mieszczanie się, co pozwala, po zbliżeniu się do powierzchni adhezji, na poszukiwanie
utworzonych już mikrokolonii. Formy swarmer cells P. mirabilis posiadają zarówno
rzęski, jak i fimbrie.
Po zbliżeniu się komórek do powierzchni na odległość mniejszą niż 1,5 nm rozpo-
czyna się adhezja nieodwracalna, polegająca na utworzeniu wiązań między powierz-
chnią a adhezynami bakterii, fimbriami czy polimerami polisacharydowymi. Rozpo-
czyna się produkcja glikokaliksu otaczającego formujący się biofilm [11].
Kolejnym etapem jest tworzenie mikrokolonii. Polega on na namnażaniu drobno-
ustrojów a także ich różnicowaniu i przejmowaniu różnych funkcji. Proces ten odbywa
się poprzez aktywację bądź hamowanie ekspresji odpowiednich genów. Komórki
przebywające w niższych warstwach biofilmu tracą rzęski. Ich metabolizm zmienia się
na beztlenowy ze względu na utrudniony dostęp do tlenu. Zahamowana jest synteza
niektórych enzymów czy toksyn. Bakterie namnażają się wolno lub przechodzą w sta-
nie anabiozy. Komórki warstw powierzchniowych biofilmu pozostają aktywne.
Umożliwia to obecność sieci kanałów w biofilmie, pozwalających na swobodny
przepływ substancji odżywczych [12].
Możliwy jest przepływ informacji między komórkami – quorum sensing. Specy-
ficzne cząsteczki sygnałowe przekazują informację o poziomie zagęszczenia
w biofilmie. Rodzaj oraz liczba cząsteczek sygnałowych, dla których komórki
posiadają receptory, informuje o liczbie bakterii tego samego gatunku znajdujących się
w błonie biologicznej. Po przekroczeniu pewnej liczby osobników i stężenia
cząsteczek sygna-łowych w środowisku, wszystkie komórki w podobnym czasie
zmieniają swój meta-bolizm. Przepływ informacji zachodzi również między
osobnikami różnych gatunków. Dzięki temu możliwa jest koordynacja procesów
zachodzących w obrębie biofilmu [13, 14].
Po osiągnięciu dojrzałości i odpowiedniej grubości biofilmu, następuje jego powol-
na degradacja. W rezultacie uwalniane są z niego komórki. Wynika to z niesprzyja-
jących warunków środowiska, wyczerpania składników odżywczych lub trudności
z ich przepływem w głąb biofilmu. Pojedyncze formy zdolne do ruchu poszukują no-
wych miejsc do zasiedlenia lub dołączają do formujących się mikrokolonii. Biofilm
rozprzestrzenia się [15].
Większość szczepów P. mirabilis ma dużą zdolność tworzenia biofilmu. Dużą rolę
w tym procesie odgrywają fimbrie oraz rzęski, którymi charakteryzują się formy
swarmer cells odpowiedzialne za wzrost mgławicowy [10].
1.3. Wpływ obecności biofilmu na proces gojenia
Prawidłowe gojenie rany przeprowadzone pod opieką lekarza opiera się na
koncepcie T. I. M. E. Rozwinięcie tego anglojęzycznego skrótu wskazuje na podsta-
wowe problemy napotykane podczas gojenia ran. Pierwszym z nich jest konieczność
oczyszczania rany z martwej tkanki, obcych elementów, resztek opatrunku czy

Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis
115
biofilmu (T, Tissue; tkanka). Drugim problemem jest obecność stanu zapalnego (I,
Inflammation), który jest naturalną odpowiedzią organizmu na uszkodzenie ciągłości
skóry. Nadmierne, przedłużające się zapalenie może świadczyć o zakażeniu rany czy
obecności biofilmu. Przy prawidłowym gojeniu się rany należy starać się o zachowanie
równowagi nawodnienia w środowisku rany (M, moisture balance; nawilżenie).
Znaczenie ma również brzeg rany (E, edge of wound) na pograniczu z tkanką zdrową.
Proces zapalny rozwijający się w ranie jest niezbędny w prawidłowym gojeniu. Jed-
nak jeżeli ten etap się przedłuża, zwłaszcza przy jednoczesnym formowaniu biofilmu,
może być niebezpieczny dla pacjenta. Biofilm powoduje zmiany w łożysku rany
i zahamowanie gojenia. Stymuluje proces zapalny [21], zaburza procesy immuno-
logiczne (osłabiając je), opóźnia ziarninowanie i naskórkowanie rany. Ze względu na
swoją strukturę biofilm jest oporny na działanie dezynfektantów. Penetracja antybio-
tyków do komórek biofilmu również jest utrudniona. Udowodniono, że minimalne
stężenie antybiotyków hamujące (minimal inhibitory concentration, MIC) wzrost
bakterii w biofilmie może być wielokrotnie wyższe niż MIC komórek planktonicznych
tego samego gatunku [14, 22]. Biofilm tworzy się w większości ran przewlekłych.
Badania z 2008 roku podają, że był on obecny w 60% badanych ran przewlekłych [23].
Zjadliwość i zdolność do wywoływania ostrych postaci zakażeń jest zależna od
bakterii tworzących biofilm.
1.4. Produkty pszczele jako środki terapeutyczne
Apiterapia to forma medycyny tradycyjnej, w której stosuje się miód, pyłek, wosk
pszczeli, mleczko pszczele, propolis, jad pszczeli i inne produkty pochodzenia pszcze-
lego. Aktualnie bardzo popularnym produktem pszczelim jest pochodzący z Nowej
Zelandii miód Manuka.
Miody Manuka wytwarzane są z pyłku roślin Leptospermum scoparium. W porów-
naniu do miodów pochodzenia polskiego różnią się zawartością sacharozy. W polskich
miodach średnia zawartość sacharozy stanowi średnio 2,3%, natomiast w miodach
nowozelandzkich 15,4%. W miodach Manuka notuje się wysoką zawartość 5-hydro-
ksymetylofurfuralu. Bardzo charakterystyczna dla tych miodów jest wysoka zawartość
polifenoli oraz związana z tym zdolność usuwania wolnych rodników, porównywalna
jedynie z miodami ciemnymi (gryczanymi, wrzosowymi, spadziowymi) [16]. Naj-
ważniejszą właściwością miodów Manuka jest wysoka zawartość metylglioksalu
(MGO), nawet do 761 mg/kg, co stanowi wartość wielokrotnie wyższą niż zawartość
metylglioksalu w innych miodach (do 24 mg/kg). Związek ten wykazuje właściwości
hamujące wzrost bakterii w stężeniu 1,1 mM [17]. Miody Manuka zawierające tak
wysoką zawartość metylglioksalu wykazywały działanie bakteriobójcze w stężeniu od
15 do 30% [17].
MGO powstaje w nieenzymatycznym procesie z dihydroksyacetonu, który wystę-
puje w nektarze zebranym z kwiatów Leptospermum scopiarum. Udowodniono, że
świeżo zebrany miód Manuka zawiera niskie stężenia MGO, ale wysokie stężenia
dihydroksyacetonu, natomiast miód przechowywany w 37°C zawierał niższe stężenie
dihydroksyacetonu, ale odpowiednio wyższe stężenie MGO [18].

Jana Przekwas, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
116
Inne składniki miodów pochodzących z roślin Leptospermum, mogące mieć
właściwości inhitibujące wzrost bakterii lub modulować tą właściwość miodu to
nadtlenek wodoru powstający w wyniku działania oksydazy glukozowej pszczół,
obecność fenoli, węglowodanów złożonych i peptydów, a także wysoka osmolarność
i nieskie wartości pH 3,5- 4,5 [19]. Obecność charakterystycznego dla Leptospermum
glikozydu syringinian metylu 4-O-β-D-gentobioza nazwanego leptozyną w miodach
Manuka koreluje z ilością metylglioksalu będącego głównym komponentem
odpowiedzialnym za jego bakteriobójcze właściwości [20].
1.5. Wpływ miodu na proces gojenia
Skrócenie czasu gojenia rany pod wpływem działania produktów pszczelich można
tłumaczyć ograniczeniem odpowiedzi zapalnej. Z jednej strony miód Manuka po-
wstrzymuje rozwój przewlekłego zapalenia przez hamowanie rozprzestrzeniania się
komórek zapalnych w ranie. Jednocześnie jednak stymuluje produkcję cytokin
prozapalnych: TNF-α, IL-1β i IL-6 aktywując w ten sposób układ immunologiczny
i wzmagając stan zapalny [24]. Pozwala to na kontynuację prawidłowego procesu
gojenia rany, pobudza proliferację fibroblastów oraz komórek nabłonka. Dodatkowo
pH miodu o wartości 3,5 do 4,5 nie tylko hamuje wzrost bakterii, ale i stymuluje
makrofagi do aktywności bakteriobójczej.
Antybakteryjne działanie miodu Manuka jest zależne od zawartości MGO [17].
Zawarte w nim fenole mają aktywność antyoksydacyjną oraz antybakteryjną [16].
Neutralizacja MGO w miodzie pochodzącym z kwiatów Leptospermum scopiarum nie
prowadzi do obniżenia działania antybakteryjnego miodu wobec P. aeruginosa, ale
powoduje obniżenie aktywności miodu wobec S. aureus oraz B. subtilis [25]. Działanie
antybakteryjne miodu Manuka nie zależy więc jedynie od MGO i jest różne wobec
różnych gatunków bakterii. Wykazano również, że w owrzodzeniach będących
powikłaniem cukrzycy jego wysokie stężenie źle wpływa na proces gojenia [26].
2. Materiały i metody
W badaniach wykorzystano 18 szczepów P. mirabilis, w tym 17 pochodzących
z wymazów z ran przewlekłych i odleżyn pacjentów Szpitala Uniwersyteckiego nr 1
im. dr A. Jurasza w Bydgoszczy. Jeden szczep, P. mirabilis 29906, pochodził z amery-
kańskiej kolekcji szczepów wzorcowych (American Tissue Culture Collection,
ATCC). Wszystkie badane szczepy wykazywały zdolność do wzrostu mgławicowego
oraz tworzenia biofilmu. Do czasu badań szczepy przechowywano w bulionie
mózgowo-sercowym (Becton Dickinson) z 20% zawartością glicerolu (Avator)
w temperaturze -70°C.
W pracy badano działanie bakteriobójcze miodu Manuka (Manuka Health)
o zawartości MGO na poziomie co najmniej 400 mg/kg.

Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis
117
2.1. Badanie działania miodu Manuka na tworzący się biofilm
Proteus mirabilis
Badane szczepy pojedynczo wysiewano z zamrożenia na agar tryptozowo-sojowy (Tryptose-Soy Agar, TSA; Becton-Dickinson). Po 24 godzinach inkubacji w tempera-
turze 37C pobierano bakterie ze strefy wzrostu rozpełzłego i zawieszano w bulionie Mueller-Hinton (Mueller-Hinton Bouillion, MHB; Becton Dickinson) w celu otrzyma-nia zawiesiny o gęstości 0,5 w skali McFarlanda.
Biofilm tworzono w polistyrenowych 96-dołkowych płytkach (Profilab). Do każdego dołka dodawano 100 μl zawiesiny bakterii i odpowiednio: 100 μl jałowego MHB do kontroli dodatniej, oraz po 100 μl miodu Manuka rozcieńczonego zbuforo-wanym roztworem soli fizjologicznej (PBS, Avantor), uzyskując stężenie końcowe miodu, odpowiednio: 5,0%, 12,5% i 25,0%. Badanie wykonano w trzech powtórze-niach. Kontrolę stanowiło 200 μl podłoża MHB.
Płytki polistyrenowe zamykano szczelnie parafilmem. Umieszczano je w wilgotnej komorze i inkubowano w 37°C. Po 24 godzinach usuwano zawartość dołków i płukano jałowym roztworem PBS.
Komórki biofilmu uwalniano z wykorzystaniem ultradźwięków (Ultrasonic DU-4, Nickel-Electro) o częstotliwości 40 kHz. Uzyskiwano zawiesinę bakterii uwolnio-nych z biofilmu. Zawiesiny rozcieńczano w szeregu a następnie posiewano 100 μl odpowiednich rozcieńczeń na stałe podłoża TSA. Hodowle inkubowano przez 24 godziny w 37°C. Wyniki w trzykrotnych powtórzeniach odczytywano zliczając ilość kolonii na podłożach i wyrażano w jednostkach tworzących kolonie (j.t.k.)/dołek.
2.2. Badanie działania miodu Manuka na wzrost mgławicowy Proteus mirabilis
W celu oceny wzrostu mgławicowego przygotowywano jałowy agar tryptozowo-sojowy (Tryptose-Soy Agar, TSA), do którego dodawano miód Manuka rozcieńczony jałowym roztworem PBS, otrzymując końcowe stężenia miodu odpowiednio: 25,0%, 12,5% i 5,0%. Agar z dodatkiem miodu w warunkach sterylnych wylewano na jałowe szalki Petriego i pozostawiono do stężenia.
Po dwóch godzinach badane szczepy posiewano punktowo na przygotowane podłoże z miodem. Kontrolę wzrostu rozpełzłego stanowiło podłoże TSA bez dodatku miodu.
Hodowle inkubowano przez 24 godziny w temperaturze 37°C a następnie przez kolejne 96 godzin w temperaturze pokojowej. Wyniki odczytywano po 24, 48 i 120 godzinach inkubacji mierząc strefę wzrostu mgławicowego. Po 24 godzinach kolonie wyrosłe na podłożach z miodem o stężeniu 5%, 12,5% i 25% przesiano na jałowe podłoża TSA bez dodatku miodu, inkubowano przez 24 godziny w 37°C a następnie przez 24 h w temperaturze pokojowej. Wyniki w trzykrotnych powtórzeniach odczyty-wano po 24 i 48 godzinach po wykonaniu posiewu.
2.3. Ocena lekowrażliwości
Wrażliwość badanych szczepów na leki przeciwdrobnoustrojowe była oceniana za pomocą kart VITEK AST-N332 (BioMėrieux). Wyniki były interpretowane zgodnie z aktualnymi rekomendacjami EUCAST.

Jana Przekwas, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
118
2.4. Analiza statystyczna
Analizę statystyczną wykonano w programie Statistica 13. Wyniki badań wpływu działania miodu Manuka na wzrost mgławicowy i formowanie biofilmu przez P. mirabilis opracowano statystycznie wykorzystując test Bonferroniego, natomiast wyniki badania związku między antybiotykoopornością badanych szczepów klinicz-nych a ich wrażliwością na miód Manuka opracowano statystycznie wykonując test ANOVA rang Kruskala-Wallisa.
3. Wyniki i wnioski
3.1. Hamowanie tworzenia biofilmu przez miód Manuka
W badaniu hamowania tworzenia biofilmu przez miód Manuka mediana jednostek tworzących kolonie/dołek (j.t.k./dołek) dla kontroli dodatniej (0%; bez dodatku miodu)
wynosiła , natomiast dla biofilmów poddanych działaniu miodu w stężeniach 5,0%, 12,5% i 25,0% odpowiednio , 2 i . Testem Bonferroniego (α=0,05) wykazano różnice statystyczne pomiędzy kontrolą dodatnią a wszystkimi badanymi stężeniami miodu Manuka. Istotne statystycznie różnice zaobserwowano także między stężeniami 5,0% miodu i 25,0% miodu [Wykres 1].
Wykres 1. Wpływ stężenia miodu Manuka na tworzenie biofilmu przez Proteus mirabilis wyrażony
w j.t.k./dołek. Symbolem „*” oznaczono wartość testu ANOVA [opracowanie własne].
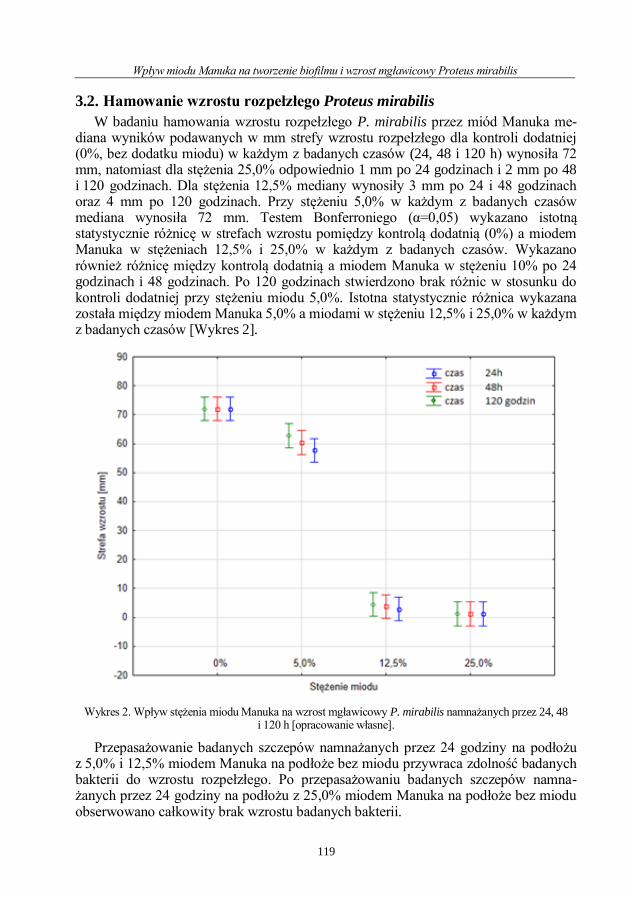
Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis
119
3.2. Hamowanie wzrostu rozpełzłego Proteus mirabilis
W badaniu hamowania wzrostu rozpełzłego P. mirabilis przez miód Manuka me-diana wyników podawanych w mm strefy wzrostu rozpełzłego dla kontroli dodatniej (0%, bez dodatku miodu) w każdym z badanych czasów (24, 48 i 120 h) wynosiła 72 mm, natomiast dla stężenia 25,0% odpowiednio 1 mm po 24 godzinach i 2 mm po 48 i 120 godzinach. Dla stężenia 12,5% mediany wynosiły 3 mm po 24 i 48 godzinach oraz 4 mm po 120 godzinach. Przy stężeniu 5,0% w każdym z badanych czasów mediana wynosiła 72 mm. Testem Bonferroniego (α=0,05) wykazano istotną statystycznie różnicę w strefach wzrostu pomiędzy kontrolą dodatnią (0%) a miodem Manuka w stężeniach 12,5% i 25,0% w każdym z badanych czasów. Wykazano również różnicę między kontrolą dodatnią a miodem Manuka w stężeniu 10% po 24 godzinach i 48 godzinach. Po 120 godzinach stwierdzono brak różnic w stosunku do kontroli dodatniej przy stężeniu miodu 5,0%. Istotna statystycznie różnica wykazana została między miodem Manuka 5,0% a miodami w stężeniu 12,5% i 25,0% w każdym z badanych czasów [Wykres 2].
Wykres 2. Wpływ stężenia miodu Manuka na wzrost mgławicowy P. mirabilis namnażanych przez 24, 48
i 120 h [opracowanie własne].
Przepasażowanie badanych szczepów namnażanych przez 24 godziny na podłożu z 5,0% i 12,5% miodem Manuka na podłoże bez miodu przywraca zdolność badanych bakterii do wzrostu rozpełzłego. Po przepasażowaniu badanych szczepów namna-żanych przez 24 godziny na podłożu z 25,0% miodem Manuka na podłoże bez miodu obserwowano całkowity brak wzrostu badanych bakterii.
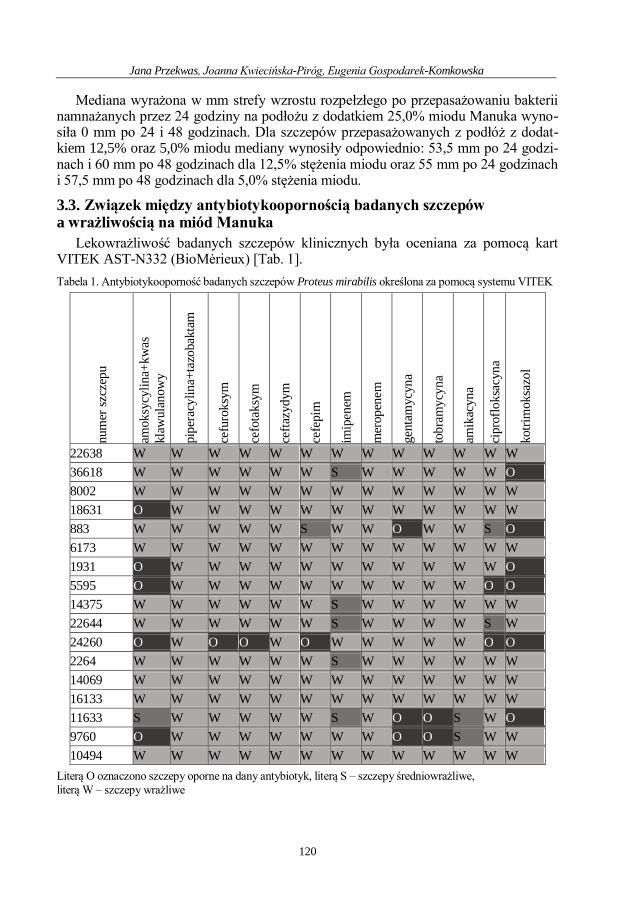
Jana Przekwas, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
120
Mediana wyrażona w mm strefy wzrostu rozpełzłego po przepasażowaniu bakterii namnażanych przez 24 godziny na podłożu z dodatkiem 25,0% miodu Manuka wyno-siła 0 mm po 24 i 48 godzinach. Dla szczepów przepasażowanych z podłóż z dodat-kiem 12,5% oraz 5,0% miodu mediany wynosiły odpowiednio: 53,5 mm po 24 godzi-nach i 60 mm po 48 godzinach dla 12,5% stężenia miodu oraz 55 mm po 24 godzinach i 57,5 mm po 48 godzinach dla 5,0% stężenia miodu.
3.3. Związek między antybiotykoopornością badanych szczepów a wrażliwością na miód Manuka
Lekowrażliwość badanych szczepów klinicznych była oceniana za pomocą kart VITEK AST-N332 (BioMėrieux) [Tab. 1].
Tabela 1. Antybiotykooporność badanych szczepów Proteus mirabilis określona za pomocą systemu VITEK
num
er s
zcze
pu
amoksy
cyli
na+
kw
as
kla
wula
now
y
pip
erac
yli
na+
tazo
bak
tam
cefu
roksy
m
cefo
taksy
m
ceft
azydym
cefe
pim
imip
enem
mer
open
em
gen
tam
ycy
na
tobra
mycy
na
amik
acyna
cipro
floksa
cyna
kotr
imoksa
zol
22638 W W W W W W W W W W W W W
36618 W W W W W W S W W W W W O
8002 W W W W W W W W W W W W W
18631 O W W W W W W W W W W W W
883 W W W W W S W W O W W S O
6173 W W W W W W W W W W W W W
1931 O W W W W W W W W W W W O
5595 O W W W W W W W W W W O O
14375 W W W W W W S W W W W W W
22644 W W W W W W S W W W W S W
24260 O W O O W O W W W W W O O
2264 W W W W W W S W W W W W W
14069 W W W W W W W W W W W W W
16133 W W W W W W W W W W W W W
11633 S W W W W W S W O O S W O
9760 O W W W W W W W O O S W W
10494 W W W W W W W W W W W W W
Literą O oznaczono szczepy oporne na dany antybiotyk, literą S – szczepy średniowrażliwe,
literą W – szczepy wrażliwe

Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis
121
Wszystkie badane szczepy były wrażliwe na działanie piperacyliny z tazobakta-mem, ceftazydymu i meropenemu. Najwięcej spośród 17 badanych szczepów okazało się opornych wobec działania amoksycyliny z kwasem klawulanowym (29% spośród badanych szczepów okazało się opornych na działanie tego leku) i kotrimoksazolu (35% szczepów badanych było oporne).
Wyniki badań po wykonaniu testu statystycznego ANOVA rang Kruskala-Wallisa nie wykazały istotnej statystycznie zależności między wrażliwością szczepów na miód a wrażliwością na antybiotyki.
4. Dyskusja
Badania przeprowadzone w niniejszej pracy wykazały, że roztwór miodu Manuka w stężeniach 12,5 i 25,0% efektywnie hamuje tworzenie biofilmu przez bakterie P. mirabilis, a także inhibuje ich wzrost rozpełzły. Mediana dla kontroli dodatniej 0% (bez wpływu miodu) wynosiła
j.t.k./dołek, podczas gdy mediana w dołkach z dodatkiem miodu w stężeniu 25,0% wynosiła j.t.k./dołek, a w stężeniu 12,5% – 2 j.t.k./dołek. Wyniki te są zbieżne z rezultatami badań uzyskanymi przez innych autorów, na przykład badania wpływu miodu Manuka na tworzący się oraz uformowany biofilm P. mirabilis i Enterobacter cloacae wykazały, że miód o zawartości MGO 250 g/kg w stężeniu 10% hamuje rozwój biofilmu przez oba badane szczepy [27]. Wykazano również, że miód Manuka w stężeniach 10% i wyższych hamuje rozwój biofilmu P. mirabilis oraz E. cloacae. Na biofilm 24-godzinny szczepów obu badanych gatunków działano 50% stężeniem miodu przez kolejne 24 godziny. Metodą z fioletem krystalicznym wykazano istotny statystycznie spadek biomasy biofilmu w porównaniu z kontrolą dodatnią. Oceniano również liczbę żywych komórek przez posiew na podłoża stałe rozcieńczeń dziesiętnych i zliczanie wyrosłych kolonii po 24-godzinnej inkubacji w 37°C. Wykazano spadek liczby żywych komórek w przypadku obu szczepów, jednak zdecydowanie większy dla P. mirabilis niż dla E. cloacae. Stężenie 50% miodu Manuka na tworzący się biofilm dawało najlepsze efekty w hamowaniu jego rozwoju. Działano również MGO w stężeniach 0,005% i 0,01% na tworzący się biofilm i 0,1% na 24-godzinny biofilm obu gatunków bakterii. Udowodniono, że MGO nie przeciwdziała tworzeniu się biofilmu, ale w użytym stężeniu redukuje liczbę komórek żywych w biofilmie 24-godzinnym P. mirabilis. Oznacza to, że MGO może dyfundować do macierzy pozakomórkowej chroniącej biofilm i w ten sposób utrudniać jego rozwój [27].
W badaniach wpływu miodu Manuka o UMF (Unique Manuka Factor) 15+ (~550 mg/kg MGO) na tworzący się biofilm i biofilm 24-godzinny [28] największe zahamo-wanie procesu jego tworzenia osiągnięto przy najwyższym stosowanym stężeniu końcowym miodu, tj. 16,7%. Nastąpiła redukcja biofilmu o 77% w ciągu 3 dni w po-równaniu z kontrolą dodatnią. Oceny biomasy biofilmu dokonano pomiarem foto-metrycznym z wykorzystaniem fioletu krystalicznego [28].
Podobne badania na szczepie P. mirabilis przeprowadzono dodając do formującego się biofilmu miód Manuka w stężeniu o połowę mniejszym niż oznaczone MIC dla tego szczepu, tj. 20%. Uzyskany biofilm barwiono fioletem krystalicznym. Wykazano 96-procentową redukcję biomasy biofilmu [29].

Jana Przekwas, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
122
Działanie miodu Manuka na formowanie biofilmu sprawdzano również w odnie-sieniu do szczepów Streptococcus pyogenes. Badano wpływ miodu w stężeniach 0%, 10%, 20% oraz 40%. Wykazano 75-procentową redukcję biofilmu przy stężeniu 10% miodu oraz odpowiednio 90- i 96-procentową redukcję przy stężeniu miodu 20 i 40% po barwieniu fioletem krystalicznym. Obraz biofilmu uformowanego przy 20% w mikroskopie świetlnym ujawnił brak formowania mikrokolonii, zwiększone rozmiary niektórych komórek i obecność długich łańcuchów komórek [30].
Badania wpływu miodu Manuka na wzrost rozpełzły P. aeruginosa przez posiew punktowy szczepów na agarze z dodatkiem miodu w stężeniach od 1,5 do 12% wykazały całkowite zahamowanie procesu przy stężeniu miodu 12% oraz zahamo-wany wzrost przy stężeniach od 1,5 do 6% w porównaniu z próbą bez dodatku miodu. Wykazano, że miód Manuka powoduje zahamowanie ekspresji genów kodujących białka budujące rzęski. Uniemożliwia przez to migrację komórek [31]. Stężenie hamujące wzrost rozpełzły wykazane dla P. aeruginosa jest zbliżone do stężenia, dla którego w niniejszych badaniach wykazano przeciwmgławicowe działanie miodu Manuka wobec P. mirabilis.
Czynniki obecne w miodzie prawdopodobnie odpowiedzialne za zahamowanie wzrostu mgławicowego P. mirabilis to wysoka osmolarność, zawartość MGO, niskie pH, aktywność antyoksydacyjna związków fenolowych. Aby stwierdzić, które czyn-niki mają krytyczny wpływ na obserwowane skutki badań, należałoby przeprowadzić badania wpływu izolowanych czynników obecnych w miodzie na wzrost mgławicowy P. mirabilis.
Wciąż prowadzone są badania in vitro i in vivo nad wpływem miodu Manuka na wzrost wielu gatunków bakterii wywołujących różne zakażenia.
Badania prowadzone przez Rabie i współpracowników mające na celu określić mechanizm działania antybakteryjnego MGO wykazały, że hamuje wzrost zarówno bakterii Gram dodatnich (B. subtilis, S. aureus), jak i Gram ujemnych (P. aeruginosa, E. coli). Badano wpływ MGO w stężeniu 0,5, 1,0 i 2,0 mM na struktury komórkowe B. subtilis i E. coli. Wykazano, że przy stężeniach 1,0 i 2,0 mM komórki posiadały mniej fimbrii oraz mniej lub całkowity brak rzęsek. Przy 2,0 mM MGO badane szczepy wykazywały brak tych struktur komórkowych, okrągły kształt komórek wynikający z ich kurczenia się oraz utratę ciągłości błony komórkowej. Tak dalece idące zmiany w komórkach powstałe pod wpływem MGO utrudniają adhezję oraz poruszanie się bakterii. Może to powodować hamowanie wzrostu mgławicowego tych gatunków [32].
Miód Manuka wykazuje właściwości bakteriobójcze nie tylko wobec Proteus spp., ale także wobec wielu innych pałeczek Gram-ujemnych, Enterococcus spp., w tym wykazujących mechanizm oporności VRE, Staphylococcus spp. opornych na metycylinę, E. coli i Enterobacter spp. wytwarzających beta-laktamazy o rozsze-rzonym spektrum substratowym (extender spectrum beta-lactamases, ESBL) [33]. W innych badaniach wykazano, że miód Manuka obniża ekspresję genów wirulencji w biofilmie tworzonym in vitro przez metycylinooporne szczepy Staphylococcus aureus, co ma wpływ na patogenność tych bakterii [34]. Wskazuje to na szerokie spektrum działania miodu Manuka oraz niewybiórcze działanie w przeciwieństwie do antybiotyków.

Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis
123
5. Wnioski
Roztwory miodu Manuka w stężeniach od 5,0 do 25,0% ograniczają tworzenie
biofilmu P. mirabilis na płytkach polistyrenowych oraz hamują wzrost rozpełzły
bakterii na podłożu agarowym. Skuteczność antybakteryjna miodu Manuka rośnie
wraz ze wzrostem jego koncentracji w roztworze.
Literatura
1. Bessa L. J., Fazii P., Giulio M. D. Bacterial isolates from infected wounds and their antibiotic susceptibility pattern: some remarks about wound infection, International Wound Journal, 12(1) (2008), s.47-52 doi:10.1111/iwj.12049.
2. Jockenhöfer F., Gollnick H., Herberger K., Isbary G., Renner R., Stücker M., Valesky E., Wollina U., Weichenthal M., Karrer S., Ross B., Heintschel von Heinegg E., Dissemond J. Current detection rates of multiresistant gram negative bacteria (3MRGN, 4MRGN) in patients with chronic leg ulcers, (2014) doi: 10.1007/s00105-014-3523-7
3. Salavastru C. M., Nedelcu L. E., Tiplica G-S. Management of leg ulcers in patients with chronic venous insufficiency: the experience of a Dermatology Clinic in Bucharest, Romania, (2012) doi: 10.1111/j.1529-8019.2012.01513.x
4. Koerber A., Schmid E. N., Buer J., Klode J, Schadendorf D., Dissemond J. Bacterial colonization of chronic leg ulcers: current results compared with data 5 years ago in a specialized dermatology department, (2010) doi: 10.1111/j.1468-3083.2010.03570.x
5. Różalski A. Kwil I., Torzewska A., Baranowska M., Stączek P. Bakterie z rodzaju Proteus – cechy i czynniki chorobotwórczości, Postępy Higieny i Medycyny Doświadczalnej (online) 61 (2007), s.204-219
6. Matusiak D. Zakażenia układu moczowego z udziałem Proteus mirabilis – rola biofilmu i inkrustacji cewnika urologicznego, Postępy mikrobiologii, 53(2) (2014), s.173–181
7. Morgenstein R. M., Szostek B., Rather P. N. Regulation of gene expression during swarmer cell differentiation in Proteus mirabilis, FEMS Microbiology Reviews, 34(5) (2010), s.753-763 https://doi.org/10.1111/j.1574-6976.2010.00229.x
8. Armbruster Ch. E., Hodges S. A., Mobley H. L. T. Initiation of swarming motility by Proteus mirabilis occurs in response to specific cues present in urine and requires excess l-glutamine, (2013) doi: 10.1128/JB.02136-12
9. Pellegrino R., Galvalisi U., Scavone P., Sosa V., Zunino P. Evaluation of Proteus mirabilis structural fimbiral proteins as antigens against urinary tract infection, FEMS Mecial Microbiology and Immunology, 36(1-2) (2003), s.103-110. https://doi.org/10.1016/S0928-8244(03)00103-2
10. Schaffer N. J., Pearson M. M. Proteus mirabilis and urinary tract infections, Microbiology Spectrum, 3(5) (2015) doi: 10.1128/microbiolspec.UTI-0017-2013.
11. Kręgiel D. Zastosowanie luminometrii w badaniach adhezji drobnoustrojów do powierzchni abiotycznych, LAB 2016 (4)
12. Kołwzan B., Analiza zjawiska biofilmu– warunki jego powstawania i funkcjonowania, Ochrona Środowiska, 33(4) (2011), s.3-14
13. Czaczyk K., Myszka K. Mechanizmy warunkujące oporność biofilmów bakteryjnych na czynniki antymikrobiologiczne, Biotechnologia 1, (76) (2007), s.40–52
14. Younis K. M., Usup G., Ahmad A. Secondary metabolites produced by marine streptomyces as antibiofilm and quorum-sensing inhibitor of uropathogen Proteus mirabilis, Environmental Science and Pollution Research, 23 (2016), s.4756–4767 doi:10.1007/s11356-015-5687-9

Jana Przekwas, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
124
15. Myszka K., Czaczyk K. Mechanizm quorum sensing jako czynnik regulujący wirulencję bakterii Gram-ujemnych, Postępy Higieny i Medycyny Doświadczalnej (online) 64 (2010), s.582-589 www.phmd.pl e-ISSN 1732-2693
16. Wilczyńska A. Skład chemiczny i właściwości antyoksydacyjne miodu manuka, Problemy Higieny i Epidemiologii 94(4) (2013), s.873-875
17. Mavric E., Wittmann S., Barth G., Henle T. Identification and quantification of methylglyoxal as the dominant antibacterial constituent of Manuka (Leptospermum scopiarum) honeys from New Zealand, (2008) doi: 10.1002/mnfr.200700282
18. Adams Ch. J., Boult Ch. H., Deadman B. J, Farr J. M., Grainger M. N. C., Manley-Harris M., Snow M. J. Isolation by HPLC and characterisation of the bioactive fraction of New Zealand manuka (Leptospermum scopiarum) honey, (2008) https://doi.org/10.1016/j.carres.2007.12.011
19. Lu J., Carter D. A., Turnbull L., Rosendale D., Hedderley D., Stephens J., Gannabathula S., Steinhorn G., Schlothauer R. C., Whitchurch C. B., Harry E. J. The effect of New Zealand Kanuka, Manuka and Clover Honeys on bacterial growth dynamics and cellular morphology varies according to the species, (2013) https://doi.org/10.1371/journal.pone.0055898
20. Kato Y., Umeda N., Maeda A. Matsumoto D., Kitamoto N., Kikuzaki H. Identification of a novel glycoside, leptosin, as a chemical marker of Manuka honey, (2012) doi: 10.1021/jf300068w
21. Lepaer D. J., Schultz G., Carville K., Fletcher J., Swanson T., Drake R. Extending the TIME concept: what have we learned in the past 10 years?, (2012) doi: 10.1111/j.1742-481X.2012. 01097.x
22. Pasich E., Walczewska M., Pasich A., Marcinkiewicz J. Mechanizm i czynniki powstawania biofilmu bakteryjnego jamy ustnej, Postępy Higieny i Medycyny Doświadczalnej (online), 67 (2013), s.736-741 e-ISSN 1732-2693
23. James G. A., Swogger E., Wolcott R., Pulcini E., Secor P., Sestrich J., Costerton J. W., Stewart P. S. Biofilms in chronic wounds, Wound Repair and Regeneration, 16(1) (2008), s.37-44
24. Miguel M. G., Antunes M. D., Faleiro M. L. Honey as a complementary medicine, (2017) doi: 10.1177/1178633717702869
25. Roberts A. E. L., Brown H. L., Jenkins R. On the antibacterial effects of Manuka honey: mechanistic insights, Research and Reports in Biology, 6 (2015), s.215-224 doi:10.2147/RRB.S75754
26. Majtan J. Methyglyoxal – a potential risk factor of Manuka honey in healing of diabetic ulcers, Evidence-Based Complementary and Alternative Medicine, (2011), Article ID 295494, 5 str. http://dx.doi.org/10.1093/ecam/neq013
27. Majtan J., Bohova J., Horniackova M., Klaudiny J. Anti-biofilm effects of honey against wound pathogens Proteus mirabilis and Enterobacter cloacae, Phytotherapy Research, 28(10) (2014), s.69-75 doi: 10.1002/ptr.4957
28. Emineke S., Cooper A.J., Fouch S. Diluted honey inhibits biofilm formation: potential application in urinary catheter management?, Journal of Clinical Pathology, 70 (2017), s.140-144
29. Abbas H. A. Comparative antibacterial and antibiofilm activities of Manuka Honey and Egyptian Clover Honey, Asian Journal of Applied Sciences, 2(2) (2014), ISSN: 2321 – 0893
30. Maddocks S. E., Lopez M. S., Rowlands R. S., Cooper R. A. Manuka honey inhibits the development of Streptococcus pyogenes biofilms and causes reduced expression of two fibronectin binding proteins, Microbiology, 158 (2012), s.781–790 doi: 10.1099/mic.0.053959-0

Wpływ miodu Manuka na tworzenie biofilmu i wzrost mgławicowy Proteus mirabilis
125
31. Roberts A. E. L., Maddocks S. E., Cooper R. A. Manuka honey reduces the motility of Pseudomonas aeruginosa by suppression of flagella-associated genes, Journal of Antimicrobial Chemotherapy, 70(3) (2015), s.716-725 doi: 10.1093/jac/dku448
32. Rabie E., Serem J. C., Oberholzer H. M., Marques Gaspar A. R., Bester M. J. How methylglyoxal kills bacteria: An ultrastructural study, Ultrastructural Pathology, 40(2) (2016), s.107-111 http://dx.doi.org/10.3109/01913123.2016.1154914
33. Tirado D. J., Hudson N. R., Maldonado C. J. Efficacy of medical grade honey against multidrug-resistant organisms of operational significance: Part I, (2014) doi: 10.1097/TA.0000000000000324
34. Jenkins R., Burton N., Cooper R. Proteomic and genomic analysis of methicillin-resistant Staphylococcus aureus (MRSA) exposed to Manuka honey in vitro demonstrated down-regulation of virulence markers, Journal of Antimicrobial Chemotherapy, 69(3) (2014), s.603-615 https://doi.org/10.1093/jac/dkt430
Wpływ miodu Manuka na tworzenie biofilmu i wzrost rozpełzły Proteus mirabilis
Streaszczenie Oporność drobnoustrojów na antybiotyki to obecnie jeden z najpoważniejszych problemów współczesnej medycyny. Wymusza to konieczność poszukiwania alternatywnych metod zwalczania drobnoustrojów, np. z zastosowaniem produktów pszczelich. W pracy oceniano wpływ miodu Manuka w stężeniu 5,0, 12,5 i 25,0% na tworzenie biofilmu oraz wzrost mgławicowy 17 szczepów P. mirabilis izolowanych z przewlekłych ran chorych leczonych w Szpitalu Uniwersyteckim im. dra A. Jurasza w Bydgoszczy oraz szczepu wzorcowego z kolekcji kultur. Biofilm poddany dobowemu działaniu badanych stężeń miodu Manuka tworzono w 96-dołkowych płytkach polistyrenowych. Po dobie inkubacji komórki biofilmu uwalniano z wykorzystaniem ultradźwięków, a uzyskane zawiesiny rozcieńczano i posiewano na stałe podłoże TSA. Zahamowanie wzrostu mgła-wicowego oceniano po 24 godzinach inkubacji w 37°C i następnie po 48 i 120 h inkubacji w temperaturze pokojowej. Wykazano, że miód Manuka w każdym z badanych stężeń ogranicza tworzenie biofilmu przez szczepy P. mirabilis. Jego działanie stawało się efektywniejsze wraz ze wzrostem stężenia. Miód Manuka ograniczał wzrost rozpełzły badanych szczepów. Przepasażowanie szczepów namnażanych na podłożu z 25% mio-dem Manuka na podłoże bez miodu uniemożliwia bakteriom wzrost mgławicowy. Słowa kluczowe: miód Manuka, biofilm, wzrost rozpełzły, Proteus mirabilis
Impact of Manuka honey on biofilm forming and swarming motility of Proteus mirabilis
Abstract Nowadays, bacteria resistance to antibiotics is one of the most important problems of modern medicine. This issue forces to look for alternative methods of fighting microorganisms, e. g. using bees’ products. In studies, we examined impact of Manuka honey in 5,0, 12,5 and 25,0% concentration on biofilm forming and swarming ability of 17 P. mirabilis strains isolated from chronic wounds of patients treated in University Hospital of dr A. Jurasz in Bydgoszcz and standard strain from culture collection. Biofilm treated with examined concentrations of Manuka honey was forming in 96-well polystyrene plates for 24 hours. After 24-hour incubation biofilm cells were liberated using sonication. Then, dilutions was made of obtained suspensions and inoculated on tryptose-soy agar. The swarming ability inhibiton was assessed after 24 hours of 37°C incubation and after 48 and 120 hours of room temperature incubation. The studies have shown that Manuka honey in each examined concentration restricts biofilm forming of P. mirabilis strains. Its activity was more effective with higher concentrations. Manuka honey also restricted swarming ability of examined P. mirabilis strains. Inoculation of strains growing on agar with addition of 25% Manuka honey on fresh tryptose-soy agar prevents the swarming ability of strains. Keywords: Manuka honey, biofilm, swarming motility, Proteus mirabilis

126
Anna Minkiewicz1, Kamila Strom
2, Bartłomiej Iwańczyk
3, Piotr Wychowański
4,
Sylwia Jarzynka5, 6
, Gabriela Olędzka7
Wpływ prądu stałego na formowanie biofilmu
Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych
1. Wstęp
W skład mikrobiomu jamy ustnej ludzi wchodzi wiele różnych grup mikroorga-
nizmów, w tym bakterie, grzyby, pierwotniaki i wirusy [1]. Bakterie stanowią najlicz-
niejszą grupę, około 700 różnych gatunków zasiedlających całą powierzchnię jamy
ustnej i gardła. W każdym mililitrze śliny znajduje się od 108 do 10
9 komórek bakterii,
występujących w postaci rozproszonych komórek – forma planktoniczna. Stanowią
one głównie mikrobiom posiadający fizjologiczną funkcję w jamie ustnej.
W grupie pacjentów ze schorzeniami jamy ustnej, szczególnie z wszczepami
stomatologicznymi, uzupełnieniami protetycznymi oraz implantami, obserwowane są
zmiany składu gatunkowego oraz liczebności poszczególnych grup mikroorganizmów
jamy ustnej. Zaburzenia mikrobiomu oraz utrudniona higiena powierzchni zbudo-
wanych z biomateriałów dentystycznych, prowadzą do rozwoju biofilmu bakteryjnego
[1, 2]. Ta przestrzennie zorganizowana struktura może powodować zapalenia dziąseł,
ze szczególnym uwzględnieniem periimplantitis, zakażenia tkanek miękkich i twar-
dych w miejscach implantacji i okołowszczepowych, co może prowadzić nawet do
usunięcia wszczepu [3,4]. Aktualnie główny problem kliniczny stanowi poimplan-
tacyjne tworzenie biofilmu na biomateriałach i tkankach okalających wszczepy, przez
szczepy Staphylococcus aureus, Staphylococcus epidermidis oraz grupę Streptococcus
mutans.
Proces kształtowania się biofilmu, także stomatologicznego (dentystycznego),
rozpoczyna się od etapu adhezji, przylegania komórek bakterii do powierzchni
biotycznych i abiotycznych. W środowisku jamy ustnej obserwuje się adhezję bakterii
1 [email protected], Zakład Biologii Medycznej, Wydział Nauki o Zdrowiu, Warszawski Uniwersytet Medyczny 2 [email protected], Zakład Biologii Medycznej, Wydział Nauki o Zdrowiu, Warszawski
Uniwersytet Medyczny [email protected], Zakład Chirurgii Stomatologicznej, Wydział Lekarsko-Dentystyczny, Warszawski Uniwersytet Medyczny 4 [email protected] Zakład Chirurgii Stomatologicznej, Wydział Lekarsko-Dentystyczny,
Warszawski Uniwersytet Medyczny 5 [email protected], Zakład Biologii Medycznej, Wydział Nauki o Zdrowiu, Warszawski Uniwersytet Medyczny, autor korespondencyjny, tel. 22 6252468 6 Zakład Mikrobiologii, Centralny Szpital Kliniczny Warszawskiego Uniwersytetu Medycznego 7 [email protected] Zakład Biologii Medycznej, Wydział Nauki o Zdrowiu, Warszawski
Uniwersytet Medyczny

Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych
127
do śluzówki dziąseł, szkliwa zębów oraz do biomateriałów stomatologicznych budu-
jących aparaty ortodontyczne i implanty. Na tym etapie, komórki bakteryjne wiążą się
ze śluzówkami i biomateriałami na zasadzie niespecyficznych, odwracalnych oddzia-
ływań sił van der Waalsa (elektrostatycznych czy hydrofobowych) [5]. W zależności
od składu gatunkowego, liczby komórek budujących biofilm oraz właściwości fizycz-
nych powierzchni, tworzący się biofilm może charakteryzować zróżnicowany stopień
przylegania. W kolejnych etapach tworzenia i dojrzewania biofilmu, jego skład może
ulec zmianie. Obserwowane są biofilmy mieszane, zbudowane z różnych rodzajów
bakterii lub różnych mikroorganizmów [5-7]. Dojrzały biofilm dentystyczny może
składać się z blisko 100 rożnych gatunków mikroorganizmów, w tym również grzy-
bów, pierwotniaków, Archaea i wirusów [2, 8]. Dojrzewanie biofilmu składa się
z następujących etapów: otoczenie mikroorganizmów macierzą składającą się między
innymi z wielocukrowych polimerów oraz białek (z ang. extracellular polimeric
substances – EPS), zmiany w procesach metabolicznych z wytworzeniem warunków
beztlenowych wewnątrz biofilmu, ścisłe nieodwracalne połączenie struktury biofilmu
z podłożem [3, 8, 9]. Wielokomórkowa struktura biofilmu utrudnia działania obronne
układu immunologicznego gospodarza oraz dostęp i przenikalność leków przeciwdrob-
noustrojowych.
Formowanie biofilmu jest związane także z charakterystyką biomateriałów
stomatologicznych. Istotną właściwością biomateriałów stosowanych w stomatologii
jest szorstkość ich powierzchni. Cecha ta umożliwia łatwiejszą adhezję bakterii do
powierzchni biomateriałów i wytwarzanie na ich powierzchni biofilmu niż tworzenie
tej struktury na powierzchni śluzówek i szkliwa [10]. Dlatego też, współczesna
implantologia stomatologiczna koncentruje się na wykorzystywaniu materiałów
wykazujących utrudnioną zdolność adhezji i tworzenia biofilmu na ich powierzchni.
Stale opracowywane są skuteczne i nieinwazyjne metody oczyszczania biomateriałów,
wszczepów i uzupełnień protetycznych z kolonizacji bakteryjnej.
Z biomateriałów stomatologicznych wykazujących niskie powikłania stomatolog-
giczne, na uwagę zasługują tytan i dwutlenek cyrkonu.
Tytan to jeden z biomateriałów wykazujących najniższy wskaźnik adhezji bakterii
do jego powierzchni. Wykazuje on najprawdopodobniej działanie bakteriostatyczne [9-
11]. Tytan i jego stopy dzięki swoim właściwościom fizykochemicznym, mechanicz-
nym oraz biologicznym wykazują dużą tolerancje w środowisku tkankowym pacjenta.
Mają niskie przewodnictwo cieplne i elektryczne nie posiadają również właściwości
magnetycznych. Tytan wykorzystywany jest między innymi w uzupełnieniach prote-
tycznych stałych: wkłady, nakłady, korony (teleskopowe, licowane porcelaną lub
innymi materiałami), mosty oraz w uzupełnieniach ruchomych: protezy szkieletowe,
płyty protez całkowitych a także do tworzenia wkładów koronowo-korzeniowych,
ćwieków okołomiazgowych [12].
Dwutlenek cyrkonu należy do grupy ceramik stosowanych powszechnie w stoma-
tologii i używanych do maskowania szkieletu metalowych koron. Wykazuje on jeden
z najniższych stopni adhezji bakterii ze wszystkich ceramik [9, 11, 13]. Materiał ten
jest od niedawna stosowany w protetyce stomatologicznej, cechuje się wysoką wytrzy-

Anna Minkiewicz, Kamila Strom, Bartłomiej Iwańczyk, Piotr Wychowański,
Sylwia Jarzynka, Gabriela Olędzka
128
małością mechaniczną, ma korzystne właściwości fizyko-mechaniczne, a przy tym jest
biokompatybilny. W stomatologii wykorzystywany jest między innymi do produkcji
implantów, łączników do implantów, wkładów koronowo-korzeniowych na
podbudowy koron i mostów oraz zamków ortodontycznych [14, 15].
Duże znaczenie kliniczne ma opracowanie metod: skutecznego leczenia zapobiega-
jącego powstawaniu biofilmu, usuwania powstałego biofilmu lub wprowadzenie
innych działań przeciwbakteryjnych zatrzymujących proces formowania biofilmu
stomatologicznego. Jedna z metod inhibicji biofilmu a także uszkodzenia jego struk-
tury, która szeroko była dyskutowana w literaturze, polega na zastosowaniu krótko-
trwałego prądu stałego, o niskim natężeniu do 10 mA, bezpiecznego dla człowieka, na
tworzący się biofilm na biomateriałach [16-19]. W trakcie opracowania są już
urządzenia stomatologiczne, które mogłyby działać w oparciu o w/w metodę. Metoda
ta może być nie tylko alternatywą dla pacjentów z lekoopornymi szczepami bakterii,
ale także może ograniczyć ilość stosowanych antybiotyków oraz zmniejszyć ilość
powikłań po antybiotykoterapii [2, 4, 6, 17]. Metoda wykorzystująca prąd stały
o niskim natężeniu może być nie tylko metodą leczniczą, ale także zapobiegającą
formowaniu biofilmu, co wiąże się ze zmniejszeniem stanów zapalnych, chorobowych
i zakażeń w obrębie jamy ustnej.
2. Cel
Celem badań była ocena in vitro wpływu prądu stałego, o niskim natężeniu na
formowanie biofilmu Staphylococcus aureus na biomateriałach stomatologicznych.
3. Materiał i metody
Materiał do badań stanowiły szczepy S. aureus pozyskane od pacjentów leczonych
w Zakładzie Chirurgii Stomatologicznej Warszawskiego Uniwersytetu Medycznego.
Zdolność tworzenia biofilmu oznaczono na krążkach z tytanu (Ti6Al4V) i cyrkonu
(ZrO2) (średnica 1 cm), biomateriałach stomatologicznych [Rys.1]. Klasyfikację
szczepów ze względu na zdolność do tworzenia biofilmu dokonano na podstawie
odczytu absorbancji (=584nm) mierzonej spektrofotometrycznie, na płytkach
titracyjnych barwionych fioletem krystalicznym. Szczepy wykazujące niską zdolność
tworzenia biofilmu oznaczono, jako mały biofilm (absorbancja ≤ 0,24) i średni biofilm
(absorbancja 0,24-0,39). Szczepy cechujące się wysoką zdolnością tworzenia biofilmu
określono, jako duży biofilm (absorbancja > 0,39) (dane niepublikowane).
Erozję i inhibicję biofilmu formowanego przez 3 wybrane szczepy Staphylococcus
aureus, wykazujące zróżnicowaną zdolność do tworzenia biofilmu, badano z użyciem
prądu stałego o natężeniu 10 mA, wykorzystując zjawisko elektrolizy, w którym anodę
stanowiły wykorzystane do doświadczenia krążki biomateriałowe. Hodowlę S. aureus
prowadzano na podłożu płynnym LB (Luria-Bertani) z dodatkiem 1% glukozy przez
24 godziny w 37°C z wytrząsaniem.

Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych
129
Rysunek 1. Krążki tytanowe (Ti6Al4V) i cyrkonowe (ZrO2) [źródło: opracowanie własne]
Badanie erozji powstałego biofilmu
Krążki tytanowe (Ti6Al4V) i cyrkonowe (ZrO2) inkubowano w hodowli S. aureus
przez 24 godziny w 37°C. W kolejnym etapie krążki z powstałym biofilmem, poddano
działaniu prądu stałego (natężenie 10 mA) w roztworze PBS, w trzech powtórzeniach
w czasie: 0 min, 10 min i 60 min. Następnie krążki płukano w 10ml roztworu PBS
poprzez wytrząsanie z użyciem wstrząsarki typu Vortex. W kolejnym etapie metodą
seryjnych rozcieńczeń w roztworze PBS, oznaczono liczbę jednostek tworzących
kolonie (JTK)/ml mikroorganizmów budujących powstały biofilm.
Badanie inhibicji tworzącego się biofilmu
Krążki tytanowe (Ti6Al4V) i cyrkonowe (ZrO2) inkubowano w hodowli S. aureus
przez 2 godziny w 37°C. Następnie krążki wraz z wstępną fazą struktury biofilmu,
poddano działaniu prądu stałego (10 mA) w roztworze PBS, w trzech powtórzeniach
w czasie: 0 min, 10 min i 60 min. Następnie krążki płukano w 10ml roztworu PBS
poprzez wytrząsanie z użyciem wstrząsarki typu Vortex. W kolejnym etapie metodą
seryjnych rozcieńczeń w roztworze PBS, oznaczono JTK/ml mikroorganizmów.
Analizę statystyczną uzyskanych danych wykonano z użyciem testu T (Statistica
13.1) z poziomem istotności p = 0,05.
4. Wyniki
Szczepy S. aureus wykazywały zróżnicowaną zdolność tworzenia biofilmu na
biomateriałach stomatologicznych, zarówno po 2, jak i po 24 godz. inkubacji. Ogólna
wyjściowa liczba komórek każdego szczepu S. aureus, w hodowli płynnej wynosiła od
6 do 6,9 JTK/ml.
W badaniach zaobserwowano spadek liczby jednostek tworzących kolonie
(JTK)/ml, w trakcie erozji oraz podczas inhibicji, zarówno w przypadku dużego,
średniego jak i małego biofilmu S. aureus powstałego na biomateriałach. Spadek
liczby komórek budujących biofilm zarówno na cyrkonie, jak i na tytanie, był zależny
od czasu ekspozycji biofilmu na prąd.

Anna Minkiewicz, Kamila Strom, Bartłomiej Iwańczyk, Piotr Wychowański,
Sylwia Jarzynka, Gabriela Olędzka
130
Średnia liczba komórek bakteryjnych tworzących 24 godz. biofilm na cyrkonie
wynosiła odpowiednio: dla biofilmu dużego (DB) – 3,7 JTK/ml, średniego (ŚB) – 1,8
JTK/ml i małego (MB) – 2,7 JTK/ml. Po 10 min erozji biofilmu na cyrkonie, ozna-
czono spadek liczby komórek, który był statystycznie istotny, odpowiednio dla BD - 3
JTK/ml (p = 0,0002), ŚB – 1,6 JTK/ml (p = 0,0001), MB – 2,2 JTK/ml (p = 0,0019), (p
< 0,05). Po kolejnych 50 min erozji oznaczono statystycznie istotną zmniejszoną liczbę
komórek, dla DB – 2,6 JTK/ml (p = 0,0135), ŚB – 1,2 JTK/ml (p = 0,0013), BM – 1,7
JTK/ml (p = 0,0001), (p < 0,05) [Wykres 1].
Po 2 godz. inkubacji S. aureus na cyrkonie, oznaczono średnią liczbę 3,7 JTK/ml
w dużym biofilmie, 2,8 JTK/ml w ŚB i 3,9 JTK/ml w BM. Po upływie 10 min inhibicji
biofilmu na cyrkonie, średnia liczba komórek w dużym biofilmie istotnie statystycznie
spadła do 3,3 JTK/ml (p = 0,0001), w ŚB – 2,4 JTK/ml (p = 0,0029), a w MB – 3,5
JKT/ml (p = 0,038), (p < 0,05). Po kolejnych 50 min inhibicji średnia liczba komórek
spadła w DB a w przypadku średniego i małego biofilmu pozostała na takim samym
poziomie. Wyniki te nie są istotne statystycznie, odpowiednio dla DB – 3,1 JTK/ml (p
= 0,054), ŚB – 2,4 JTK/ml (p = 0,112), MB – 3,5 JTK/ml (p = 0,097), (p > 0,05)
[Wykres 2].
W 24 godz. hodowli S. aureus na krążkach tytanowych, oznaczono średnią liczbę
3,7 JTK/ml w dużym biofilmie, 2,7 JTK/ml w średnim oraz 2,2 JTK/ml w małym.
W wyniku 10 min erozji biofilmu na tytanie, zaobserwowano statystycznie istotny
spadek liczby komórek bakteryjnych tworzących DB do 3 JTK/ml (p = 0,0001), ŚB –
2,6 JTK/ml (p = 0,0097) , MB – 1,3 JTK/ml (p = 0,028), (p < 0,05). Po kolejnych 50
min erozji biofilmu oznaczono spadek JTK/ml istotny statystycznie jedynie dla
średniego – 2,3 JTK/ml (p = 0,0018) i małego biofilmu – 1,3 JTK/ml (p = 0,0016)
[Wykres 3].
Z kolei, w trakcie inhibicji dużego, średniego i małego biofilmu S. aureus na
krążkach tytanowych, oznaczono statystycznie istotny spadek liczby komórek
tworzących kolonie w odniesieniu do czasu działania prądu o niskim natężeniu: po 10
min odpowiednio, DB – 3,3 JTK/ml (p = 0,0002), ŚB – 2,4 JTK/ml (p = 0,0047), MB
– 1,6 JTK/ml (p = 0,014); po kolejnych 50 min odpowiednio, DB – 2,9 JTK/ml (p =
0,029), ŚB – 2,2 JTK/ml (p = 0,0086), MB – 1,5 JTK/ml (p = 0,0013), (p < 0,05).
[Wykres 4].
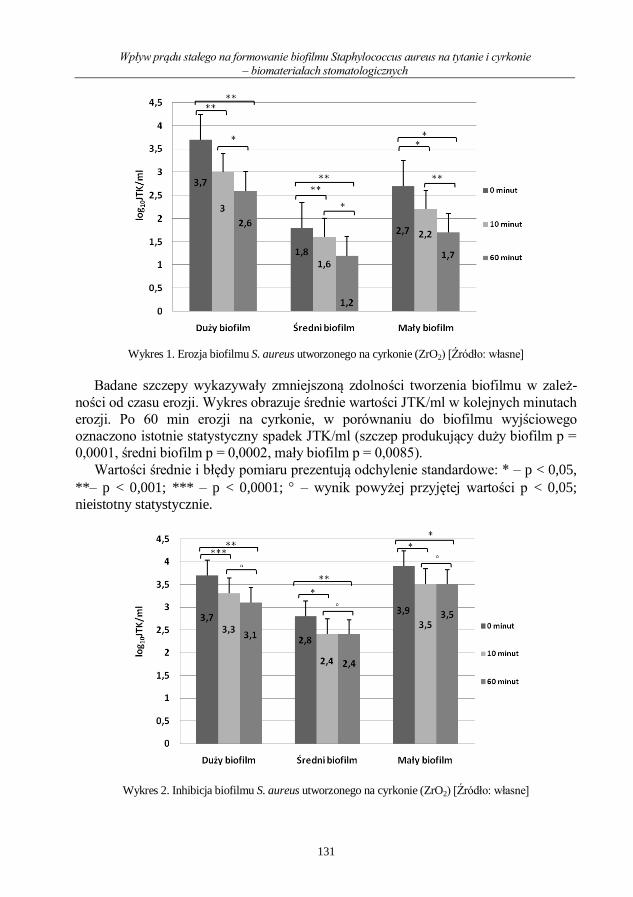
Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych
131
Wykres 1. Erozja biofilmu S. aureus utworzonego na cyrkonie (ZrO2) [Źródło: własne]
Badane szczepy wykazywały zmniejszoną zdolności tworzenia biofilmu w zależ-
ności od czasu erozji. Wykres obrazuje średnie wartości JTK/ml w kolejnych minutach
erozji. Po 60 min erozji na cyrkonie, w porównaniu do biofilmu wyjściowego
oznaczono istotnie statystyczny spadek JTK/ml (szczep produkujący duży biofilm p =
0,0001, średni biofilm p = 0,0002, mały biofilm p = 0,0085).
Wartości średnie i błędy pomiaru prezentują odchylenie standardowe: * – p < 0,05,
**– p < 0,001; *** – p < 0,0001; – wynik powyżej przyjętej wartości p < 0,05;
nieistotny statystycznie.
Wykres 2. Inhibicja biofilmu S. aureus utworzonego na cyrkonie (ZrO2) [Źródło: własne]
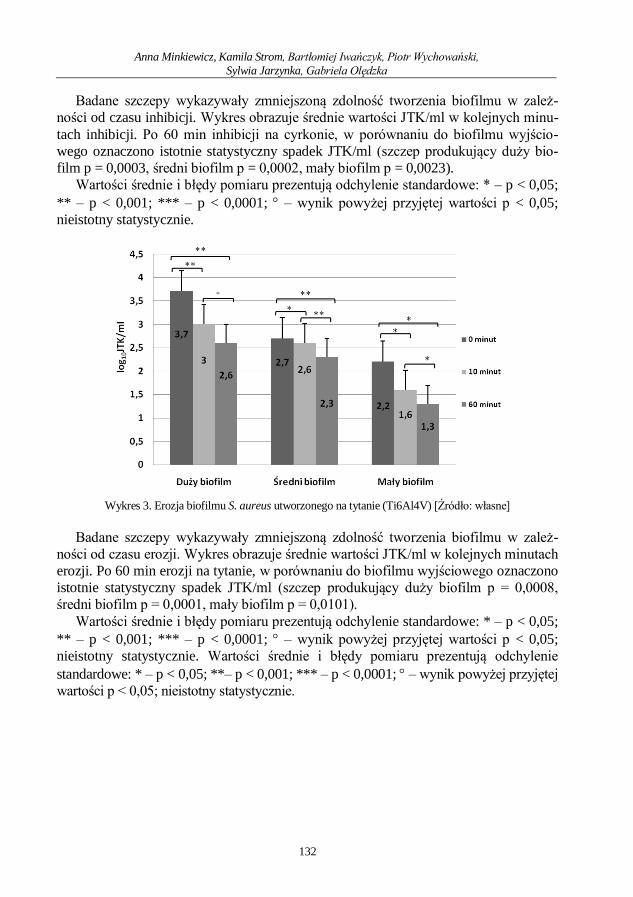
Anna Minkiewicz, Kamila Strom, Bartłomiej Iwańczyk, Piotr Wychowański,
Sylwia Jarzynka, Gabriela Olędzka
132
Badane szczepy wykazywały zmniejszoną zdolność tworzenia biofilmu w zależ-
ności od czasu inhibicji. Wykres obrazuje średnie wartości JTK/ml w kolejnych minu-
tach inhibicji. Po 60 min inhibicji na cyrkonie, w porównaniu do biofilmu wyjścio-
wego oznaczono istotnie statystyczny spadek JTK/ml (szczep produkujący duży bio-
film p = 0,0003, średni biofilm p = 0,0002, mały biofilm p = 0,0023).
Wartości średnie i błędy pomiaru prezentują odchylenie standardowe: * – p < 0,05;
** – p < 0,001; *** – p < 0,0001; – wynik powyżej przyjętej wartości p < 0,05;
nieistotny statystycznie.
Wykres 3. Erozja biofilmu S. aureus utworzonego na tytanie (Ti6Al4V) [Źródło: własne]
Badane szczepy wykazywały zmniejszoną zdolność tworzenia biofilmu w zależ-
ności od czasu erozji. Wykres obrazuje średnie wartości JTK/ml w kolejnych minutach
erozji. Po 60 min erozji na tytanie, w porównaniu do biofilmu wyjściowego oznaczono
istotnie statystyczny spadek JTK/ml (szczep produkujący duży biofilm p = 0,0008,
średni biofilm p = 0,0001, mały biofilm p = 0,0101).
Wartości średnie i błędy pomiaru prezentują odchylenie standardowe: * – p < 0,05;
** – p < 0,001; *** – p < 0,0001; – wynik powyżej przyjętej wartości p < 0,05;
nieistotny statystycznie. Wartości średnie i błędy pomiaru prezentują odchylenie
standardowe: * – p < 0,05; **– p < 0,001; *** – p < 0,0001; – wynik powyżej przyjętej
wartości p < 0,05; nieistotny statystycznie.
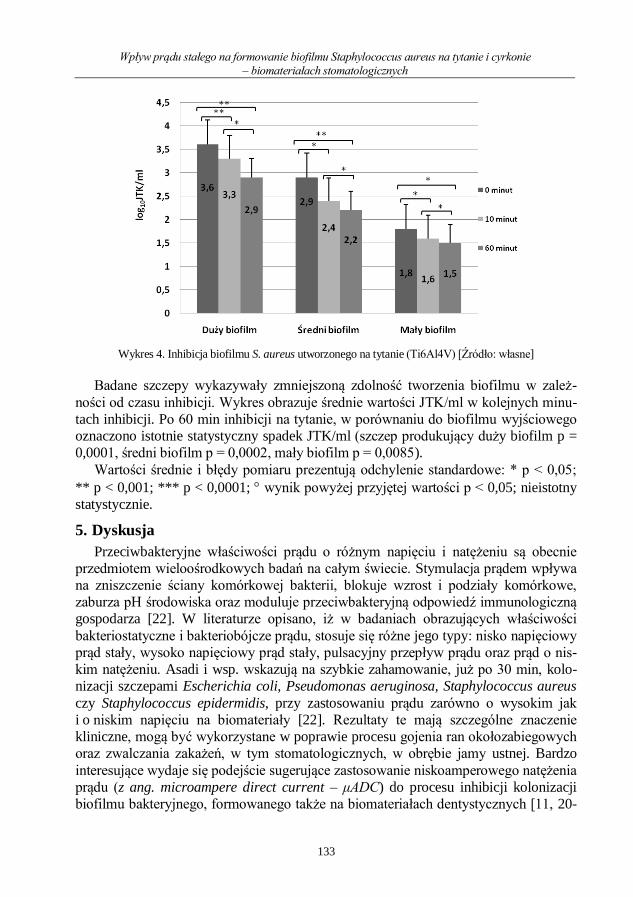
Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych
133
Wykres 4. Inhibicja biofilmu S. aureus utworzonego na tytanie (Ti6Al4V) [Źródło: własne]
Badane szczepy wykazywały zmniejszoną zdolność tworzenia biofilmu w zależ-
ności od czasu inhibicji. Wykres obrazuje średnie wartości JTK/ml w kolejnych minu-
tach inhibicji. Po 60 min inhibicji na tytanie, w porównaniu do biofilmu wyjściowego
oznaczono istotnie statystyczny spadek JTK/ml (szczep produkujący duży biofilm p =
0,0001, średni biofilm p = 0,0002, mały biofilm p = 0,0085).
Wartości średnie i błędy pomiaru prezentują odchylenie standardowe: * p < 0,05;
** p < 0,001; *** p < 0,0001; wynik powyżej przyjętej wartości p < 0,05; nieistotny
statystycznie.
5. Dyskusja
Przeciwbakteryjne właściwości prądu o różnym napięciu i natężeniu są obecnie
przedmiotem wieloośrodkowych badań na całym świecie. Stymulacja prądem wpływa
na zniszczenie ściany komórkowej bakterii, blokuje wzrost i podziały komórkowe,
zaburza pH środowiska oraz moduluje przeciwbakteryjną odpowiedź immunologiczną
gospodarza [22]. W literaturze opisano, iż w badaniach obrazujących właściwości
bakteriostatyczne i bakteriobójcze prądu, stosuje się różne jego typy: nisko napięciowy
prąd stały, wysoko napięciowy prąd stały, pulsacyjny przepływ prądu oraz prąd o nis-
kim natężeniu. Asadi i wsp. wskazują na szybkie zahamowanie, już po 30 min, kolo-
nizacji szczepami Escherichia coli, Pseudomonas aeruginosa, Staphylococcus aureus
czy Staphylococcus epidermidis, przy zastosowaniu prądu zarówno o wysokim jak
i o niskim napięciu na biomateriały [22]. Rezultaty te mają szczególne znaczenie
kliniczne, mogą być wykorzystane w poprawie procesu gojenia ran okołozabiegowych
oraz zwalczania zakażeń, w tym stomatologicznych, w obrębie jamy ustnej. Bardzo
interesujące wydaje się podejście sugerujące zastosowanie niskoamperowego natężenia
prądu (z ang. microampere direct current – μADC) do procesu inhibicji kolonizacji
biofilmu bakteryjnego, formowanego także na biomateriałach dentystycznych [11, 20-

Anna Minkiewicz, Kamila Strom, Bartłomiej Iwańczyk, Piotr Wychowański,
Sylwia Jarzynka, Gabriela Olędzka
134
22]. Według wielu autorów tą metodą można uzyskać dużo większy efekt bakterio-
statyczny niż w przypadku zastosowania innych metod eradykacji biofilmu [22-24].
Podobne, obiecujące wyniki uzyskiwane są przy zastosowaniu prądu stałego o niskim
natężeniu 5-20 mA (z ang. low intensity direct current – LIDC).
W badaniach własnych zastosowano LIDC prąd stały o niskim natężeniu 10 mA.
Oznaczono działanie prądu na tworzenie (inhibicja) i zwalczanie (erozja) biofilmu S.
aureus formowanego na powierzchni biomateriałów tytanowych i cyrkonowych.
Bakteriobójcze działanie 10 mA prądu zaobserwowano już po 10 minutach jego
wpływu (p < 0,05). W przypadku badania erozji biofilmu, największy spadek liczby
jednostek tworzących kolonie oznaczono dla dużego biofilmu zarówno na cyrkonie,
jak i na tytanie, z 3,7 do 3 JTK/ml (p = 0,001), po 10 minutach. Obserwowany istotny
statystycznie spadek liczby komórek tworzących biofilm w krótkim czasie oraz
nieinwazyjność testowanej metody, stwarza potencjalną możliwość jej wykorzystania
w profilaktyce oraz terapii u pacjentów z uzupełnieniami stomatologicznymi. Del Pozo
i wsp. zaobserwowali również zmniejszoną żywotność szczepów S. epidermidis, S.
aureus i P. aeruginosa po stymulacji prądem stałym o niskim natężeniu od 0,002 mA
do 2 mA [24]. Podobne wyniki uzyskali Maadi i wsp. dla szczepów P. aeruginosa
[25]. Z kolei wyniki badań przeprowadzonych przez Petrofsky i wsp. pokazują większą
skuteczność przeciwbakteryjną, szczególnie wobec P. aeruginosa, prądu zmiennego
o niskim natężeniu 5-20 mA [26]. W literaturze nie obserwowano, jak dotąd, wzrostu
bakterii po zastosowaniu prądu stałego o niskim natężeniu.
Metoda wykorzystana w badaniach własnych może mieć szczególne znaczenie
w zwalczaniu biofilmów tworzonych na biomateriałach stomatologicznych u pacjen-
tów z periimplantitis (zapalenie tkanek wokół implantu) [27]. W badaniach własnych
do porównania właściwości prądu stałego na tworzący się biofilm wybrano dwa bio-
materiały o podobnych właściwościach adhezyjnych: tytan i dwutlenek cyrkonu.
Niektórzy badacze, w tym Sahrmann i wsp., wykazali bakteriobójcze działanie po 10
min działania prądu stałego o niskim natężeniu 10 mA i 15 mA, na biofilm tworzony
na implantach tytanowych [28,29]. Autorzy badali 40,5 godzinny biofilm tworzony
przez paciorkowce Streptococcus oralis i Streptococcus anginosus oraz beztlenowce
Actinomyces oris, Fusobacterium nucleatum subsp. nucleatum, Veillonella dispar,
Campylobacter rectus, Prevotella intermedia oraz Porphyromonas gingivalis, które
stanowią znaczny odsetek mikrobiomu jamy ustnej. W pracy obserwowano 3 i 4-
krotne zmniejszenie liczby żywych komórek bakterii [28,29]. W badaniach własnych
zaobserwowano podobną zależność. Podczas erozji biofilmu na krążkach tytanowych,
oznaczono ogólny spadek liczby komórek tworzących biofilm zarówno dużego z 3,7
do 2,6 JTK/ml (p < 0,001), średniego z 2,7 do 2,3 JTK/ml (p < 0,01), jak i małego
biofilmu z 2,2 do 1,3 JTK/ml (p < 0,05). Liczba komórek istotnie zmniejszała się
również po 60 min inhibicji biofilmu na tytanie, zarówno dużego z 3,6 do 2,9 JTK/ml
(p < 0,05), średniego z 2,9 do 2,2 JTK/ml (p < 0,001), jak i małego biofilmu z 1,8 do
1,5 JTK/ml (p < 0,05).
Wysoką zdolność tworzenia biofilmu przez S. aureus zaobserwowano również na
cyrkonie (duży biofilm). Zarówno w dużym, średnim i małym biofilmie na cyrkonie

Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych
135
podczas erozji jak i inhibicji oznaczono spadek liczby komórek po upływie już 10 min,
a także po upływie 60 min. W dostępnej literaturze nie odnotowano innych, po za
własnymi, badań oceniających przeciwbakteryjne działanie prądu na biofilm formo-
wany na cyrkonie. Jednakże inni badacze wykazują możliwość tworzenia biofilmu
bakteryjnego na powierzchniach cyrkonowych [30,31]. Wyniki badań własnych
wskazują na skuteczne działanie prądu stałego o niskim natężeniu na inhibicję i erozję
biofilmu bakteryjnego na cyrkonie.
Uzyskane rezultaty pokazują możliwe wykorzystanie prądu stałego o niskim natę-
żeniu w profilaktyce i leczeniu zwiększonej kolonizacji i zakażeń bakteriami tworzą-
cymi biofilm zarówno na śluzówkach, uzębieniu, elementach kostnych jak i biomate-
riałach sztucznych mających zastosowanie w stomatologii. W kolejnym etapie badań
warto ocenić działanie bakteriobójcze prądu stałego o niskim natężeniu na tworzenie
biofilmu na innych biomateriałach stosowanych w stomatologii. Obiecujące wyniki
badań in vitro skłaniają również do przeprowadzenia badań in vivo w grupie pacjentów
z przewlekłymi schorzeniami jamy ustnej oraz leczonych protetycznie i implanto-
logicznie.
6. Wnioski
Wysoką zdolność tworzenia biofilmu przez szczepy S. aureus uzyskano na
cyrkonie.
Prąd stały o niskim napięciu może hamować już po 10 minutach tworzenie
biofilmu S. aureus, szczególnie na wyrobach tytanowych.
Literatura
1. Dewhirst F.E., Chen T., Izard J., Paster B.J., Tanner A.C.,Yu W-H., Lakshmanan A., Wade
W.G., The human oral microbiome, Journal of Bacteriology., 192 (2010), s. 5002-5017.
2. Satpathy S., Sen S.K., Pattanaik S., Raut S., Review on bacterial biofilm: An universal
cause of contamination, Biocatalysis and Agricultural Biotechnology., 7 (2016), s. 56-66.
3. Huang R., Li M., Gregory R.L., Bacterial interactions in dental biofilm, Virulence., 2
(2011), s. 435-444.
4. Hanif A., Qureshi Z., Sheikh Z., Rashid H., Complications in implant dentistry, European
Journal of Dentistry., 11 (2017), s. 135-140.
5. Hori K., Matsumoto S., Bacterial adhesion: From mechanism to control, Biochemical
Engineering Journal., 48 (2010), s. 424-434.
6. Roberts C.G., The role of biofilms in reprocessing medical devices, American Journal of
Infection Control., 41 (2013) s. 77-80.
7. Hojo K., Nagaoka S., Ohshima T., Maeda N., Bacterial interactions in dental biofilm
development, Journal of Dental Research., 88 (2009), s. 982-990.
8. Do T., Devine D.A., Marsh P.D., Oral biofilms: molecular analysis, challenges, and future
prospects in dental diagnostics, Cosmetic and Investigational Dentistry., 5 (2013), s. 11-19.
9. Subramani K., Jung R.E., Molenberg A., Hammerle C.H., Biofilm on dental implants: A review
of the literature, International Journal of Oral & Maxillofacial Implants ., 4 (2009), s. 616-26.
10. Rosentritt M., Hahnel S., Gröger G., Mühlfriedel B., Bürgers R., Handel G., Adhesion of
Streptococcus mutans to Various Dental Materials in a Laminar Flow Chamber System,

Anna Minkiewicz, Kamila Strom, Bartłomiej Iwańczyk, Piotr Wychowański,
Sylwia Jarzynka, Gabriela Olędzka
136
Wiley Periodicals, Inc. Journal of Biomedical Materials Research Part B: Applied
Biomaterials., 86(B), (2008), s. 36-44.
11. Scarano A., Piattelli M., Caputi S., Piattelli A., Bacterial Adhesion on Commercially Pure
Titanium and Zirconium Oxide Disks: An In Vivo Human Study, Journal of
Periodontology., 75 (2004), s. 292-296.
12. Jedynak B., Mierzwińska-Nastalska E., Tytan – właściwości i zastosowanie w protetyce
stomatologicznej., Dental Forum., 1 (2013), 41.
13. Bremer F., Grade S, Kohorst P, Stiesch M., In vivo biofilm formation on different dental
ceramics, Quintessence International., 42 (2011), s. 565-574.
14. Lasek K., Okoński P., Mierzwińska-Nastalska E., Tlenek cyrkonu – właściwości fizyczne
i zastosowanie kliniczne, Protetyka Stomatologiczna., 59( 6), (2009), s. 415-422.
15. Dejak B., Kacprzak M., Suliborski B, Śmielak B., Struktura i niektóre właściwości ceramik
dentystycznych stosowanych w uzupełnieniach pełnoceramicznych w świetle literatury,
Protetyka Stomatologiczna., 56, (6), (2006), s. 471-477.
16. Mohn D., Matthias Zehnder M., Stark W.J., Imfeld T., Electrochemical Disinfection of
Dental Implants – a Proof of Concept, PLoS One., 6 (2011), e16157.
17. del Pozo J.L., Rouse M.S., Mandrekar J.N., Steckelberg J.M., Patel R., The electricidal effect:
reduction of Staphylococcus and Pseudomonas biofilms by prolonged exposure to low-
intensity electrical current, Antimicrob Agents Chemotherapy., 53 (1), (2009), s. 41-45.
18. Ruiz-Ruigomez M., Badiola J., Schmidt-Malan S.M., Greenwood-Quaintance K., Karau
M.J., Brinkman C.L., Mandrekar J.N., Patel R., Direct electrical current reduces bacterial
and yeast biofilm formation, Int J Bacteriol. 2016:9727810, (2016), doi:
10.1155/2016/9727810.
19. Brinkman C.L., Schmidt-Malan S.M., Karau M.J., Greenwood-Quaintance K., Hassett D.J.,
Mandrekar J.N., Patel R., Exposure of bacterial biofilms to electrical current leads to cell
death mediated in part by reactive oxygen species, PLOS One, 11(12), (2016), doi:
10.1371/journal.pone.0168595.
20. Liu W.K., Brown M.R., Elliott T.S., Mechanisms of the bactericidal activity of low amperage
electric current (DC), Journal of Antimicrobial Chemotherapy., 39 (6), (1997), s. 687-95.
21. Sultana S.T., Babauta J.T., Beyenal H., Electrochemical biofilm control: a review,
Biofouling. 31 (9-10), (2015), s. 745-58.
22. Asadi M.R., Torkaman G., Bacterial Inhibition by electrical stimulation, Advances in
wound care., 3 (2), ( 2014), s. 91-97.
23. Daeschlein G., Assadian O., Kloth L.C., Meinl C., Ney F., Kramer A., Antibacterial
activity of positive and negative polarity low-voltage pulsed current (LVPC) on six typical
Gram-positive and Gram-negative bacterial pathogens of chronic wounds, Wound Repair
and Regeneration. 15 (3), (2007), s. 399-403.
24. Del Pozo J.L., Rouse M.S., Mandrekar J.N., Fernandez Sampedro M., Steckelberg J.M.,
Patel R., Effect of electrical current on the activities of antimicrobialagents against
Pseudomonas aeruginosa, Staphylococcus aureus, and Staphylococcus epidermidis
biofilms, Antimicrobial Agents and Chemotherapy., 53 (2009), s. 35.
25. Maadi H., Haghi M., Delshad R., Kangarloo H., Mohammadnezhady M.A., Hemmatyar
G.R., Effect of alternating and direct currents on Pseudomonas aeruginosa growth in vitro,
African Journal of Biotechnol., 9 (2010) s. 6373.
26. Petrofsky J., Laymon M., Chung W., Collins K., Yang T.N., Effect of electrical stimulation
on bacterial growth, The Journal of Neurological Surgery., 31 (2008) s. 43.

Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie
– biomateriałach stomatologicznych
137
27. Merriman H.L., Hegyi C.A., Albright-Overton C.R., Carlos J., Putnam R.W., Mulcare J.A., A comparison of four electrical stimulation types on Staphylococcus aureus growth in vitro, Journal of Rehabilitation Research & Development., 41 (2004), s. 139.
28. Sahrmann P., Zehnder M., Mohn D.,Meier A., Imfeld T., Thurnheer T., Effect of low direct current on anaerobic multispecies biofilm adhering to a titanium implant surface, Clinical Implant Dentistry and Related Research., 16 (4), (2014), s. 552-556.
29. Sahrmann P., Mohn D., Zehnder M., Stark W.J., Imfeld T., Weber F.E., Effect of direct current on surface structure and cytocompatibility of titanium dental implants, The International Journal of Oral & Maxillofacial Implants., 29 (3), (2014), s. 735-742.
30. Chrzanowska N., Załęska-Radziwiłł M., The impacts of aluminum and zirconiumnano-oxides on planktonic and biofilm bacteria, Desalination and Water Treatment., 52 (19-21) (2014), s. 3680-3689.
31. Scotti R., Zanini Kantorski K., Scotti N., Monaco C., Valandro L.F., Bottino M.A., Early biofilm colonization on polished- and glazed-zirconium ceramicsurface. Preliminary results, Minerva Stomatologica., 55 (9), (2006), s. 493-502.
Wpływ prądu stałego na formowanie biofilmu Staphylococcus aureus na tytanie i cyrkonie – biomateriałach stomatologicznych
Streszczenie W przebiegu leczenia stomatologicznego, formowanie biofilmu bakteryjnego na biomateriałach, może prowadzić do zakażeń w obrębie jamy ustnej. Celem badań była ocena in vitro wpływu prądu stałego, o niskim natężeniu na formowanie biofilmu Staphylococcus aureus na biomateriałach stomatologicznych. Materiał do badań stanowiły szczepy S. aureus izolowane od pacjentów, leczonych w Zakładzie Chirurgii Stomatologicznej Warszawskiego Uniwersytetu Medycznego. Zdolność tworzenia biofilmu oznaczono na krążkach tytanowych i cyrkonowych. Erozję i inhibicję powstałego biofilmu badano dla 3 wybranych szczepów S. aureus, z użyciem prądu stałego o natężeniu 10 mA, w trzech powtórzeniach w czasie (0 min, 10 min i 60 min). Metodą seryjnych rozcieńczeń oznaczono liczbę drobnoustrojów. Badane szczepy wykazywały zróżnicowaną zdolność tworzenia biofilmu. Erozję na cyrkonie i tytanie badano na 24-godz. biofilmie, a inhibicje na 2 godzinnym. W badaniach potwierdzono zdolność S. aureus do formowania zróż-nicowanego biofilmu na tytanie i cyrkonie – biomateriałach wykorzystywanych w stomatologii, a także hamujący wpływ prądu stały o niskim natężeniu, na tworzenie biofilmu S. aureus, szczególnie na krążkach tytanowych. Słowa kluczowe: biofilm, biomateriały, natężenie prądu, tytan, cyrkon
Effect of low direct current on Staphylococcus aureus biofilm formation on titanium and zirconium – dental biomaterials
Abstract In the course of the dental treatment, the forming of the bacterial biofilm on biomaterials can cause infection inside of mouth. The objective of this study was evaluation of the in vitro influence of the low intensity direct current (LIDC) on Staphylococcus aureus biofilm forming on the dental biomaterials. The study material was S. aureus strains isolated from patients, treated at Department of Oral Surgery Medical University of Warsaw. The ability to create the biofilm was indicated on titanium and zirconium discs. Erosion and inhibition of the biofilm was examined for 3 chosen strains of S. aureus, with using the low intensity direct current about 10 mA intensity, in three repeating time (0 min, 10 min and 60 min). The numbers of microorganisms were determined using the agar dilution method. Examined strains demonstrated the diversified ability to create the biofilm. Erosion on zirconium and titanium was being examined on 24 h biofilm and for inhibition for 2 h. Conclusions is that S. aureus is forming the diversified biofilm on titanium and zirconium – biomaterials used in dentistry. Low intensity direct current can inhibit creating of the Staphylococcus aureus biofilm after 10 min, particularly on titanium products. Keywords: biofilm, biomatrials, low direct currenty, titanium, zirconium

138
Julia Szułczyńska1
Wykorzystanie preparatów zawierających
nanocząsteczki do usuwania biofilmu bakteryjnego
1. Wstęp
Poszczególne mikroorganizmy charakteryzują się właściwościami adhezyjnymi
tworząc przy tym trwałą, rozbudowaną strukturę jaką jest biofilm, nazywany również
błoną biologiczną. Biofilm może występować zarówno na powierzchniach ożywionych
jak i abiotycznych [1]. Składa się on z licznych mikrokolonii, które są oddzielone od
siebie siecią kanalików, dzięki którym usuwane są produkty przemiany materii oraz
dostarczane są składniki pokarmowe [1]. Ponadto, mikrokolonie mogą się składać
z wielu różnych gatunków mikroorganizmów. Struktura biofilmu bakteryjnego nieus-
tannie ulega przemianom ze względu na zachodzące w nim procesy, zarówno wew-
nątrz jak i na zewnątrz [2]. Dodatkowo, drobnoustroje bytujące w błonie biologicznej
generują chemiczne cząsteczki sygnałowe, które umożliwiają komunikację między
mikroorganizmami tego samego gatunku jak i między innymi [1]. Zjawisko to, nazy-
wane quorum sensing, umożliwia mikrobom kontrolę istotnych procesów życiowych
tj. tworzenie biofilmu, bioluminescencji czy wirulencji [3].
1.1. Charakterystyka biofilmu bakteryjnego
Liczne doniesienia literaturowe [5, 6] wskazują, że proces kształtowania biofilmu
można podzielić na charakterystyczne fazy. Początkowo czysta powierzchnia
stosunkowo szybko zostaje pokryta związkami organicznymi takimi jak polisacharydy
czy białka [6]. Jest to tak zwana warstwa kondycjonująca, która umożliwia mikroorga-
nizmom możliwość adhezji do powierzchni, za pomocą sił elektrostatycznych
i oddziaływań międzycząsteczkowych (sił van der Waals’a) [6]. Kolejny etap obejmuje
coraz intensywniejsze przyczepianie się komórek z udziałem fimbrii i/lub wydzie-
lanych polimerów [6]. Następnie komórki te rozmnażają się tworząc przy tym jedno-
warstwową strukturę [6]. Powstają mikrokolonie drobnoustrojów, które syntetyzują
zewnątrzkomórkowe polimery niezbędne do podtrzymania wzrostu biofilmu [6]. Jest
to dojrzała forma biofilmu. W następstwie komórki znajdujące się w biofilmie mogą
ulec rozproszeniu. Proces powstawania biofilmu bakteryjnego został zaprezentowany
na rysunku 1. W literaturze wyróżnia się również odmienne fazy formowania się bio-
filmu bakteryjnego dla bakterii Gram-ujemnych i Gram-dodatnich. Etapy powstawania
biofilmu w zależności od budowy ściany komórkowej bakterii zostały przedstawione
w tabeli 1 [7].
1 [email protected], SKN Inventum, Katedra Przyrodniczych Podstaw Jakości, Wydział
Towaroznawstwa, Uniwersytet Ekonomiczny w Poznaniu, www.ue.poznan.pl.

Wykorzystanie preparatów zawierających nanocząsteczki do usuwania biofilmu bakteryjnego
139
Tabela 1. Etapy powstawania biofilmu bakteryjnego
Bakterie Gram-dodatnie
np. Staphylococcus aureus, Bacillus
subtilis
Bakterie Gram-ujemne
np. Pseudomonas aeruginosa,
Escherichia coli
1. Początkowa adhezja komórek 1. Początkowy kontakt komórek
z powierzchnią
2. Wewnątrzkomórkowa agregacja 2. Odwracalna adhezja
3. Dojrzewanie biofilmu 3. Nieodwracalna adhezja
4. Dyspersja biofilmu 4. Dojrzewanie i rozproszenie się
biofilmu bakteryjnego
Źródło: Opracowanie własne na podstawie [7]
Badania wskazują, że błona biologiczna złożona jest z ok. 97% wody, od 2% do 5%
z komórek mikroorganizmów, z polisacharydów – od 1% do 2% [7]. Oprócz tego w jej
skład wchodzą białka, w tym enzymy (<1%-2%), cząsteczki DNA lub RNA (<1%-
2%) oraz związane bądź wolne jony [7].
Rozpatrując aspekt formowania się biofilmu bakteryjnego kluczową rolę pełnią
możliwości przyczepności komórek do różnych powierzchni. Zdecydowanie, szorstka
i pofałdowana struktura jak i hydrofobowość powierzchni ułatwiają adhezję komórek
[2]. Również stężenie substancji odżywczych, brak środków przeciwdrobnoustro-
jowych, temperatura i prędkość przepływu wody mogą istotnie wpływać na wzrost
błony biologicznej [2]. Ponadto właściwości komórek drobnoustrojów nie pozostają
bez znaczenia. Występowanie flagelli, fimbrii, zewnątrzkomórkowych polimerów oraz
hydrofobowość powierzchni komórek przyczyniają się do zintensyfikowania
właściwości adhezyjnych mikroorganizmów [2]. Przykładowo, Pseudomonas aerugi-
nosa charakteryzuje się czterema strukturami powierzchniowymi, które sprzyjają
adhezji komórek [4, s. 326]:
rzęski,
fimbrie,
lipopolisacharyd (LPS),
alginian – śluzowy egzopolisacharyd tworzący wokół bakterii grubą ochronną
otoczkę.
Biofilm bakteryjny jest złożony głównie z komórek bakteryjnych oraz z zewnątrz-
komórkowych polimerów o dużej masie cząsteczkowej, które są syntetyzowane przez
te komórki (ang. Extracellular Polymeric Substances) [2]. Zewnątrzkomórkowe
polimery (EPS) są najczęściej zbudowane z polisacharydów [2]. Wykazują szereg
właściwości niezbędnych do funkcjonowania biofilmu bakteryjnego. Struktura zew-
nątrzkomórkowych polimerów umożliwia tworzenie licznych wiązań wodorowych,
które zapobiegają odwodnieniu biofilmu [2]. Dodatkowo, przyczyniają się do zwięk-
szenia oporności w stosunku do środków przeciwdrobnoustrojowych, poprzez utrud-
nienie masowego transportu antybiotyków [2]. Najprawdopodobniej za pomocą two-
rzenia bezpośrednich wiązań z danymi antybiotykami [2].

Julia Szułczyńska
140
Komórki bakterii bytujące w głębszych warstwach błony biologicznej przeksz-
tałcają swoje czynności życiowe, ze względu na mniejszą ilość substratów pokarmo-
wych czy tlenu [1]. Skutkuje to zmniejszeniem tempa wzrostu oraz przechodzeniem
w stan podobny do anabiozy [1]. Taka forma bytowania komórek powoduje większą
odporność na działanie środków przeciwdrobnoustrojowych [1]. Należy dodać, że
bakterie znajdujące się w różnych warstwach biofilmu charakteryzują się różną
podatnością na działanie antybiotyków czy środków dezynfekujących [1]. Spowol-
nienie metabolizmu komórek znajdujących się w dojrzałym biofilmie bakteryjnym
przyczynia się do nieskutecznego działania antybiotyków [1], ponieważ funkcjo-
nowanie antybiotyków polega na przekształceniu szlaków metabolicznych komórki
bakteryjnej [11]. Ponadto, długookresowe działanie antybiotyków na błonę biolo-
giczną, może przyczyniać się do wystąpienia punktowych mutacji genów, które zwięk-
szają oporność mikroorganizmów [1].
Charakterystyczną cechą komórek znajdujących się w biofilmie jest ich oporność
na liczne środki dezynfekujące, surfaktanty czy antybiotyki. Mikroorganizmy te mogą
być nawet 1000 razy bardziej oporne niż komórki znajdujące się w zawiesinie [1].
Błony biologiczne mogą tworzyć się na różnych powierzchniach abiotycznych tj.
szklanych, drewnianych, metalowych czy z tworzywa sztucznego. Mogą również
formować się w naturalnych systemach wodnych lub występować w instalacjach
wodno-kanalizacyjnych, czy nawet na sprzętach medycznych tj. cewniki, zastawki
sercowe, rozruszniki lub protezy [1, 7, 14]. Redukcja biofilmu bakteryjnego jest klu-
czowa z punktu widzenia bezpieczeństwa produkcji m.in. w przemyśle spożywczym,
gdzie kontakt żywności z biofilmem może przyczynić się do skażenia produktu i być
czynnikiem chorobotwórczym. Jednym ze sposobów usuwania biofilmu bakteryjnego
jest stosowanie preparatów zawierających nanocząsteczki, które są tematem zain-
teresowanie niniejszej pracy.
1.2. Nanocząsteczki
Nanocząsteczka jest to fragment materii, której przynajmniej jeden z trzech wymia-
rów przestrzennych jest mniejszy niż 100 nm [8, s. 766]. Ze względu na swój niewielki
rozmiar, nanocząsteczki mogą zostać wchłonięte przez komórki organizmów żywych,
powodując przy tym m.in. zaburzenia w proliferacji komórek czy blokowanie kanałów
jonowych [8]. Interesujące właściwości fizyko-chemiczne nanocząsteczek srebra są
sprzężone z dużą powierzchnią aktywną cząsteczki [9].
Nanocząsteczki srebra wykazują silnie działanie przeciwdrobnoustrojowe. Bak-
teriobójcze właściwości nanosrebra skutkują wzrostem przepuszczalności błony ko-
mórkowej bakterii, szkodliwym działaniem na DNA oraz zaburzeniami w funkcjo-
nowaniu kanałów jonowych komórki [9]. Oddziaływanie nanosrebra na komórki
mikroorganizmów sprawia, że produkty przemiany materii kumulują się wewnątrz
komórki. Innym działaniem jest uniemożliwienie syntezy białek [9]. Liczne donie-
sienia naukowe świadczą o właściwościach nanosrebra do hamowania bądź usuwania
biofilmu bakteryjnego wytworzonego przez Pseudomonas aeruginosa, Staphylococcus

Wykorzystanie preparatów zawierających nanocząsteczki do usuwania biofilmu bakteryjnego
141
epidermidis czy Streptococcus mutans, który poprzez skomplikowaną strukturę jest
trudny do usunięcia [9, 10].
Miedź posiada silne właściwości biobójcze w stosunku do licznych gatunków
bakterii czy grzybów strzępkowych. Udokumentowano wiele badań naukowych
dotyczących właściwości przeciwdrobnoustrojowych nanocząsteczek miedzi
w szerokim spektrum działania tj. K. pneumonie, E. coli, P. aeruginosa czy M. luteus
[13]. Dowiedziono też, że w przypadku A. niger, A. flavus, i C. albicans nanomiedź
posiada właściwości fungistatyczne, aczkolwiek zahamowanie wzrostu jest mniejsze
niż w przypadku bakterii [13].
Nanocząsteczki miedzi i srebra mogą być swoistą alternatywą w stosunku do
tradycyjnych metod hamowania rozwoju mikroorganizmów i usuwania biofilmu
bakteryjnego, ze względu na szerokie spektrum działania na bakterie i grzyby.
2. Metodyka
2.1. Przygotowanie mikroorganizmów
W artykule wykorzystano zarówno bakterie Gram-dodatnie – Staphylococcus
aureus i Bacillus subtilis, jak i Gram-ujemne – Pseudomonas aeruginosa oraz
Escherichia coli. Hodowla P. aeruginosa i S. aureus była prowadzona na bulionie
odżywczym. Natomiast, hodowla B. subtilis i E. coli na płynnym podłożu Tripticasein
Soy Broth (TSB). Do badań wykorzystano hodowle o gęstości optycznej równej 3
w skali McFarland’a.
2.2. Preparaty z nanocząsteczkami Cu i Ag
W badaniu wykorzystano preparaty handlowe z nanocząsteczkami Cu i Ag
o stężeniu wyjściowym 2000 ppm. Badane roztwory rozcieńczono na płytkach titracyj-
nych, także zakres stężeń plasował się od 2000 ppm do 0,06 ppm. Rozcieńczenia te
wyglądały następująco: 2000 ppm, 1000 ppm, 500 ppm, 250 ppm, 125 ppm, 62,5 ppm,
31,25 ppm, 15,63 ppm, 8 ppm, 4 ppm, 2 ppm, 1 ppm, 0,5 ppm, 0,25 ppm, 0,13 ppm
i 0,06 ppm. W próbach kontrolnych wykorzystano sterylną wodę.
2.3. Ocena stopnia usuwania biofilmu przez preparaty z nanocząsteczkami
Ocenę właściwości antybiofilmowych preparatów zawierających nanocząsteczki
miedzi i srebra wykonano z wykorzystaniem 96-dołkowych jałowych mikropłytek
z płaskim dnem. Celem przeprowadzonych badań była ocena stopnia usunięcia
biofilmu bakteryjnego wytworzonego przez bakterie Gram-ujemne i Gram-dodatnie tj.
E. coli, P. aeruginosa, S. aureus i B. subtilis. Przebieg badania prezentował się nastę-
pująco.
W pierwszej kolejności zaaplikowano po 50 µl pożywki odpowiedniej dla danych
bakterii (tj. bulion odżywczy dla P. aeruginosa i S. aureus oraz TSB dla E. coli i B.
subtilis). Następnie, naniesiono po 50 µl zawiesiny mikroorganizmów. Mikroorga-
nizmy zostały wprowadzone do pożywki za pomocą pipety wielokanałowej. Płytki
titracyjne inkubowano w 37°C przez okres 72 godzin. Kolejnym krokiem było prze-
mycie sterylną wodą każdego dołka, w celu usunięcia komórek bakterii, które nie

Julia Szułczyńska
142
wykazały właściwości adhezyjnych do powierzchni. Mikropłytki z wytworzonym
biofilmem suszono w 60°C przez 45 minut. Następnie zaaplikowano 100 µl kolejnych,
dwukrotnych rozcieńczeń preparatów z nanocząsteczkami miedzi i srebra. Rozcień-
czenia przygotowano na równoległej płytce tak, aby zakres stężeń wynosił od 2000
ppm do 0,063 ppm. Po 15 minutach preparaty usunięto a płytki przepłukano sterylną
wodą usuwając z nich pozostałość preparatów. Ponownie pozostawiono je w 60°C
przez okres 45 minut. Dalszy etap polegał na barwieniu komórek przyczepionych do
powierzchni płytek za pomocą roztworu fioletu krystalicznego, który wprowadzano
w ilości 100 µl i pozostawiano przez 15 minut. Po usunięciu barwnika, płytki titracyjne
suszono w 60°C przez 30 minut. Następnie, każdą studzienkę przemyto i wprowa-
dzono 125 µl roztworu alkoholu etylowego. Ostatnim etapem był pomiar spektrofoto-
metryczny przy długości fali 600 nm. Schemat mikropłytki przedstawiono na rysunku 2.
Rysunek 1. Schemat 96-dołkowej płytki titracyjnej z preparatem nanosrebra i nanomiedzi
Źródło: opracowanie własne
3. Analiza wyników
Preparat zawierający nanocząsteczki Cu wykazywał się największą zdolnością
redukcyjną w przypadku wszystkich badanych bakterii. Preparat ten w największym
stopniu usuwał biofilm dla bakterii B. subtilis i E. coli. W przypadku bakterii B.
subtilis zaobserwowano skuteczne usuwanie powstałego biofilmu w całym zakresie
stężeń, a skuteczność wynosiła od 80,99% do 63,31%. Natomiast, w przypadku bak-
terii E. coli skuteczne usuwanie biofilmu przez preparat z nanomiedzią głównie
w zakresie wyższych stężeń tj. od 500 ppm do 2000 ppm, chociaż proces zachodził
efektywnie również w niższych stężeniach, od 0,25 ppm do 8 ppm. W przypadku S.

Wykorzystanie preparatów zawierających nanocząsteczki do usuwania biofilmu bakteryjnego
143
aureus preparat z nanocząsteczkami miedzi stosunkowo intensywnie redukował
powstałą błonę biologiczną w niskich stężeniach, od 2 do 0,06 ppm, chociaż silne
usuwanie biofilmu było widoczne również w stężeniu stężeń: od 1000 do 2000 ppm.
W odniesieniu do P. aeruginosa badane preparaty redukowały błonę biologiczną
w najmniejszym stopniu (maksymalny stopień redukcji wyniósł = 4,92% dla Cu).
Aczkolwiek, nanopreparat zawierający miedź wykazywał się większą skutecznością.
Nanopreparat zawierający srebro w największym stopniu oddziaływał na redukcję
biofilmu w stosunku do B. subtilis w całym zakresie stężeń. Największą redukcję
biofilmu bakteryjnego nanosrebro wykazywało w zakresie stężeń od 2000 ppm do 500
ppm (od 80,74% do 80,50%). Najsłabszą aktywność preparat przejawiał przy dwóch
najniższych badanych stężeniach – redukcja biofilmu wyniosła ok. 67%). Działanie
redukcyjne nanocząsteczek srebra w przypadku biofilmu wytworzonego przez E. coli
było relatywnie umiarkowane i wahało się od 29,92% do 57,87%. Jednakże, w zak-
resie stężeń od 4 ppm do 0,5 ppm nanocząsteczki srebra skuteczniej usuwały błonę
biologiczną niż nanocząsteczki miedzi. Optymalne stężenie nanosrebra w przypadku
bakterii E.coli w przypadku badanego preparatu wynosiło 1 ppm.
Najmniejszą skuteczność nanopreparat srebra wykazał w stosunku do biofilmu
bakteryjnego wytworzonego przez S. aureus i P. aeruginosa. W tych dwóch przypad-
kach działał on dopiero od pewnego zakresu stężeń (dla S. aureus od 8 ppm, a dla P.
aeruginosa od 250 ppm). W wyższych stężeniach preparatu nie zaobserwowano
działania redukującego.
Należy zauważyć, że P. aeruginosa wytworzył najbardziej trwały i oporny biofilmu
bakteryjny spośród czterech badanych mikroorganizmów. Jest to konsekwencją posia-
dania aż czterech struktur zewnątrzkomórkowych sprzyjających adhezji komórek do
powierzchni m.in. fimbrie, alginian. Maksymalny procent usunięcia biofilmu przez
nanocząsteczki wynosił zaledwie 4,92%. Natomiast, w przeciwieństwie do P. aeru-
ginosa, B. subtilis uformował najmniej trwałą błonę biologiczną. Była ona stosunkowo
łatwo usuwana, na poziomie ok. 70-80%, przez oba badane preparaty. Jak również
przez wodę, która redukowała go w 68,27%.
Jako wariant kontrolny zastosowano sterylną wodę do usuwania biofilmu bakte-
ryjnego. W zależności od badanego mikroorganizmu woda redukowała błonę biolo-
giczną na następującym poziomie; dla P. aeruginosa w wysokości 0,61%, dla E.coli
w 36,95%, dla S. aureus w 2,34% oraz dla B. subtilis w 68,27%.
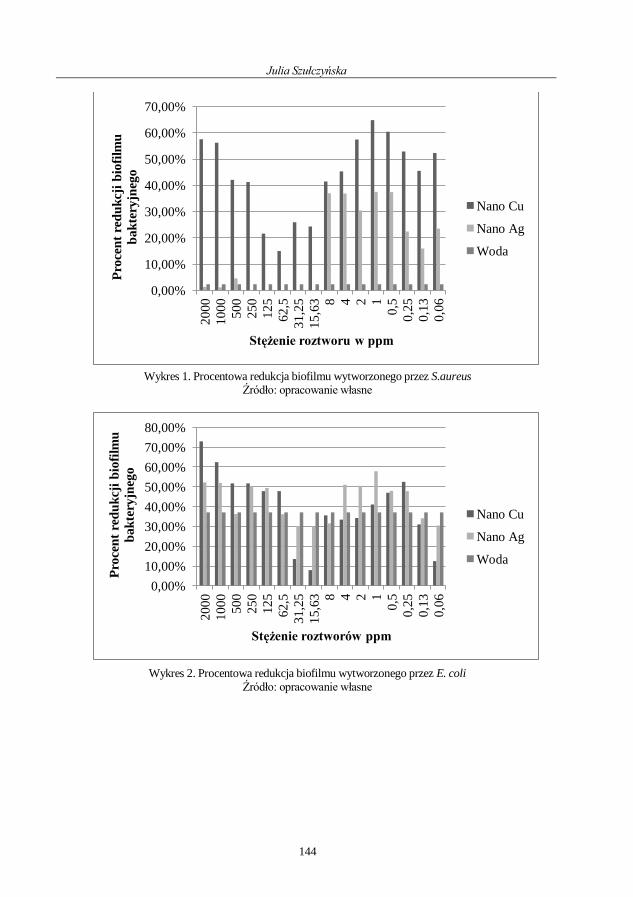
Julia Szułczyńska
144
Wykres 1. Procentowa redukcja biofilmu wytworzonego przez S.aureus
Źródło: opracowanie własne
Wykres 2. Procentowa redukcja biofilmu wytworzonego przez E. coli
Źródło: opracowanie własne
0,00%
10,00%
20,00%
30,00%
40,00%
50,00%
60,00%
70,00%
2000
1000
500
250
125
62,5
31,2
5
15,6
3
8
4
2
1
0,5
0,2
5
0,1
3
0,0
6
Pro
cen
t re
du
kcj
i b
iofi
lmu
bak
tery
jneg
o
Stężenie roztworu w ppm
Nano Cu
Nano Ag
Woda
0,00%
10,00%
20,00%
30,00%
40,00%
50,00%
60,00%
70,00%
80,00%
2000
1000
500
250
125
62,5
31,2
5
15,6
3
8
4
2
1
0,5
0,2
5
0,1
3
0,0
6
Pro
cen
t re
du
kcj
i b
iofi
lmu
bak
tery
jneg
o
Stężenie roztworów ppm
Nano Cu
Nano Ag
Woda
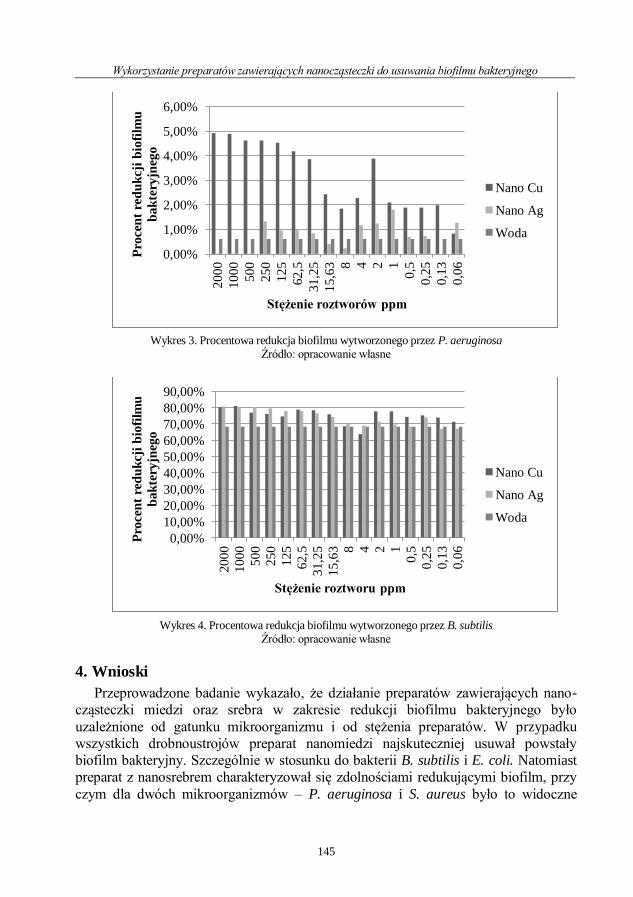
Wykorzystanie preparatów zawierających nanocząsteczki do usuwania biofilmu bakteryjnego
145
Wykres 3. Procentowa redukcja biofilmu wytworzonego przez P. aeruginosa
Źródło: opracowanie własne
Wykres 4. Procentowa redukcja biofilmu wytworzonego przez B. subtilis
Źródło: opracowanie własne
4. Wnioski
Przeprowadzone badanie wykazało, że działanie preparatów zawierających nano-
cząsteczki miedzi oraz srebra w zakresie redukcji biofilmu bakteryjnego było
uzależnione od gatunku mikroorganizmu i od stężenia preparatów. W przypadku
wszystkich drobnoustrojów preparat nanomiedzi najskuteczniej usuwał powstały
biofilm bakteryjny. Szczególnie w stosunku do bakterii B. subtilis i E. coli. Natomiast
preparat z nanosrebrem charakteryzował się zdolnościami redukującymi biofilm, przy
czym dla dwóch mikroorganizmów – P. aeruginosa i S. aureus było to widoczne
0,00%
1,00%
2,00%
3,00%
4,00%
5,00%
6,00%
2000
1000
500
250
125
62,5
31,2
5
15,6
3
8
4
2
1
0,5
0,2
5
0,1
3
0,0
6 P
roce
nt
red
uk
cji
bio
film
u
bak
tery
jneg
o
Stężenie roztworów ppm
Nano Cu
Nano Ag
Woda
0,00%
10,00%
20,00%
30,00%
40,00%
50,00%
60,00%
70,00%
80,00%
90,00%
2000
1000
500
250
125
62,5
31,2
5
15,6
3
8
4
2
1
0,5
0,2
5
0,1
3
0,0
6 P
roce
nt
red
uk
cji
bio
film
u
bak
tery
jneg
o
Stężenie roztworu ppm
Nano Cu
Nano Ag
Woda

Julia Szułczyńska
146
dopiero od pewnego zakresu stężeń. Początkowe, wysokie stężenia nanosrebra nie
demonstrowały właściwości redukujących.
Najsilniejsze działanie antybiofilmowe odnotowano dla nanopreparatu miedzi
w stosunku do B. subtilis i wynosiło ono 80,40%. Brak jakiegokolwiek działania elimi-
nującego biofilm bakteryjny stwierdzono dla preparatu zawierającego nanosrebro
w przypadku P. aeruginosa i S. aureus w początkowych zakresach stężeń.
Ponadto, błona biologiczna wytworzona przez B. subtilis cechowała się łatwą
redukcją zarówno przez oba badane nanopreparaty, jak i przez wodę. Natomiast,
biofilm bakteryjny ukształtowany przez P. aeruginosa był najbardziej oporny i niewra-
żliwy na działanie zastosowanych preparatów nanosrebra i nanomiedzi.
Podsumowując, badane handlowe preparaty zawierające nanocząsteczki miedzi
i srebra różnią się właściwościami redukującymi biofilmu bakteryjny. Jednakże, mogą
być alternatywą dla konwencjonalnych środków stosowanych do usuwania błon
biologicznych.
Literatura
1. Czaczyk K., Myszka K., Mechanizmy warunkujące oporność biofilmów bakteryjnych na
czynniki antymikrobiologiczne, Biotechnologia, 76, (2007), s. 40-52.
2. Donlan R. M., Biofilms: Microbial Life on Surfaces, Emerging Infectious Diseases, 8,
(2002), s. 881-890.
3. Matejczyk M., Suchowierska M., Charakterystyka zjawiska quorum sensing i jego
znaczenie w aspekcie formowania i funkcjonowania biofilmu w inżynierii środowiska,
budownictwie, medycynie oraz gospodarstwie domowym, Budownictwo i Inżynieria
Środowiskowa, 2, (2011), s. 71-75.
4. Murray P. R., Rosenthal K. S., Pfaller M. A., Mikrobiologia, Wydawnictwo Elsevier.
5. Sauer K., Camper A. K., Ehrlich G. D., Costerton J. W., Davies D. G., Pseudomonas
aeruginosa displays multiple phenotypes during development as a biofilm, Journal of
Bacteriology, 184, s. 1140-1154.
6. Wimpenny J., Structural determinants in biofilm formation, (2005), s. 35-51, [w:] Evans L.
V., Biofilms: Recent Advances in their Study and Control, (2005), wyd. Taylor & Francis,
Holandia.
7. Vasudevan R., Biofilms: Microbial Cities of Scientific Significance, Journal of
Microbiology & Experimentation, 1 (3), (2014), s. 1-16.
8. Runowski M., Nanotechnologia – nanomateriały, nanocząstki i wielofunkcyjne
nanostruktury typu rdzeń/powłoka, Chemik, 68, (2014).
9. Pokrowiecki R., Mielczarek A., Wybrane przykłady zastosowania nano-cząsteczek srebra
w procedurach medycznych, Nowa Stomatologia, 3, (2012), s. 117 - 121.
10. Kalishwaralal K., Barathmanikant S., Pandian S. R. K., Deepak V., Gurunathan S., Silver
nanoparticles impede the biofilm formation by Pseudomonas aeruginosa and Staphylococcus
epidermidis, Colloid and Surfaces B: Biointerfaces, 79 (2), (2010), 340-344.
11. Dzierżanowska D., Antybiotykoterapia praktyczna, Wyd. α-medica Press, (2000), s. 73-96,
Bielsko-Biała.
12. DeAlba-Montero I., Guajardo-Pacheco J., Moralez-Sanchez E., Araujo-Martinez R.,
Loredo-Beccera G. M., Martinez-Castanon G. A., Ruiz F., Compean Jasso M. E.,
Antimicrobial Properties of Copper Nanoparticles and Amino Acid Chelated Copper

Wykorzystanie preparatów zawierających nanocząsteczki do usuwania biofilmu bakteryjnego
147
Nanoparticles Produced by Using a Soya Extract, Bioinorganic Chemistry and
Applications, (2017), s. 1-6.
13. Ramyadevi J., Jeyasubramanian K., Marikani A., Rajakumar G., Rahuman A. A., Synthesis
of antimicrobial activity of copper nanoparticles, Materials Letters, 71, (2012), s. 114-116. 14. Stoodley-Hall L., Costerton J. W., Stoodley P., Bacterial Biofilms: from the Natural
Environment to Infectious Diseases, Nature Reviews: Microbiology, 2, (2004), s. 95-108.
Wykorzystanie preparatów zawierających nanocząsteczki
do usuwania biofilmu bakteryjnego
Streszczenie Celem przeprowadzonych badań była ocena stopnia usunięcia biofilmu bakteryjnego przez dwa handlowe
preparaty zawierające nanocząsteczki Cu i Ag o stężeniu 2000 ppm. W opracowaniu posłużono się
zarówno bakteriami gram-dodatnimi i gram-ujemnymi tj. Pseudomonas aeruginosa, Escherichia coli,
Staphylococcus aureus i Bacillus subtilis. Hodowle mikroorganizmów P. aeruginosa oraz S. aureus prowadzono na bulionie odżywczym, natomiast
mikroorganizmy E. coli i B. subtilis były hodowane na płynnym podłożu TSB (Tripticasein Soy Broth).
Wszystkie bakterie inkubowano na 96-dołkowych płytkach titracyjnych z płaskim dnem przez 72 godziny
w 37°C. Powstały biofilm bakteryjny przepłukano preparatami handlowymi z nanocząsteczkami Cu i Ag w różnym zakresie stężeń (od 2000 ppm do 0,063 ppm). Następnie, mikropłytki barwiono roztworem
fioletu krystalicznego w celu umożliwienia pomiaru absorbancji. Do oceny usunięcia biofilmu bakte-
ryjnego zastosowano pomiar spektrofotometryczny przy długości fali 600 nm.
Oba badane preparaty handlowe zawierające nanocząsteczki usuwały powstały biofilm bakteryjny. Stopień usunięcia biofilmu był uzależniony od rodzaju użytego preparatu oraz od jego stężenia. Ponadto,
w przypadku różnych mikroorganizmów stopień usunięcia biofilmu był różny.
Słowa kluczowe: biofilm bakteryjny, nanopreparaty, nanosrebro, nanomiedź, Pseudomonas aeruginosa,
Bacillus subtilis, Escherichia coli, Staphylococcus aureus
The use of nanoparticle preparations in remove of bacterial biofilm
Abstract The purpose of this study was to evaluate the degree of bacterial biofilm removal by commercial prepa-
rations containing nanoparticle of Cu and Ag at the concentration of 2000 ppm. Both gram-positive and
gram-negative bacteria were used in the paper. The following microorganisms were tested: Pseudomonas
aeruginosa, Escherichia coli, Staphylococcus aureus and Bacillus subtilis. P. aeruginosa and S. aureus were cultured on nutrient broth, whereas E. coli and B. subtilis were grown on
TSB medium (Tripticasein Soy Broth). All bacteria were incubated in 96-well flat bottomed titration plates
for 72 hours at 37°C. The formed bacterial biofilm was rinsed was rinsed Cu and Ag nanopreparations at
different range of concentrations (from 2000 ppm to 0,063ppm). Subsequently the microplates were stained with a crystalline violet solution to allow absorbance measure. Spectrophotometric measurement at
wavelength of 600 nm were used to assess the removal of bacterial biofilm.
Both tested nanopreparations removed bacterial biofilm. However the degree of biofilm removal dependent
on the type of nanopreparations and on its range of concentrations. In addition, the level of biofilm removal was various for different microorganisms.
Keywords: bacterial biofilm, nanopreparations, nano-zinc, nano-copper, Pseudomonas aeruginosa,
Bacillus subtilis, Escherichia coli, Staphylococcus aureus

148
Aldona Gajek1, Joanna Kwiecińska-Piróg
2, Eugenia Gospodarek-Komkowska
2
Ocena wytwarzania biofilmu przez szczepy Proteus
mirabilis izolowane z ran przewlekłych
1. Wstęp
1.1. Charakterystyka Proteus mirabilis
Pałeczki Proteus mirabilis są jednym z czterech gatunków rodzaju Proteus. Zostały
opisane po raz pierwszy przez Gustawa Hausera w 1885 roku [1]. Początkowo rodzaj
Proteus stanowiły dwa gatunki: P. mirabilis i P. vulgaris. Obecnie w obrębie rodzaju
wyróżnia się cztery gatunki: P. mirabilis, P. vulgaris, P. penneri i P. hauseri oraz trzy,
nienazwane gatunki genomowe [1].
Pałeczki Proteus spp. należą do oportunistycznych patogenów człowieka. P.
mirabilis jest trzecim w kolejności patogenem (po Escherichia coli i Klebsiella pniu-
moniae) izolowanym z zakażeń układu moczowego. Są to przede wszystkim zakażenia
u pacjentów z wadami anatomicznymi w obrębie układu moczowego, jak również u
pacjentów cewnikowanych [2]. Drobnoustroje te mogą także wywoływać zakażenia
ran, szczególnie o przewlekłym charakterze, a także zakażenia kości, sepsę czy zapa-
lenie opon mózgowo-rdzeniowych [1, 3].
Najczęściej izolowanym gatunkiem tego rodzaju z zakażeń człowieka jest P.
mirabilis. Chorobotwórczość pałeczek gatunku jest związana z obecnością licznych
czynników wirulencji. Należą do nich: zdolność do przylegania do powierzchni
(uwarunkowana głównie występowaniem fimbrii lub afimbrialnych adhezyn), inwa-
zyjność, zjawisko rozpełzłego (mgławicowego) wzrostu, aktywność hemolityczna,
zdolność do rozkładu mocznika, właściwości proteolityczne oraz wytwarzanie endo-
toksyny (lipopolisacharydu, LPS) [1, 3].
Czynniki wirulencji pozwalają bakteriom z rodzaju Proteus skutecznie skolo-
nizować wybraną tkankę lub powierzchnię biomateriału, co może zapoczątkować
tworzenie biofilmu [1].
Biofilm stanowią wzajemnie komunikujące się drobnoustroje osiadłe na określo-
nym podłożu. Przylegają one do siebie i są osłonięte warstwą egzopolisacharydu.
Biofilm może być utworzony przez jeden lub wiele gatunków drobnoustrojów [1].
Tworzenie biofilmu jest procesem kilkuetapowym, który rozpoczyna wiązanie
komórek drobnoustrojów z powierzchnią kolonizowaną. Początkowo adhezja jest
odwracalna i polega na słabych oddziaływaniach o charakterze hydrofobowym. Po
adhezji odwracalnej dochodzi do nieodwalanego związania komórek ze stałą po-
1 Studenckie Koło Naukowe przy Katedrze i Zakładzie Mikrobiologii Collegium Medicum im. Ludwika Rydygiera w Bydgoszczy, Uniwersytet Mikołaja Kopernika w Toruniu, ul. Skłodowskiej-Curie 9, 85-094
Bydgoszcz 2 Katedra i Zakład Mikrobiologii Collegium Medicum im. Ludwika Rydygiera w Bydgoszczy, Uniwersytet
Mikołaja Kopernika w Toruniu, ul. Skłodowskiej-Curie 9, 85-094 Bydgoszcz

Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis izolowane z ran przewlekłych
149
wierzchnią. Komórki drobnoustrojów ulegają zmianom fenotypowym związanym
z aktywacją i wygaszaniem genów. Dochodzi do wytworzenia warstwy egzopolisa-
charydowej i wykształcenia wielu mikrokolonii – podstawowych jednostek struktural-
nych biofilmu. Między mikrokoloniami tworzą się kanały, umożliwiające przepływ
wody, substancji odżywczych, produktów metabolizmu oraz płynów ustrojowych [1, 4].
Kolejny etap stanowi tworzenie się dojrzałego biofilmu. Jest on złożony z kilku
warstw: wewnętrznej, stykającej się z powierzchnią stałą; warstwy podstawowej,
w której komórki przylegają do siebie; oraz warstwy powierzchniowej, charakte-
ryzująca się luźnym ułożeniem komórek drobnoustrojów. Dzięki dostępowi do tlenu
i substancji odżywczych, komórki warstwy powierzchniowej charakteryzują się szyb-
kim metabolizmem, jednak są najbardziej narażone na działanie układu immunolo-
gicznego gospodarza czy też stosowanych środków bakteriobójczych lub bakterio-
statycznych. Komórki warstwy powierzchniowej izolują komórki wewnętrznych
warstw biofilmu przed wpływem związków przeciwdrobnoustrojowych, jednak mają
ograniczony dostęp do związków odżywczych. Konsekwencją tego jest zwolnienie
tempa metabolizmu, a nawet przejście w stan „uśpienia” [1, 4].
Poszczególne komórki drobnoustrojów w biofilmie mogą wysyłać i odbierać
cząsteczki sygnałowe, dzięki czemu możliwa jest komunikacja pomiędzy drobnoustro-
jami oraz reagowanie na zagęszczenie populacji. Zjawisko to określane jest mianem
quorum sensing. Efektem quorum sensing jest skoordynowana reakcja populacji
komórek biofilmu, prowadząca m.in. do rozpoczęcia procesu przekształcania się form
osiadłych w planktoniczne oraz odrywania się ich od biofilmu i przemieszczania
w inne miejsca, co jest określane mianem dyspersji biofilmu [1, 4].
Zdolność tworzenia biofilmu sprzyja rozwojowi i utrzymywaniu się zakażeń,
szczególnie związanych z wykorzystaniem materiałów sztucznych, takich jak cewniki
naczyniowe i do pęcherza moczowego, stenty moczowodowe lub sterczowe, implanty
penisa, implanty jąder, zastawki serca, protezy tchawicy [2].
Przypuszcza się, że trudności terapeutyczne leczenia zakażeń o etiologii P. mira-
bilis mogą być związane z obecnością biofilmu. Biofilm stanowi dla bakterii ochronę
przed układem immunologicznym gospodarza, np. poprzez utrudnianie fagocytozy,
zaburzanie, zahamowanie opsonizacji oraz zmniejszenie penetracji przeciwciał, a także
leków przeciwdrobnoustrojowych [2, 5, 6].
1.2. Rany przewlekłe
Rany przewlekłe, włączając cukrzycowe owrzodzenia stóp oraz żylne owrzodzenia
stóp, stanowią ogólnoświatowy problem zdrowotny. Wysoce prawdopodobne jest, iż
drobnoustroje kolonizujące rany przewlekłe wytwarzają biofilm [7].
Raną przewlekłą określa się ranę, której proces gojenia nie przebiega prawidłowo
i nie kończy się w ciągu trzech miesięcy od jej pojawienia się, pomimo stosowania
odpowiedniego leczenia [8]. Rany o charakterze przewlekłym mogą utrzymywać się
do kilku lat [8].
Zakażenie rany przewlekłej można podejrzewać, gdy po jej właściwym oczysz-
czeniu przez okres powyżej dwóch tygodni rana nie goi się, występuje słabe ziarni-

Aldona Gajek, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
150
nowanie, dochodzi do zmian zabarwienia tkanki, nasilenia dolegliwości bólowych,
pojawienia się cuchnącego zapachu, narastania obszarów martwicy, wzmożonego
drenażu wydzieliny, a także wzrostu ocieplenia tkanek dookoła rany. Najbardziej
specyficznymi z wymienionych objawów, świadczącymi o zakażeniu są: pojawienie
się pęknięć w obrębie ziarniny oraz nasilenie dolegliwości bólowych [9, 10].
Kliniczne rozpoznanie zakażenia rany przewlekłej następuje po stwierdzeniu co
najmniej czterech z wymienionych objawów: pogorszenie stanu psychicznego pacjenta,
gorączka (≥38°C), zaczerwienienie lub wzmożona ciepłota skóry w pobliżu rany, obrzęk,
drenaż surowiczy, zlokalizowany ból lub tkliwość. Kryteria te zostały określone dla
zaawansowanych wiekiem osób przebywających w domach opieki [9, 11].
Rany przewlekłe stanowią rezerwuar lekoopornych drobnoustrojów ze względu na
kolonizację bakteryjną oraz częstą antybiotykoterapię tych zakażeń [9, 12]. W około
4% ogólnej liczby zakażeń ran wykazano obecność pałeczek z rodzaju Proteus [1].
2. Cel pracy
Pałeczki P. mirabilis są izolowane z zakażeń ran przewlekłych, w których powsta-
waniu, jak się przypuszcza, duże znaczenie odgrywa obecność drobnoustrojów zdol-
nych do tworzenia biofilmu. Z tego powodu celem niniejszej pracy była ocena wytwa-
rzania struktury biofilmu przez szczepy P. mirabilis izolowane z zakażeń ran o przew-
lekłym charakterze.
3. Metodyka
3.1. Materiał
W badaniach wykorzystano 36 szczepów P. mirabilis wyosobnionych od pacjentów
Szpitala Uniwersyteckiego nr 1 im. dra A. Jurasza w Bydgoszczy leczonych w 2016
roku. Szczepy były identyfikowane do gatunku za pomocą spektrometrii mas Maldi-
Tof (Microflex; Bruker). Na podstawie profili białkowych badanych szczepów wykreś-
lono w programie MBT Explorer (Bruker) dendrogram podobieństwa szczepów.
W badaniach intensywności tworzenia biofilmu użyto dwóch szczepów wzorco-
wych z Amerykańskiej Kolekcji Szczepów Wzorcowych (American Type Culture
Collection, ATCC): E. coli ATCC 35218, stanowiącego kontrolę ujemną (szczep słabo
wytwarzający biofilm) oraz P. mirabilis ATCC 29906 jako kontrolę dodatnią (izolat
tworzący biofilm). Do czasu badań szczepy przechowywano w bulionie mózgowo-
sercowym (BHI, Becton Dickinson) z 20% zawartością glicerolu (Avator) w tem-
peraturze -70°C.
3.2. Ocena lekowrażliwości i wytwarzania beta-laktamaz o rozszerzonym
spektrum substratowym
Lekowrażliwość badanych szczepów na amoksycylinę z kwasem klawulanowym
(AMC), piperacyline z tazobaktamem (TZP), cefuroksym (CXM), cefotaksym (CTX),
ceftazydym (CAZ), cefepim (FEP), imipenem (IMP), meropenem (MEM), genta-
mycynę (GEN), amikacynę (AMK), tobramycynę (TOB), ciprofloksacynę (CIP)
i trimetoprim/sulfametoksazol (SXT) oceniano z wykorzystaniem kart AST-N332

Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis izolowane z ran przewlekłych
151
i systemu VITEK2 Compact. Wyniki interpretowano zgodnie z aktualnymi rekomen-
dacjami EUCAST (European Committee on Antimicrobial Susceptibility Testing) [13].
Występowanie beta-laktamaz o rozszerzonym spektrum substratowym (extended
spectrum beta-lactamses, ESBL) badano zgodnie z zaleceniami KORLD (Krajowego
Ośrodka ds. Oceny Lekowrażliwości Drobnoustrojów) metodą synergizmu dwóch
krążków (double disk synergic test, DDST), stosując krążki z amoksycyliną i kwasem
klawulanowym, ceftazydymem, cefepimem i cefotaksymem (BioMaxima) [14].
3.3. Ocena wytwarzania biofilmu
Szczepy po rozmrożeniu namnażano przez 24 godziny w temperaturze 37°C na
podłożu agarowo-tryptozowym (TSA; Becton Dickinson). Z pojedynczych kolonii
wykonywano zawiesinę w bulionie tryptozowo-sojowym (TSB; Becton Dickinson),
następnie hodowlę inkubowano w temperaturze 37°C. Po dobie inkubacji wyko-
nywano zawiesinę w 0,85% roztworze NaCl o gęstości 0,5 w skali MacFarlanda.
Zawiesiną badanych szczepów uzupełniano studzienki 96-dołkowych płytek polisty-
renowych (Profilab) w trzech powtórzeniach dla każdego szczepu (po 4 μl/studzienkę).
Równolegle z wykorzystaniem tych samych zawiesin badanych szczepów
przygotowywano dwie płytki (A, B). Następnie dołki uzupełniono do objętości 200 μl
jałowym TSB i inkubowano przez 24 godziny w temperaturze 37°C. Wielkość two-
rzonego biofilmu oceniano w dołkach płytki A metodą kolorymetryczną z wykorzys-
taniem fioletu krystalicznego (Cristal Violet, CV; Avantor) przy długości fali 570 nm.
W dołkach płytki B badano aktywność metaboliczną biofilmu poprzez pomiar
absorbancji (λ=470 nm) formazanu (produktu metabolizmu chlorku 2,3,5-trifeny-
lotetrazolu, TTC; Avantor).
Dalsze procedury dotyczące barwienia fioletem krystalicznym oraz formazanem
były przeprowadzone zgodnie z metodyką Kwiecińska-Piróg i wsp. [15]. Na podstawie
sumy średniej (s) i trzech odchyleń standardowych (SD) wartości absorbancji próby tła
ustalono wartość (X), powyżej której szczepy uznawano za tworzące biofilm.
Wielokrotności wartości X stanowiły granice przedziałów, pomiędzy szczepami słabo,
średnio i silnie tworzącymi biofilm.
Tab. 1.Wartości graniczne absorbancji
Stopień tworzenia biofilmu Wartości graniczne absorbancji
TTC CV
Brak <0,053(X) <0,074 (X)
Słaby ≥0,053(X) ˄ ≤0,212 (5X) ≥0,074 (X) ˄ ≤0,148 (2X)
Średni >0,212 (5X) ˄ ≤0,424 (10X) >0,148 (2X) ˄ ≤0,222 (3X)
Silny >0,424 (10X) >0,222 (3X)
Objaśnienia: TTC – wyniki uzyskane metodą pomiaru absorbancji formazanu, CV – wyniki uzyskane metodą
pomiaru absorbancji fioletu krystalicznego
Wyniki absorbancji zostały zapisane w programie KC4 (BioTek), a następnie zeb-
rane w programie Microsoft Office Excel (2013). Opracowanie statystycznie wyników
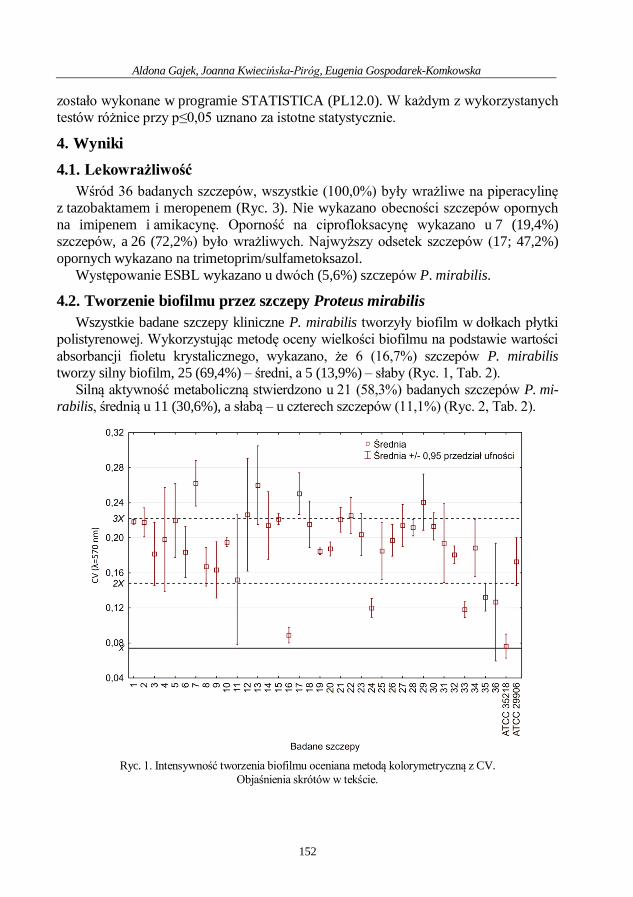
Aldona Gajek, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
152
zostało wykonane w programie STATISTICA (PL12.0). W każdym z wykorzystanych
testów różnice przy p≤0,05 uznano za istotne statystycznie.
4. Wyniki
4.1. Lekowrażliwość
Wśród 36 badanych szczepów, wszystkie (100,0%) były wrażliwe na piperacylinę
z tazobaktamem i meropenem (Ryc. 3). Nie wykazano obecności szczepów opornych
na imipenem i amikacynę. Oporność na ciprofloksacynę wykazano u 7 (19,4%)
szczepów, a 26 (72,2%) było wrażliwych. Najwyższy odsetek szczepów (17; 47,2%)
opornych wykazano na trimetoprim/sulfametoksazol.
Występowanie ESBL wykazano u dwóch (5,6%) szczepów P. mirabilis.
4.2. Tworzenie biofilmu przez szczepy Proteus mirabilis
Wszystkie badane szczepy kliniczne P. mirabilis tworzyły biofilm w dołkach płytki
polistyrenowej. Wykorzystując metodę oceny wielkości biofilmu na podstawie wartości
absorbancji fioletu krystalicznego, wykazano, że 6 (16,7%) szczepów P. mirabilis
tworzy silny biofilm, 25 (69,4%) – średni, a 5 (13,9%) – słaby (Ryc. 1, Tab. 2).
Silną aktywność metaboliczną stwierdzono u 21 (58,3%) badanych szczepów P. mi-
rabilis, średnią u 11 (30,6%), a słabą – u czterech szczepów (11,1%) (Ryc. 2, Tab. 2).
Ryc. 1. Intensywność tworzenia biofilmu oceniana metodą kolorymetryczną z CV.
Objaśnienia skrótów w tekście.
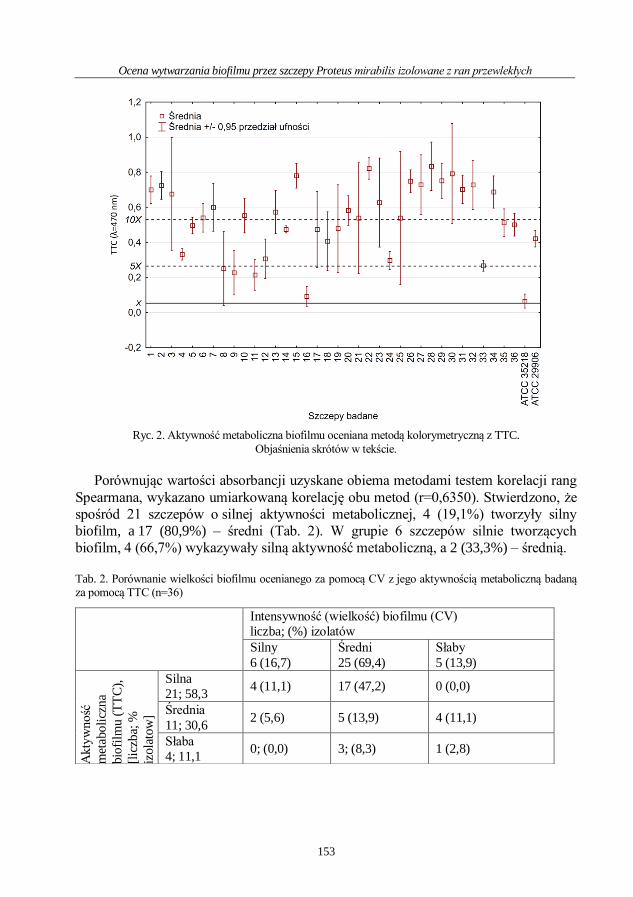
Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis izolowane z ran przewlekłych
153
Ryc. 2. Aktywność metaboliczna biofilmu oceniana metodą kolorymetryczną z TTC.
Objaśnienia skrótów w tekście.
Porównując wartości absorbancji uzyskane obiema metodami testem korelacji rang
Spearmana, wykazano umiarkowaną korelację obu metod (r=0,6350). Stwierdzono, że
spośród 21 szczepów o silnej aktywności metabolicznej, 4 (19,1%) tworzyły silny
biofilm, a 17 (80,9%) – średni (Tab. 2). W grupie 6 szczepów silnie tworzących
biofilm, 4 (66,7%) wykazywały silną aktywność metaboliczną, a 2 (33,3%) – średnią.
Tab. 2. Porównanie wielkości biofilmu ocenianego za pomocą CV z jego aktywnością metaboliczną badaną za pomocą TTC (n=36)
Intensywność (wielkość) biofilmu (CV)
liczba; (%) izolatów
Silny
6 (16,7)
Średni
25 (69,4)
Słaby
5 (13,9)
Akty
wność
met
aboli
czna
bio
film
u (
TT
C),
[lic
zba;
%
izola
tow
]
Silna
21; 58,3 4 (11,1) 17 (47,2) 0 (0,0)
Średnia
11; 30,6 2 (5,6) 5 (13,9) 4 (11,1)
Słaba
4; 11,1 0; (0,0) 3; (8,3) 1 (2,8)

Aldona Gajek, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
154
4.3. Dendrogram podobieństwa szczepów P. mirabilis
Na podstawie profili białkowych wykonano dendrogram podobieństwa badanych
szczepów (Rys. 3), w którym wyróżniono dwie główne gałęzie. Pierwsza z nich (A)
obejmuje 13 (36,1%), a druga (B) – 23 (63,9%) badane szczepy P. mirabilis.
Wśród 13 izolatów zaliczonych do gałęzi A, 2 tworzyły silny biofilm, 9 – średni,
a 2 – słaby. Spośród 23 izolatów z gałęzi B, słaby biofilm tworzyły 3 szczepy, 16 –
średni, a 4 – silny. Porównując wartości absorbancji formazanu stwierdzono niższą
medianę absorbancji dla szczepów w gałęzi A (0,512) niż w gałęzi B (0,582), ale
różnice nie były istotne statystycznie (p=0,2813; test U Manna-Whithey’a). Wartości
median absorbancji CV były zbliżone do siebie w obu gałęziach (A=0,198, B=0,197).
Wśród 13 szczepów wyodrębnionych w gałęzi A było więcej izolatów wrażliwych
na amoksycylinę z kwasem klawulanowym (84,6%), ceftazydym (100,0%), imipenem
(69,2%), wszystkie badane aminoglikozydy (gentamycyna – 92,3%, tobramycyna –
100,0%, amikacyna – 100,0% szczepów), ciprofloksacynę (76,9%) i trimeto-
prim/sulfametoksazol (53,8%) niż wśród izolatów wyszczególnionych w gałęzi B
(odpowiednio: 65,2%, 95,7%, 60,9%, 78,3%, 82,6%, 82,6%, 69,6% i 52,2%
szczepów). Uzyskane różnice odsetka szczepów wrażliwych i niewrażliwych
(średniowrażliwych i opornych) w obu gałęziach dendrogramu nie były istotne
statystycznie (Tab. 3). Wszystkie szczepy oporne na tobramycynę (3; 100%)
i ciprofloksacynę (7; 100,0%) i większość szczepów opornych na amoksycylinę
z kwasem klawulanowym (7; 87,5%) oraz gentamycynę (4; 80%) należały do izolatów
zakwalifikowanych do gałęzi B.
5. Dyskusja
Wśród szczepów Proteus mirabilis obserwuje się wzrost oporności na antybiotyki,
również na leki „ostatniej szansy”, np. z grupy karbapenemów [16]. W niniejszej pracy
nie wykazano jednak obecności szczepów opornych na meropenem oraz imipenem.
Może to być wynikiem pochodzenia szczepów z zakażeń ran przewlekłych, w leczeniu
których rzadko stosuje się leki tej grupy. Podobne wyniki zostały uzyskane przez
Mihai i wsp. [17], którzy oceniali wrażliwość bakterii, w tym jednego szczepu P.
mirabilis, izolowanych z przewlekłych zmian skórnych. Spośród 7 badanych przez
Mihai i wsp. [17] szczepów z rodziny Enterobacteriaceae, wszystkie były wrażliwe na
imipenem, meropenem i ertapenem. Nie stwierdzono również obecności szczepów
opornych na amikacynę [17], podobnie jak w badaniach własnych.
Według Omar i wsp. [18] wrażliwość na ceftazydym wśród szczepów P. mirabilis
wynosi około 96%, na cefotaksym – około 82%, na ciprofloksacynę – około 54%. W niniejszej pracy otrzymano bardzo podobne wyniki wrażliwości badanych szcze-
pów na cefalosporyny, odpowiednio: 97% i 94%. Wykazano natomiast wyższy odsetek
(72%) szczepów wrażliwych na ciprofloksacynę.
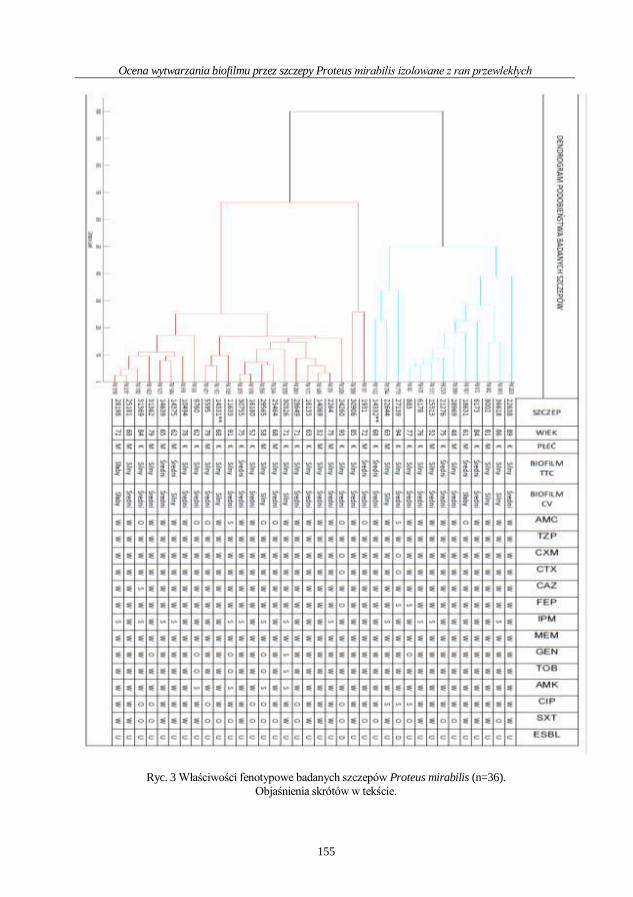
Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis izolowane z ran przewlekłych
155
Ryc. 3 Właściwości fenotypowe badanych szczepów Proteus mirabilis (n=36).
Objaśnienia skrótów w tekście.
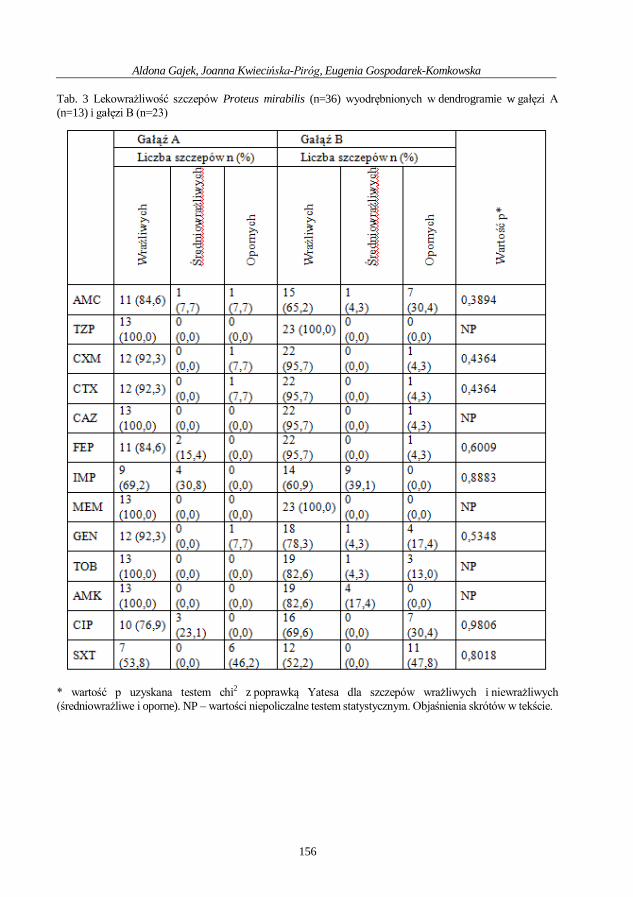
Aldona Gajek, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
156
Tab. 3 Lekowrażliwość szczepów Proteus mirabilis (n=36) wyodrębnionych w dendrogramie w gałęzi A
(n=13) i gałęzi B (n=23)
* wartość p uzyskana testem chi2 z poprawką Yatesa dla szczepów wrażliwych i niewrażliwych
(średniowrażliwe i oporne). NP – wartości niepoliczalne testem statystycznym. Objaśnienia skrótów w tekście.

Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis izolowane z ran przewlekłych
157
Wang i wsp. [19] zlokalizowali plazmidowy gen kodujący oporność na ciproflo-ksacynę u szczepu P. mirabilis o obniżonej wrażliwość na ten chemioterapeutyk. Szczep pozostawał wrażliwy na leki przeciwdrobnoustrojowe, poza trimeto-primem/sulfametoksazolem, co koreluje z uzyskanymi w niniejszej pracy wynikami. Wszystkie oporne na ciprofloksacynę szczepy P. mirabilis były również oporne na trimetoprim/sulfametoksazol. Szczepy te występowały wyłącznie w jednej (B) gałęzi dendrogramu, co jest prawdopodobnie związane z obecnością w komórce bakteryjnej białka odpowiedzialnego za występowanie oporności.
Wszystkie szczepy oporne na tobramycynę i większość szczepów opornych na gentamycynę zostało zakwalifikowanych do jednej gałęzi dendrogramu (B). Może to być efektem obecności plazmidu zawierającego geny zaangażowane w oporność P. mirabilis na aminoglikozydy, który został zbadany przez Wachino i wsp. [20]. Produktem tego genu jest białko, którego widmo mogło zostać uwzględnione podczas tworzenia dendrogramu badanych w niniejszej pracy szczepów, co będzie przed-miotem dalszych badań.
Saito i wsp. [21] zaobserwowali, iż wśród szczepów P. mirabilis izolowanych z moczu oporność na ciprofloksacynę jest związana m.in. z produkcją ESBL. W badaniach własnych wykazano obecność jedynie dwóch szczepów z mechanizmem oporności typu ESBL, z których tylko jeden był oporny na ciprofloksacynę. Badania Biendo i wsp. [22] wykazały, że szczepy wytwarzające ESBL charakteryzują się opornością na trimetoprim/sulfametoksazol. W badaniach własnych obydwa szcze-py wytwarzające ESBL były oporne na kotrimoksazol.
Wszystkie badane w przedstawianej pracy szczepy P. mirabilis tworzyły biofilm. Podobne wyniki uzyskali Mihai i wsp. [17], którzy oceniali zdolność tworzenia briofil-mu przez pałeczki z rodziny Enterobacteriaceae wyizolowane z ran przewlekłych, w tym P. mirabilis. Jest to również zbieżne z wynikami uzyskanymi przez De i wsp. [23]. Dwa izolowane przez De i wsp. [23] szczepy P. mirabilis tworzyły biofilm o umiarkowanym stopniu. W badaniach własnych również zaobserwowano, że najwięcej szczepów P. mirabilis (69,4%) tworzyło biofilm z umiarkowaną intensyw-nością.
W przedstawianej pracy oceniano wielkość tworzonego biofilmu, jak również jego aktywność metaboliczną. Wykazano, że biofilm większości szczepów P. mirabilis (58,3%) wykazywał silną aktywność metaboliczną. Podobny odsetek szczepów P. mirabilis, których biofilm wykazywał silną i bardzo silną aktywność metaboliczną (61,5%) stwierdzono we wcześniejszych badaniach [15].
Podsumowując, szczepy z gatunku P. mirabilis izolowane z ran przewlekłych posiadają zdolność tworzenia biofilmu z różną intensywnością i o różnej aktywności metabolicznej.
Literatura:
1. Różalski A., Kwil I., Torzewska A., Baranowska M., Stączek P. Bakterie z rodzaju Proteus – cechy i czynniki chorobotwórczości, Postępy Higieny i Medycyny Doświadczalnej., 61 (2007), s. 204-219.
2. Kwiecińska-Piróg J., Skowron K., Zniszczol K., Gospodarek E. The assessment of Proteus mirabilis susceptibility to ceftazidime and ciprofloxacin and the impact of these antibiotics

Aldona Gajek, Joanna Kwiecińska-Piróg, Eugenia Gospodarek-Komkowska
158
at subinhibitory concentrations on Proteus mirabilis biofilms, BioMed Research International., 2013 (2013), s. 930876.
3. Endimiani A. Luzzaro F, Brigante G, Perilli M, Lombardi G, Amicosante G, Rossolini GM, Toniolo A. Proteus mirabilis bloodstream infections: risk factors and treatment outcome related to the expression of extended-spectrum β-lactamases, Antimicrobial Agents and Chemotherapy., 49 (2005), s. 2598-2605.
4. Jones S. M., Yerly J., Hu Y., Ceri H., Martinuzzi R. Structure of Proteus mirabilis biofilms grown in artificial urine and standard laboratory media, FEMS Microbiology Letters, 268 (2007), s. 16-21.
5. Bartoszewicz M., Rygiel A. Biofilm jako podstawowy mechanizm zakażenia miejsca operowanego — metody prewencji w leczeniu miejscowym, Chirurgia Polska., 8 (2006), s. 171-178.
6. Jacobsen S. M. Shirtliff M. E. Proteus mirabilis biofilms and catheter-associated urinary tract infections, Virulence, 2 (2011), s. 460-465.
7. James G. A., Swogger E., Wolcott R., deLancey Pulcini E., Secor P., Sestrich J., Costeron J. W., Stewart P. S. Biofilms in chronic wounds, Wound Repair and Regeneration., 16 (2008), s. 37-44.
8. K. Kirketerp-Møller, K. Zulkowski, G. James, Chronic Wound Colonization, Infection, and Biofilms, w Biofilm Infections, Springer, New York, NY, (2011), s. 11-24.
9. Hryniewicz W., Kulig J., Ozorowski T., Mól A., Kulig P., Wąchol D. Stosowanie antybiotyków w wybranych zakażeniach skóry i tkanek miękkich, Ministerstwo Zdrowia., http://www.antybiotyki.edu.pl/pdf/rekomendacje-stosowanie-ant-w-wybranych-zak-skory.pdf, (2013).
10. National Pressure Ulcer Advisory Panel, European Pressure Ulcer Advisory Panel and Pan Pacific Pressure Injury Alliance. Prevention and Treatment of Pressure Ulcers: Quick Reference Guide. Emily Haesler (Ed.). Cambridge Media: Perth, Australia; (2014).
11. High K. P., Bradley S. F., Gravenstein S., Mehr D. R., Quagliarello V. J., Richards C., Yoshikawa T. T. Clinical practice guideline for the evaluation of fever and infection in older adult residents of long-term care facilities: 2008 Update by the Infectious Diseases Society of America, Clinical Infectious Diseases., 48 (2009), s. 149-71.
12. Siddiqui A. R., Bernstein J. M. Chronic wound infection: facts and controversies, Clinics in Dermatology., 28 (2010), s. 519-526.
13. http://www.korld.edu.pl/pdf/eucast/EUCAST_breakpoints_tlumaczenie_versja_6_2016.pdf 14. http://www.korld.edu.pl/pdf/02-Rek2009-Paleczki_z_rodziny_Enterobacteriaceae.pdf 15. Kwiecinska-Piróg J., Bogiel T., Skowron K., Wieckowska E., Gospodarek E. Proteus
mirabilis biofilm - qualitative and quantitative colorimetric methods-based evaluation, Brazilian Journal of Microbiology., 45 (2015), s. 1423-1431.
16. Szabo D., Paterson D. L. Proteus – infectious disease and antimicrobial agents, Proteus species., http://www.antimicrobe.org/b226.asp, (2012).
17. Mihai M. M., A. Holban M., Giurcaneanu C. Identification and phenotypic characterization of the most frequent bacterial etiologies in chronic skin ulcers, Romanian Journal of Morphology & Embryology., 55 (2014), s. 1401-1408.
18. Omar A., Wright J. B., Schultz G., Burrell R., Nadworny P. Microbial biofilms and chronic wounds, Microorganisms., 5 (2017).
19. Wang M., Guo Q., Xu X., Wang X., Ye X., Wu S., Hooper D. Wang M., New Plasmid-Mediated Quinolone Resistance Gene, qnrC, Found in a Clinical Isolate of Proteus mirabilis, Antimicrobial Agents and Chemotherapy, 5 (2009), s. 1892-1897.
20. Wachino J., Yamane K., Shibayama K., Kurokawa H., Shibata N., Suzuki S., Doi Y., Kimura K., Ike Y., Arakawa Y. Novel Plasmid-Mediated 16S rRNA Methylase, RmtC, Found in a Proteus mirabilis Isolate Demonstrating Extraordinary High-Level Resistance

Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis izolowane z ran przewlekłych
159
against Various Aminoglycosides, Antimicrobial Agents and Chemotherapy, 1 (2006), s. 178-184.
21. Saito R., Okugawa S., Kumita W., Sato K., CHida T., Okamura N., Moriya K., Koike K. Clinical epidemiology of ciprofloxacin-resistant Proteus mirabilis isolated from urine samples of hospitalised patients, Clinical Microbiology and Infection., 12 (2007), s. 1204-1206.
22. Bienido M., Thomas D., Laurans G., Hamdad-Daoudi F., Canarelli B., Rousseau F., Castelain S., Eb F. Molecular diversity of Proteus mirabilis isolates producing extended-spectrum β-lactamases in a French university hospital, Clinical Microbiology and Infection., 5 (2005), s. 395-401.
23. De A., Raj H. J., Haldar J., Mukherjee P., Maiti P. K. Biofilm colonization in chronic treatment refractory infections presenting with discharging sinuses: a study in a tertiary care hospital of Eastern India, Journal of Laboratory Physicians., 9 (2017), s. 125-131.
Ocena wytwarzania biofilmu przez szczepy Proteus mirabilis izolowane z ran przewlekłych
Streszczenie Rany przewlekłe diagnozowane są w Polsce u ponad 500 000 chorych rocznie. U ponad 50% z nich roz-poznawane jest zakażenie i włączana antybiotykoterapia, często nieskuteczna. Jest to związane, m. in., ze zdolnością drobnoustrojów do tworzenia struktury biofilmu, który chroni drobnoustroje. Obecność biofilmu wykazano również w zakażeniach ran przewlekłych, w których etiologii znaczącą rolę odgrywają Gram-ujemne pałeczki z gatunku Proteus mirabilis. Celem pracy była ocena wytwarzania biofilmu przez 36 szczepów P. mirabilis izolowanych z zakażonych ran przewlekłych. Wykorzystano dwie metody kolorymetryczne oparte na pomiarze absorbancji fioletu krystalicznego oraz formazanu (produkt redukcji chlorku 2,3,5-trójfenylotetrazoliowego). Kontrolę dodatnią stanowił szczep wzorcowy P. mirabilis ATCC 29906, tworzący biofilm, a ujemną – szczep słabo tworzący biofilm – E. coli ATCC 35218. Na podstawie wyników absorbancji utworzono przedziały intensywności tworzenia biofilmu i jego aktywności metabolicznej. Wykazano, że wszystkie badane szczepy tworzyły biofilm. Silną aktywność metaboliczną stwierdzono u 21 (58,3%) badanych szczepów P. mirabilis, natomiast wysoką absorbancję fioletu krystalicznego, korespondującą z wielkością tworzonego biofilmu, wykryto u 6 (16,7%) szczepów. Porównując obie stosowane metody testem korelacji rang Spearmana (Statistica 12.0 PL), wykazano umiarkowaną korelację obu metod (r=0,6350). Słowa kluczowe: Proteus mirabilis, biofilm, rany przewlekłe
Evaluation of biofilm by Proteus mirabilis strain isolated from chronic wounds
Abstract Chronic wounds are diagnosed in Poland at over 500,000 people every year. In more than 50% of them infections are diagnosed and treatment with antibiotics, often ineffective. It may be related, among others, with the ability of microorganisms to formation biofilm structure. The biofilm is found in chronic wounds infections, where the presence of Proteus mirabilis is significant. The aim of the study was to evaluate the biofilm formation of 36 P. mirabilis strains isolated from chronic wounds infections. In the study were used two colorimetric methods based on measuring the absorbance of crystal violet and formazan (reduction product of 2,3,5-triphenyltetrazolium chloride). The reference strain of P. mirabilis ATCC 29906, forming a biofilm, was a positive control, and E. coli ATCC 35218, weakly forming biofilm, was a negative control. On the basis of the absorbance value, the ranges of the intensity of biofilm formation and metabolic activity were established. It was shown that all of studied strains formed biofilm. Strong metabolic activity was found in 21 (58,3%) of P. mirabilis strains. High absorbance of crystal violet, corresponding to biofilm biomass, was observed in 6 (16,7%). Comparing these two methods using Spearmans correlation (Statistica 12.0 PL) demonstrated moderate correlation between both used methods (r = 0.6350). Keywords: Proteus mirabilis, biofilm, chronic wounds,

160
Jolanta Małajowicz1, Ewa Majewska
2, Dorota Nowak
3
Porównanie wydajności syntezy gamma-dekalaktonu
w hodowlach ciągłych mutanta drożdży
Yarrowia lipolytica MTLY40-2p
1. Wprowadzenie
Gamma-dekalakton (rys. 1) jest wewnątrzcząsteczkowym estrem kwasu 4-
hydroksydekanowego. Ma postać oleistej, bezbarwnej cieczy i charakteryzuje się
intensywnym zapachem kremowo-brzoskwiniowym, wyczuwalnym już przy bardzo
niskim stężeniu – ok. 0,088 pmm w roztworach wodnych. Występuje naturalnie
w wielu owocach (truskawki, brzoskwinie, ananasy i inne) oraz produktach mlecznych
fermentowanych [1]. Z uwagi na posiadany status GRAS (ang. Generally Recognized
as Safe) powszechnie wykorzystywany jest w przemyśle spożywczym i kosme-
tycznym, jako jeden ze składników aromatów.
OO
Rysunek 1. Wzór gamma-dekalaktonu [opracowanie własne]
Pomimo iż gamma-dekalakton jest naturalnym składnikiem nuty zapachowej wielu
owoców, to ze względu na niewielką w nich jego zawartość, ekstrakcja bądź destylacja
z naturalnych źródeł nie jest opłacalna. Dlatego też aktualna produkcja gamma-
dekalaktonu bazuje głównie na metodach chemicznych lub biotechnologicznych.
Wadą metod chemicznych jest często ich wieloetapowość i brak enancjoselektywności.
Metody biotechnologiczne, pomimo że charakteryzują się niewielką wydajnością, to
jednak umożliwiają syntezę pożądanego enancjomeru (R) i odpowiadają na zapo-
trzebowanie konsumentów odnośnie wykorzystania składników pochodzenia natu-
ralnego w żywności.
Biotechnologiczna produkcja laktonów bazuje na procesach oksydacyjnej
degradacji kwasów tłuszczowych. Najlepiej poznanym szlakiem tych procesów jest β-
oksydacja, zachodząca w peroksysomach drożdży [2]. Możliwość wykorzystania
procesu β-oksydacji w biotransformacji kwasów tłuszczowych odkryto już w 1963
roku. Przy zastosowaniu drożdży Candida tropicalis, zespół Okui i wsp. [3] prze-
1 [email protected], Katedra Chemii, Wydział Nauk o Żywności, Szkoła Główna Gospodarstwa
Wiejskiego w Warszawie, www.wnoz.sggw.pl 2 [email protected], Katedra Chemii, Wydział Nauk o Żywności, Szkoła Główna Gospodarstwa
Wiejskiego w Warszawie, www.wnoz.sggw.pl 3 [email protected], Katedra Inżynierii Żywności i Organizacji Produkcji, Wydział Nauk o Żywności,
Szkoła Główna Gospodarstwa Wiejskiego w Warszawie, www.wnoz.sggw.pl

Porównanie wydajności syntezy gamma-dekalaktonu w hodowlach ciągłych mutanta drożdży
Yarrowia lipolytica MTLY40-2p
161
prowadził rozkład kwasu rycynolowego do C16, C14, C12, dekalaktonu i kwasu 2-
hydroksyoktanowego. Od momentu tego odkrycia prowadzone są nieustannie badania
laboratoryjne nad wykorzystaniem różnych gatunków mikroorganizmów w wydajnej
biosyntezie laktonów, w tym szczególnie gamma-dekalaktonu.
Obecnie gamma-dekalakton syntetyzowany jest głównie na drodze biotransformacji
kwasu rycynolowego. Przekształcanie tego kwasu do gamma-dekalaktonu polega na
odszczepieniu od karboksylowego końca ośmiowęglowego fragmentu, które następuje
w wyniku czterech cykli β-oksydacji. Powstały w reakcji kwas 4-hydroksydekanowy
ulega wewnątrzcząsteczkowej estryfikacji do gamma-dekalaktonu (rys. 2) Sprzyja-
jącymi warunkami laktonizacji kwasu jest niskie pH i podwyższona temperatura [4].
Rysunek 2. Schemat przekształcania kwasu rycynolowego do gamma-dekalaktonu,
opracowanie własne na podstawie [4]
Biotransformacja kwasu rycynolowego do gamma-dekalaktonu prowadzona była
dotychczas przy użyciu różnych mikroorganizmów: Yarrowia lipolytica [5, 6],
Sporobolomyces odorus [7], Sporidiobolus salmonicolor [8], Sporidiobolus ruineni [9],
Rhodotorula aurantiaca [10] i innych. Gatunek Yarrowia lipolytica charakteryzuje się
jednak najwyższą wydajnością w produkcji gamma-dekalaktonu i posiada największą
liczbę zidentyfikowanych genów, kodujących enzymy specjalizujące się w degradacji
hydrofobowych substratów [11]. W komórkach Yarrowia lipolytica występuje sześć
oksydaz acylo-CoA (Aox1 do Aox6 kodowanych przez geny POX1 do POX6). Geny
POX wykazują różną preferencję względem długości degradowanego łańcucha
węglowego. Oksydaza Aox2 wykazuje specyficzność względem długołańcuchowych
substratów, zaś Aox3 jest aktywna w stosunku do związków krótkołańcuchowych.

Jolanta Małajowicz, Ewa Majewska, Dorota Nowak
162
Udowodniono, iż trzy pozostałe enzymy – Aox1, Aox4 oraz Aox5 przejawiają bardzo
słabą aktywność względem większości dostępnych substratów [12]. Stąd też wszelkie
modyfikacje genetyczne bazują głównie na manipulacji w obrębie genów POX2
i POX3.
W ramach niniejszej pracy podjęto próbę porównania produktywności gamma-
dekalaktonu przez zmodyfikowany szczep drożdży Yarrowia lipolytica MTLY40-2p
namnażany w kolbach i bioreaktorze. Modyfikacja tego szczepu polegała na usunięciu
trzech (spośród pięciu) aktywnych oksydaz acyl-CoA – Aox3, Aox4 oraz Aox5.
Szczególne znaczenie w tej modyfikacji miało ograniczenie aktywności Aox3, która
odpowiedzialna jest za kontynuację procesu oksydacji za dziesięciowęglowym
łańcuchem węglowym, a tym samym przyczynia się do degradacji wytworzonego
gamma-dekalaktonu.
2. Materiał i metodyka badań
W badaniach wykorzystano modyfikowany genetycznie szczep drożdży Yarrowia
lipolytica MTLY40-2p pochodzący z Kolekcji Kultur Drobnoustrojów Laboratorium
GPMA Uniwersytetu Burgundzkiego w Dijon we Francji. Szczep przechowywano
w 10% roztworze glicerolu w warunkach głębokiego zamrożenia – 80°C.
2.1. Warunki hodowli mikroorganizmów
W celu przygotowania prekultury, komórki drożdży Yarrowia lipolytica MTLY40-
2p przeszczepiano wstępnie na szalki Petriego, zawierające stałe podłoże YPGA,
(skład podłoża: ekstrakt drożdżowy 10 g/dm3; pepton 20 g/ dm
3; glukoza 20 g/dm
3;
agar 20 g/dm3) po czym namnażano przez 48h w temperaturze 27°C. Wyhodowane
komórki wykorzystywano następnie do zaszczepienia prekultury (podłoże YPG:
ekstrakt drożdżowy 10 g/dm3; pepton 20 g/dm
3; glukoza 20 g/dm
3 – objętość 50 cm
3).
Prekulturę namnażano przez 24 h w temperaturze 27°C przy poziomie mieszania 140
rpm. Po tym czasie 1 cm3 zawiesiny przeszczepiano na właściwe podłoże biotrans-
formacyjne.
2.2. Reakcja biotransformacji oleju rycynowego do gamma-dekalaktonu
Reakcję biotransformacji prowadzono w kolbach o objętości całkowitej 500 cm3
lub bioreaktorze BIOFLO 3000 (New Brunswick Scientific Edison, N.I., USA)
o objętości całkowitej 5 dm3. W obu hodowlach stosowano podłoże zawierające
w swym składzie olej rycynowy – 100 g/dm3; pepton 20 g/dm
3 i surfaktant – Tween 20
– 0,5 g/dm3. Hodowlę prowadzono przez okres 7 dni, w temperaturze 27°C, przy
prędkości mieszania 140 rpm. W bioreaktorze hodowla była dodatkowo napowietrzana
na poziomie 0,2 vvm z wykorzystaniem sprężonego powietrza podawanego przez
barboter.
2.3. Ekstrakcja gamma-dekalaktonu
Stężenie gamma-dekalaktonu w podłożu analizowano w każdym dniu trwania
reakcji, przez okres 7 dni. Z mieszaniny reakcyjnej pobierano 1,5 cm3 fazy wodnej lub
olejowej, kolejno dodawano 50 µl 1 M kwasu solnego (kwas dodawany celem zaha-

Porównanie wydajności syntezy gamma-dekalaktonu w hodowlach ciągłych mutanta drożdży
Yarrowia lipolytica MTLY40-2p
163
mowania aktywności enzymów i laktonizacji kwasu 4-hydroksydekanowego) oraz 20
µl gamma-undekalaktonu (C11) – stosowanego jako standard wewnętrzny. Całość
ekstrahowano za pomocą 1,5 cm3 eteru dietylowego.
2.4. Analiza chromatograficzna gamma-dekalaktonu
Po ekstrakcji, próbki analizowano metodą chromatografii gazowej. Analizę prowa-
dzono na aparacie YL 6100 Young Lin Instrument z wykorzystaniem kolumny
kapilarnej typu BPX 70 (30 m x 0,25 mm). Gazem nośnym w analizach był azot,
temperatura detektora wynosiła 280°C, przepływ ustalono na poziomie 1,1 cm3/min.
Czas trwania analizy wynosił łącznie 16 min. Czasy retencji standardu wewnętrznego
– gamma-undekalaktonu (C11) oraz gamma-dekalaktonu (C10) wynosiły odpowiednio
8,9 min oraz 10,2 min.
2.5. Pomiar gęstości optycznej OD600
Każdego dnia trwania hodowli z podłoża biotransformacyjnego pobierano próbę
celem pomiaru gęstości optycznej komórek drożdży. 1 cm3 pobranej zawiesiny
poddawano wirowaniu, stosując mikrowirówkę typu Eppendorf 5418 (ROTH). Super-
natant zlewano znad osadu, a osad zawieszano w 1 cm3 wody destylowanej. Dla tak
przygotowanej próbki, po odpowiednim jej rozcieńczeniu, dokonywano pomiaru
gęstości optycznej (OD) z zastosowaniem spektrofotometru UV-VIS 1800 EnviSense
przy długości fali 600 nm. Pomiar wykonywano w 3 powtórzeniach.
2.6. Pomiar suchej masy drożdży
Podczas reakcji biotransformacji analizowano plon suchej masy drożdży. Z podłoża
biotransformacyjnego do gilzy pobierano 10 cm3 zawiesiny. Celem oddzielenia
komórek od supernatantu, komórki odwirowywano z prędkością 8000 obr/min na
wirówce typu MPW 351 przez okres 5 minut. Po przepłukaniu komórek wodą desty-
lowaną i ponownym odwirowaniu, były one suszone w suszarce, w temperaturze ok.
100˚C do momentu uzyskania stałej masy. Pomiar wykonywano w dwóch powtó-
rzeniach, podając jako wynik plon suchej masy drożdży na dm3 podłoża biotrans-
formacyjnego.
2.7. Pomiar żywotności komórek drożdży
Celem pomiaru żywotności komórek, z podłoża biotransformacyjnego pobierano
próbę o objętości 1 cm3 i rozcieńczano ją odpowiednio w jałowym roztworze soli
fizjologicznej. Kolejno pobierano 10 µl właściwie rozcieńczonej zawiesiny drożdży
i zaszczepiano nią powierzchniowo stałe podłoże YPGA na płytce Petriego. Zaszcze-
pione płytki inkubowano w temperaturze pokojowej przez około 48 h, po czym zli-
czano liczbę kolonii drożdży wyrosłych na płytkach. Doświadczenie prowadzono
w dwóch powtórzeniach podając w wyniku końcowym liczbę jtk (jednostek
tworzących kolonię)/cm3.
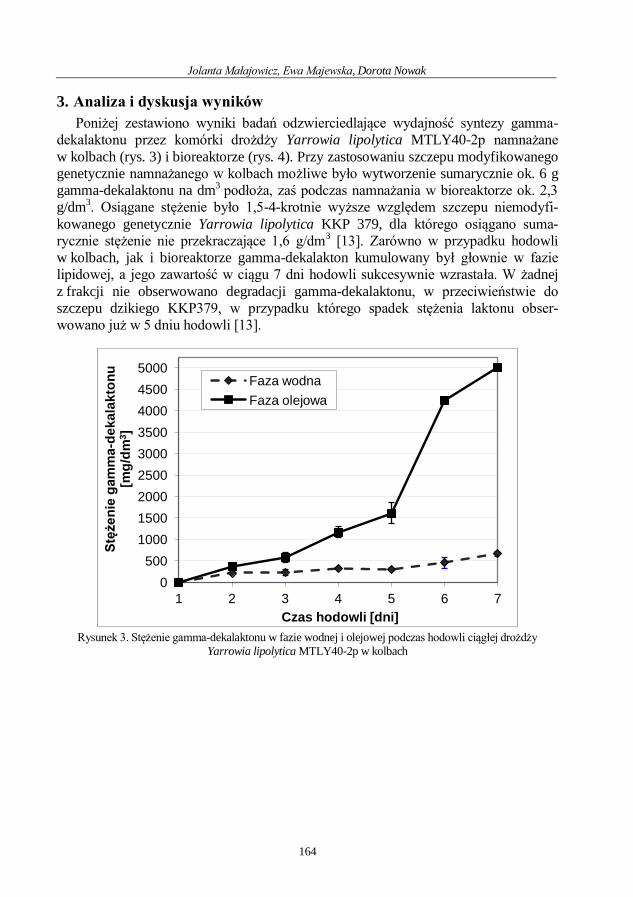
Jolanta Małajowicz, Ewa Majewska, Dorota Nowak
164
3. Analiza i dyskusja wyników
Poniżej zestawiono wyniki badań odzwierciedlające wydajność syntezy gamma-
dekalaktonu przez komórki drożdży Yarrowia lipolytica MTLY40-2p namnażane
w kolbach (rys. 3) i bioreaktorze (rys. 4). Przy zastosowaniu szczepu modyfikowanego
genetycznie namnażanego w kolbach możliwe było wytworzenie sumarycznie ok. 6 g
gamma-dekalaktonu na dm3
podłoża, zaś podczas namnażania w bioreaktorze ok. 2,3
g/dm3. Osiągane stężenie było 1,5-4-krotnie wyższe względem szczepu niemodyfi-
kowanego genetycznie Yarrowia lipolytica KKP 379, dla którego osiągano suma-
rycznie stężenie nie przekraczające 1,6 g/dm3 [13]. Zarówno w przypadku hodowli
w kolbach, jak i bioreaktorze gamma-dekalakton kumulowany był głownie w fazie
lipidowej, a jego zawartość w ciągu 7 dni hodowli sukcesywnie wzrastała. W żadnej
z frakcji nie obserwowano degradacji gamma-dekalaktonu, w przeciwieństwie do
szczepu dzikiego KKP379, w przypadku którego spadek stężenia laktonu obser-
wowano już w 5 dniu hodowli [13].
Rysunek 3. Stężenie gamma-dekalaktonu w fazie wodnej i olejowej podczas hodowli ciągłej drożdży
Yarrowia lipolytica MTLY40-2p w kolbach
0
500
1000
1500
2000
2500
3000
3500
4000
4500
5000
1 2 3 4 5 6 7
Stę
że
nie
gam
ma-d
ek
ala
kto
nu
[m
g/d
m3]
Czas hodowli [dni]
Faza wodna
Faza olejowa
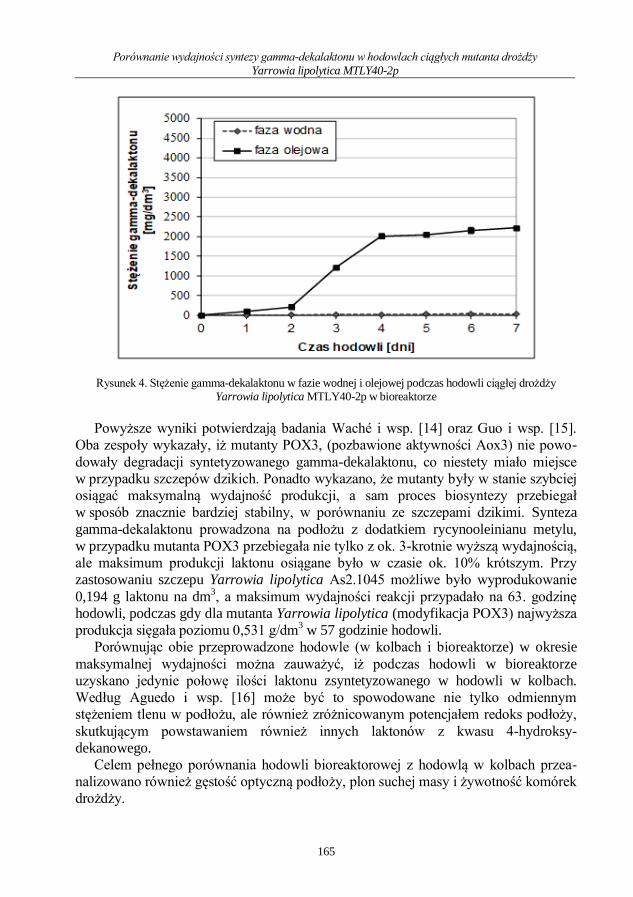
Porównanie wydajności syntezy gamma-dekalaktonu w hodowlach ciągłych mutanta drożdży
Yarrowia lipolytica MTLY40-2p
165
Rysunek 4. Stężenie gamma-dekalaktonu w fazie wodnej i olejowej podczas hodowli ciągłej drożdży Yarrowia lipolytica MTLY40-2p w bioreaktorze
Powyższe wyniki potwierdzają badania Waché i wsp. [14] oraz Guo i wsp. [15].
Oba zespoły wykazały, iż mutanty POX3, (pozbawione aktywności Aox3) nie powo-
dowały degradacji syntetyzowanego gamma-dekalaktonu, co niestety miało miejsce
w przypadku szczepów dzikich. Ponadto wykazano, że mutanty były w stanie szybciej
osiągać maksymalną wydajność produkcji, a sam proces biosyntezy przebiegał
w sposób znacznie bardziej stabilny, w porównaniu ze szczepami dzikimi. Synteza
gamma-dekalaktonu prowadzona na podłożu z dodatkiem rycynooleinianu metylu,
w przypadku mutanta POX3 przebiegała nie tylko z ok. 3-krotnie wyższą wydajnością,
ale maksimum produkcji laktonu osiągane było w czasie ok. 10% krótszym. Przy
zastosowaniu szczepu Yarrowia lipolytica As2.1045 możliwe było wyprodukowanie
0,194 g laktonu na dm3, a maksimum wydajności reakcji przypadało na 63. godzinę
hodowli, podczas gdy dla mutanta Yarrowia lipolytica (modyfikacja POX3) najwyższa
produkcja sięgała poziomu 0,531 g/dm3 w 57 godzinie hodowli.
Porównując obie przeprowadzone hodowle (w kolbach i bioreaktorze) w okresie
maksymalnej wydajności można zauważyć, iż podczas hodowli w bioreaktorze
uzyskano jedynie połowę ilości laktonu zsyntetyzowanego w hodowli w kolbach.
Według Aguedo i wsp. [16] może być to spowodowane nie tylko odmiennym
stężeniem tlenu w podłożu, ale również zróżnicowanym potencjałem redoks podłoży,
skutkującym powstawaniem również innych laktonów z kwasu 4-hydroksy-
dekanowego.
Celem pełnego porównania hodowli bioreaktorowej z hodowlą w kolbach przea-
nalizowano również gęstość optyczną podłoży, plon suchej masy i żywotność komórek
drożdży.
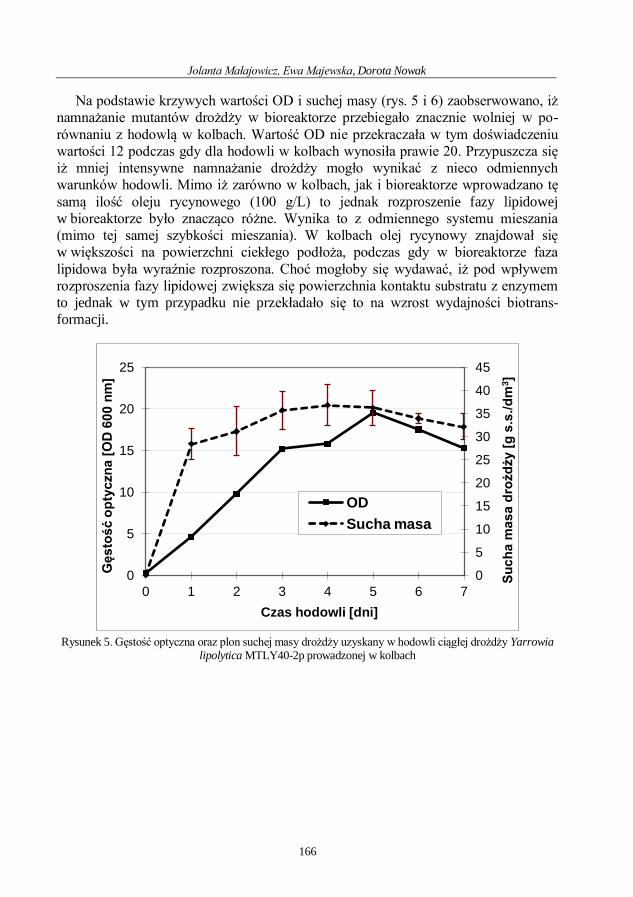
Jolanta Małajowicz, Ewa Majewska, Dorota Nowak
166
Na podstawie krzywych wartości OD i suchej masy (rys. 5 i 6) zaobserwowano, iż
namnażanie mutantów drożdży w bioreaktorze przebiegało znacznie wolniej w po-
równaniu z hodowlą w kolbach. Wartość OD nie przekraczała w tym doświadczeniu
wartości 12 podczas gdy dla hodowli w kolbach wynosiła prawie 20. Przypuszcza się
iż mniej intensywne namnażanie drożdży mogło wynikać z nieco odmiennych
warunków hodowli. Mimo iż zarówno w kolbach, jak i bioreaktorze wprowadzano tę
samą ilość oleju rycynowego (100 g/L) to jednak rozproszenie fazy lipidowej
w bioreaktorze było znacząco różne. Wynika to z odmiennego systemu mieszania
(mimo tej samej szybkości mieszania). W kolbach olej rycynowy znajdował się
w większości na powierzchni ciekłego podłoża, podczas gdy w bioreaktorze faza
lipidowa była wyraźnie rozproszona. Choć mogłoby się wydawać, iż pod wpływem
rozproszenia fazy lipidowej zwiększa się powierzchnia kontaktu substratu z enzymem
to jednak w tym przypadku nie przekładało się to na wzrost wydajności biotrans-
formacji.
Rysunek 5. Gęstość optyczna oraz plon suchej masy drożdży uzyskany w hodowli ciągłej drożdży Yarrowia
lipolytica MTLY40-2p prowadzonej w kolbach
0
5
10
15
20
25
30
35
40
45
0
5
10
15
20
25
0 1 2 3 4 5 6 7
Su
ch
a m
as
a d
rożd
ży [
g s
.s./
dm
3]
Gęs
tość
op
tyczn
a [
OD
60
0 n
m]
Czas hodowli [dni]
OD
Sucha masa
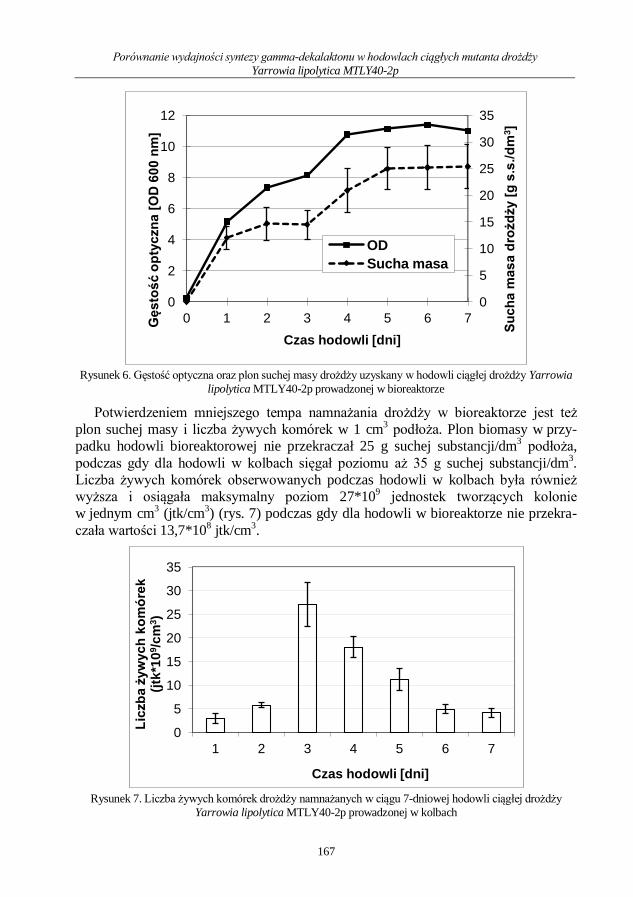
Porównanie wydajności syntezy gamma-dekalaktonu w hodowlach ciągłych mutanta drożdży
Yarrowia lipolytica MTLY40-2p
167
Rysunek 6. Gęstość optyczna oraz plon suchej masy drożdży uzyskany w hodowli ciągłej drożdży Yarrowia
lipolytica MTLY40-2p prowadzonej w bioreaktorze
Potwierdzeniem mniejszego tempa namnażania drożdży w bioreaktorze jest też
plon suchej masy i liczba żywych komórek w 1 cm3 podłoża. Plon biomasy w przy-
padku hodowli bioreaktorowej nie przekraczał 25 g suchej substancji/dm3 podłoża,
podczas gdy dla hodowli w kolbach sięgał poziomu aż 35 g suchej substancji/dm3.
Liczba żywych komórek obserwowanych podczas hodowli w kolbach była również
wyższa i osiągała maksymalny poziom 27*109 jednostek tworzących kolonie
w jednym cm3 (jtk/cm
3) (rys. 7) podczas gdy dla hodowli w bioreaktorze nie przekra-
czała wartości 13,7*108 jtk/cm
3.
Rysunek 7. Liczba żywych komórek drożdży namnażanych w ciągu 7-dniowej hodowli ciągłej drożdży
Yarrowia lipolytica MTLY40-2p prowadzonej w kolbach
0
5
10
15
20
25
30
35
0
2
4
6
8
10
12
0 1 2 3 4 5 6 7
Su
ch
a m
as
a d
rożd
ży [
g s
.s./
dm
3]
Gę
sto
ść
op
tyc
zn
a [
OD
60
0 n
m]
Czas hodowli [dni]
OD
Sucha masa
0
5
10
15
20
25
30
35
1 2 3 4 5 6 7
Lic
zb
a ż
yw
yc
h k
om
óre
k
(jtk
*10
9/c
m3)
Czas hodowli [dni]

Jolanta Małajowicz, Ewa Majewska, Dorota Nowak
168
4. Podsumowanie
Przeprowadzone doświadczenia pozwoliły zaobserwować, iż biotransformacje
prowadzone z zastosowaniem szczepu Yarrowia lipolytica MTLY40-2p modyfiko-
wanego genetycznie pozwalają na uzyskanie od 1,5 do 4-krotnie wyższego stężenia
gamma-dekalaktonu w podłożu hodowlanym, w porównaniu z wcześniej testowanym
przez autorów szczepem dzikim KKP379. Biotransformacja oleju rycynowego skut-
kuje kumulacją gamma-dekalakton głównie (w ok. 90-95%) w fazie lipidowej.
Hodowla bioreaktorowa przyczynia się do ponad 2-krotnie niższego stężenia aromatu
(2,25 g/dm3) w porównaniu z hodowlą w kolbach (5 g/dm
3). Jest to prawdopodobnie
wynikiem zróżnicowanego tempa wzrostu i namnażania się komórek drożdży na
skutek odmiennego natlenienia i zemulgowania podłoży. Przeanalizowane parametry
gęstości optycznej wskazują iż w hodowli bioreaktorowej maksymalny poziom
zmętnienia podłoża wynosił ok. 12, podczas gdy dla hodowli w kolbach wartość OD
kształtowała się na poziomie ok. 20. Potwierdzeniem wyższego tempa namnażania się
mikroorganizmów w kolbach jest również plon suchej masy drożdży, wynoszący
ponad 35 g/dm3, podczas gdy dla hodowli bioreaktorowej sięgał niecałych 25 g
s.s./dm3.
Zebrane w ramach powyższych doświadczeń wyniki stanowią pilotażową analizę
i potwierdzają potencjał możliwości syntezy gamma-dekalaktonu z udziałem szczepu
modyfikowanego genetycznie MTLY-402p. W dalszych pracach badawczych
z udziałem tego szczepu warto podjąć próbę optymalizacji hodowli bioreaktorowej,
kładąc szczególny nacisk na poziom natlenienia podłoża oraz stężenie i poziom
zemulgowania substratu lipidowego – oleju rycynowego.
Literatura
1. Schrader J., Etschmann M.M.W., Sell D., Hilmer J.M., Rabenhorst J. Applied biocatalysis
for the synthesis of natural flavour compounds – current industrial processes and future
prospects, Biotechnology Letters, 26, (2005), s. 463-472.
2. Fickers P., Benetti P.H., Waché Y., Marty A., Mauersberger S., Smit M.S., Nicaud J.M.
Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential
applications, FEMS Yeast Research, 5 (6-7), (2005), s. 527-543.
3. Okui S., Uchiyama M., Mizugaki M. Metabolic conversion of ricinoleic acidy by a certain
microorganism to 8-D-(+)-hydroxy-tetradesc-cis-5-enoic acid, Journal of Biochemistry, 53
(4), (1966), s. 265-270.
4. Waché Y., Aguedo M., Nicaud J-M., Belin J-M. Catabolism of hydroxyacids and
production of lactones by the yeast Yarrowia lipolytica, Applied Microbiology and
Biotechnology, 61, (2003), s. 393-404.
5. Waché Y., Aguedo M., Choquet A., Gatfield I.L., Nicaud J-M., Belin J-M. Role of β-
oxidation enzymes in γ-decalactone production by the yeast Yarrowia lipolytica, Applied
Environmental Microbiology, 67 (12), (2001), s. 5700-5704.
6. Waché Y., Aguedo M., LeDall M.T., Nicuad J-M., Belin J-M. Optimization of Yarrowia
lipolytica's β-oxidation pathway for γ-decalactone production, Journal of Molecular
Catalysis B - Enzymatic, 19 (20), (2002), s. 347-351.

Porównanie wydajności syntezy gamma-dekalaktonu w hodowlach ciągłych mutanta drożdży
Yarrowia lipolytica MTLY40-2p
169
7. Lee S.L., Lin S.J., Chou C.C. Production of γ-decalactone by Sporobolomyces odorus in
jar fermentors as affected by pH, aeration and fed-batch technique, Journal of
Fermentation and Bioengineering, 80 (2), (1995), s. 195-199.
8. Lee S.L., Cheng H.Y., Chen W.C., Chou C.C. Effect of physical factors on the production
of γ-decalactone by immobilized cells of Sporidiobolus salmonicolor, Process
Biochemistry, 34, (1999), s. 845-850.
9. Blin-Perrin C., Molle D., Duffose L., Le-Quere J.L., Viel C., Mauvais G., Feron G.
Metabolism of ricinoleic acid into γ-decalactone: β-oxidation and long chain acyl
intermediates of ricinoleic acid in the genus Sporidiobolus sp., Fems Microbiology Letters,
188, (2000), s. 69-74.
10. Alchihab M., Destain J., Aguedo M., Majad L., Ghalfi H., Wathelet J-P., Thonart P.
Production of γ-decalactone by a psychrophilic and mesophilic strain of the yeast
Rhodotorula aurantica, Applied Biochemistry and Biotechnology, 158 (1) (2009), s. 41-50.
11. Fickers P., Benetti P.H., Waché Y., Marty A., Mauersberger S., Smit M.S., Nicaud J.M.
Hydrophobic substrate utilisation by the yeast Yarrowia lipolytica, and its potential
applications, FEMS Yeast Research, 5 (6-7), (2005), s. 527-543.
12. Waché Y., Aguedo M., Le Dall M.T., Nicaud J.M., Belin JM. Optimization of Yarrowia
lipolytica’s β-oxidation pathway for γ-decalactone production, Journal of Molecular
Catalysis B: Enzymatic. 19-20, (2002), s. 234-351.
13. Krzyczkowska J. The use of castor oil in the production of γ-decalactone by Yarrowia
lipolytica KKP 379, Chemine Technologija 3 (61), (2012), s. 58-61.
14. Waché Y., Aguedo M., Choquet A., Gatfield I.L., Nicaud J.-M., Belin JM. Role of β-
Oxidation Enzymes in γ-Decalactone Production by the Yeast Yarrowia lipolytica, Applied
and Environmental Microbiology. 67 (12), (2001), s. 5700-5704.
15. Guo Y., Feng C., Song H., Wang Z., Ren Q., Wang R. Effect of POX3 gene disruption
using self-cloning CRF1 cassette in Yarrowia lipolytica on the γ-decalactone production,
World Journal of Microbiology and Biotechnology, 27 (2011), s. 2807-2812.
16. Aguedo M., Ly M. H., Belo I., Teixeira J. A., Belin JM., Waché Y. Production of aroma
compounds from lipids, Food Technology and Biotechnology, 42 (4) (2004), s. 327-336.
Porównanie wydajności syntezy gamma-dekalaktonu w hodowlach ciągłych
mutanta drożdży Yarrowia lipolytica MTLY40-2p
Streszczenie
Gamma-dekalakton jest wewnątrzcząsteczkowym estrem kwasu 4-hydroksydekanowego, o intensywnym,
brzoskwiniowo-tłuszczowym zapachu. Powszechnie wykorzystywany jest w przemyśle spożywczym i kosmetycznym, jako jeden ze składników aromatów. Biotechnologiczne metody jego syntezy bazują na
procesach degradacji kwasów tłuszczowych, głównie kwasu rycynolowego, który jest głównym skład-
nikiem oleju rycynowego pozyskiwanego z nasion byliny rącznika pospolitego (łac. Ricinus communis).
W biotechnologicznej syntezie gamma-dekalaktonu stosowane są głównie drożdże Yarrowia lipolytica, posiadające status GRAS i największą liczbę zidentyfikowanych genów, kodujących enzymy specja-
lizujące się w degradacji hydrofobowych substratów.
Celem badań była próba porównania produktywności gamma-dekalaktonu w hodowlach ciągłych mutanta
drożdży Yarrowia lipolytica MTLY40-2p namnażanego w kolbach i bioreaktorze. Modyfikacja tego szczepu polegała na usunięciu trzech (spośród pięciu) aktywnych oksydaz acyl-CoA. W badaniach
przeanalizowano stężenie gamma-dekalaktonu w fazie wodnej i olejowej obu hodowli, by porównać je
z produktywnością szczepów dzikich, dokonano oznaczenia gęstości optycznej podłoży, plonu suchej
masy drożdży oraz oceniono żywotność komórek drożdży.

Jolanta Małajowicz, Ewa Majewska, Dorota Nowak
170
Na podstawie przeprowadzonych doświadczeń zaobserwowano, iż przy zastosowaniu szczepu mody-
fikowanego genetycznie namnażanego w kolbach uzyskano sumarycznie ok. 6 g gamma-dekalaktonu na
dm3 podłoża, zaś podczas namnażania w bioreaktorze jedynie ok. 2,3 g/dm3. Niezależnie jednak od warunków hodowli uzyskane stężenie było 1,5-4-krotnie wyższe względem szczepu niemodyfikowanego
genetycznie Yarrowia lipolytica KKP379, dla którego osiągano sumarycznie stężenie nie przekraczające
1,6 g/dm3. Podczas reakcji biotransformacji oleju rycynowego przez komórki drożdży Yarrowia lipolytica
MTLY40-2p gamma-dekalakton kumulowany był głównie (w ok. 90-95%) w fazie lipidowej. Hodowla bioreaktorowa skutkowała ponad 2-krotnie niższym stężeniem aromatu (2,25 g/dm3) w porównaniu
z hodowlą w kolbach (5 g/dm3). Było to po części wynikiem zróżnicowanego tempa wzrostu i namnażania
się komórek drożdży. Maksymalna gęstość optyczna podczas hodowli bioreaktorowej wynosiła ok. 12,
podczas gdy dla hodowli w kolbach wartość OD kształtowała się na poziomie 20. Słowa kluczowe: gamma-dekalakton, Yarrowia lipolytica, biotransformacje.
The comparison of the gamma-decalactone production in continuous culture
of yeast Yarrowia lipolytica mutants MTLY 40-2p
Abstract Gamma-decalactone, the lactone of 4-hydroxydecanoic acid, is the most widely used flavor lactone
exhibiting an oily-peachy aroma. It is commonly applied in food and cosmetic industry as one of the
flavourings component. It can be obtained biotechnologically by the biotransformation of ricinoleic acid,
the main constituent of castor oil, a natural and non-toxic oil, obtained from the seed of the castor plant, Ricinus communis. In biotechnological synthesis of gamma-decalactone mainly yeast Yarrowia lypolitica
are used, which is considered as non-pathogenic and GRAS strain. In addition, this microorganism has also
the ability to produce lipases that play an important role in the hydrolysis of castor oil.
The purpose of this work was the biosynthesis of gamma-decalactone by genetically modified yeast strain Yarrowia lipolytica MTLY 40-2p (the modification was done within the genes POX1-POX5 which
encodes acyl-CoA oxidases – enzymes active in β-oxidation). In this experiment, flask and bioreactor
fermentations were compared for the production of gamma-decalactone.
It has been observed that with the use of genetically modified strain the yield of the synthesis of gamma-decalactone carried out in flask was 6 g of gamma-decalactone per 1 dm3 of the medium, whereas in
bioreactor the yield was 2.3 g/dm3. Regardless of the reaction condition the concentration of the gamma-
decalactone was 1.5-4-fold higher than the concentration obtained with the use of non-genetically modified
strain Yarrowia lipolytica KKP379 for which it was only 1.6g/dm3. During the reaction catalyzed by Yarrowia lipolytica MTLY-40-2p gamma-decalactone was mainly accumulated in lipid phase (90-95%).
The bioreactor culture produced twofold less gamma-decalactone (the concentration was 2.25 g/dm3) than
the flask culture (5 g/dm3), which was the result of significant differences in growth dynamics and the
proliferation of yeast cells. The maximum optical density of bioreactor culture was 12, whereas optical density of flask culture was about 20.
Keywords: gamma-decalactone, Yarroiwa lipolytica, biotransformations.

171
Magdalena Kręcidło1, Teresa Krzyśko-Łupicka
2
Rozprzestrzenianie się bioaerozoli mikrobiologicznych
– przegląd badań naukowych
1. Wstęp
Powietrze jest wektorem do przemieszczania się mikroorganizmów saprofitycznych
i patogennych. Mikroorganizmy takie jak bakterie, grzyby i wirusy przemieszczają się
wraz z wieloma innymi cząstkami obecnymi w bioaerozolu [1]. Badania wskazują, że
ekspozycja na wysokie stężenie bioaerozolu może powodować w ciężkich przypad-
kach niewydolność oddechową, infekcje mikrobiologiczne, reakcje układu oddecho-
wego, a nawet nowotwory. Sugeruje się, że ekspozycja dzieci w wczesnym dzieciń-
stwie na bioaerozol zawierające endotoksyny bakteryjne przyczynia się do rozwoju
egzemy i astmy [2]. Procesy przebiegające w środowisku, rolnictwo, przemysł, aktyw-
ność biotechnologiczna, przetwarzanie i składowanie odpadów – generują duże ilości
bioaeozoli mikrobiologicznych, które trafiają do atmosfery. Również w budynkach
mieszkalnych, miejscach pracy i użytku publicznego bioaerozole mikrobiologiczne
generowane są przez ludzi i zwierzęta. Szacuje się, że więcej niż 30% martwych
komórek skóry obecnych w powietrzu przenosi bakterie z rodzaju Staphylococcus [3].
Pomimo, że bioaerozol może stanowić poważne zagrożenie dla zdrowia ludzi i zwie-
rząt, czy upraw roślin, to ciągle jeszcze sposoby jego monitorowania są mało dokładne,
drogie lub wymagają skomplikowanej analizy [4]. Natomiast samo rozprzestrzenianie
się zanieczyszczeń mikrobiologicznych w bioaerozolu jest zjawiskiem trudnym do
wychwycenia.
Celem pracy jest zbiorcze przedstawienie dotychczasowych badań dotyczących
źródeł bioaerozoli mikrobiologicznych oraz dróg ich rozprzestrzeniania się.
2. Metodyka badań
Prace przeglądową przygotowano poprzez syntetyczną analizę danych litera-
turowych zgodnie z standardami naukowymi. Przegląd prac dokonano poprzez bazę
danych PubMed (dostęp na 28/03/2017). Zdefiniowano następujące kategorie poję-
ciowe oraz słowa kluczowe, które wykorzystano następnie w poszukiwaniu prac
związanych z tematem: (((bioaerosol OR biological dust OR organic dust OR airborne)
AND (fungi OR mould OR bacteria)) AND (spread OR spreading OR dissemination
OR dispersal OR distribution OR emission)) AND (building OR agriculture OR
atmosphere OR outdoor OR indoor OR air OR model) Przegląd literatury ograniczono
do prac polskich i anglojęzycznych z ostatnich 10 lat, czyli od roku 2007 do roku 2017.
1 [email protected], Samodzielna Katedra Biotechnologii i Biologii Molekularnej,
Wydział Przyrodniczo-Techniczny, Uniwersytet Opolski, www.uniopole.pl 2 [email protected], Samodzielna Katedra Biotechnologii i Biologii Molekularnej,
Wydział Przyrodniczo-Techniczny, Uniwersytet Opolski, www.uniopole.pl

Magdalena Kręcidło, Teresa Krzyśko-Łupicka
172
Do zarządzania źródłami bibliograficznymi użyto programu Mendeley Descop (wersji
1.17.6). Selekcja publikacji przebiegała według następujących kryteriów:
Kryteria włączenia publikacji do monografii: badania dotyczyły rozprzestrze-
niania się bioaerozoli w powietrzu atmosferycznym lub wewnątrz budynków/
badania były związane z rozprzestrzenianiem się/ emisją bioaerozoli wewnątrz
budynku lub w powietrzu atmosferycznym
Kryteria wyłączenia publikacji z listy: badania nie dotyczyły bioaerozoli/ badania
były opublikowane w innym języku niż język angielski lub polski/ praca doty-
czyła przeglądu literatury/ praca dotyczyła wirusów/ praca dotyczyła dezynfekcji
i metody oczyszczania powietrza
Z monografii wyłączono publikacje dotyczące możliwych dróg rozprzestrzeniania
się chorób wirusowych. Problem związany z rozprzestrzenianiem się chorób wiru-
sowych drogą kropelkową stanowi bardzo szerokie zagadnienie, a celem pracy nie
było przedstawienie danych epidemiologicznych, ani modeli rozprzestrzeniania się
epidemii.
3. Wyniki
3.1. Analiza bibliometryczna
Po wpisaniu wybranej kategorii pojęciowej w bazie danych PUBMED (dostęp na
28.03.2017) znaleziono 767 prac, z czego 370 mieściło się w założonych w metodyce
latach 2017-2007. Od 2007 roku liczba prac dotyczących bioaerozoli mikrobiolo-
gicznych rosła. Najwięcej, bo 50 prac badawczych opublikowano w 2015 roku, nato-
miast najmniej w roku 2007 (n=18). 52% wszystkich publikacji stanowiły prace
badawcze związane z bioaerozolem mikrobiologicznym. 16% prac dotyczyła roz-
przestrzeniania się bioaerozoli oraz źródeł ich emisji, a kolejne 36 % artykułów
traktowała o występowaniu, różnorodności i liczebności mikroorganizmów w bioa-
erozolach pozyskiwanych z różnych źródeł zarówno wewnątrz budynków jak i w po-
wietrzu atmosferycznym (Tabela 1.). Najwięcej uwagi naukowcy poświęcali zagad-
nieniom związanym z powietrzem atmosferycznym, budynkami użyteczności pub-
licznej, przemysłem spożywczym i rolnictwem. Wśród budynków użyteczności pub-
licznej najczęściej badane były pomieszczenia takie jak: szkoły i uniwersytety,
szpitale, ośrodki opieki zdrowotnej oraz biblioteki i archiwa [5-12] Prace związane
z przemysłem spożywczym i rolnictwem najczęściej dotyczyły zakładów mięsnych,
mleczarni, hodowli bydła mlecznego oraz ferm i zakładów przetwórstwa drobiu [4, 12-
16]. W przypadku powietrza atmosferycznego najczęściej badane i porównywane byłt
składy bioaerozoli pomiędzy obszarem rolniczym, a zurbanizowanym [17, 18].
Różnorodność mikroorganizmów w bioaerozolu pochodzącym z różnych miast [16,
19-21]. Poza tym badania dotyczyły również bioaerozoli emitowanych z mórz i oce-
anów, czy dróg przenoszenia się kurzu biologicznego znad Sahary [22, 23]. Najwięcej
prac opisujących źródła emisji bioaerozoli dotyczyło kompostowni, składowisk
odpadów i oczyszczalni ścieków [24, 25].
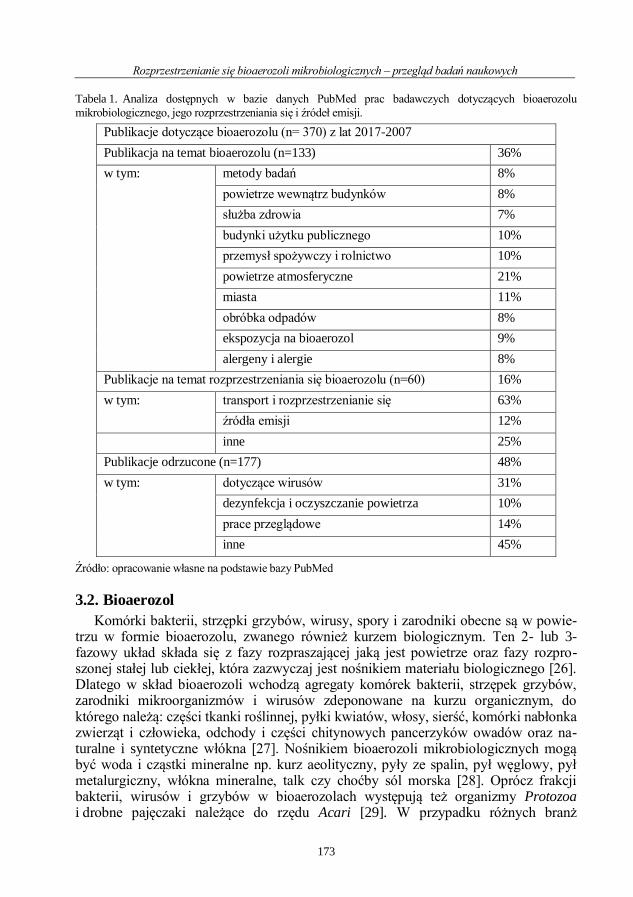
Rozprzestrzenianie się bioaerozoli mikrobiologicznych – przegląd badań naukowych
173
Tabela 1. Analiza dostępnych w bazie danych PubMed prac badawczych dotyczących bioaerozolu
mikrobiologicznego, jego rozprzestrzeniania się i źródeł emisji.
Publikacje dotyczące bioaerozolu (n= 370) z lat 2017-2007
Publikacja na temat bioaerozolu (n=133) 36%
w tym: metody badań 8%
powietrze wewnątrz budynków 8%
służba zdrowia 7%
budynki użytku publicznego 10%
przemysł spożywczy i rolnictwo 10%
powietrze atmosferyczne 21%
miasta 11%
obróbka odpadów 8%
ekspozycja na bioaerozol 9%
alergeny i alergie 8%
Publikacje na temat rozprzestrzeniania się bioaerozolu (n=60) 16%
w tym: transport i rozprzestrzenianie się 63%
źródła emisji 12%
inne 25%
Publikacje odrzucone (n=177) 48%
w tym: dotyczące wirusów 31%
dezynfekcja i oczyszczanie powietrza 10%
prace przeglądowe 14%
inne 45%
Źródło: opracowanie własne na podstawie bazy PubMed
3.2. Bioaerozol
Komórki bakterii, strzępki grzybów, wirusy, spory i zarodniki obecne są w powie-trzu w formie bioaerozolu, zwanego również kurzem biologicznym. Ten 2- lub 3-fazowy układ składa się z fazy rozpraszającej jaką jest powietrze oraz fazy rozpro-szonej stałej lub ciekłej, która zazwyczaj jest nośnikiem materiału biologicznego [26]. Dlatego w skład bioaerozoli wchodzą agregaty komórek bakterii, strzępek grzybów, zarodniki mikroorganizmów i wirusów zdeponowane na kurzu organicznym, do którego należą: części tkanki roślinnej, pyłki kwiatów, włosy, sierść, komórki nabłonka zwierząt i człowieka, odchody i części chitynowych pancerzyków owadów oraz na-turalne i syntetyczne włókna [27]. Nośnikiem bioaerozoli mikrobiologicznych mogą być woda i cząstki mineralne np. kurz aeolityczny, pyły ze spalin, pył węglowy, pył metalurgiczny, włókna mineralne, talk czy choćby sól morska [28]. Oprócz frakcji bakterii, wirusów i grzybów w bioaerozolach występują też organizmy Protozoa i drobne pajęczaki należące do rzędu Acari [29]. W przypadku różnych branż
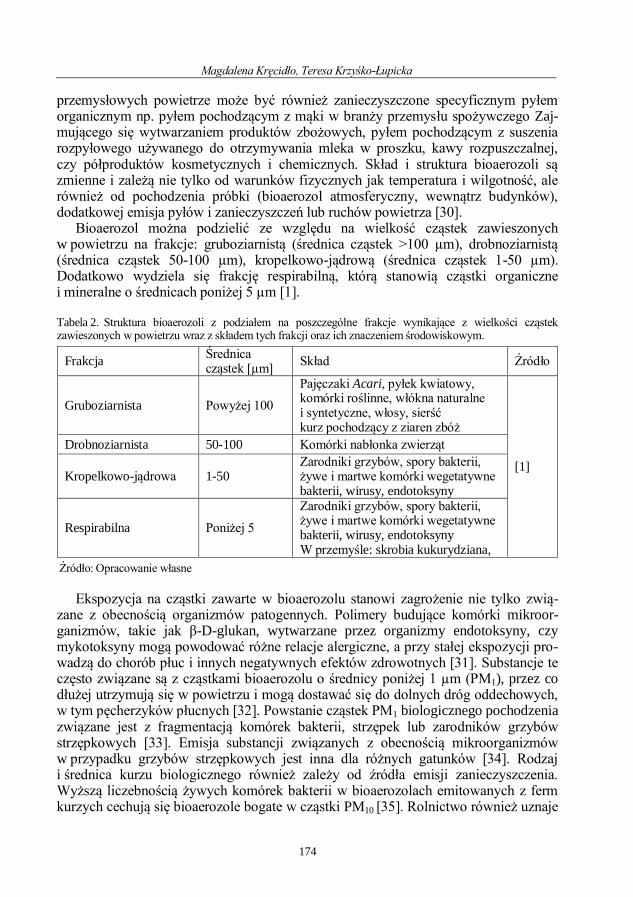
Magdalena Kręcidło, Teresa Krzyśko-Łupicka
174
przemysłowych powietrze może być również zanieczyszczone specyficznym pyłem organicznym np. pyłem pochodzącym z mąki w branży przemysłu spożywczego Zaj-mującego się wytwarzaniem produktów zbożowych, pyłem pochodzącym z suszenia rozpyłowego używanego do otrzymywania mleka w proszku, kawy rozpuszczalnej, czy półproduktów kosmetycznych i chemicznych. Skład i struktura bioaerozoli są zmienne i zależą nie tylko od warunków fizycznych jak temperatura i wilgotność, ale również od pochodzenia próbki (bioaerozol atmosferyczny, wewnątrz budynków), dodatkowej emisja pyłów i zanieczyszczeń lub ruchów powietrza [30].
Bioaerozol można podzielić ze względu na wielkość cząstek zawieszonych w powietrzu na frakcje: gruboziarnistą (średnica cząstek >100 µm), drobnoziarnistą (średnica cząstek 50-100 µm), kropelkowo-jądrową (średnica cząstek 1-50 µm). Dodatkowo wydziela się frakcję respirabilną, którą stanowią cząstki organiczne i mineralne o średnicach poniżej 5 µm [1].
Tabela 2. Struktura bioaerozoli z podziałem na poszczególne frakcje wynikające z wielkości cząstek zawieszonych w powietrzu wraz z składem tych frakcji oraz ich znaczeniem środowiskowym.
Frakcja Średnica cząstek [µm]
Skład Źródło
Gruboziarnista Powyżej 100
Pajęczaki Acari, pyłek kwiatowy, komórki roślinne, włókna naturalne i syntetyczne, włosy, sierść kurz pochodzący z ziaren zbóż
[1]
Drobnoziarnista 50-100 Komórki nabłonka zwierząt
Kropelkowo-jądrowa 1-50 Zarodniki grzybów, spory bakterii, żywe i martwe komórki wegetatywne bakterii, wirusy, endotoksyny
Respirabilna Poniżej 5
Zarodniki grzybów, spory bakterii, żywe i martwe komórki wegetatywne bakterii, wirusy, endotoksyny W przemyśle: skrobia kukurydziana,
Źródło: Opracowanie własne
Ekspozycja na cząstki zawarte w bioaerozolu stanowi zagrożenie nie tylko zwią-zane z obecnością organizmów patogennych. Polimery budujące komórki mikroor-ganizmów, takie jak β-D-glukan, wytwarzane przez organizmy endotoksyny, czy mykotoksyny mogą powodować różne relacje alergiczne, a przy stałej ekspozycji pro-wadzą do chorób płuc i innych negatywnych efektów zdrowotnych [31]. Substancje te często związane są z cząstkami bioaerozolu o średnicy poniżej 1 µm (PM1), przez co dłużej utrzymują się w powietrzu i mogą dostawać się do dolnych dróg oddechowych, w tym pęcherzyków płucnych [32]. Powstanie cząstek PM1 biologicznego pochodzenia związane jest z fragmentacją komórek bakterii, strzępek lub zarodników grzybów strzępkowych [33]. Emisja substancji związanych z obecnością mikroorganizmów w przypadku grzybów strzępkowych jest inna dla różnych gatunków [34]. Rodzaj i średnica kurzu biologicznego również zależy od źródła emisji zanieczyszczenia. Wyższą liczebnością żywych komórek bakterii w bioaerozolach emitowanych z ferm kurzych cechują się bioaerozole bogate w cząstki PM10 [35]. Rolnictwo również uznaje

Rozprzestrzenianie się bioaerozoli mikrobiologicznych – przegląd badań naukowych
175
się za branżę, w której najczęściej dochodzi do fragmentacji komórek mikroorga-nizmów i emisji pyłów PM1 i PM10. Związane jest to z przetwarzaniem i wykorzysty-waniem surowców roślinnych. W bioaerozolach pochodzenia rolniczego dominują grzyby strzępkowe, są one również bogatsze w β-D-glukan w przeciwieństwie do bioa-erozoli pochodzących z budynków mieszkalnych [36].
3.3. Źródła bioaerozoli
Przy badaniach dotyczących rozprzestrzeniania się bioaerozoli ważnym elementem jest odnalezienie źródła ich emisji. W zależności od tego, czy badania dotyczą mikro-klimatu pomieszczeń, czy powietrza atmosferycznego źródła emisji bioaerozolu są różne.
W powietrzu atmosferycznym źródła emisji bioaerozolu mikrobiologicznego mo-żemy podzielić na antropogeniczne i naturalne. Do tych pierwszych będą należeć aglomeracje miejskie, rolnictwo, sortownie i składowiska odpadów, kompostownie, punkty uzdatniania wody, oczyszczalnie ścieków, przemysł spożywczy, zwłaszcza zakłady mięsne [37]. Źródła bioaerozolu mikrobiologicznego związane z hodowlą zwierząt, czy utylizacją odpadów i recyklingiem mogą generować duże ilości mikro-organizmów patogennych. Promieniowce oraz Aspergillus fumigatus stanowią dominującą część mikrobioty emitowanej w bioaerozolu z różnych kompostowni [27, 38]. Zanieczyszczenie mikrobiologiczne emitowane z otwartych kompostowni może przenosić się na odległość do 300 m od miejsca emisji [37, 39]. Naturalnym źródłem zanieczyszczeń mikrobiologicznych może być kurz aeolityczny, zbiorniki wodne, opady deszczu, rośliny i zwierzęta [17].
Jednym z głównych źródeł bioaerozolu jest również człowiek. Czynności takie jak kaszlanie, rozmawianie czy oddychanie generują aerozol, w tym cząstki o średnicy poniżej 10 µm, które szybko rozprzestrzeniają się w powietrzu i uważane są za główny nośnik patogenów przenoszonych drogą kropelkową [1].
Długość pozostawania cząstki aerozolu wodnego, czy śluzu generowane przez człowieka i zwierzęta w powietrzu zależy nie tylko od ich średnicy, ale także od wilgotności względnej powietrza. W warunkach suchego powietrza (0% wilgotności względnej) generowany bioaerozol może ulegać ewaporacji zanim zdążył opaść na powierzchnię [40]. Podobną relację pomiędzy wilgotnością względną, wielkością cząstek, a trajektorią ich transportu w powietrzu potwierdził Nichol i współautorzy. Jednym z mechanizmów generowania bioaerozolu jest zaburzenie powierzchni między fazami [41].
3.4. Powietrze atmosferyczne
Oszacowanie składu bioaerozolu atmosferycznego jest dużym wyzwaniem dla naukowców, zwłaszcza, że najnowsze badania wskazują, że akumulacja cząstek biologicznych w powietrzu odgrywa ważną rolę środowiskową. Istnieje silna korelacja pomiędzy strukturą mikrobiologicznej frakcji bioaerozolu, a właściwościami fizyko-chemicznymi mas powietrza [42]. Kurz biologiczny odgrywa ważną rolę w zmianach klimatu, może wpływać na zdrowie ludzkie i produktywność rolnictwa. Ponad 33% kryształków lodu powstaje poprzez powstawanie biologicznych jąder kondensacji lodu INC (Ice Condensation Nuclei). Mikroorganizmy obecne w powietrzu atmosfer-

Magdalena Kręcidło, Teresa Krzyśko-Łupicka
176
rycznym mogą być również jądrami kondensacji chmur CNC (Cloud Condensation Nuclei). Niektórzy naukowcy uważają, że mogą również prowadzić aktywną biodegra-dację odgrywając ważna rolę w procesach zachodzących w atmosferze i zmieniając właściwości mas powietrza [42, 43]. Migracja mikrobiologicznej frakcji bioaerozolu wraz z masami powietrza jest bardzo poważnym problemem epidemiologicznym. Aeoliczny kurz znad Sahary może przenosić się wiele tysięcy kilometrów. Oprócz cząstek mineralnych zawiera również żywe komórki mikroorganizmów, w tym grzy-bów np. A, niger, A. fumigatus czy bakterii Staphylococcus gallinarum. Istnieją realne obawy, że wraz z kurzem biologicznym mogą przenosić się wirusy, bakterie i grzyby będące patogenami dla roślin, zwierząt i człowieka w rezultacie powodując roz-przestrzenianie się chorób [44]. Rozprzesrzenianie się zarodników grzybów strzęp-kowych w powietrzu wraz z wiatrem i ich różnorodność jest zależna od pory dnia, warunków meteorologicznych oraz sezonu [45, 46]. Liczebność i różnorodność mikroorganizmów w powietrzu atmosferycznym maleje w sezonie zimowym. Liczba Gram-dodatnich bakterii przetrwalnikujących rośnie w warunkach wysokiej tempe-ratury i niskiej wilgotności, co jest prawdopodobnie związane z dehydratacją obecnych na roślinach i w glebie bakterii, która indukuje wytwarzanie spor i ich rozproszenie się wraz z ruchami powietrza [47].
Różnorodność mikroorganizmów w bioaerozolach różnych regionów geograficz-nych jest zagadnieniem w dalszym ciągu mało poznanym. Podobnie jak czynniki ją determinujące [48]. Pomimo znacznych różnic w składzie bioaerozoli w różnych rejonach świata to można jednak wyznaczyć mikroorganizmy dominujące w powietrzu atmosferycznym. Wśród grzybów strzępkowych rodzajem najczęściej opisywanym w bioaerozolu powietrza atmosferycznego jest rodzaj Cladosporium [45, 49]. Natomiast bioaerozole pochodzące z oceanów cechują się wyższą liczebnością bakterii niż bioaerozol atmosferyczny znad lądów [50].
3.5. Powietrze wewnątrz budynków mieszkalnych i użytku publicznego
Badania różnorodności, struktury i składu mikrobiomu pomieszczeń oraz mikro-klimatu budynków cieszą się coraz większym zainteresowaniem. Na podstawie tego typu badań dotyczących mikroklimatu pomieszczeń szacuje się narażenie ludzi na szkodliwe dla zdrowia czynniki biologiczne. Powstają również normy mające na celu minimalizację narażenia pracowników, czy ludzi przebywających w danych warun-kach na szkodliwe dla zdrowia bioaerozole. Dotychczas jeszcze nie ustalono jak prze-bywanie w pomieszczeniu o odpowiednim mikrobiomie wpływa na zdrowie czło-wieka. Natomiast wiadomo, że nieodpowiednie warunki mikroklimatyczne pomiesz-czeń mogą prowadzić do negatywnych skutków zdrowotnych takich jak np. syndrom chorego budynku [51]. Pomiędzy przebywającymi w budynku ludźmi, a bioaerozolem występuje współzależność. Nie tylko mikroorganizmy obecne w powietrzu wpływają na zdrowie przebywających w pomieszczeniu ludzi, ale również obecność człowieka, jako źródła emisji bioaerozolu, kształtuje strukturę mikrobiomu pomieszczenia [3].

Rozprzestrzenianie się bioaerozoli mikrobiologicznych – przegląd badań naukowych
177
3.6. Powietrze w budynkach przemysłowych
W warunkach przemysłowych głównym generatorem bioaerozoli są zakłady prze-mysłu spożywczego. Związane jest to z wykorzystaniem i przetwarzaniem surowców roślinnych zawierających dużo biodegradowalnych węglowodanów, jak i surowców pochodzenia zwierzęcego bogatych w białko. W tej branży przemysłu zanieczysz-czenia mikrobiologiczne w produkcie gotowym mogą prowadzić do zatruć pokarmo-wych u konsumentów. Zachowanie bezpieczeństwa żywności wymaga od producen-tów monitorowania czystości mikrobiologicznej surowców, produktów i środowiska produkcji. Podobne obostrzenia dotyczą przemysłu farmaceutycznego i kosmetyce-nego. Branże te wymagają nie tylko wprowadzenia skutecznych działań dezynfekcyj-nych i prewencyjnych, ale również metod szybkiego wykrywania źródła emisji zanie-czyszczenia, które niejednokrotnie może nie pochodzić z surowców, od pracowników lub z procesu produkcji. Przykładem jest bioaerozol zawierający bakterie Listeria innacua generowany w zakładach przetwórstwa drobiu. Nieodpowiedni proces mycia powierzchni produkcyjnej może prowadzić do emisji aerozolu zwierającego bakterie L. innacua pochodzące z odpływu [52].
Emisja bioaerozoli w przemyśle spożywczym i rolnictwie stanowi problem nie tylko ze względu na możliwość kontaminacji żywności. Poważnym problem stanowi rozprzestrzenianie się genów antybiotykoodporności wśród bakterii. Wydawałoby się, że zagadnienie to dotyczy jedynie służby zdrowia, a także składowisk odpadów i oczyszczalni ścieków. Niestety istnieje coraz więcej doniesień na temat emisji multi-opornych szczepów bakterii Escherichia coli, Staphylococcus, czy Enterococcus przez przemysł spożywczy i rolnictwo, a zwłaszcza fermy drobiu i hodowle bydła [53-55]. 21,8% izolatów E. coli pochodzących z ferm bydła mlecznego wykazywała oporność na przynajmniej jeden z testowanych antybiotyków, a 14% z nich wykazywała fenotyp multioporności [56].
4. Podsumowanie i wnioski
Wraz z rozwojem metod analizy i próbkowania bioaerozoli oraz kurzu biolo-gicznego rośnie liczba badań dotyczących tych zagadnień. Niemniej struktura bioaerozoli oraz ich wspólne oddziaływanie z człowiekiem nie zostały jeszcze do końca poznane. Interesującym zagadnieniem jest wpływ bioaerozoli mikrobiolo-gicznych na zmiany klimatyczne, kształtowanie się chmur burzowych, czy opady śniegu. Przemieszczanie się mikroorganizmów wraz z prądami morskimi oraz skład i struktura bioaerozoli w poszczególnych strefach klimatycznych są coraz częściej opisywane (21% analizowanych prac). Problem dla naukowców stanowi analiza dróg rozprzestrzeniania się bioaerozoli oraz ocena źródeł ich emisji, gdyż ten typ prac stanowił jedynie 16 % ze wszystkich analizowanych artykułów. Ważną rolę wśród prac dotyczących kurzu biologicznego i mikroorganizmów zdeponowanych w po-wietrzu są doniesienia o negatywnych skutkach oddziaływania bioaerozoli oraz informacje o ekspozycji zawodowej. Za najbardziej problematyczne źródła emisji bioaerozoli uważa się przemysł spożywczy, rolnictwo, oczyszczalnie ścieków i skła-dowiska odpadów. Coraz więcej jest również niepokojących doniesień o rozprzestrze-nianiu się w powietrzu mikroorganizmów wykazujących fenotyp multioporności na leki.

Magdalena Kręcidło, Teresa Krzyśko-Łupicka
178
Literatura
1. Gralton J., Tovey E., McLaws ML. The role of particle size in aerosolised pathogen transmission, Journal of Infection., 62 (2011), s. 1-13.
2. Hsu NY., Lee CC., Wang JY., Li YC., Chang HW., Chen CY., Bornehag CG., Wu PC., Sundell J., Su HJ. Predicted risk of childhood allergy, asthma, and reported symptoms using measured phthalate exposure in dust and urine, Indoor Air., 22 (2012), s. 186-199.
3. Lax S., Smith DP., Hampton-Mercell J., Owens SM., Handley KM., Scott NM., Gibbons SM., Larsen P., Shogan BD., Weiss S., Metcalf JL., Ursell LK., Vazquez-Baeza Y., Van Treuren W., Hasan NA., Gibson MK., Colwell R., Dantas G., Knight R., Gilbert JA. Longitudinal analysis of microbial interaction between humans and the indoor environment, Science., 345 (2014), s. 1048-1052.
4. Brandl H., Fricker-Feer C., Ziegler D., Mandal J., Stephan R., Lehner A. Distribution and identification of culturale airborne microorganisms in a Swiss milk processing facility, Journal of Dairy Science., 97 (2014), s. 240-246.
5. Godwin C., Batterman S. Indoor air quality in Michigan schools, Indoor Air., 17 (2007), s. 109-121.
6. Aydogdu H., Asan A. Airborne fungi in child day care centers in Edirne City, Turkey, Environmental Monitoring and Assessment., 147 (2008), s. 423-44.
7. Aydogdu H., Asan A., Tatman Otkun M. Indoor and outdoor airborne bacteria in child day-care centers in Edirne City (Turkey), seasonal distribution and influence of meteorological factors, Environmental Monitoring and Assessment., 164 (2010), s. 53-66.
8. Harkawy A., Górny RL., Ogierman L., Wlazło A., Ławniczek-Wałczyk A., Niesler A. Bioaerosol assessment in naturally ventilated historical library building with restricted per-sonnel access, Annals of Agricultural and Environmental Medicine., 18 (2011), s. 323-329.
9. Chung FF., Lin HL., Liu HE., Lien AS., Hsiao HF., Chou LT., Wan GH. Aerosol distribution during open suctioning and long-term surveillance of air quality in a respiratory care center within a medical center, Respiratory Care., 60 (2015), s. 30-37.
10. Hospodsky D., Yamamoto N., Nazaroff WW., Miller D., Gorthala S., Peccia J. Characterizing airborne fungal and bacterial concentrations and emission rates in six occupied children's classrooms, Indoor Air., 25 (2015), s. 641-652.
11. Skóra J., Gutarowska B., Pielech-Przybylska K., Stępień Ł., Pietrzak K., Piotrowska M., Pietrowski P. Assessment of microbiological contamination in the work environments of museums, archives and libraries, Aerobiologia (Bologna)., 31 (2015), s. 389-401.
12. Diefenbach H., Hartung E., Sundrum A., Lohmeyer M., Rieger MA. Airborne biological hazards in different pig fattening systems, International Journal of Immunopathology and Pharmacology., 20 (2007), s. 45-49.
13. Abd-Elall AM., Mohamed ME., Awadallah MA. Potential airborne microbial hazards for workers on dairy and beef cattle farms in Egypt, Veterinaria Italiana., 45 (2009), s. 275-285.
14. Duan H., Chai T., Liu J., Zhang X., Qi C., Gao J., Wang Y., Cai Y., Miao Z., Yao M., Schlenker G. Source identification of airborne Escherichia coli of swine house surroundings using ERIC-PCR and REP-PCR, Environmental Research., 109 (2009), s. 511-517.
15. Berrang ME., Frank JF. Generation of airborne Listeria innocua from model floor drains, Journal of Food Protection., 75 (2012), s. 1328-31.
16. Gao M., Qiu T., Jia R., Han M., Song Y., Wang X. Concentration and size distribution of viable bioaerosols during non-haze and haze days in Beijing, Environmental Science and Pollution Research International., 22 (2015), s. 4359-4368.
17. Oliveira M., Ribeiro H., Delgado L., Fonseca J., Castel-Branco MG., Abreu I. Outdoor allergenic fungal spores: comparison between an urban and a rural area in northern Portu-gal, Journal of Investigational Allergology and Clinical Immunology., 20 (2010), s. 117-1128.

Rozprzestrzenianie się bioaerozoli mikrobiologicznych – przegląd badań naukowych
179
18. Lepers C., Dergham M., Armand L., Billet S., Verdin A., Andre V., Pottier D., Courcot D., Shirali P., Sichel F. Mutagenicity and clastogenicity of native airborne particulate matter samples collected under industrial, urban or rural influence, Toxicology in Vitro., 28 (2014), s. 866-874.
19. Ceter T., Pinar NM. Atmospheric concentration of fungus spores in Ankara and the effect of meteorological factors in 2003 period, Microbiological Biulletin., 43 (2009), s. 627-638.
20. Erkara IP., Ilhan S., Oner S. Monitoring and assessment of airborne Cladosporium Link and Alternaria Nées spores in Sivrihisar (Eskisehir), Turkey, Environmental Monitoring and Assessment., 148 (2009), s. 477-84.
21. Cao C., Jiang W., Wang B., Fang J., Lang J., Tian G., Jiang J., Zhu TF. Inhalable microorganisms in Beijing's PM2.5 and PM10 pollutants during a severe smog event, Environmental Science and Technology., 48 (2014), s. 1499-1507.
22. Cho BC., Hwang CY. Prokaryotic abundance and 16S rRNA gene sequences detected in marine aerosols on the East Sea (Korea), FEMS Microbiological Ecology., 76 (2011), s. 327-341.
23. Rosselli R., Fiamma M., Deligios M., Pintus G., Pellizzaro G., Canu A., Duce P., Squartini A., Muresu R., Cappuccinelli P. Microbial immigration across the Mediterranean via airborne dust, Scientific Reports., 5 (2015), s. 163-176.
24. Fischer G., Albrecht A., Jäckel U., Kämpfer P. Analysis of airborne microorganisms, MVOC and odour in the surrounding of composting facilities and implications for future investi-gations, International Journal of Hygiene and Environmental Health., 211 (2008), s. 132-142.
25. Han Y., Li L., Liu J. Characterization of the airborne bacteria community at different distances from the rotating brushes in a wastewater treatment plant by 16S rRNA gene clone libraries, Journal of Environmental Science (China)., 25 (2013), s 5-19.
26. Afanou KA., Straumfors A., Skogstad A., Nilsen T., Synnes O., Skaar I., Hjeljord L., Tronsmo A., Green BJ., Eduard W. Submicronic fungal bioaerosols: high-resolution microscopic characterization and quantification, Applied and Environmental Microbiology., 80 (2014), s. 7122-7130.
27. Kaarakainen P., Rintala H., Meklin T., Kärkkäinen P., Hyvärinen A., Nevalainen A. Concentrations and diversity of microbes from four local bioaerosol emission sources in Finland, Journal of the Air & Waste Management Association., 61 (2011), s. 370-372.
28. Li M., Qi J., Zhang H., Huang S., Li L., Gao D. Concentration and size distribution of bioaerosols in an outdoor environment in the Qingdao coastal region, Science of Total Environment., 409 (2011), s. 3812-3819.
29. Tovey ER., Liu-Brennan D., Garden FL., Oliver BG., Perzanowski MS., Marks GB. Time-Based Measurement of Personal Mite Allergen Bioaerosol Exposure over 24 Hour Periods, PLoS One., 18 (2016), s. 1-16.
30. Poole JA., Dooley GP., Saito R., Burrell AM., Bailey KL., Romberger DJ., Mehaffy J., Reynolds SJ. Muramic acid, endotoxin, 3-hydroxy fatty acids, and ergosterol content explain monocyte and epithelial cell inflammatory responses to agricultural dusts, Journal of Toxicology and Environmental Health., 73 (2010), s. 684-700.
31. Leleu C., Menotti J., Meneceur P., Choukri F., Sulahian A., Garin YJ., Denis JB., Derouin F. Bayesian development of a dose-response model for Aspergillus fumigatus and invasive aspergillosis, Risk Analysis., 33 (2013), s. 1441-1453.
32. Singh A., Kesavachandran CN., Kamal R., Bihari V., Ansari A., Azeez PA., Saxena PN., Ks AK., Khan AH. Indoor air pollution and its association with poor lung function, microalbuminuria and variations in blood pressure among kitchen workers in India: a cross-sectional study, Environmental Health., 16 (2017), s. 33-45.
33. Seo SC., Grinshpun SA., Iossifova Y., Schmechel D., Rao C., Reponen T. A New Field Compatible Methodology for the Collection and Analysis of Fungal Fragments, Aerosol Science and Technology., 41 (2007), s. 794-803.

Magdalena Kręcidło, Teresa Krzyśko-Łupicka
180
34. Seo SC., Reponen T., Grinshpun SA. Size fractionated (1→3)-β-D-glucan Concentrations Released from Different Moldy Materials, Science of Total Environment., 407 (2008), s. 806-814.
35. Zhao Y., Zhao D., Ma H., Liu K., Atilgan A., Xin H. Environmental assessment of three egg production systems - Part III: Airborne bacteria concentrations and emissions, Poultry Science., 95 (2016), s. 1473-1481.
36. Singh U., Levin L., Grinshpun SA., Schaffer C., Adhikari A., Reponen T. Influence of home characteristics on airborne and dustborne endotoxin and β-D-glucan, Journal Environmental Monitoring., 13 (2011), s. 3246-3253.
37. Kaarakainen P., Meklin T., Rintala H., Hyvärinen A., Kärkkäinen P., Vepsalainen A., Hirvonen MR., Nevalainen A. Seasonal Variation in Airborne Microbial Concentrations and Diversity at Landfill, Urban and Rural Sites Clean., 36 (2008), s. 556-563.
38. Taha MPM., Drew GH., Longhurst PJ., Smith R., Pollard SJT. Bioaerosol Releases from Compost Facilities: Evaluating Passive and Active Source Terms at a Green Waste Facility for Improved Risk Assessments, Atmospheric Environment., 40 (2006), s. 1159-1169.
39. Grisoli P., Rodolfi M., Villani S., Grignani E., Cottica D., Berri A., Picco AM., Dacarro C. Assessment of Airborne Microorganism Contamination in an Industrial Area Characterized by An Open Composting Facility and a Wastewater Treatment Plant, Environental Research., 109 (2009), s. 135-142.
40. Nicas M., Nazaroff WW., Hubbard A. Toward understanding the risk of secondary airborne infection: emission of respirable pathogens, Journal of Occupational and Environmental Hygiene., 2 (2005), s.143-154.
41. Nichol K., Bigelow P., O’Brien-Pallas L., McGeer A., Manno M., Holness DL. The individual, environmental, and organizational factors that influence nurses’ use of facial protection to prevent occupational transmission of communicable respiratory illness in acute care hospitals, American Journal of Infection Control., 36 (2008), s. 481-487.
42. Yoo K., Lee TK., Choi EJ., Yang J., Shukla SK., Hwang SI., Park J. Molecular approaches for the detection and monitoring of microbial communities in bioaerosols: A review, Journal of Environmental Science (China)., 51 (2017), s. 234-247.
43. Šantl-Temkiv T., Finster K., Dittmar T., Hansen BM., Thyrhaug R., Nielsen NW., Karlson UG. Hailstones: a window into the microbial and chemical inventory of a storm cloud, PLoS One., 8 (2013), s. 1-7.
44. Joung YS., Ge Z., Buie CR. Bioaerosol generation by raindrops on soil, Nature Communications., 7-8 (2017), s. 1-10.
45. Savage D., Barbetti MJ., MacLeod WJ., Salam MU., Renton M. Seasonal and diurnal patterns of spore release can significantly affect the proportion of spores expected to undergo long-distance dispersal, Microbial Ecology., 63 (2012), s. 578-585.
46. Pashley CH., Fairs A., Free RC., Wardlaw AJ. DNA analysis of outdoor air reveals a high degree of fungal diversity, temporal variability, and genera not seen by spore morphology, Fungal Biolgy., 116 (2012), s. 214-224.
47. Brodie EL., DeSantis TZ., Parker JPM., Zubietta IX., Piceno YM., Andersen GL. Urban aerosols harbor diverse and dynamic bacterial populations, Proceedings of the National Academy of Sciences USA., 104 (2007), s. 299-304.
48. Womack AM., Bohannan BJM., Green JL. Biodiversity and biogeography of the atmosphere, Philosophical Transactions of the Royal Society B: Biological Sciences., 365 (2010), s. 129-145.
49. Oliveira M., Ribeiro H., Delgado JL., Abreu I. The effects of meteorological factors on airborne fungal spore concentration in two areas differing in urbanisation level, International Journal of Biometeorology., 53 (2009), s. 61-73.
50. Seifried JS., Wichels A., Gerdts G. Spatial distribution of marine airborne bacterial communities, Microbiology Open., 4 (2015), s. 475-490.

Rozprzestrzenianie się bioaerozoli mikrobiologicznych – przegląd badań naukowych
181
51. Fisk WJ., Lei-Gomez Q., Mendell MJ. Meta-analyses of the associations of respiratory health effects with dampness and mold in homes, Indoor Air., 17 (2007), s. 284-296.
52. Berrang ME., Frank JF. Generation of airborne Listeria innocua from model floor drains, Journal of Food Protection., 75 (2012), s. 1328-1331.
53. Muzslay M., Moore G., Turton JF., Wilson AP. Dissemination of antibiotic-resistant enterococci within the ward environment: the role of airborne bacteria and the risk posed by unrecognized carriers, American Journal of Infection Control., 41 (2013), s. 57-60.
54. Liu D., Chai T., Xia X., Gao Y., Cai Y., Li X., Miao Z., Sun L., Hao H., Roesler U., Wang J. Formation and transmission of Staphylococcus aureus (including MRSA) aerosols carrying antibiotic-resistant genes in a poultry farming environment, Science of Total Environment., 426 (2012), s. 139-145.
55. Friese A., Schulz J., Zimmermann K., Tenhagen BA., Fetsch A., Hartung J., Rösler U. Occurrence of livestock-associated methicillin-resistant Staphylococcus aureus in Turkey and broiler barns and contamination of air and soil surfaces in their vicinity, Applied and Environmental Microbiology., 79 (2013), s. 2759-2766.
56. Navajas-Benito EV., Alonso CA., Sanz S., Olarte C., Martínez-Olarte R., Hidalgo-Sanz S., Somalo S., Torres C. Molecular characterization of antibiotic resistance in Escherichia coli strains from a dairy cattle farm and its surroundings, Journal of the Science of Food and Agriculture., 11 (2016), s. 362-365.
Rozprzestrzenianie się bioaerozoli mikrobiologicznych – przegląd badań naukowych
Streszczenie Powietrze jest jednym z ważniejszych ośrodków rozprzestrzeniania się zanieczyszczeń mikrobiologicz-nych. Komórki bakterii, strzępki grzybów, zarodniki i wirusy mogą być przenoszone przy udziale cząstek stałych, takich jak kurz, włosy, martwy naskórek lub w formie aerozolu złożonej z trójfazowej mieszaniny azeotropowej. Celem pracy jest zbiorcze przedstawienie dotychczasowych badań dotyczących źródeł bioaerozoli mikro-biologicznych oraz dróg jego rozprzestrzeniania. Ważnym elementem składającym się na poznanie składu mikrobioty zawieszonej w powietrzu jest ocena potencjalnych źródeł zanieczyszczenia mikrobiologicznego. W przypadku, gdy środowiskiem badań jest pomieszczenie zamknięte – źródłem mikroorganizmów mogą być zwierzęta, aktywność ludzi, systemu wentylacji, rośliny, żywność czy woda. W przypadku atmosfery obecność mikroorganizmów może być związana z mikrobiotą naturalnie występującą w powietrzu danego środowiska oraz zanieczyszczeniem mikrobiologicznym przeważnie o podłożu antropogenicznym. Słowa kluczowe: bioaerozol, zanieczyszczenia mikrobiologiczne, źródła, powietrze atmosferyczne, powietrze wewnątrz budynków
Dispersal of microbiological contaminations in the air – a review
Abstract The air is one of the most important sources of microbial spread. Bacterial cells, fungal mycelium, spores and viruses may be transportation by solid particles as dust, hairs, dead epidermis cells or in the form of solution composited from three-phases azeotrope mixture. The aim of the study was to perform the consolidated studies about researches about sources and disemission ways of the microbiological bioaerosols. The most important issue, which contribute on the knowledge about composition of airborne microbiota is assessment of potential emission sources. In the case of indoor environment the source of microorganism may be animals and human activity, ventilation systems, plants, food or water. Outdoor the sources of bioaerosols could be divide into the naturals and anthropogenic sources. Keywords: bioaerosol, microbial contamination, sources, outdoor air, indoor air

182
Ewa Brągoszewska1, Monika Rajtor
2, Józef S. Pastuszka
3
Porównanie wartości stężeń
oraz wyników identyfikacji aerozolu bakteryjnego,
otrzymanych przy użyciu dwóch różnych aspiratorów
1. Wprowadzenie
Pojęcie „bioaerozol” definiuje się jako układ dwufazowy, składający się z zawie-
szonych w powietrzu cząstek biologicznych, którymi mogą być np. wirusy, bakterie,
czy też grzyby mikroskopijne [1-3]. Najmniejszymi spośród biocząstek są wirusy,
których średnica osiąga od 0,02 µm do 0,3 µm. Średnica komórek bakteryjnych jest
większa i wynosi ona od 1 do 2 µm. Natomiast średnica grzybów mikroskopijnych
(w zależności od gatunku) od 2 do 8 µm [4-8].
Wyznaczenie stężeń bioaerozolu stanowi złożony proces, obejmujący między
innymi, zliczanie cząstek biologicznych w ustalonej objętości powietrza, oraz ich
identyfikację, ale zawsze pierwszym i podstawowym etapem jest pobór zawieszonych
w powietrzu biocząstek odpowiednio dobranym urządzeniem. Metody poboru aerozoli
biologicznych determinowane są przez ich fizyczne oraz biologiczne właściwości [9].
Zagadnienie poprawności pomiarów stężeń bioaerozoli zostało po raz pierwszy
podjęte w latach 1990-1991 na Uniwersytecie Cincinnati, w USA. Opracowane zostały
wówczas podstawowe kryteria pomiarowe wiążące poszczególne typy aspiratorów
z rodzajami mikroorganizmów oraz czas poboru z oczekiwanym stężeniem bioaerozoli
[3, 10].
Chociaż, w ocenie zanieczyszczenia mikrobiologicznego powietrza zaleca się
stosowanie metod wolumetrycznych, polegających na aktywnym pobieraniu próbki
powietrza o określonej objętości, to szereg laboratoriów stosuje wciąż jako metodę
podstawową w ocenie stężenia bioaerozoli – metodę Kocha [9, 11, 12]. Metoda ta jest
wykorzystywana ze względu na jej prostotę, szybkość oraz niski koszt badania. Polega
na obliczeniu stężeń bioaerozoli na podstawie pomiaru depozycji. Przeliczenie to
opiera się na założeniu, że na 100 cm² powierzchni pożywki agarowej, w ciągu 5 minut
osadza się taka ilość mikroorganizmów, jaka znajduje się w 10 dm³ powietrza. Jednak
nie istnieje żadna ścisła zależność fizyczna pomiędzy wielkością stężenia, a opadem
cząstek, dlatego też metoda Kocha jest z założenia niewłaściwa. Pobór cząstek
bioaerozolu metodą depozycji jest uzasadniony jedynie wtedy, gdy do oceny skażenia 1 [email protected], Katedra Ochrony Powietrza, Wydział Inżynierii Środowiska i Energetyki,
Politechnika Śląska, www.polsl.pl 2 [email protected], Katedra Mikrobiologii, Wydział Biologii i Ochrony Środowiska, Uniwersytet Śląski,
www.us.edu.pl 3 [email protected], Katedra Ochrony Powietrza, Wydział Inżynierii Środowiska i Energetyki,
Politechnika Śląska, www.polsl.pl

Porównanie wartości stężeń oraz wyników identyfikacji aerozolu bakteryjnego,
otrzymanych przy użyciu dwóch różnych aspiratorów
183
mikrobiologicznego powietrza wystarczająca jest informacja o poziomie opadu
bioaerozolu, lub wiedza o składzie gatunkowym mikroorganizmów [13].
Zatem, aby uzyskać informację dotyczącą stężenia aerozolu bakteryjnego i grzy-
bowego należy posługiwać się odpowiednio dobranym urządzeniem do aktywnego
poboru mikroorganizmów zawieszonych w powietrzu. Zdecydowana większość
przyrządów do wyznaczania stężeń bioaerozolu wykorzystuje wytrącanie biocząstek
z pobranej objętości powietrza na skutek oddziaływania na strugę powietrza siły
zewnętrznej, zwykle siły bezwładności. Wytrącane cząstki biologiczne osadzają się na
powierzchni odpowiednio dobranego podłoża mikrobiologicznego, rozlanego wcześniej
do szalek Petriego, gdzie po określonym czasie inkubacji rozrastają się w kolonie, a ich
ilość dzielona jest przez objętość powietrza pobranego przez impaktor.
2. Cel pracy
Celem niniejszej pracy było porównanie wyników przeprowadzonej analizy
ilościowej i jakościowej bakterii tworzących kolonie na pożywkach stałych, pobranych
przy użyciu dwóch różnych impaktorów: jednostopniowego Air Ideal (bioMérieux
Marcy-l'Etoile, France) oraz referencyjnego 6-stopniowego kaskadowego impaktora
typu Andersena (Tisch Environmental, USA).
3. Metodyka badań
Pomiary zostały przeprowadzone w okresie marzec-kwiecień w centrum Gliwic, na
terenie kampusu Politechniki Śląskiej (50°17'37.1"N 18°40'54.9"E). Gliwice to miasto
położone w południowej Polsce, w województwie śląskim, na zachodnim krańcu
Górnośląskiego Okręgu Przemysłowego.
Podczas pomiarów rejestrowano parametry meteorologiczne, przy użyciu
przenośnej stacji pogodowej (Oregon Scientific, USA) (tab.1).
Tabela 1. Parametry meteorologiczne mierzone w trakcie poboru próbek
Wybrany parametr meteorologiczny Średnia Min Max
Temperatura powietrza [°C] 14,2 11,1 18,8
Ciśnienie atmosferyczne [hPa] 1005,33 992,25 1010,92
Prędkość wiatru [m/s] 5,3 4,42 7,01
Wilgotność powietrza [%] 48,4 41 56
Źródło: Opracowanie własne
Do poboru próbek wykorzystano metodę wolumetryczną, która polegała na zasysaniu przez aspirator znanej objętości powietrza uderzającego z dużą prędkością w powierzchnię pożywek agarowych. W badaniach wykorzystano 6-stopniowy impaktor typu Andersena (Tisch Environmental, USA), który składa się z sześciu nakładanych na siebie segmentów/stopni (ze średnicami odcięcia: 7,0; 4,7; 3,3; 2,1; 1,1; 0,65 μm), posiadających coraz to mniejsze otwory wlotowe. Do impaktora dołączona jest pompa, która zapewnia stałe natężenie przepływu powietrza w trakcie pomiarów wynoszące 28,3 l/min. Pobór za pomocą impaktora Andersena trwał 10

Ewa Brągoszewska, Monika Rajtor, Józef S. Pastuszka
184
minut. Jest to optymalny czas poboru, gdyż po dłuższym okresie czasu następuje utwardzanie agaru na skutek wysychania jego powierzchni, a to z kolei powoduje zaniżenie wyznaczonego stężenia aerozolu bakteryjnego [3].
W badaniach posługiwano się także jednostopniowym impaktorem Air Ideal (bioMérieux Marcy-l'Etoile, France), który pobiera cząstki o średnicy od 3 μm. Przepływ powietrza w impaktorze wynosił 100 l/min, a prędkość zderzenia cząsteczek o powierzchnię podłoża 20 m/s. Czas trwania pomiaru impaktorem Air Ideal wynosił 3 min.
Mikroorganizmy pobierane były na powierzchnię pożywki agarowej rozlanej na szalkach Petriego. Podłożem odpowiednim dla wzrostu bakterii był agar odżywczy TSA, do którego dodawano cycloheksymid hamujący wzrost grzybów. Po poborze szalki Petriego zostały umieszczone w cieplarce (w temperaturze 22°C, 4-6 dni).
Następnie zliczano wyrosłe kolonie i wyznaczono stężenie, które dla stricte żywych bakterii jest niezwykle trudne do określenia. Jako wyróżnik stężenia bioaerozolu przyjęto ilość wyrosłych kolonii na podłożach stałych, odniesioną do jednostki objętości powietrza, jtk/m
3 (jednostki tworzące kolonie w 1m³ powietrza).
Przeprowadzono także identyfikację wyizolowanych szczepów bakterii. Bakterie identyfikowano do gatunku w oparciu o profil komórkowych kwasów tłuszczowych z wykorzystaniem systemu identyfikacji mikroorganizmów opracowanego przez firmę MIDI [14,15]. W celu izolacji kwasów tłuszczowych, 40 mg biomasy bakteryjnej uzyskanej z 24 godzinnej hodowli poddano saponifikacji. Zawiesinę komórek gotowano przez pół godziny w roztworze 13% NaOH w mieszaninie metanolu i wody (1:1, v:v), worteksując ją po pierwszych 10 min inkubacji we wrzącej łaźni wodnej. Uwolnione kwasy tłuszczowe poddano metylacji mieszaniną metanolu i 6N kwasu solnego (1:1, v:v), w 80°C przez 10 min, a następnie ekstrakcji mieszaniną heksanu i eteru metylowo-butylowego (1:1, v:v). Fazę organiczną przemyto 10% roztworem NaOH. Ekstrakty analizowano na chromatografie gazowym Hewlett Packard 6890, wyposażonym w kolumnę kapilarną Ultra 2. Gazem nośnym był azot. Analizę uzys-kanych profili tłuszczowych przeprowadzono w oparciu o bibliotekę TSBA40 bazy Sherlock 5.0 systemu identyfikacji organizmów (MIS – microbial identyfication system). Bakterie zaklasyfikowano do gatunku przy prawdopodobieństwie identy-fikacji (PI) >75%, natomiast przy PI >60%, do rodzaju. Wyniki o PI<60% odrzucono.
4. Analiza wyników badań
4.1. Analiza ilościowa
Średnie całkowite stężenie aerozolu bakteryjnego pobieranego 6-stopniowym impaktorem Andersena wynosiło 513 CFU/m
3. Minimalne stężenie bakterii pobie-
ranych tym urządzeniem było równe 230 CFU/m3, a maksymalne 869 CFU/m
3. Przy
czym, średnie stężenie frakcji respirabilnej (cząstki o średnicy aerodynamicznej dae<3,3μm) wynosiło 107 CFU/m
3, co stanowiło aż 20% średniego stężenia całko-
witego. Z kolei, średnie stężenie bakterii w powietrzu atmosferycznym otrzymane przy
użyciu jednostopniowego aspiratora Air Ideal wynosiło 293 CFU/m3, było zatem
prawie o dwa razy mniejsze od tego, które otrzymano przy użyciu impaktora Ander-sena. Wyniki przedstawiono w tabeli nr 2.

Porównanie wartości stężeń oraz wyników identyfikacji aerozolu bakteryjnego,
otrzymanych przy użyciu dwóch różnych aspiratorów
185
Tabela 2. Stężenia aerozolu bakteryjnego w Gliwicach
Całkowite stężenie aerozolu
bakteryjnegooraz jego dwóch frakcji
rozmiarowych w CFU/m³
Andersen Air
Ideal
Frakcja
respirabilna
(dae<3,3μm)
Frakcja
gruba
(dae>3,3μm)
Średnia 513 293 107 407
Minimum 230 143 46 169
Maximum 869 607 198 749
Odchylenie standardowe 190 140 49 193
Źródło: Opracowanie własne, gdzie dae – średnica aerodynamiczna
Wartości stężeń aerozolu bakteryjnego pobranego przy wykorzystaniu impaktora Andersena są prawie dwukrotnie wyższe od stężeń otrzymanych przy użyciu impak-tora Air Ideal, co z pewnością spowodowane jest znacznie większą średnicą odcięcia (średnicą graniczną) tego drugiego urządzenia w stosunku do impaktora Andersena. Wartości te wynoszą odpowiednio 3 μm (Air Ideal) i 0,65 μm (impaktor Andersena). Oznacza to, że cząstek bakteryjnych w zakresie średnic 0,65-3 μm impaktor Air Ideal nie wychwytuje. Dane te są zgodne z wynikami uzyskanymi w Gliwicach w latach 2011-2015 [12, 16, 17].
Uzyskane wyniki wskazują, że jednostopniowy impaktor Air Ideal można bez ograniczeń używać raczej w badaniach skriningowych. Należy podkreślić, że impaktor ten jest bardzo lekki, wygodny w obsłudze, posiada własne zasilanie i może być auto-matycznie włączany (wyłącza się zawsze samoczynnie).
Udział frakcji respirabilnej bioaerozolu (cząstki o średnicy aerodynamicznej dae<3,3μm) może osiągać wartość aż do 40% stężenia całkowitego. Zatem, jeśli nie znany jest rozkład ziarnowy bioaerozolu w środowisku, a jego stężenie całkowite powinno być wyznaczone z dużą dokładnością, to do pomiarów należy wybrać aspi-rator o średnicy odcięcia nie większej niż 1 µm [5].
4.2. Analiza jakościowa
Najpowszechniejsze grupy bakterii stanowiły bakterie Gram-dodatnie tworzące endospory (5 gatunków), reprezentowane głównie przez laseczki tlenowe z rodzaju Bacillus, a także Gram-dodatnie ziarniaki z rodzaju Micrococcus, z Micococcus luteus jako ich najczęstszym przedstawicielem. Są to mikroorganizmy zazwyczaj licznie występujące w bioaerozolu, w związku z ich powszechnym występowaniem w przy-rodzie i wysoką opornością na zewnętrzne czynniki fizykochemiczne. U Bacillus, oporność ta wynika z umiejętności wytwarzania przetrwalników, natomiast u Micro-coccus jest związana z produkcją barwników karotenoidowych, które chronią komórki przed szkodliwym wpływem promieniowania UV. Wyniki te są zgodne z wynikami badań prowadzonymi w latach 2011-2016 w Gliwicach, podczas których przepro-wadzona została identyfikacja bakterii zawieszonych w powietrzu atmosferycznym, przy wykorzystaniu metody biochemicznej, tj. testów API [16-19].
Jednakże, przeprowadzona identyfikacja aerozolu bakteryjnego występującego w powietrzu atmosferycznym Gliwic, wskazuje większą różnorodność wśród identyfi-kowanych bakterii w próbach pobranych przy użyciu impaktora Andersena (10 gatunków), w porównaniu do Air Ideal (5 gatunków). Wyniki przedstawiono w tabeli nr 3.

Ewa Brągoszewska, Monika Rajtor, Józef S. Pastuszka
186
Tabela 3. Jakościowa charakterystyka aerozolu bakteryjnego pobranego przy użyciu dwóch urządzeń
Zidentyfikowane gatunki/rodzaje bakterii Impaktor Andersena Air Ideal
Micrococcus luteus + +
Micrococcus lylae + -
Micrococcus sp. + -
Rhodococcus sp. + -
Kocuria kristinae + -
Kocuria sp. + -
Staphylococcus sp. - +
Bacillus cereus + -
Bacillus megaterium + -
Bacillus pumilus + -
Bacillus sp. + +
Virgibacillus sp. + -
Paenibacillus pabuli - +
Paenibacillus sp. + +
Microbacterium luteolum + -
Microbacterium lacticum - +
Microbacterium sp. - +
Mycobacterium sp. + -
Corynebacterium callunae + -
Nocardia brasiliensis + -
Nocardia sp. - +
Arthrobacter sp. + -
Yersinia frederiksenii + -
Cellulomonas sp. + -
Pauciomonas sp. + -
Pseudomonas agarici - +
Pseudomonas putida - +
Źródło: Opracowanie własne, gdzie + zidentyfikowano; - nie zidentyfikowano

Porównanie wartości stężeń oraz wyników identyfikacji aerozolu bakteryjnego,
otrzymanych przy użyciu dwóch różnych aspiratorów
187
5. Podsumowanie
Średnie stężenie bakterii tworzących kolonie na pożywkach stałych, otrzymane
przy użyciu jednostopniowego impaktora Air Ideal było rzędu 3x10² CFU/m³
i było ono prawie dwa razy mniejsze od średniego stężenia aerozolu bakteryjnego
otrzymanego przy użyciu 6-stopniowego impaktora Andersena (5x10²). Jest to
z pewnością spowodowane znacznie większą średnicą odcięcia Air Ideal,
w stosunku do impaktora Andersena. Wartości te wynoszą odpowiednio 3 μm
(Air Ideal) i 0,65 μm (impaktor Andersena). Oznacza to, że cząstek bakteryjnych
w zakresie średnic od 0,65 do 3 μm impaktor Air Ideal nie wychwytuje.
Wykazano większą różnorodność wśród identyfikowanych bakterii w próbach
pobranych przy użyciu 6-stopniowego impaktora Andersena (10 gatunków), niż
w próbach pobranych przy użyciu 1-stopniowego Air Ideal (5 gatunków).
Jeśli nie znany jest rozkład ziarnowy bioaerozolu, a jego stężenie całkowite
powinno być wyznaczone z dużą dokładnością, to do pomiarów należy wybrać
aspirator o średnicy odcięcia nie większej niż 1 µm.
Podziękowania
Praca została wykonana w ramach grantu BKM/533/RIE-2/2016 finansowanego
przez Ministerstwo Nauki i Szkolnictwa Wyższego.
Literatura
1. Douwes, J., Thorne, P., Pearce, N., Heederik, D. Bioaerosol health effects and exposure
assessment: Progress and prospects, Annals of Occupational Hygiene, 3 (2003), s. 187-200.
2. Haas, D., Unteregger, M., Habib, J., Galler, H., Marth, E., Reinthaler, F. F. Exposure to
bioaerosol from sewage systems, Water, Air, & Soil Pollution, 207 (2010), s. 49-56.
3. Nevalainen, A., Willeke, K., Liebhaber, F., Pastuszka, J. S., Burge, H., Henningson, E.
Bioaerosol sampling, Aerosol Measurement: Principles, Techniques, and Applications,
Willeke, K., Baron, P., Red., Van Nostrand Reinhold, New York (1993), s. 471-492.
4. Kołwzan, B., Adamiak, W., Grabas, K., Adam Pawełczyk Podstawy mikrobiologii
w ochronie środowiska, Oficyna Wydawnicza Politechniki Wrocławskiej, Wrocław (2005).
5. Pastuszka, J. Narażenie na aerozole ziarniste, włókniste i biologiczne (bakterie i grzyby
mikroskopijne) populacji generalnej Górnośląskiego Okręgu Przemysłowego, Oficyna
Wydawnicza Politechniki Wrocławskiej, Wrocław (2001).
6. Walser, S. M., Gerstner, D. G., Brenner, B., Bünger, J., Eikmann, T., Janssen, B., Kolb, S.,
Kolk, A., Nowak, D., Raulf, M., Sagunski, H., Sedlmaier, N., Suchenwirth, R., Wiesmüller,
G., Wollin, K. M., Tesseraux, I., Herr, C. E. W. Evaluation of exposure-response
relationships for health effects of microbial bioaerosols - A systematic review, International
Journal of Hygiene and Environmental Health, 218 (2015), s. 577-589.
7. Nasir, Z. A., Colbeck, I. Assessment of bacterial and fungal aerosol in different residential
settings, Water, Air, & Soil Pollution, 211 (2010), s. 367-377.
8. Szewczyk, E. Ćwiczenia z mikrobiologii dla studentów kosmetologii, Uniwersytet
Medyczny w Łodzi, Łódź (2014).
9. Gołofit-Szymczak, M., Ławniczek-Walczyk, A., Górny, R. Ilościowa i jakościowa kontrola
szkodliwych czynników biologicznych w środowisku pracy, Podstawy i Metody Oceny
Środowiska Pracy, 2 (2013), s. 5-17.

Ewa Brągoszewska, Monika Rajtor, Józef S. Pastuszka
188
10. Nevalainen, A., Pastuszka, J., Liebhaber, F., Willeke, K. Performance of bioaerosol
samplers: collection characteristics and sampler design considerations, Atmospheric
Environment, Part A, General Topics, 26 (1992), s. 531-540.
11. Górny, R. L. Biological aerosols – a role of hygienic standards in the protection of
environment and health, Medycyna Środowiskowa, 13 (2010), s. 41-51.
12. Pastuszka, J. S., Iwasiewicz, P., Brągoszewska, E. Preliminary testing of a new bioaerosol
sampler developed for the measurements of low and medium concentration levels,
Environment Protection Engineering, 39 (2013), s. 129-138.
13. Canha, N., Almeida, S. M., Freitas, M. D. C., Wolterbeek, H. T. Assessment of bioaerosols
in urban and rural primary schools using passive and active sampling methodologies,
Archives of Environmental Protection, 41 (2015), s. 11-22.
14. Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids,
Technical Note, 101 (2001), s. 1-6.
15. Frostegard, A., Baath, E. The use of phospholipid fatty acid analysis to estimate bacterial
and fungal biomass in soil, Biology and Fertility of Soils, 22 (1996), s. 59-65.
16. Brągoszewska, E., Kowalski, M., Pastuszka, J. S. Narażenie ludzi na aerozol bakteryjny
występujący w powietrzu atmosferycznym Gliwic w okresie wiosennym, Janiszewska, M.,
Bilska, S., Red., Biomedyczny przegląd naukowy, 1 (2016), s. 305-314.
17. Brągoszewska, E. Aerozol bakteryjny w powietrzu atmosferycznym Gliwic i jego udział
w całkowitym narażeniu ludzi na bakterie wchłaniane drogą inhalacyjną, Rozprawa
Doktorska, Gliwice (2014).
18. Brągoszewska, E., Mainka, A., Pastuszka, J. S. Bacterial and Fungal Aerosols in Rural
Nursery Schools in Southern Poland, Atmosphere, 7 (2016).
19. Brągoszewska, E., Mainka, A., Pastuszka, J. S. Bacterial aerosols in an urban nursery
school in Gliwice, Poland: a case study, Aerobiologia, 32 (2016), s. 469-480.
Porównanie wartości stężeń oraz wyników identyfikacji aerozolu bakteryjnego,
otrzymanych przy użyciu dwóch różnych aspiratorów
Streszczenie
Do oceny stężenia bioaerozoli stosuje się różnorodne aspiratory oraz metody poboru, co skutkuje tym, że
dla identycznych środowisk i porównywalnych warunków pomiarów otrzymywane są bardzo różniące się od siebie wyniki. Celem niniejszej pracy było porównanie wyników przeprowadzonej analizy ilościowej
i jakościowej bakterii tworzących kolonie na pożywkach stałych. Aerozol bakteryjny pobierany był
równocześnie przy użyciu dwóch różnych impaktorów: 1-stopniowego aspiratora cząstek biologicznych
Air Ideal (bioMérieux) oraz 6-stopniowego impaktora typu Andersena (Tisch Environmental, USA). Bio-aerozol pobierany był z powietrza atmosferycznego w okresie marzec-kwiecień 2016 r., na terenie kampusu
Politechniki Śląskiej w Gliwicach. Wykazano, że średni poziom stężeń aerozolu bakteryjnego otrzymany
przy pomocy impaktora Andersena był prawie dwa razy wyższy od średniego stężenia bakterii otrzy-
manego przy użyciu jednostopniowego impaktora Air Ideal. Spowodowane jest to prawdopodobnie zna-cznie większą średnicą odcięcia (średnicą graniczną) tego drugiego urządzenia w stosunku do impaktora
Andersena. Wartości te wynoszą odpowiednio 0,65 μm (impaktor Andersena) i 3 μm (Air Ideal). Oznacza
to, że cząstek bakteryjnych w zakresie średnic od 0,65 do 3 μm jednostopniowy impaktor Air Ideal nie
wychwytuje. Przeprowadzono także identyfikację gatunkową wybranych prób, która wykazała, iż większa różnorodność wśród identyfikowanych bakterii występowała w próbach pobranych przy użyciu impaktora
Andersena.
Słowa kluczowe: bioaerozol, powietrze atmosferyczne, aerozol bakteryjny, identyfikacja bakterii

Porównanie wartości stężeń oraz wyników identyfikacji aerozolu bakteryjnego,
otrzymanych przy użyciu dwóch różnych aspiratorów
189
Comparison of the results of concentrations and identification of bacterial
aerosol, obtained using by two different aspirators
Abstract To evaluate the concentration of bioaerosols by various aspirators and collection methods resulting in very
different results for the same environments and comparable measurement conditions. The purpose of this
work was to compare the quantitative and qualitative results of airborne bacteria collected simultaneously
using by two different impactors: single-stage biological particle impactor Air Ideal (bioMérieux, France) and six-stage Andersen cascade impactor (Tisch Environmental, USA). Bioaerosol was collected from the
atmospheric air in March-April 2016, on the campus of the Silesian University of Technology in Gliwice.
The average level of bacterial aerosol obtained by the Andersen impactor was nearly two times higher than
the average bacterial concentration achieved with the single-stage Air Ideal impactor. This is the probably of the larger cut-off diameter of the second device in relation to the Andersen impactor. These values are
respectively 0.65 μm (Andersen Impactor) and 3 μm (Air Ideal). This means that the bacterial particles in
the range from 0.65 to 3 μm single-stage Air Ideal do not collect. The species identification of selected
samples was also performed, which showed that greater diversity among the identified bacteria found in samples using by Anderson impactor.
Keywords: bioaerosol, atmospheric air, bacteria aerosol, bacteria identification

190
Sylwia Magdalena Duda1, Monika Garbacz
2
Ocena aktywności mikroorganizmów
z wykorzystaniem matrycy czujników gazu
1. Wprowadzenie
Działalność mikroorganizmów jest nierozerwalnie związana z gospodarką wodno-
ściekową. Jednym z najszerzej stosowanych czynników procesowych w oczyszczal-
niach ścieków na całym świecie jest osad czynny, czyli kłaczkowata zawiesina
zawierająca różne gatunki organizmów żywych, pełniących określoną rolę w procesie
oczyszczania ścieków [1, 2].
Eksploatatorzy oczyszczalni oceniają procesy zachodzące w poszczególnych
urządzeniach na podstawie parametrów fizykochemicznych ścieków. Zalicza się do
nich głownie: biologiczne zapotrzebowanie na tlen (BZT), chemiczne zapotrzebowanie
na tlen (ChZT), zawiesinę ogólną (Zog) oraz związki azotu i fosforu. W literaturze
omówione zostały już zależności pomiędzy wysokością wskaźników zanieczyszczeń
a uciążliwością zapachową badanego medium [3-5].
Omawiane obiekty są jednocześnie jednymi z najbardziej istotnych emiterów
związków zapachowo-czynnych do atmosfery na wszystkich etapach pracy. Identy-
fikacja i określenie stężenia związków uciążliwych zapachowo jest trudne. Uciążli-
wość zapachową często analizuje się z wykorzystaniem olfaktometrii dynamicznej.
Metoda te jednak nie jest pozbawiona wad, dlatego też co raz częściej wykorzystywane
są matryce wieloczujnikowe, które razem z innymi elementami współtworzą matryce
wieloczujnikowe, czyli imitację ludzkiego zmysłu. Nie brakuje doniesień o stosowaniu
urządzeń tego typu w obiektach inżynierskich służących oczyszczaniu ścieków [4].
Celem niniejszej pracy jest omówienie możliwej korelacji pomiędzy wskazaniami
matrycy czujników a klasycznie określanym parametrem ścieków, czyli BZT5 [5-8].
Na podstawie biochemicznego zapotrzebowania na tlen można szacować aktywność
mikroorganizmów. Wielkość zużycia tlenu może informować o zachodzących
procesach, a także ich intensywności.
2. Biochemiczne zapotrzebowanie na tlen
W ściekach występuje zazwyczaj wiele substancji organicznych, dlatego też byłaby
niemożliwa klasyfikacja każdej z osobna. W praktyce określa się grupy substancji.
Związki organiczne zawarte w omawianym medium oznacza się za pomocą ilości
zużytego tlenu – mówimy wtedy o wyznaczaniu BZT lub ChZT (Chemiczne Zapo-
1 [email protected], SKN FOR&AGAINST, Wydział Inżynierii Środowiska, Politechnika Lubelska, ul. Nadbystrzycka 40B, Lublin 20-618, tel: +48 815384784 2 [email protected], Koło Młodych Inżynierów PZiTS, Wydział Inżynierii Środowiska, Politechnika
Lubelska, ul. Nadbystrzycka 40B, Lublin 20-618, tel: +48 815384784
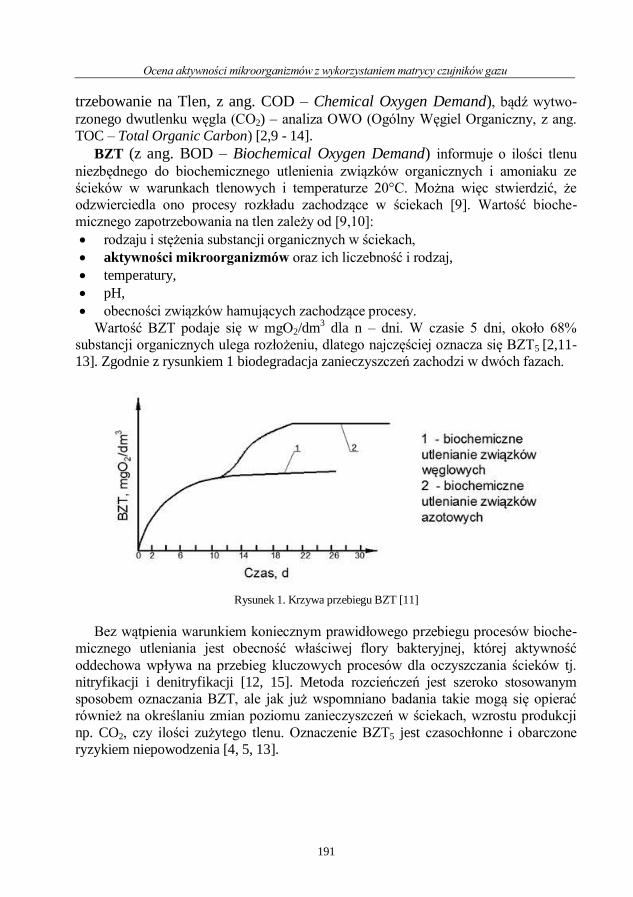
Ocena aktywności mikroorganizmów z wykorzystaniem matrycy czujników gazu
191
trzebowanie na Tlen, z ang. COD – Chemical Oxygen Demand), bądź wytwo-
rzonego dwutlenku węgla (CO2) – analiza OWO (Ogólny Węgiel Organiczny, z ang.
TOC – Total Organic Carbon) [2,9 - 14].
BZT (z ang. BOD – Biochemical Oxygen Demand) informuje o ilości tlenu
niezbędnego do biochemicznego utlenienia związków organicznych i amoniaku ze
ścieków w warunkach tlenowych i temperaturze 20°C. Można więc stwierdzić, że
odzwierciedla ono procesy rozkładu zachodzące w ściekach [9]. Wartość bioche-
micznego zapotrzebowania na tlen zależy od [9,10]:
rodzaju i stężenia substancji organicznych w ściekach,
aktywności mikroorganizmów oraz ich liczebność i rodzaj,
temperatury,
pH,
obecności związków hamujących zachodzące procesy.
Wartość BZT podaje się w mgO2/dm3 dla n – dni. W czasie 5 dni, około 68%
substancji organicznych ulega rozłożeniu, dlatego najczęściej oznacza się BZT5 [2,11-
13]. Zgodnie z rysunkiem 1 biodegradacja zanieczyszczeń zachodzi w dwóch fazach.
Rysunek 1. Krzywa przebiegu BZT [11]
Bez wątpienia warunkiem koniecznym prawidłowego przebiegu procesów bioche-
micznego utleniania jest obecność właściwej flory bakteryjnej, której aktywność
oddechowa wpływa na przebieg kluczowych procesów dla oczyszczania ścieków tj.
nitryfikacji i denitryfikacji [12, 15]. Metoda rozcieńczeń jest szeroko stosowanym
sposobem oznaczania BZT, ale jak już wspomniano badania takie mogą się opierać
również na określaniu zmian poziomu zanieczyszczeń w ściekach, wzrostu produkcji
np. CO2, czy ilości zużytego tlenu. Oznaczenie BZT5 jest czasochłonne i obarczone
ryzykiem niepowodzenia [4, 5, 13].

Sylwia Magdalena Duda, Monika Garbacz
192
3. Metody pomiaru uciążliwości zapachowych w gospodarce wodno-
ściekowej
Analizy jakościowe powietrza wykonuje się poprzez różne metody. Żadna z nich
jednak nie jest w pełni automatycznym systemem ciągłych pomiarów próbek powiet-
rza. Rysunek 2 przedstawia podział metod pomiaru oddziaływań zapachowych. Meto-
da sensoryczna opiera się na naturalnym działaniu ludzkiego zmysłu i określaniu stop-
nia uciążliwości zapachu, metody analityczne z kolei wykorzystują urządzenia
(instrumenty) do rozpoznawania składników związków zapachowo-czynnych. Ostatnia
z wymienionych metod, czyli analiza sensoryczno-instrumentalna polega na wykony-
waniu pomiarów instrumentami, które w dużym uproszczeniu naśladują ludzki zmysł
węchu.
Rysunek 2. Podział metod oceny oddziaływania zapachowego [16, 17]
Omawiane metody można klasyfikować ze względu na wykorzystywane metody
badawcze [17, 18]:
pomiary bezpośrednie – pomiary emisji i imisji (w pewnej odległości od źródła),
modelowanie dyspersji zanieczyszczeń w powietrzu – bazuje na wynikach
wspomnianych pomiarów.
3.1. Olfaktometria dynamiczna
Do oceny parametrów jakościowych i ilościowych mieszanin gazów szeroko
wykorzystywana jest i dynamicznie się rozwijająca chromatografia gazowa. Niestety ta
niezwykle czuła (niski próg detekcji) i dokładna metoda nie informuje o uciążliwości
zapachowej, a co za tym idzie stopniu zanieczyszczenia ścieków i pośrednio aktyw-
ności mikroorganizmów [4, 19]. Tak więc szeroko wykorzystywaną metodą oceny
uciążliwości związków zapachowo-czynnych jest olfaktometria dynamiczna.
Istotą oceny uciążliwości zapachowej w tej procedurze jest subiektywna opinia
specjalnie wybranych oraz przygotowanych testerów, co wiążę się z wielogodzinną
pracą nielicznych certyfikowanych jednostek w kraju. Aby wyniki były bardziej
wiarygodne zawsze uwzględnia się ocenę większej ilości osób. Wrażliwość węchową
badaczy ocenia się na podstawie reakcji na n-butanol (odorant odniesienia) [17].
Ponadto pomimo stosunkowo niskich kosztów inwestycyjnych na aparaturę

Ocena aktywności mikroorganizmów z wykorzystaniem matrycy czujników gazu
193
(w porównaniu np. do chromatografów), zdecydowanie kosztochłonna okazuję się
praca zespołu [4].
Olfaktometry dynamiczne są urządzeniami, w których badane próbki rozcieńczane są
czystym powietrzem, a zespół badawczy ocenia tak przygotowany strumień [19, 20].
Pomimo, iż początki prac na omawianą metodą badań sięgają końca XIX
w. dopiero w latach osiemdziesiątych XX w. pojawiły się pierwsze standaryzacje
Amerykańskiego Stowarzyszenia Badań i Materiałów, a także Niemieckiego Stowa-
rzyszenia Inżynierów. W tym samym czasie rozpoczął pracę Komitet Techniczny
funkcjonujący przy Europejskim Komitecie Normalizacyjnym, co poskutkowało
powstaniem olfaktometrycznej normy europejskiej w 2001 r., a dalej jej przetłu-
maczenia na język polski w 2007 r. [17].
3.2. Matryca czujników gazu
Inną stosowaną metodą są systemy oparte na wskazaniach czujników imitujących
ludzi zmysł węchu. Czujniki te sprawdzane są pod katem wielu odorantów.
System czujników połączonych z jednostką obliczeniową (np. komputerem lub
mikrokontrolerem), gdzie następuje analiza sygnałów oraz ich interpretacja tworzą
elektroniczny nos. Model elektronicznego nosa z 8 czujnikami gazu widoczny jest na
rysunku 3.
Rysunek 3. Widok matrycy czujników gazu po zdemontowaniu pokrywy przedniej [4]
Dzięki stosowaniu wielu sensorów o różnej czułości i selektywności istnieje możli-
wość stworzenia każdorazowo niepowtarzalnej kombinacji sygnałów tzw. gas
fingerprint (z ang. gazowy odcisk palca), charakterystycznej dla próbki gazu. W zależ-
ności od rodzaju stosowanych czujników (specyficznych lub niespecyficznych), mogą
one rozpoznawać poszczególne składniki próby bądź też grupy składników, co daje
możliwość powiązania odczytów z różnymi parametrami. Po otrzymaniu danych
pomiarowych stosuje się specjalne dobrane metody analityczne do pozyskania
konkretnych informacji o badanym gazie [5]. Do analiz wybierany jest zbiór profili,
będących odpowiedziami poszczególnych elementów matrycy. Różne czujniki
rozpoznają rozmaite związki, a co za tym idzie wytwarzają odmienne sygnały, które
razem tworzą pewną kombinację [4, 21, 22]. Dlatego, aby matryca była właściwie

Sylwia Magdalena Duda, Monika Garbacz
194
skalibrowana przydatna jest informacja na temat możliwości występowanie pewnych
związków w gazie poddawanym analizie [17].
3.2.1. Czujniki gazu i zasada ich działania
Według rysunku 4 zasadniczo wyróżnia się cztery grupy czujników stosowanych
w technologii elektronicznego nosa. Różnią się one między sobą m.in. czasem reakcji,
czułością i selektywnością reakcji na poszczególne gazy [4].
Rysunek 4. Podział czujników gazu stosowanych w matrycach [4]
Z punktu widzenia pracy matrycy wieloczujnikowej istotny jest również aspekt
rozmieszczenia czujników w stosunku do przepływającego strumienia badanej próbki.
Wyróżniamy wtedy [4, 23]:
matryce szeregowe – mieszanina przepływająca przepływa kolejno przez
wszystkie czujniki zainstalowane na matrycy, co czasem powoduje powstawanie
sygnałów zakłócających.
matryce równoległe w konstrukcji tej czujniki gazu rozmieszczone są tak, że
tworzą okrąg, a centralnie dopływający do komory gaz dociera w tym samym
momencie do każdego z odbiorników.
Uściślając, w omawianych matrycach wieloczujnikowych szeroko stosowane są
czujniki gazu typu MOS (z ang. Metal-Oxide-Semiconductor – metal-tlenek-półprze-
wodnik), wykorzystujące półprzewodnikowe tlenki metalu, najczęściej dwutlenek cyny
(SnO2) z różnymi domieszkami. Na powierzchni używanego spieku dwutlenku cyny
dochodzi do zmiany przewodnictwa (zjawisko chemisorpcji) co umożliwia wykonywa-
nie pomiarów, dzieje się tak ponieważ gaz oraz elektrony z półprzewodnika wytwa-
rzają wiązanie i ostatecznie następuje zmiana przewodnictwa elektrycznego. Częścią
omawianego procesu jest również powstanie m.in. cząsteczek CO2, CO, czy H2O [4].
Istotne i zarazem bardzo trudne jest uzależnienie wahań przewodnictwa od stężenia
badanych gazów. Można tego dokonać stosując poniższy wzór (1):
(1)
gdzie: RS – zmiana przewodnictwa elektrycznego, A – stała, c – stężenie gazu,
α – nachylenie krzywej RS.
Czujniki typu MOS klasyfikować można według grubości powłoki gazo czułej:
cienkowarstwowe (od 5 nm do 2 μm) – powłoka nakładana jest na czujnik
techniką rozpylania lub naparowywania,

Ocena aktywności mikroorganizmów z wykorzystaniem matrycy czujników gazu
195
grubowarstwowe (od 10 μm do 300 μm) – materiał nanoszony jest w postaci
pasty, kolejnym etapem przygotowania czujnika jest wypalenie w wysokiej
temperaturze tlenków metali.
W celu poprawy selektywności, czyli zdolności wyłapywania konkretnych zanie-
czyszczeń w omawianej warstwie gazoczułej stosuje się domieszki metali szlachetnych
tj. złoto, srebro.
3.2.2. Opracowywanie odczytanych danych
Zbiory sygnałów z matrycy wieloczujnikowej są złożone i muszą być analizowane
za pomocą odpowiednich systemów rozpoznawania wzorca. Według zgromadzonych
danych najszerzej wykorzystywane są następujące metody PCA (z ang. Principal
Component Analysis – Analiza Głównych Składowych) lub ANN (z ang. Artifficial
Neural Network – Sztuczne Sieci Neuronowe) [4, 24 - 26].
W dużym uproszczeniu metoda statystyczna PCA polega na grupowaniu danych
poprzez dobraniu nowych zmiennych opisujących jak najdokładniej rozważany profil
informacji z matrycy. Metoda ta zmniejsza wielowymiarowość zebranych danych,
czyli jest metodą kompresji danych i tym samym umożliwia pokazanie ich na
wykresach. W wyniku przekształceń cześć pierwotnych danych zanika, ale istnieje
możliwość odwrócenia transformacji i powrót do informacji wyjściowych. Co ważne
analiza PCA pozwala na wdrożenie jej w strukturę sterującą i prowadzenie transfor-
macji pomiarów jeszcze w trakcie trwania pomiarów uciążliwości zapachowej [27].
Sztuczne Sieci Neuronowe według części autorów są uznawane za lepsze roz-
wiązanie [24, 25]. Technika ta bazuje na skomplikowanej sieci połączonych neuronów
występującej w ludzkim mózgu, gdzie dendryty gromadzą informacje z sąsiednich
komórek i w pewien sposób przetransformowane dane są przekazywane dalej poprzez
aksony. Można więc powiedzieć, że ANN funkcjonuje opierając się na wykreowanym
modelu matematycznym z przetwarzaniem danych według wyżej opisanego schematu
[28, 29]. W celu sprzężenia ANN z matrycą wieloczujnikową wykonuje się tzw.
uczenie według różnych algorytmów. Ilość warstw neuronów dobiera się w zależności
od stopnia skomplikowania relacji danych wejściowych i wyjściowych. Tak skorelo-
wany układ jest porównywany z ludzkim zmysłem powonienia [4, 30].
4. Korelacja wskazań matrycy i klasycznych wskaźników zanieczyszczeń
ścieków
Kluczowym zagadnieniem wydaje się odpowiedź na pytanie, czy na podstawie
poznanego składu fazy gazowej (powietrze) można określić parametry fazy ciekłej
(ścieki)? Otóż na rynku dostępne są respirometry, czyli urządzenia oceniające BZT na
podstawie próbki powietrza. Natomiast system elektronicznego nosa jak już
wspomniano szeroko wykorzystywany jest nie tylko do badania uciążliwości zapa-
chowych ścieków, ale również pośredniej oceny parametrów fizykochemicznych
ścieków – BZT, ChZT, OWO, Zog. Ogromną zaletą matryc w stosunku do respirome-
trów jest fakt, iż umożliwiają one szacowanie kilku kluczowych parametrów badanego
medium w kontekście eksploatacji obiektu [31].

Sylwia Magdalena Duda, Monika Garbacz
196
Onkal-Engin i in. w 2010 r. badał za pomocą matrycy czujników gazu próbki po-
wietrza z różnych miejsc oczyszczalni ścieków komunalnych i przemysłowych oraz
klasyfikował je wykorzystując ANN. Zespół stwierdził, że ogólne wyniki wskazują na
to, że metodę tę można użyć do klasyfikacji próbek ścieków, a także elektroniczny nos
może być wykorzystany jako urządzenie do monitorowania biochemicznych
zapotrzebowania na tlen [32].
Według wyników badań z 2014 r., których celem był rozwój technik ostrzega-
wczych przed zakłóceniami procesów oczyszczania ścieków z wykorzystaniem
elektronicznego nosa. Zespół prowadzący eksperyment zauważył, że wraz ze wzrostem
stężenie substancji zapachowo-czynnych maleje sygnał płynący z czujników gazu,
a „zaprogramowana” sieć neuronowa określa stopień uciążliwości zapachowej [5]. Do
takich samych wniosków doszedł w 2016 r. zespół z Politechniki Lubelskiej, który
przeprowadził badania uciążliwości zapachowej w trakcie wymuszonej awarii oraz
stabilnej pracy laboratoryjnego rektora SBR z osadem czynnym. Wykorzystali oni
matrycę równoległą z czujnikami typu MOS i zaimplementowaną sztuczną siecią
neuronową [4, 21]. Przytoczone prace potwierdzają zasadność używania matryc
wieloczujnikowych do monitoringu procesów zachodzących w trakcie oczyszczania
ścieków. Mogą one rozpoznawać poszczególne etapy i stany pracy oczyszczalni
(warunki tlenowe/beztlenowe; ścieki surowe/oczyszczone) [5].
Badania zespołu z Politechniki Lubelskiej (Guz i in. z 2014) dowiodły, iż możliwe
jest stworzenie sieci neuronowej o parametrach pozwalających przybliżyć wartość
ChZT i Zog (zawiesina ogólna) w ściekach [33].
5. Podsumowanie i wnioski
Biochemiczne zapotrzebowanie tlenu informuje o jakości procesu oczyszczania
ścieków, a co za tym idzie o aktywności mikroorganizmów w trakcie oczyszczania
biologicznego. Innymi równie ważnymi wskaźnikami poziomu zanieczyszczeń są
ChZT oraz OWO, które jest ściśle związane z BZT. Co więcej znając rodzaj badanego
medium możemy określić wzajemne stosunki wartości poszczególnych wskaźników.
Tradycyjne oznaczenia są dokładne, ale trwają długo i niejednokrotnie wymagają
dużych nakładów pracy. Bez wykonania pełnej analizy niemożliwe jest stwierdzenie
bądź wykluczenie obecności zanieczyszczeń, czy substancji toksycznych kluczowych
dla poprawnego działania części biologicznej obiektów służących oczyszczaniu
ścieków.
Matryce wieloczujnikowe można stosować w obiektach o dużej uciążliwości
zapachowej m.in. obiektach gospodarki wodno-ściekowej. Urządzenia te mogą
analizować różne gazy, także te wytwarzane w trakcie przemian metabolicznych
drobnoustrojów. Matryca czujników gazu wykonuje badania szybko, stosunkowo tanio
oraz w powtarzalny sposób, pod warunkiem częstej kalibracji. Jest to metoda właściwa
dla tak zwanych badań przesiewowych, czyli informujących o występowaniu bądź nie
zanieczyszczeń na poziomie alarmującym. Prowadzenie badań online umożliwia
prowadzenie ciągłego monitoringu w obiektach inżynierskich, a zatem tradycyjne

Ocena aktywności mikroorganizmów z wykorzystaniem matrycy czujników gazu
197
oznaczenia (drogie i skomplikowane) są wykonywane w momencie zanotowania
konkretnych zanieczyszczeń.
Podsumowując matryce wieloczujnikowe są ciągle rozwijającą się metodą
o szerokim zastosowaniu. Ciągłe pomiary nad zwierciadłem ścieków umożliwiają
pozyskanie informacji o przebiegu realizowanych procesów. Jest to rozwiązanie, które
uzupełnia i wspomaga tradycyjne sposoby oznaczania wskaźników zanieczyszczeń.
Ponad to praca omawianych urządzeń i wyniki z prowadzonych badań służą tworzeniu
systemów wczesnego ostrzegania i pomagają w modelowaniu procesów oczyszczania
ścieków.
Literatura
1. Lenart A., Kowalska A., Wykorzystanie osadu czynnego w oczyszczaniu ścieków,
KOSMOS Problemy Nauk Biologicznych, 61 (2012), s. 677-689
2. Janosz-Rajczyk M., Badania wybranych procesów oczyszczania ścieków, Wydawnictwo
Politechniki Częstochowskiej, Częstochowa 2008.
3. Łomotowski J., Szpindor A., Nowoczesne systemy oczyszczania ścieków, Arkady,
Warszawa 1999.
4. Guz Ł., Łagód G., Suchorab Z., Sobczuk H., Kurek E., Application of e-nose to smell
nuisance evaluation of watste water treatment plant devices, Monografie Politechnika
Lubelska, Lublin 2016.
5. Guz Ł., Łagód G., Jaromin-Gleń K., Sobczuk H., Wykrywanie zakłóceń procesu
oczyszczania ścieków w bioreaktorze porcjowym z wykorzystaniem e-nosa, Proceedings of
ECopole, Vol 8, No 1. (2014), s. 147-153.
6. Capelli L., Sironi S., Centola P., Del Rosso R., Grande MI., Electronic noses for the
continuous monitoring of odours from a wastewater treatment plant at specific receptors:
Focus on training methods, Sens Act B, 131 (2008), s. 442-452.
7. Onkal-Engina G., Demir I., Engin SN, Determination of the relationship between sewage
odour and BOD by neural networks, Environ Modell Softw, 20 (2005), s. 843-850.
8. Guz Ł., Suchorab Z., Sobczuk H., Pomiar odorów za pomocą przenośnego miernika z
matrycą półprzewodnickowych czujników gazu, Przem Chem, 89 (4) (2010), s. 378-381.
9. Sanak - Rydlewska S., Metody oczyszczania ścieków miejskich, AGH Uczelniane
Wydawnictwa Naukow-Dydaktyczne, Kraków 2009.
10. Sadecka Z., Podstawy biologicznego oczyszczania ścieków, Wydawnictwo "Seidel-
Przywecki" Sp. z o.o., Bydgoszcz 2010.
11. Stier E., Fischer M., Podręczny poradnik eksploatacji oczyszczalni ścieków, Wydawnictwo
„Seidel-Przywecki” Sp. z o.o., Bydgoszcz 1998.
12. Miksch K., Sikora J., Biotechnologia ścieków, Wydawnictwo Naukowe PWN, Warszawa
2010.
13. Miksch K., Czerska B., Grabińska-Sota E., Kalka J., Surmacz-Górska J., Biotechnologia
ścieków, Wydawnictwo Politechniki Śląskiej, Gliwice 2000.
14. Henze M., Harremoes P., Jansen J. C, Arvin E., Oczyszczanie ścieków. Procesy
biologiczne i chemiczne, Wydawnictwo Politechniki Świętokrzyskiej, Kielce 2002.
15. Głodniok M., Korczak K., Zdebik D., Metodyczne aspekty analizy aktywności oddechowej
mikroorganizmów osadu czynnego w odniesieniu do możliwości optymalizacji pracy
biologicznej oczyszczalni ścieków, Prace Naukowe GIG Górnictwo i Środowisko, 1 (2010),
s. 5-13.

Sylwia Magdalena Duda, Monika Garbacz
198
16. Gostelow P., Persons S.A., Stuetz R.M., Odour maesurements for sewage treatment works,
Water Research, vol. 35 no. 3 (2001), s. 579-597.
17. Barczak R., Analiza porównawcza olfaktometrii terenowej z innymi metodami badawczymi
w ocenie oddziaływania zapachowego oczyszczalni ścieków, rozprawa doktorska,
Warszawa 2014.
18. Witherspoon J.R., Barnes J.L., Comparison of methods used to measure odour at
wastewater treatment plant fencelines, Environmental Odour Management International
Conference, Cologne (2004), s. 363-375.
19. Witkiewicz Z., Hetper J., Chromatografia gazowa, Wydawnictwa Naukowo-Techniczne,
Warszawa 2001.
20. Kośmider J., Pomiary stężenia zapachowego metodą olfaktometrii dynamicznej zgodnie z
PN-EN 13725:2007, Wodociągi i Kanalizacja, 10 (2007), s. 34-35.
21. Guz Ł., Łagód G., Jaromin-Gleń K., Guz E., Sobczuk H., Assessment of batch bioreactor odour
nuisance using an e-nose, Desalination and Water Treatment, 57 (3) (2016), s. 1327-1335.
22. Sobczuk H., Guz Ł., Pomiary zanieczyszczeń odorowych wieloczujnikowym urządzeniem z
tlenkowymi czujnikami gazu, Rocznik Ochrony Środowiska, 13 (2011), s. 1531-1542.
23. Nakamoto T., Odor Handling and Delivery Systems, In: Pearce T.C., Schiffman S.S., Nagle
H.T., Gardner J.W., (Eds.), Handbook of Machine Olfaction Electronic Nose Technology,
Viley-Vch., Weinheim (2006).
24. Llobet E., Brezmes J., Vilanova X., Sueiras J. E., Correig X., Qualitative and quantitative
analysis of volatile organic compounds using transient and steady-state response of a thick-
film tin oxide gas sensor array, Sensors and Actuators B: Chemical, 41 (1997), s. 13-21.
25. Haugen J. E., Kvaal K., Electronic nose and artificial neural network, Meat Science, 49(1)
(1998), s. 273-286.
26. Gardner J. W., Bartlett P. N., A brief history of electronic noses, Sensors and Actuators B,
18 (1-3) (1994), S. 210-212.
27. Krzanowski W. J., Principles of Multivariate Analysis, Oxford University Press, New York
(2000).
28. Smolarz A., Kotyra A., Wójcik W., Ballester J., Advanced diagnostics of industrial
pulverized coal burner using optical methods and artificial intelligence, Experimental
Thermal and Fluid Science, 43 (2012), s. 83-89.
29. Osowski S., Sieci neuronowe do przetwarzania informacji, Oficyna Wydawnicza P. W.,
Warszawa 2000.
30. El-Din A. G., Smith D. W., A neural network model to predict the wastewater inflow
incorporating rainfall events, Water Research, 36 (5) (2002), s. 1117-1124.
31. Guz Ł., Łagód G., Jaromin-Gleń K., Suchorab Z., Sobczuk H., Bieganowski A.,
Application of Gas Sensor Arrays in Assessment of Wastewater Purification Effects,
Sensors, 15 (2015), s. 1-21.
32. G. Onkal-Engin, I. Demir, S.N. Engin, Determination of the relationship between sewage
odour and BOD by neural networks, Environ. Model. Softw. 20 (2005) 843–850.
33. Guz Ł., Łagód G., Jaromin-Gleń K., Sobczuk H., Metoda Pośredniego okreslania wartości
ChZT i zawiesiny w bioreaktorach z osadem czynnym za pomocą matrycy czujników
nieselektywnych, Materiały Konferencyjne, Kijów 2014.

Ocena aktywności mikroorganizmów z wykorzystaniem matrycy czujników gazu
199
Ocena aktywności mikroorganizmów z wykorzystaniem matrycy czujników gazu
Streszczenie
Obiekty służące oczyszczaniu ścieków są emiterami związków zapachowo-czynnych do atmosfery na ró-
żnych etapach swojej pracy, co z kolei w znacznej mierze związane jest z aktywnością mikroorganizmów.
Uciążliwość zapachową analizować można za pomocą różnych technik – na przykład olfaktometrii dynamicznej, chromatografii gazowej jak również z wykorzystaniem matryc wieloczujnikowych. Emisja
związków zapachowo-czynnych jest również związana z poziomem zanieczyszczenia, które wyrażane
może być za pomocą biologicznego zapotrzebowania na tlen (wskaźnika BZT5). Wykazano, że parametry
określające jakość ścieków odzwierciedlają aktywność mikroorganizmów osadu czynnego, a co za tym idzie efektywność procesów oczyszczania.
W pracy omówiono korelację pomiędzy wskazaniami matrycy czujników a standardowymi parametrem
jakości ścieków: BZT5, ChZT, Zog. W niniejszym opracowaniu dokonano przeglądu metodyki analiz
odorów i odorantów, a w szczególności charakterystyki pracy matryc czujników gazu. Omówiono także wyniki prowadzonych badań, pod katem powiązania wskazań elektronicznego nosa i parametrów fizy-
kochemicznych badanego medium. Udowodniono, iż możliwe jest szacowanie wartości wspomnianych
parametrów na podstawie wskazań matrycy wieloczujnikowej, a co za tym idzie uciążliwości zapachowej
ścieków. E-nosy są ciągle rozwijającą się metodą o szerokim zastosowaniu m.in. w gospodarce wodno-ściekowej.
Jest to rozwiązanie, które wspomaga i uzupełnia tradycyjne sposoby oznaczania wskaźników zanieczysz-
czeń badanego medium.
Słowa kluczowe: BZT5, uciążliwość zapachowa, matryca czujników gazu, elektroniczny nos
Evaluation of microbial activity using a gas sensor array
Abstract Sewage treatment plants are one of the odour-active compounds emitters at all stages of purification, which
is closely related to the activity of microorganisms. Odour nuisance can be analyzed by a variety of
techniques – for example, dynamic olfactometry, gas chromatography and by multi-sensor matrices. The
odor nuisances of sewage are also linked to the contamination level, which can be considered as a biological oxygen demand (BOD5) – a kind of microbial activity measurement, informing about the
processes of transformation and biodegradation of impurities.
The aim of the study is to discuss the correlation between the results of multi-sensor matrices tests and
standard wastewater quality parameters: BOD5 and COD. The current national and foreign literature on odor and odorant analysis has been reviewed, paying special attention to work characteristics of gas sensor
matrics. The results of the study were also discussed, in terms of connection between e-nose indications
and wastewater quality parameters.
The conducted research allowed to draw conclusions concerning the connection of readings from the gas sensor matrics with biological oxygen demand, treated as an indicator of impurity and odor nuisance of the
tested medium.
Keywords: BOD5, E-Nose, odor nuisance

200
Monika Garbacz1, Sylwia Magdalena Duda
2
Syndrom chorego budynku
oraz jego wpływ na zdrowie mieszkańców
w kontekście mikrobiologii środowiska wewnętrznego
1. Wprowadzenie
W dzisiejszych czasach Syndrom Chorego Budynku zalicza się do trudnych proble-
mów dotyczących sektora budowlanego. Ochrona przed korozją biologiczną stanowi
szerokie zagadnienie w branży budowlanej, jednakże łączenie problemów technicz-
nych, jak i biologicznych stwarza nie lada wyzwanie dla współczesnych inżynierów.
Wzrastająca liczba ludzi podatnych na alergie wywołane grzybami pleśniowymi
powoduje większe zainteresowanie tym problemem oraz jego wpływem na stan
zdrowia człowieka. Mykologia budowlana zwana patologią budynków (ang. building
pathology) stanowi interdyscyplinarną gałąź wiedzy i praktyki łączącą w sobie
problemy biologii z problemami budowlano-technicznymi [1].
Według Światowej Organizacji Zdrowia (WHO) jakość powietrza wewnątrz
pomieszczenia wpływa w znaczny sposób na zdrowie i samopoczucie człowieka.
Wynika to głównie z faktu, że ludzie w wielu krajach spędzają nawet 90% czasu wew-
nątrz budynków. Badania naukowe potwierdzają, że powietrze wewnątrz pomieszczeń
jest bardziej zanieczyszczone w porównaniu z powietrzem zewnętrznym. Najbardziej
charakterystycznym przykładem nieprawidłowej jakości powietrza wewnątrz
pomieszczeń jest Syndrom Chorego Budynku (SBS ang. Sick Building Syndrome). Jest
to jeszcze niezbyt dobrze poznany temat związany z problemami zdrowotnymi osób
przebywających w budynkach. Syndrom Chorego Budynku możne zostać uznany za
nowa jednostkę chorobotwórczą, powodującą poważne zagrożenia zdrowotne.
Określenie Sick Building Syndrome pojawiło się w pismach medycznych pod koniec
lat 70 ubiegłego wieku. Ściśle ujmując pojecie to stanowi kombinację różnych
dolegliwości zdrowotnych, których występowanie związane jest z przebywaniem ludzi
w konkretnych pomieszczeniach zamkniętych. Początkowo problem ten dotyczył
głownie pomieszczeń biurowych z czasem jednak zauważono, że może on dotyczyć
również pomieszczeń mieszkalnych [2].
Celem artykułu jest analiza zagadnienia Syndromu Chorego Budynku wynikającego
ze złego zaprojektowania czy użytkowania budynku. Zwrócono również dużą uwagę
na objawy przebywania w „chorych pomieszczeniach zamkniętych”. W 1982 r.
Światowa Organizacja Zdrowia (WHO) uznała Syndrom Chorego Budynku za
zdefiniowany problem zagrażający zdrowiu człowieka. WHO za „chory budynek”
1 [email protected], Koło Młodych Inżynierów PZiTS, Wydział Inżynierii Środowiska, Politechnika
Lubelska, ul.Nadbystrzycka 40 B, Lublin 20-618, Tel.+48815384784 2 [email protected], SKN FOR&AGAINST, Wydział Inżynierii Środowiska, Politechnika Lubelska
ul.Nadbystrzycka 40 B, Lublin 20-618, Tel.+48815384784

Syndrom chorego budynku oraz jego wpływ na zdrowie mieszkańców w kontekście mikrobiologii
środowiska wewnętrznego
201
uznaje taki, w którym więcej niż 1/3 użytkowników uskarża się na złe warunki czy
niekorzystny mikroklimat. Do najczęstszych dolegliwości należą bóle głowy
i problemy z oddychaniem, a o SBS można mówić, gdy ich przyczyną jest klimat
wewnętrzny budowli lub jej wady konstrukcyjne. Dolegliwości są tym silniejsze, im
dłużej przebywa się w chorym budynku [3].
2. Przyczyny zespołu chorego budynku
Zespół Chorego Budynku występuje u osób przebywających w pomieszczeniach
zamkniętych, w których panuje niekorzystna jakość powietrza. Na ten stan budynku
może wpływać wiele rożnych czynników. Najczęstszą przyczyną pojawienia się SBS-u
jest obecność zanieczyszczeń mikrobiologicznych (bakterii, pleśni, grzybów, pyłków
roślin), zanieczyszczeń chemicznych (wyemitowanych przez materiały budowlane,
urządzenia biurowe lub inne wyposażenie techniczne ) a także niewłaściwie zapro-
jektowana lub wykonana wentylacja czy klimatyzacja. Zdarza się, że do przyczyn SBS-
u zalicza się również różnego rodzaju hałasy, wibracje, zjawiska radiacyjne czy
elektrostatyczne, które uniemożliwiaj normalne funkcjonowanie osób przebywających
w pomieszczeniu [3].
Do zanieczyszczeń mikrobiologicznych zlicza się głównie grzyby, pleśnie, bakterie
oraz inne drobnoustroje rozwijające się na pożywce materiałów budowlanych (Rysu-
nek 1). Mikroorganizmy te zagrożenie zdrowia człowieka w sposób bezpośredni lub
przez wytwarzanie trujących substancji tzw. mykotoksyn – bardzo niebezpiecznych dla
zdrowia człowieka produktów przemiany materii grzybów. Porażenie mikrobie-
logiczne budynków odpowiada za większość problemów zdrowotnych związanych
z jakością powietrza wewnętrznego [4].
Rysunek 1. Mikroorganizmy pokrywające powierzchnie elewacji budynku użyteczności publicznej
[opracowanie własne]

Monika Garbacz, Sylwia Magdalena Duda
202
Poznanie przyczyn Syndromu Chorego Budynku wymaga zbadania i analizy
konkretnych przypadków budowlanych. W ubiegłych latach problem zagrzybienia
występował głównie w starych, zwilgoconych lub nieocieplonych budynkach, jednak
coraz częściej pojawia się w nowo budowanych obiektach. Najpoważniejszym źródłem
porażenia budynków są grzyby. Mimo znacznego rozwoju społeczeństwa zarówno pod
względem higienicznym, jak i technicznym nadal niepokojącym problemem są
zagrzybione budynki [3].
Grzyby oraz pleśnie stanowią poważne zagrożenia dla zdrowia i życia ludzi przeby-
wających w zamkniętych pomieszczeniach budowlanych, do których zalicza się:
mieszkania, biurowce (Rysunek 2), placówki oświaty, służby zdrowia oraz wszelkiego
rodzaju budynki stanowiące zamkniętą przestrzeń. Często dochodzi do pogorszenia
stanu zdrowia ludzi przebywających w tego typu pomieszczeniach – jest to zespól
chorób nazywanych w literaturze BRI – (ang. Building Related Illness) [3]. Może być
to powodowane niehigienicznym użytkowaniem lokali, dotyczy to m.in. trzymaniem
zwierząt w pomieszczeniach, brak wentylacji lub zwyczaju przewietrzania pomiesz-
czeń. Jednakże najczęstszą przyczyną porażenia mikrobiologicznego budynków są
wady konstrukcji budowlanych, które stwarzają dogodne warunki do rozwoju grzybów
w tym pleśni, wytwarzających niebezpieczne toksyny [1].
Rysunek 2. Mikroorganizmy pokrywające powierzchnie elewacji budynku biurowego [opracowanie własne]
Warunki sprzyjające rozwojowi mikroorganizmów to przede wszystkim wilgotność względna przekraczająca 60%, tworzenie się wilgotnej pary lub powłoki na powierzch-ni ścian i okien oraz dodatnie temperatury. Powietrze staje się wilgotne w momencie osiągnięcia temperatury punktu rosy. Spowodowane jest to niedostateczną izolacją przegród budowlanych, gdy na zewnątrz panuje niska temperatura i przenika do

Syndrom chorego budynku oraz jego wpływ na zdrowie mieszkańców w kontekście mikrobiologii
środowiska wewnętrznego
203
wnętrza przegród budowlanych. W pomieszczenia, które są intensywnie ogrzewane dochodzi do skroplenia wilgoci na elementach przegrody. Jest to dogodny klimat do rozwoju grzybów [5].
Zanieczyszczenia mikrobiologiczne występują w postaci bioareozoli, czyli zawie-szonych w powietrzu cząstek biologicznych. Znajdują się w nim różnorodne, pocho-dzące z wielu źródeł mikroorganizmy i cząsteczki substancji organicznej. Źródło bio-areozoli stanowią ludzie i zwierzęta zamieszkujące mieszkanie. Zanieczyszczenia występujące w dużym stężeniu wytwarzane przez grzyby i pleśnie, pochodzą głównie od organizmów rosnących w mieszkaniu [5].
Kolejną zasadniczą kwestią jest niedostateczne wietrzenie pomieszczeń lub utrud-niona wentylacja. Jest to czynnik sprzyjający wzrostowi stężenia składników bioareo-zoli, powstającego w przestrzeni zamkniętej. Dalszą konsekwencją tego problemu jest wzrost zwilgocenia pomieszczeń w wyniku zbyt wolnego usuwania pary wodnej obecnej w powietrzu (Rysunek 3).
Rysunek 3. Porażenie mikrobiologiczne występujące w pomieszczeniach zamkniętych [opracowanie własne]
Woda jest produktem przemiany materii ludzi i zwierząt oraz spalania substancji organicznej. Para wodna powstaje podczas zwykłych czynności m.in. przygotowanie posiłku czy utrzymywanie higieny osobistej. W gospodarstwach domowych powstaje jej niewyobrażalna ilość. Jak podają źródła [6]: 4 osobowa rodzina w gospodarstwie domowym emituje do powietrza w mieszkaniu ok. 14-15 litrów wody dziennie, skali roku daje to ponad 5 ton.
Do najczęstszych błędów budowlanych, jeszcze na etapie projektowania zalicza się głównie niewłaściwy dobór materiałów oraz zabezpieczeń chroniących budynek przed wilgocią i zmianą temperatury, zwłaszcza dotyczy to piwnic i stropów. Już na etapie realizowania budowy popełnia się wiele błędów, które w późniejszym czasie wpływają niekorzystnie na stan budynku. Do najczęstszych zalicza się: skrócony czas realizacji inwestycji, montaż szczelnej stolarki budowlanej, zastosowanie ociepleniowych systemów, które w znacznym stopniu utrudniają wysychanie ścian obiektu budow-lanego [4].

Monika Garbacz, Sylwia Magdalena Duda
204
Źródłem zanieczyszczeń chemicznych w budynkach są najczęściej materiały budowlane (kleje, izolacje, lakiery, farby), urządzenia biurowe, wyposażenie tech-niczne oraz wszelkiego rodzaju środki chemiczne. Do najczęściej występujących sub-stancji szkodliwych zalicza się: toluen, ksylen, formaldehyd, benzen i styren. Trujący formaldehyd wydzielają niektóre wykładziny i płyty wiórowe, z których produkuje się meble. Farby mogą zawierać metale ciężkie, a wykorzystywania w budownictwie rakotwórczego azbestu zakazano dopiero w latach 90. ub. w.
Niesprawna wentylacja lub jej całkowity brak prowadzi do niewłaściwej cyrkulacji powietrza wewnątrz obiektu. Wentylacja prawidłowo zaprojektowana i wykonana powinna odprowadzić zużyte powietrze oraz dostarczyć świeże. Jej podstawowym zadaniem jest usuwanie wydychanego przez ludzi dwutlenku węgla – w powietrzu dobrej jakości może go być maksymalnie 0,1% objętości. W większych stężeniach szkodzi zdrowiu, może nawet być zabójczy. Prawidłowa zawartość tlenu to minimum 20%. W źle wentylowanej sypialni może spaść do 17%, co także jest groźne dla zdrowia.
Aby usprawnić wentylacje można zamontować nawiewniki w oknach lub nawiew-niki ścienne. Przy zakupie okien warto zwrócić uwagę na takie, w których nawiewniki są zamontowane już przez producenta [1]. Kolejnym ważnym celem wietrzenia pomieszczeń jest usuwanie pary wodnej i utrzymywanie wilgotności powietrza między 40 a 60%. Jeśli powietrze będzie za suche – pojawią się problemy z oddychaniem czy drapanie w gardle. Gdy będzie zbyt wilgotne – stworzymy dobre warunki dla rozwoju mikroorganizmów.
Wraz z postępem technicznym pojawił się jeszcze jeden problem – dogodnym miejscem dla rozwoju drobnoustrojów okazały się instalacje klimatyzacyjne i wenty-lacyjne. Występujące powszechnie w nieczyszczonej klimatyzacji bakterie z rodza-ju Legionella, które oddziaływają niekorzystnie na zdrowie człowieka.
Efekt postępu technicznego w branży budowlanej spowodował również negatywne konsekwencje. Mianowicie, budowane są zbyt szczelne budynki, w których jest za ciepło i brakuje świeżego powietrza. Zbyt wysoka temperatura powietrza, wilgotność oraz koncentracja zanieczyszczeń powoduje spadek wydajności i produktywności człowieka [1].
3. Objawy Syndromu Chorego Budynku
Według Światowej Organizacji Zdrowia (WHO) jakość powietrza występującego wewnątrz pomieszczeń zamkniętych intensywniej wpływa na zdrowie ludzi i zwierząt, w przeciwieństwie do powietrze zewnętrznego, które jakby mogło się wydawać jest bardziej zanieczyszczone [1].
Najbardziej charakterystycznym przykładem niewłaściwej jakości powietrza w pomieszczeniach jest Syndrom Chorego Budynku (SBS), którego główne objawy to: zapalenie błony śluzowej, astma, zapalenie oskrzeli i krtani, a także migreny, rozdraż-nienie, zaburzenia koncentracji prowadzące do zmniejszenia motywacji i efektywności pracy [4].
Stan zdrowia człowieka uzależniony jest od czasu narażenia od rodzaju czynnika oraz liczby mikroorganizmów obecnych w powietrzu. Najczęstszym przenośnikiem czynników biologicznych w powietrzu są bioareozole, zawierające elementy biolo-

Syndrom chorego budynku oraz jego wpływ na zdrowie mieszkańców w kontekście mikrobiologii
środowiska wewnętrznego
205
giczne, krople wody oraz inne zanieczyszczenia. Człowiek jest narażony na bezpo-średni kontakt z bioarezolem, przez układ oddechowy, skórę, oczy oraz włosy. Jednak-że stopień narażenia jak i przebieg choroby jest indywidualny i zależy od wielu czyn-ników m.in. od cech osobniczych, przebytych chorób, przebytych terapiach zwiększa-jących szanse na infekcje [9].
W czasie oddychania zarodniki grzybów dostają się do płuc i u osób uczulonych powodują poważne choroby płuc. Wielu zamieszkujących je lokatorów odczuwa różne dolegliwości (senność, zmęczenie, nudności, zawroty głowy, drażliwość, zaburzenia pamięci, ociężałość, szybkie i łatwe męczenie się, długotrwałe bóle i częste przezię-bienia, podrażnienie błon śluzowych oczu, nosa, krtani, gardła, zaczerwienienie skóry, objawy zbliżone do astmy: ucisk w klatce piersiowej, duszności) [1].
Syndrom Chorego Budynku spowodowany jest również występowaniem w powietrzu Mikrobiologicznych Lotnych Związków Organicznych (MVOC), myko-toksyn, alergenów, zarodników i fragmentów grzybów. Główne objawy to: zapalenie błony śluzowej, astma, zapalenie oskrzeli i krtani, a także migreny, rozdrażnienie, zaburzenia koncentracji prowadzące do zmniejszenia motywacji i efektywności [10].
Listę objawów, związanych z przebywaniem w chorym budynku, WHO ogłosiła w 1987 r. Znalazły się na niej:
bóle i zawroty głowy, omdlenia, mdłości, utrudnione oddychanie, nienaturalne zmęczenie.
dolegliwości alergiczne: zapalenie śluzówek, przewlekłe zapalenia krtani, oskrzeli, suchy kaszel.
objawy związane z samopoczuciem: migreny, rozdrażnienie, zaburzenia koncentracji.
przesuszenie, zaczerwienienie i łuszczenie skóry twarzy, rąk, uszu. Z problemem Syndromu Chorego Budynku łączone są również poważne
schorzenia, na które ludzie zapadają w wyniku przebywania w zanieczyszczonym środowisku wewnętrznym, np.:
astmę oskrzelową,
chorobę legionistów, gorączkę klimatyzacyjną,
choroby nowotworowe, pojawiające się w następstwie oddziaływania dymu tytoniowego, azbestu, radonu.
Jednakże aby móc je odróżnić od uciążliwego, ale nie tak poważnego Syndromu Chorego Budynku, nazwano je Zespołem Chorób Związanych z Budynkiem (Building Related Illness).
Przy diagnozowaniu Syndromu Chorego Budynku znaczenie ma ustępowanie objawów po opuszczeniu chorego budynku i to, czy wystąpiły one u innych osób w nim przebywających. WHO zauważa, że SBS pojawia się na ogół w budynkach nowych lub modernizowanych (biurowych i mieszkalnych) oraz w obiektach ze skomplikowanymi systemami wentylacyjnymi [2].
4. Metody oceny stopnia zanieczyszczenia środowiska wewnętrznego
Do podstawowych metod oceny stopnia zanieczyszczenia środowiska wew-nętrznego zalicza się: metodę mikrobiologiczną, chemiczną oraz szybką detekcję za pomocą matrycy wieloczujnikowej (tzw. elektronicznego nosa).

Monika Garbacz, Sylwia Magdalena Duda
206
Metoda mikrobiologiczna Metoda mikrobiologiczna oceny jakości powietrza wewnętrznego może zostać
podzielona na metodę tradycyjną (mykologiczną) oraz molekularną. Ocena mykologiczna opiera się na makroskopowej i mikroskopowej obserwacji
zebranych próbek, oznaczanie wilgoci w zebranych próbkach, ocenie wydajności wentylacji i innych instalacji sanitarnych. Metoda ta oparta jest na określaniu obecności grzybów pleśniowych w próbkach powietrza lub wymazach, obliczaniu liczby kolonii oraz określeniu gatunku lub rodzaju. Wśród tradycyjnych technik można wyróżnić:
metodę sedymentacji Kocha,
metodę filtracji przez wychwytywanie żywych i martwych grzybów,
metoda kaskady zderzeniowej, która opiera się na przepływie zanieczyszczonego powietrza przez małe otwory [10].
Impaktor kaskadowy (Rysunek 4) jest to urządzenie służące do oznaczania określonych rozkładów wielkości wszelkich pyłów tworzących się w aerozolu. Przy pomocy odpowiednio wyprofilowanych kanałów, które prowadzą dany aerozol, pyły o różnych średnicach osadzają się w różnych miejscach [12].
Rysunek 4. Impaktor kaskadowy [opracowanie własne]
Zastosowanie tradycyjnych technik mikrobiologicznych jest czasochłonne i może trwać nawet do 43 dni.
Do metod molekularnych wykrywania porażeń mikrobiologicznych należy metoda PCR (Reakcja łańcuchowa polimerazy), opierająca się na amplifikacji fragmentu DNA w wyniku podgrzewania i ochładzania próbki w warunkach laboratoryjnych. PCR jest metodą stosunkowo kosztowną oraz czasochłonną jednakże zastosowanie tej metody oferuje poznanie jednoznacznych wyników [13].

Syndrom chorego budynku oraz jego wpływ na zdrowie mieszkańców w kontekście mikrobiologii
środowiska wewnętrznego
207
Metody chemiczne Metody chemiczne umożliwiają rozpoznanie wybranych markerów będących
produktami metabolitów takich jak, mykotoksyny czy lotne związki organiczne. Wśród wszystkich metod najbardziej popularne to:
chromatografia gazowa-spektrometria masowa,
wysokosprawna chromatografia cieczowa. Obie metody wymagają skomplikowanego procesu przygotowania próbki, które
wymaga dużo pracy ręcznej. Szybka detekcja za pomocą matrycy wieloczujnikowej Szybka detekcja pozwalają na bezpośrednie oszacowanie możliwości zagrożenia
porażeniem mikrobiologicznych powietrza wewnętrznego. Metoda ta wykorzystywana w celu sprawdzenia stanu zanieczyszczenia budynku i wyborze odpowiedniego obiek-ty do dalszych badań metodami tradycyjnymi. Najbardziej powszechną techniką wśród metod wczesnego wykrywania jest czujnik gazu nazywane e-nosa.
Technika ta jest stosowana w wielu dziedzinach zastosowań, głównie w branżach medycznych, przemysłu farmaceutycznego, kosmetyce, przemyśle spożywczym i wielu innych obszarach [10].
Matryca wieloczujnikowa stanowi podstawę działania elektronicznego nosa. Jest urządzeniem wieloczujnikowym wykorzystywanym do badań jakości powietrza, służącym również do rozpoznawania określonych produktów ze względu na ich zapach. Wieloczujnikowe analizatory gazów służą do monitoringu specyficznych zanieczyszczeń powietrza. Układy wieloczujnikowe znalazły również zastosowanie w pomiarach stopnia porażenia mikrobiologicznego budynków. Zastosowaniem sys-temów wieloczujnikowych charakteryzują się niższym kosztem aparatury i wykony-wania analiz w porównaniu z chromatografią czy olfaktometrią [14].
Matryca zbudowana jest z płyty, na której umieszcza się kilka, kilkanaście lub kilkadziesiąt różnych czujników, reagujących mało selektywnie na obecność różnych związków lub grup związków chemicznych.
Profile zapachowe z matrycy czujnikowej są wielowymiarowe i musza być anali-zowane za pomocą systemów rozpoznawania wzorca jak np. analiza głównych składowych PCA (Principal Component Analysis), PLS (Partial Least Squares), PDA (Funcional Discriminant Analysis) czy sztuczne sieci neuronowe ANN (Artifficial Neural Network). ANN według niektórych autorów są efektywniejsze do klasyfikacji i predykcji złożonych nieliniowych zbiorów danych niż wielowymiarowe metody analizy [10].
Elektroniczny nos stanowi biologiczną analogię zmysłu powonienia. Szybka detekcja umożliwia rozpoznanie różnych rodzajów cząstek zawartych w otoczeniu lub ich cech tj: zasadowość, kwasowość, specyficzne wiązania chemiczne. Informacje te pozwalają na określenie chemicznego składu otocznia.
E-nosy mogą być konstruowanie przy użyciu różnorakich detektorów w tym detektorów optoelektronicznych i akustoelektronicznych. Zasada działania e-nosa opiera się prymitywnym modelu biologicznego analizatora węchowego. Pole czujników elektronicznego nosa (E-Nose Sensor Field) tworzy zwykle kilkanaście lub kilkadziesiąt różnych elementów. Są to najczęściej czujniki konduktometryczne

Monika Garbacz, Sylwia Magdalena Duda
208
i piezoelektryczne. Istotą działania czujników konduktometrycznych są zmiany elek-trycznego przewodnictwa materiału aktywnego, np. półprzewodzących tlenków metali (MOS, Metal Oxide Sensors) lub niektórych polimerów, zachodzące pod wpływem sorpcji określonych zanieczyszczeń powietrza.
Niezbędne zróżnicowanie wrażliwości poszczególnych czujników – receptorów na różne rodzaje związków chemicznych osiąga się przez wprowadzanie różnych domie-szek do półprzewodzących tlenków metali lub pokrywanie kryształów piezoelek-trycznego kwarcu filmem wysokowrzących cieczy (np. fazy stacjonarne dla GC). Pola sensorów mogą zawierać również inne rodzaje czujników, np. czujniki potencjo-metryczne, optoelektroniczne i akustoelektroniczne lub bioczujniki [14].
5. Podsumowanie
Klimat pomieszczeń, w których człowiek spędza znaczną część swojego czasu (tj. ponad 80% doby), ma znaczny wpływ na jego zdrowie i samopoczucie. Często projektanci i użytkownicy nie zdają sobie sprawy, jak niewłaściwe zaprojektowanie i wykonanie instalacji klimatyzacji czy wentylacji może obniżyć poziom życia i stać się nieoczekiwanym źródłem różnego rodzaju schorzeń i chorób. Syndromu Chorego Budynku uznany za nową jednostkę chorobotwórcza stanowi poważne zagrożenie dla życia i zdrowia ludzi przebywających w pomieszczeniach porażonych mikrobio-logicznie. Prawidłowe zaprojektowanie i wykonanie konstrukcji budowlanych, a także dalsze użytkowanie w higieniczny sposób stanowi podstawę do uwarunkowania właściwej jakości powietrza wewnątrz pomieszczeń.
Literatura
1. Ważnega J., Karysia J. Ochrona budynków przed korozja biologiczną, Wydawnictwo Arkady 2001, s. 53-66.
2. Raw G.J. Sick Building Syndrome: a review of the evidence on causes and solutions, HSE Contract Research Report 1992
3. Pastuszka J.S., Lis D.O., Wlazło A., Łudzeń-Izbińska B. Badanie narażenia na bioaerozole bakteryjne i grzybowe w środowisku mieszkalnym w warunkach Polski, Praca Statusowa Instytutu Medycyny Pracy i Zdrowia Środowiskowego w Sosnowcu, 2001-2004.
4. Zyska. B., Zagrożenia biologiczne w budynkach, Arkady 1999. 5. Mniszek W., Rogiński J. Wady konstrukcyjne budynków przyczyną zagrzybienia
pomieszczeń, Zeszyt Naukowy Wyższej Szkoły Zarządzania Ochrona Pracy W Katowicach, nr 1(3)/2007, s. 31-44.
6. Przybyłowicz E., Słowek G., Wpływ wilgoci na skuteczność ociepleń i mikroklimat wnętrza budynków mieszkalnych, IX Sympozjum Naukowo-Techniczne, Ochrona przed korozją, 10s/A/2007.
7. Belanger K., Bracken M. B., Leaderer B. P. The relation between fungal propagules in indoor air and Home characteristic, Allergy 2001, 56: 419-424.
8. Matkowski K., Materiały budowlane ścian a ryzyko wzrostu grzybów w budynkach, polskie Stowarzyszenie Mykologów Budownictwa, Wrocław 11/2015.
9. Rogiński J. Korozja w budownictwie, WSZOP, Katowice 2005. 10. Szponar B., Mikroby i biotoksyny, czyli co czyha na małych i duzych użytkowników
przestrzeni publicznej, X Ogólnopolska Konferencja Problemy jakości powietrza wewnętrznego w Polsce, Warszawa 2009.
11. Suchorab Z., Sobczuk H., Guz Ł., Łagód G. Gas sensors array as a device to classify mold threat of the buildings, Lublin 2015.

Syndrom chorego budynku oraz jego wpływ na zdrowie mieszkańców w kontekście mikrobiologii
środowiska wewnętrznego
209
12. Suchorab Z., Sobczuk H., Guz Ł., Łagód G. The Possibility of Building Classification for Mould Threat Using Gas Sensors Array, Sensors Lublin 2015.
13. Guntarowska B., Grzyby pleśniowe w środowisku komunalnym i w miejscu pracy - istotne zagrożenia zdrowotne, Med. Pr. 55(3): 257-266.
14. Żukiewicz-Sobczak, W., Sobczak, P., Imbor, K., Kra-sowska, E., Zwoliński, J., Horoch, A., Wojtyła, A. & Piątek, J. 2012. Zagrożenia grzybowe w budynkach i w mieszkaniach – wpływ na organizm człowieka (In Polish), Med. Og. N. Zdrow. 18 (2): 141-
15. Alvarez A.J., Buttner M.P., Use of solid-phase PCR for enhanced detection of air borne microorganisms, Appl.Environ.Microbiol. 60, 374-376.
16. Guz Ł., Metodyczne aspekty pomiaru lotnych zanieczyszczeń powietrza za pomocą urządzenia wieloczujnikowego, Sensors. 15:1-21.
Syndrom chorego budynku oraz jego wpływ na zdrowie mieszkańców w kontekście mikrobiologii środowiska wewnętrznego
Streszczenie W dzisiejszych czasach problem „chorego budynku” stanowi istotne zagadnienie w branży budowlanej. Pojęcie SBS oznacza zwiększony poziom chemicznego i biologicznego zanieczyszczenia powietrza, który negatywnie wpływa na samopoczucie osób przebywających w pomieszczeniach zamkniętych. Jest to jeszcze niezbyt dobrze poznany temat związany z problemami zdrowotnymi, coraz częściej uznawany za nową jednostkę chorobotwórczą. Podstawowym warunkiem niezbędnym do skutecznej analizy zagad-nienia Syndromu Chorego Budynku jest szczegółowe poznanie jego przyczyn oraz konsekwencji nie-sionych dla ludzi. Główną przyczyną pojawienia się Zespołu Chorego Budynku jest wprowadzenie innowacyjnych tech-nologii oraz zastosowanie nowych materiałów do budowy ścian zewnętrznych obiektów budowlanych czy poprawy wydajności wentylacji naturalnej. W wielu przypadkach powodem SBS jest wzrost wilgotności przegród budowlanych, ale również nadmierna wilgotność w pomieszczeniach, która staje sie przyczyna powstawania porażenia mikrobiologicznego. Te czynniki stwarzają odpowiednie warunki do rozwoju pleśni, która wpływa niekorzystnie na trwałość budynku, jego stan techniczny oraz stan zdrowia mieszkańców. Do oceny stopnia porażenia mikrobiologicznego służą metody: mikrobiologiczna, chemiczna oraz szybka detekcja za pomocą matrycy wieloczujnikowej. Matryca wieloczujnikowa stanowi podstawę technologii elektronicznego nosa, który może być także wykorzystywany do oceny jakości powietrza w pomiesz-czeniach zamkniętych. Słowa kluczowe: SBS, chory budynek, e-nos
Syndrome of a sick building and its impact on the health of its inhabitants in the context of microbiology of the internal environment
Abstract Nowadays, the problem of "sick building" is an important issue in the construction industry. The term SBS means increased levels of chemical and biological air pollution that negatively affect the well-being of people in closed rooms. This is not yet well known topic related to health problems, more and more considered as a new disease unit. The basic prerequisite for effective analysis of the Sick Building Syndrome is a thorough understanding of its causes and consequences for people. The main reason for the emergence of the Illness Building Group is the introduction of innovative technologies and the use of new materials for building exterior walls of buildings or improving the efficiency of natural ventilation. In many cases, the cause of SBS is the increase in the humidity of the building partitions, but also the excessive humidity in the rooms that causes the formation of microbial paralysis. These factors create suitable conditions for the development of mold, which adversely affects the durability of the building, its technical condition and the health of its inhabitants. The microbiological, chemical and fast detection methods using a multi-sensor array are used to assess microbial infection. The multi-sensor matrix is the basis of the electronic nose technology, which can also be used to assess indoor air quality. Keywords: SBS, sick building, e-nose

210
Marta Wyzińska1, Jerzy Grabiński
2, Marzena Mikos-Szymańska
3
Porażenie kłosów przez grzyby z rodzaju Fusarium
1. Wstęp
Choroby grzybowe są ważnym czynnikiem ograniczającym plonowanie zbóż.
W ostatnich latach w Polsce jak i na świecie, coraz większego znaczenia nabierają
grzyby z rodzaju Fusarium. Pod względem ekonomicznym, najgroźniejszą chorobą
powodowaną przez grzyby z rodzaju Fusarium jest fuzarioza kłosów. Najczęściej
występuje w pszenicy, ale jest zagrożeniem dla wszystkich zbóż. Szkodliwość jej
polega na obniżeniu plonowania i pogorszeniu jakości ziarna, ale również na
wytworzeniu przez grzyby szkodliwych metabolitów – mikotoksyn.
Na zainfekowanie ziarna pszenicy przez Fusarium spp., oprócz czynników pogodo-
wych i poziomu odporności żywiciela, wpływają także agrotechnika, zmianowanie
roślin, nawożenie i stosowanie środków ochrony roślin.
Celem niniejszej pracy jest przedstawienie przyczyn i skutków porażenia kłosów
zbóż przez grzyby z rodzaju Fusarium na podstawie przeglądu literatury.
2. Fuzarioza kłosów – opis choroby
Fuzarioza kłosów – Fusarium spp., jest najbardziej rozpowszechnioną chorobą
w uprawie pszenicy. Powodowana jest przez grzyby należące do rodzaju Fusarium
głównie F. culmorum i F. graminearum [1, 2]. Grzyby z rodzaju Fusarium, jako
grzyby anamorficzne są nazywane grzybami pleśniowymi (czyli nie tworzą one
zarodników). Powszechnie występują w glebie, szczególnie żyznej, gdzie bytują jako
saprofity, które żywią się martwą materią organiczną. Są one dobrze przystosowane do
zmieniających się warunków atmosferyczno-glebowych i cechują się dużą tolerancją
w stosunku do czynników abiotycznych środowiska, a także rozwijają się w dużym
zakresie temperatur, tj. 0-30°C [3]. Fuzarioza kłosów występuje na wszystkich ga-
tunkach zbóż w naszej strefie klimatycznej (pszenica, żyto, pszenżyto, owies,
jęczmień, kukurydza). Choroba ta, ma największe znaczenie w uprawie pszenicy
(fuzarioza kłosów) [4]. Grzyby z rodzaju Fusarium oprócz fuzariozy kłosów mogą
powodować także szereg innych chorób: zgorzel siewek, zgniliznę korzeni czy zgorzel
podstawy źdźbła [5-9]. Fuzarioza kłosów może powodować znaczne straty plonu
ziarna (od 5 do 60%), głównie przez obniżenie masy tysiąca ziaren. Zainfekowanie
kłosów przez Fusarium skutkuje nie tylko zniżką plonu ale także pogorszeniem jakości
ziarna, poprzez zanieczyszczenie go szkodliwymi dla zdrowia mikotoksynami [10-14].
1 [email protected], Zakład Uprawy Roślin Zbożowych, Instytut Uprawy Nawożenia
i Gleboznawstwa – Państwowy Instytut Badawczy w Puławach, www.iung.pulawy.pl 2 [email protected],Zakład Uprawy Roślin Zbożowych, Instytut Uprawy Nawożenia i Gleboznawstwa
– Państwowy Instytut Badawczy w Puławach, www.iung.pulawy.pl 3 [email protected], Zakład Nawozów, Instytut Nowych Syntez Chemicznych,
www.ins.pulawy.pl

Porażenie kłosów przez grzyby z rodzaju Fusarium
211
W zależności od gatunku, podłoża, na którym rosną, a także warunków hodowli,
grzyby z rodzaju Fusarium mogą różnić się morfologią. Strzępki grzybni występują
obficie, mają watowatą lub śluzowatą strukturę, w kolorach od białej, przez różową,
żółtą, karminową do fioletowej, a nawet ciemnobrązowej. Barwa strzępek jest cechą
gatunkową, ale wpływa na nią również skład podłoża, na którym grzyby rosną, a kon-
kretnie, dostępność cukrów i innych składników pokarmowych [3, 15].
Grzyby z rodzaju Fusarium tworzą trzy typy zarodników: makrokonidia, mikro-
konidia i chlamydospory, które są ważną cechą diagnostyczną. Nazwa sierpik odnosi
się właśnie do kształtu wielokomórkowych, dużych makrokonidiów. Mniejsze, zwykle
jednokomórkowe, rzadko podzielone przegrodami mikrokonidia są owalne, wydłużone
lub cytrynkowate i występują tylko u niektórych gatunków. Chlamydospory uważane
są za trzeci rodzaj zarodników będących jednocześnie organami przetrwalnikowymi
grzyba, wytwarzanymi w okresach niekorzystnych dla jego wzrostu [3, 15].
3. Grzyby z rodzaju Fusarium
Istnieje wiele gatunków grzybów zaliczanych do rodzaju Fusarium. Ze względu na
powszechność występowania na terenie Polski oraz znaczenie dla rolnictwa na uwagę
zasługują następujące grzyby: F. graminearum, F. avenaceum, F. sporotrichioides, F.
poae, F. oxysporum i F. culmorum. Gatunki te różnią się wyglądem grzybni, zasiedla-
nym żywicielem czy produkowanymi mikotoksynami.
F. graminearum poraża głównie zboża drobnoziarniste, kukurydzę i inne trawy.
Jest jednym z dominujących fitopatogenów na terenie Polski, ale występuje także na
innych kontynentach, również w klimacie tropikalnym [3, 16]. Uważany jest za silnie
toksynotwórczy, może produkować zearalenon, fuzarynę C oraz związki z grupy
trichotecenów, które powodują ostre i przewlekłe zatrucia, w niektórych przypadkach
kończące się śmiercią [3].
F. avenaceum występuje na całym świecie i poraża wiele gatunków roślin, m.in.
zboża drobnoziarniste i kukurydzę [15]. Wykazuje słabsze toksyczne działanie niż F.
graminearum, ale przy masowym występowaniu może spowodować duże straty
w rolnictwie. F. avenaceum jest producentem moniliforminy, bowerycyny, fuzaryny C
oraz żółtego barwnika awenaceiny Y o właściwościach antybiotycznych [15, 16].
F. sporotrichioides bytuje głównie w glebie, ale także poraża rośliny, szczególnie
zboża i kukurydzę. Zaliczany jest do grzybów silnie toksynotwórczych, produkuje T-2
toksynę, HT-2 toksynę, neosolaniol i wiele innych pochodnych w dużej ilości [16].
F. poae jest patogenem charakterystycznym dla klimatu umiarkowanego.
Izolowany jest zwykle z owsa, jęczmienia, pszenicy i żyta [17]. Szczepy z gatunku F.
poae odznaczają się niską toksycznością. Mogą produkować w niewielkich ilościach
fuzarynę X, a także substancje z grupy trichotecenów, tj. niwalenol, T-2 toksynę,
diacetoksyscirpenol [16, 17]. F. poae przybierają kolor białoróżowy, karminowoczer-
wony, czasami barwę żółtą do jasnobrązowej. Charakterystyczną cechą tego gatunku
jest zapach kolonii przypominający woń kwiatów lub dojrzałych brzoskwiń. Mikro-
konidia występują obficie, są kuliste, owalne oraz cytrynowate, najczęściej jedno-
komórkowe [3].

Marta Wyzińska, Jerzy Grabiński, Marzena Mikos-Szymańska
212
F. oxysporum występuje powszechnie i poraża wiele roślin m.in. zboża. Spotykany
często jako grzyb towarzyszący innym patogenom z tego rodzaju, tj. F. graminearum
i F. culmorum. Przybiera barwy od białej, przez kremową i różową do purpurowej,
a nawet fioletowej [3]. F. culmorum, jest uważany za gatunek silnie patogenny oraz
wysoce toksyczny. Jako mikroorganizm kosmopolityczny występuje na różnych
szerokościach geograficznych i poraża wiele gatunków roślin, w tym zboża i kuku-
rydzę. Strzępki mają kolor od białych i różowych, do czerwonych, z odcieniem żółtym
do jasnobrązowego [3].
4. Przyczyny i skutki występowania fuzariozy kłosów w zbożach
W ostatnich latach choroby grzybowe zbóż występują w wielu rejonach świata i są
uważane za jeden z głównych czynników wpływających na wielkość i jakość uzys-
kanych plonów. W ciągu całego okresu wegetacji rośliny zbożowe atakowane są przez
wiele patogenów. Za podstawową przyczynę występowania chorób grzybowych
(w tym także fuzariozy kłosów) uważa się ograniczone zmianowanie na polu. Na ogół
zboża uprawiane są po sobie a to skutkuje nagromadzeniem się chorobotwórczych
patogenów w glebie [18-22]. Na występowanie chorób grzybowych mają wpływ także
uproszczenia w uprawie roli [23], wysoki poziom nawożenia azotem oraz uprawa
w monokulturze [24]. Występowanie fuzariozy kłosów zależy od warunków panu-
jących w fazie kłoszenia i kwitnienia pszenicy. Często występujące w tym okresie opa-
dy i związana z nimi wysoka wilgotność powietrza sprzyjają rozwojowi Fusarium spp.
Dużą rolę w rozwoju tej choroby odgrywa genotyp.
Choroby wywołane przez grzyby są niezwykle istotnym czynnikiem biologicznym
wpływającym na wielkość i jakość uzyskanych plonów. Wywołują one szereg niepo-
żądanych skutków, które wpływają na obniżenie plonowania, a także pogarszają jakość
ziarna [24, 25]. Jedną z groźniejszych chorób grzybowych występujących w zasiewach
zbóż jest fuzarioza kłosów. Powoduje obniżenie plonowania, wynikające z mniejszego
ciężaru 1000 ziaren, liczby ziaren z kłosa i masy ziaren z kłosa. Grzyby powodują
zmniejszenie wartości handlowej i konsumpcyjnej upraw poprzez zmianę składu
chemicznego ziarna [26, 27]. Zmiany składu chemicznego ziarna oraz jego właści-
wości fizycznych mogą stanowić podstawę do wykrywania porażenia. Grzyby
z rodzaju Fusarium mają zdolność tworzenia mikotoksyn, skutkiem czego jest aku-
mulacja toksyn w ziarnie jeszcze przed żniwami. Do najbardziej miko toksyno-
twórczych gatunków zalicza się F. graminearum i F. culmorum. W Polsce w ziarnie
zbóż najczęściej występują: deoksyniwalenol, niwalenol, zearalenon i moniliformina
[1, 28, 29].
5. Mikotoksyny fuzaryjne
Mikotoksyny fuzaryjne charakteryzują się różnym stopniem toksyczności. Są
szkodliwe dla ludzi i zwierząt. Do organizmu mogą przenikać przez przewód pokar-
mowy, ale również przez inhalację i przez skórę. Negatywny wpływ na rośliny prze-
jawia się zamieraniem komórek, zahamowaniem wzrostu, chlorozami. Powodują rów-
nież zaburzenia mitozy i mają wpływ na metabolizm białkowy u roślin. Mikotoksyny

Porażenie kłosów przez grzyby z rodzaju Fusarium
213
produkowane są w okresie wegetacji roślin ale również podczas ich zbioru, transportu
i przechowywania [30, 31].
Za najważniejsze miko toksyny fuzaryjne uznano trichoteceny i zearalenon.
Zearalenon (ZEA) tworzony jest głównie przez F. culmorum i F. graminearum. Tri-
choteceny zostały podzielone pod względem budowy na 4 grupy: A, B, C i D. Najpo-
wszechniej występują trichoteceny zaliczone do grupy A (T-2 toksyna, HT-2 toksyna)
i B (deoksyniwalenol, niwalenol, 3-acetylo-DON, 15-acetylo DON, fuzarenon,
trichotecyna).
Mikotoksyny wykazują działanie ogólnoustrojowe, uszkadzają wątrobę, nerki,
powodują choroby serca i układu krwionośnego, prowadzą również do uszkodzeń
skóry i błon śluzowych, wywołują uszkodzenia centralnego układu nerwowego,
powodują obrzęki płuc, zakłócają gospodarkę hormonalną, obniżają odporność
organizmu, a także powodują powstawanie nowotworów [32].
6. Badania dotyczące występowania fuzariozy kłosów w zbożach
Nasilenie występowania fuzariozy kłosów zależy w dużym stopniu od warunków
pogody [33, 34], przedplonu [35, 36], nawożenia azotem oraz sposobu zwalczania
chwastów [37-39]. Według Doohana i in. [40] stopień porażenia kłosów przez
fuzariozę w dużym stopniu zależy od warunków pogodowych (temperatury i wilgot-
ności). Grzyby namnażają się co skutkuje znacznym ubytkiem plonu, a także pogor-
szeniem jego jakości [40-44]. Na rozwój tego patogena wpływa także agrotechnika:
przygotowanie pola, zmianowanie, nawożenie oraz środki ochrony roślin [42, 45-49].
Warunki pogody są czynnikiem, który w większym stopniu determinuje infekcję ziarna
pszenicy przez Fusarium, niż zróżnicowanie systemów uprawy roli. Wyniki badań
uzyskane przez Czabana i in. [50] potwierdzają, że rozprzestrzenianie fuzariozy
kłosów oraz zasiedlenie ziarna pszenicy ozimej przez grzyby z rodzaju Fusarium
zależy głównie od czynników pogodowych. Ocena ryzyka wystąpienia fuzariozy oraz
modele, które mają na celu prognozowanie wystąpienia tej choroby są oparte na
warunkach pogodowych w okresie od kwitnienia do fazy wczesnej dojrzałości
mlecznej [40]. Także technologia uprawy pszenicy ma wpływ na zasiedlenie kłosów
przez Fusarium. Czaban i in. [50] wskazują, że najsilniej porażone były kłosy oraz
ziarno pszenicy pochodzącej z obiektów, gdzie zastosowano intensywną technologię
uprawy. Zasiedlenie ziarna pszenicy orkisz [51] oraz jęczmienia jarego [52] przez
grzyby z rodzaju Fusarium było niższe w przypadku systemu ekologicznego w porów-
naniu do systemów integrowanego i konwencjonalnego.
Badania innych autorów [43, 53] potwierdzają, że warunki pogodowe były
czynnikiem silniej wpływającym na infekcję ziarna przez grzyby z rodzaju Fusarium,
niż zróżnicowanie systemów uprawy. Kiecana i in. [17, 54] stwierdzili, że efektem
zakażenia kłosów pszenicy ozimej w fazie kwitnienia przez Fusarium spp. jest obok
zniżki plonu, zmniejszenie liczby ziaren z kłosa oraz masy 1000 ziaren. Łukanowski
i Sadowski [55] wykazali, że ziarniaki pszenicy ozimej uprawianej w systemie
ekologicznym były znacznie słabiej zasiedlone przez grzyby z rodzaju Fusarium niż
w systemie integrowanym i konwencjonalnym. W innych badaniach [52, 56] zasied-

Marta Wyzińska, Jerzy Grabiński, Marzena Mikos-Szymańska
214
lenie ziarna zbóż przez grzyby z rodzaju Fusarium było również niższe w systemie
ekologicznym w porównaniu do integrowanego i konwencjonalnego.
Wprowadzenie resztek pożniwnych do gleby może zwiększać ryzyko wystąpienia
chorób wywoływanych przez Fusarium. Sallas i Dill-Macky [57, 58] dowodzą, że
wprowadzenie słomy pszenicy do gleby, na głębokość 5 cm sprzyja rozwojowi F.
graminearum. Skutkowało to zwiększeniem inokulum F. graminearum w glebie
i w powietrzu wewnątrz łanu pszenicy w okresie kwitnienia i wczesnej dojrzałości
woskowej, a także powodowało większą infekcję kłosów i ziarniaków.
Stosowanie fungicydów determinuje zasiedlenie ziarna pszenicy przez grzyby
z rodzaju Fusarium. Skuteczność tych substancji w zwalczaniu fuzariozy kłosów,
a także infekcji ziarniaków jest bardzo zmienna [59, 60], ponieważ fungicydy w dużym
stopniu mogą być nieefektywne. Sikora i in. [61] w dwuletnich badaniach (gdzie
infekcja odbyła się w sposób sztuczny) nie stwierdzili obniżenia stopnia zasiedlenia
ziarna pszenicy ozimej przez F. culmorum po zastosowaniu fungicydu Artea 330 EC
(zawierającego propikonazol + cyprokonazol). Gaurilcikiene i in. [59] twierdzą, że
fungicydy zwiększają procent ziarniaków pszenicy zainfekowanych przez Fusarium
spp. Niekorzystne działanie fungicydów może być spowodowane wyeliminowaniem
innych grzybów, przez co grzyby z rodzaju Fusarium mają ułatwiony rozwój [62, 63].
Wiewióra i in. [64] przeprowadzili analizę mikologiczną ziarna różnych gatunków
zbóż pochodzących z uprawy ekologicznej. Badania wykazały, że ziarna były licznie
zasiedlone przez grzyby. Wśród oznaczonych gatunków największy udział miały F.
poae i F. tricinctum, a ponadto na ziarnie owsa F. sporotrichioides i F. avenaceum, na
ziarnie jęczmienia F. avenaceum, a na ziarnie pszenicy F. graminearum.
Należy podkreślić, że porażenie przez Fusarium spp. uzyskane w doświadczeniu
dla ziarna ekologicznego jest niższe w porównaniu do wyników otrzymanych przez
innych badaczy [65, 66] dla innych systemów uprawy. Baturo [65] obserwowała
wysokie porażenie przez grzyby z rodzaju Fusarium ocenione na poziomie 65,9%
w przypadku ziarna pochodzącego z systemu integrowanego i 66,7% dla pocho-
dzącego z uprawy konwencjonalnej. Ponadto donosi ona także, że w zależności od
tego czy ziarno pochodziło z pól produkcyjnych, czy z poletek doświadczalnych
porażenie to wahało się od 22,2 do 66,7% [52]. Sadowski i in. [66] obserwowali mniej
zasiedlonych ziarniaków przez te patogeny w systemie ekologicznym w porównaniu
do innych systemów uprawy.
Góral i in. [67] badali wpływ przedplonu na nasilenie fuzariozy kłosów oraz
zawartość miko toksyn fuzaryjnych w ziarnie pszenicy jarej. Przedplon stanowiła
kukurydza na ziarno oraz rzepak ozimy. W latach, w których wystąpiły warunki
sprzyjające rozwojowi fuzariozy kłosów stwierdzono zwiększone nasilenie choroby na
kłosach pszenicy wysianej po kukurydzy. Ziarno z tych stanowisk zawierało więcej
DNA Fusarium oraz miko toksyn fuzaryjnych w porównaniu do ziarna ze stanowiska
po rzepaku. Dominującym sprawcą fuzariozy kłosów był gatunek F. graminearum.
Uzyskane wyniki wskazują, że uprawa pszenicy na stanowisku, na którym przed-
plonem była kukurydza może zwiększać zagrożenie fuzariozą kłosów. Wynika on
z faktu, że resztki pożniwne kukurydzy czyli duże fragmenty nierozdrobnionych łodyg

Porażenie kłosów przez grzyby z rodzaju Fusarium
215
i kolb są doskonałym miejscem dla rozwoju i przezimowania grzybów z rodzaju
Fusarium.
Rozwój choroby na kłosach pszenicy oraz stężenie mikotoksyn w ziarnie były
bardzo silnie uzależnione od warunków pogodowych. Opady oraz wilgotność względ-
na powietrza w okresie kłoszenia i kwitnienia miała decydujący wpływ na uzyskane
wyniki badań.
7. Podsumowanie
Z dotychczas przeprowadzonych badań wynika, że zarówno sposób uprawy roli jak
i technologia uprawy mają wpływ na występowanie fuzariozy kłosów. Nie bez
znaczenia jest również sposób zagospodarowania słomy na polu. Pozostawienie jej
stwarza warunki do rozwoju wielu patogenów. Występowanie tej choroby w dużym
stopniu zależne jest również od warunków pogodowych w czasie wzrostu i rozwoju
roślin ale również od doboru odmiany do siewu, ponieważ istnieją odmiany mniej lub
bardziej odporne na ten patogen. Fuzarioza kłosów powoduje duże straty w plonie,
a także wpływa na pogorszenie jakości uzyskanego ziarna, poprzez zanieczyszczenie
go szkodliwymi mikotoksynami.
Literatura
1. Bottalico A. Fusarium diseases of cereals: Species complex and related mycotoxin profiles,
European Journal of Plant Pathology, 80 (1998), s. 85-103.
2. Bottalico A., Perrone G. Toxigenic Fusarium species and mycotoxins associated with head
blight in small-grain cereals in Europe, European Journal of Plant Pathology, 108 (2002),
s. 611-624.
3. Wolny-Koładka K. Grzyby z rodzaju Fusarium – występowanie, charakterystyka i
znaczenie w środowisku, KOSMOS Problemy Nauk Biologicznych, Tom 63, 4 (2014):
4. Góral T., Foremska E., Chełkowski J., Arseniuk E. Charakterystyka odmian pszenżyta,
pszenicy i żyta pod względem odporności i tolerancji na porażenie kłosa przez Fusariumspp.,
Biuletyn Instytutu Hodowli i Aklimatyzacji Roślin, 195/196 (1995), s. 251-259.
5. Kiecana J. Fuzarioza kłosów pszenżyta, Roczniki Nauk Rolniczych, Seria E, Ochrona
Roślin, 16(2) (1986), s. 59-67.
6. Łacicowa B., Sułek-Pięta D., Wagner A. Grzyby z rodzaju Fusarium porażające kłosy
pszenicy ozimej, Zeszyty Problemowe Postępów Nauk Rolniczych, 307 (1987), s. 165-173.
7. Mańka M. Patogeniczność wybranych gatunków z rodzaju Fusarium dla siewek zbóż,
Roczniki Akademii Rolniczej, Poznań, Rozprawy Naukowe, 201 (1989), s. 1-63.
8. Bojarczuk M., Krel E., Bojarczuk J. Zgorzel siewek oraz zgnilizna korzeni i podstawy
źdźbła powodowane przez grzyby z rodzaju Fusarium w pszenicy ozimej, Hodowla Roślin
i Aklimatyzacja, 35(5/6) (1991), s. 72-92.
9. Polley R. W., Turner J. A. Surveys of stem base diseases and fusarium ear diseases in
winter wheat in England, Wales and Scotland, 1989-1990, Annals of Applied Biology, 126
(1995), s. 45-59.
10. Kiecana I. Badania nad fuzariozą kłosów jęczmienia jarego (Hordeumvulgare L.) z
uwzględnieniem podatności odmian i zawartości mikotoksyn w ziarnie, Rozprawy
Naukowe, Akademia Rolnicza, Lublin, 161 (1994), s. 1-49.

Marta Wyzińska, Jerzy Grabiński, Marzena Mikos-Szymańska
216
11. Kiecana I., Wojciechowski S., Chełkowski J. Reaction of Winter Wheat Cultivars to
Fusarium avenaceum (Fr.) Sacc. and Fusarium culmorum (W. G. Sm.) under Different
Localities, Polish Agricultural Annual, Seria E, 16(1-2) (1997), S. 61-65.
12. Lemmens M., Krska R., Buerstmayr H., Josephs R., Schumacher R., Grausgruber H.,
Ruckenbauer P. Fusarium head blight reaction and accumulation of deoxynivalenol,
moniliformin and zearalenone in wheat grains, Cereal Research Communications, 31(3-4)
(2003), s. 407-411.
13. Logrieco A., Rizzo A., Ferracane R., Ritieni A. Occurrence of Beauvericin and Enniatins
in Wheat Affected by Fusarium avenaceum Head Blight, Applied and Environmental
Microbiology, 68(1) (2002), s. 82-85.
14. Mirocha C. J., Yu Hui, Evans C. K., Kolaczkowski E. K., Dillmacky R. Chemistry and
physiology of deoxynivalenol in pathogenesis, Cereal Research Comminication, 25(3/1)
(1997), s. 309-313.
15. Kwaśna H., Chełkowski J., Zajkowski P. Grzyby (Mycota). Tom XXII. Sierpik (Fusarium).
Polska Akademia Nauk, Flora Polska. Warszawa (1991).
16. Leslie J. F., Summerell B. A., Bullock S. The Fusarium laboratory manual. Waley-
Blackwell, USA (2006).
17. Kiecana I., Mielcarek R., Perkowski J., Goliński P. Porażenie wiech przez Fusarium poae (Peck)
Wollenw oraz zawartość mikotoksyn w ziarnie owsa, Acta Agrobotanica 58 (2005): 91-102.
18. Bailey K., Lazarovits G. Suppressing soil-borne diseases with residue management and
organic amendmebts, Soil and Tillage Research, 72 (2003), s. 169-180.
19. Bojarczuk M., Bojarczuk J. Fitosanitarna ocena wartości przedplonów roślin zbożowych,
Fragmenta Agronomica, 5(1) (1988), s. 5-24.
20. Colbach N., Huet P. Modeling the frequency and severity of root and foot diseases in winter
wheat monocultures, European Journal of Agronomy, 4 (1995), s. 217-227.
21. Smagacz J., Martyniuk S. Porażenie podstawy źdźbła i korzeni pszenicy ozimej uprawianej
po różnych przedplonach prze patogeny ze szczególnym uwzględnieniem Gaeumannomyces
graminis, Postępy w Ochronie Roślin, 41(2) (2001), s. 745-746.
22. Smiley R. W., Collins H. P., Rasmussen P. E. Diseases of wheat in long-term agronomic
experiments at Pendelton, Oregon, Plant Disease, 80(7) (1996), s. 813-820.
23. Korbas M., Horoszkiewicz-Janka J., Jajor E. Uproszczone systemy uprawy a występowanie
sprawców chorób, Postępy w Ochronie Roślin, 48(4) (2008), s. 1431-1438.
24. Horoszkiewicz-Janka J., Jajor E., Korbas M. Występowanie chorób pszenicy ozimej w
zależności od wybranych czynników agrotechnicznych, Postępy w Ochronie Roślin, 52 (4)
(2012), s. 998-1004.
25. Marks M., Kurowski T. P., Makowski P. Zdrowotność pszenicy ozimej w zależności od
jednorocznego sposobu ugorowania, Fragmenta Agronomica, 4(96) (2007), s. 11-17.
26. Boyacioglu D., Hettiarachchy N. S. Changes In some biochemical componets of wheat grain
that infected with fusarium graminearum, Journal of Cereal Science, 21(1995). s. 57-62.
27. Wąsowicz E. Changes of chemical grain components, especially lipids, during their
deterioration by fungi, In: Cereal Grains, Mycotoxins, Fungi and Quality I During and
Sorag (Eds J. Chełkowski). Elsvier, Amstrdm (1991).s. 259-280.
28. Marasas W. F. O., Nelson P. E., Toussoun T. A. Toxigenic Fusarium species. Idetity and
mycotoxicology, Pensylvania State University Press, University Park, PA, USA(1984).
29. Perkowski J. Badania zawartości toksyn fuzaryjnych w ziarnie zbóż, Roczniki Akademii
Rolniczej w Poznaniu, Rozprawy Naukowe, Zeszyt 295 (1998).

Porażenie kłosów przez grzyby z rodzaju Fusarium
217
30. Creepy E. Update of survey, relation and toxic effects of mycotoxins In Europe. Toxicol,
Lett. 127 (2002). s. 19-28.
31. Packa D. Mikotoksyny zagrożeniem dla ludzi, zwierząt i roślin. Rolnicze ABC 1 (2006).
s. 12-14.
32. Chełkowski J. Mycotoxins, mycotoxigenicfungi and mycotoxicoses, SGGW – AR(1985).
33. Korbas M. Choroby i szkodniki zbóż, Wydawnictwo Multum Poznań (1999), s. 111.
34. Szwejkowski Z., Kurowski T. P. Badania wpływu czynników pogodowych na stopień
inwazyjności patogenów grzybowych w środowisku na przykładzie pszenicy ozimej,
Przegląd Naukowy Inżynieria i Kształtowanie Środowiska, 26(1) (2009), s. 102-108.
35. Kurowski T. P. Mikołajska J., Wojciechowska-Kot. Zdrowotność podstawowych zbóż w
uprawie monokulturowej, [W:] Ekologiczne procesy w monokulturowych uprawach zbóż,
Wydawnictwo Naukowe Uniwersytetu Adama Mickiewicza, Poznań, (1990), s. 223-231.
36. Woźniak A. Studia nad plonowaniem, zachwaszczeniem i zdrowotnością pszenżyta jarego,
pszenicy jarej oraz jęczmienia jarego w płodozmianach i krótkotrwałej monokulturze na
glebie rędzinowej środkowowschodniej Lubelszczyzny, Rozprawy Naukowe Akademii
Rolniczej, Lublin, 247 (2001).
37. Czuba R. Dolistne dokarmianie zbóż azotem w połączeniu ze stosowaniem pestycydów,
Postępy w Ochronie Roślin, 11(1988), s. 12-15.
38. Kurowski T. P., Rogalski L., Czajka W. Zdrowotność i plonowanie roślin zbożowych opryski-
wanych roztworem mocznikowo-fungicydowym, Materiały Sympozium „Biotyczne środo-
wisko uprawne a zagrożenie chorobowe roślin”, 7-9 września, Olsztyn, (1993), s. 255-262.
39. Woźniak A. Wpływ przedplonów na plonowanie, zachwaszczenie i zdrowotność pszenżyta
jarego, Biuletyn Instytutu Hodowli i Aklimatyzacji Roślin, 221 (2002), s. 35-43.
40. Doohan F. M., Brennan J., Cooke B. M. Influence of climatic factors on Fusarium species
pathogenic to cereals, European Journal of Plant Pathology, 109 (2003), s: 755-768.
41. Arseniuk E., Góral T. Fuzarioza kłosów – czynniki sprawcze i gospodarcze znaczenie
choroby, IV Forum Zbożowe, www.pin.org.pl/hrin/txt/2005/3-6.rtf, (2005).
42. Champeil A., Doré T., Fourbet J. F. Fusarium head blight: epidemiological origin of the
effects of cultural practices on head blight attacks and the production of micotoxins by
Fusarium in wheat grains, Plant Science, 166 (2004), s. 1389-1415.
43. Parry D. W., Jenkinson P., McLeod L. Fusarium ear blight (scab) in small grains –
areview, Plant Pathology, 44 (1995), s. 207-238.
44. Xu X. M., Parry D. W., Nicholson P., Thomsett A. A., Simpson D., Edwards S. G., Cooke
B. M., Doohan F. M., Brennan J. M., Moretti A., Tocco G., Mule G., Hornok L., Giczey
G., Tatnell J. Predominance and association of pathogenic fungi causing Fusarium ear
blight in wheat in four European countries, European Journal of Plant Pathology,112
(2005), s. 143-154.
45. Edwards S. G. Influence of agricultural practices on fusarium infection of cereals and
subsequent contamination of grain by trichothecene mycotoxins, Toxicology Letters, 153
(2004), s: 29-35.
46. Jalli M., Parikka P. Fusarium infection of heads and stems under different cultivation
practices, Proceedings of the COST SUSVAR Fusarium workshop: Fusarium deseases in
cereals – potential impact from sustainable cropping systems, 1-2 June, 2007 Valence,
Hungary, s: 38-40.
47. Łukanowski A., Sadowski C. Settlement of spring wheat kernels by Fusarium spp. In
organic systems as compared with other cropping systems, 3rd International FHB
Symposium, Szeged, Hungary, (2008), s: 581-583.

Marta Wyzińska, Jerzy Grabiński, Marzena Mikos-Szymańska
218
48. Pirgozliev S.R., Edwards S.G., Hare M.C. Jenkinson P. Strategies for the control of Fusarium
head blight in cereals, European Journal of Plant Pathology, 109 (2003), s: 731-742.
49. Podolska G. Kształtowanie cech jakościowych ziarna pszenicy poprzez technologię
produkcji, Studia i Raporty IUNG – PIB, 9 (2007), S: 55-64.
50. Czaban J., Wróblewska B., Sułek A., Podolska G. Wpływ różnych technologii produkcji
pszenicy ozimej na zasiedlenie jej ziarna przez grzyby z rodzaju Fusarium, Polish Journal
of Agronomy, 5 (2011), s. 11-20.
51. Kurowski T. P., Wysocka U. Fungi colonizing grain of winter spelt grown under two
production systems, Phytopathologia, 54 (2009), s: 45-52.
52. Baturo A., Łukanowski A., Kuś J. Comparison of health status of winter wheat and spring
barley grain cultivated in organic, integrated and conventional systems and monoculture,
Proceedings of the First World Conference on Organic Seed “Challenges and Opportunities
of Organic Agriculture and the Sees Industry”, 5-7 July, 2004, FAO Headquarters, Rome,
Italy, s: 128-132.
53. Bierzele B., Meier A., Hindorf H., Krӓmer J., Dehne H. W. Epidemiology of Fusarium
infection and deoxynivalenol content in winter wheat in the Rhineland, Germany. European
Journal of Plant Pathology. 108 (2002). s. 667-673.
54. Kiecana I., Mielniczuk E., Cegiełko M. Badania podatności kłosow pszenicy ozimej
(Triticumaestivum L.) na porażenie przez Fusarium avenaceum (Fr.) Sacc. Acta
Agrophysica. 8 (2006).s. 629-636.
55. Łukanowski A., Sadowski C. Settlement of grain of winter wheat grown in organic and
other systems by fusarium spp. and determination of their potential micotoxigenic ability
with PCR assay, 3 rd International Seed Health Conference of Seed Pathology
Microorganisms on Seeds – Harmfulness and Control Book of Abstracts, 6-8 września
Bydgoszcz (2006). s. 29-30.
56. Kurowski T. P., Wysocka U. Fungi colonizing grain of winter spelt grown under two
production systems, Phytopathologia. 54 (2009). s. 45-52.
57. Sallas B., Dill-Macky R. Colonization of Wheat Cultivars Fusarium graminearum at
Harvest and in Overwintered Residues. in Proceedings of the 2005 National Fusarium
Heat Blight Forum Milwaukee, Wisconsin 11-13 December. (2005a). s. 140-143.
58. Sallas B., DillMacky R. Effect of Residue Menagement and Host Resistance on the
Epidemiology of Fusarium Heat Blight. in Proceedings of the 2005 National Fusarium
Heat Blight Forum Milwaukee, Wisconsin 11-13 December. (2005b). s. 144-147. 2005b
59. Gaurilčikiene I. Butkute B., Mankevičiene A. A multi aspect comparative investigation on
thr use of strobilurin and triazole – based fungicides for winter wheat disease control, in
Odile Carisse (ed) Fungicides, InTech (2010). s. 69-94.
60. Henriksen B. Elen O. Natural Fusarium grain infection level in wheat, barley and oat after
application of fungicides and herbicides, Journal of Phytopathology. 153 (2005). s. 214-220.
61. Sikora H. Banachowska J., Doleżych D. Badania nad ochroną pszenicy ozimej przed
fuzariozą kłosów wywołaną przez Fusarium culmorum w warunkach sztucznej infekcji,
Progres of Plant Protection/ Postępy w Ochronie Roślin. 47(2007). S. 314-318.
62. Müllenborn C., Steiner U., Ludwig M., Oerke E. C. Effect of fungicides of the complex of
Fusarium species and saprophytic fungi colonizing wheat kernels, European Journal of
Plant Pathlogy. 120 (2008). s. 157-166.
63. Muthomi J. W., Riungu G. M., Ndung’u J. K., Narla R D. African Crop Science
Conference Proceedings. 8 (2007). s. 863-867.

Porażenie kłosów przez grzyby z rodzaju Fusarium
219
64. Wiewióra B., Mańkowski D., Bulińska-Radomska Z. Zdrowotność ziarna zbóż
pochodzącego z ekologicznej produkcji nasiennej, Biuletyn Instytutu Hodowli i
Aklimatyzacji Roślin. 251 (2009). s. 29-39.
65. Baturo A. Head Healthiness and fungus composition of spring barley harvested grain
cultivated under organic, integrated and conventional farming systems, Phytopathology
Journal. 26 (2002). s. 73-83.
66. Sadowski C., Lenc L., Kuś J. Fuzarioza kłosow I grzyby rodzaju Fusarium zasiedlające
ziarno pszenicy orkisz uprawianych w systemie ekologicznym, Journal of Research and
Applications In Agricultural Engineering. 55 (2008). s. 79-83.
67. Góral T., Ochodzki P., Walentyn-Góral D., Justesen A. F., Jorgensen L. N. Wpływ
przedplonu oraz warunków pogodowych na porażenie kłosów pszenicy jarej przez grzyby
z rodzaju Fusarium oraz zawartość mikotoksyn w ziarnie, Biuletyn Instytutu Hodowli
i Aklimatyzacji Roślin. 265 (2012). s. 11-21.
Porażenie kłosów przez grzyby z rodzaju Fusarium
Streszczenie
Niniejsza praca dotyczy porażenia kłosów przez grzyby z rodzaju Fusarium. Opisano grzyby wywołujące powstawanie fuzariozy kłosów, mikotoksyny, które są związkami produkowanymi przez te grzyby, a także
przyczyny i skutki tej choroby. Występowanie fuzariozy uzależnione jest przede wszystkim od warunków
pogody, ale również od sposobu uprawy, efektywności zastosowanych fungicydów i odporności gospo-
darza. Do skutków fuzariozy kłosów należy zaliczyć obniżenie poziomu plonowania, a także pogorszenie jakości ziarna. Jest to choroba, która dotyka wszystkie gatunki zbóż.
Słowa kluczowe: fuzarioza kłosów, choroby grzybowe, mikotoksyny, zboża
Fusarium fungi infections of cereal ears
Abstract
This paper deals with ear infections caused by Fusarium fungi. Fungi causing the formation of Fusarium
head blight (FHB) and mycotoxins that are compounds produced by these fungi, and also causes and effects of this disease have been described. The occurrence of FHB incidence depends mainly on weather
conditions but also on the agronomic practices, effectiveness of fungicides used and host resistance.
Negative effects of Fusarium head blight are the decrease in yielding and the deterioration of grain quality.
It is the disease that affects all cereal species. Keywords: Fusarium head blight, fungi disease, mycotoxins, cereals

220
Wiktor Pszczółkowski1, Zdzisława Romanowska-Duda
2, Agata Pszczółkowska
3
Porównanie wrażliwości i dynamiki wzrostu dwóch
szczepów Chlorella sp. hodowanych na odciekach
po fermentacji metanowej z biogazowni rolniczej
1. Wprowadzenie
Na świecie prowadzone są intensywne prace nad rozwojem technologii produkcji
biomasy jednokomórkowych glonów na skalę przemysłową. Działania te mają na celu
maksymalne wykorzystanie potencjału mikroglonów, w tym jako źródła cennych
składników chemicznych, biopaliw i pożywienia. Glony będąc organizmami fotoauto-
troficznymi wiążą dwutlenek węgla z atmosfery, a w uprawie masowej mogą
utylizować CO2 bezpośrednio z dymów i gazów wylotowych z instalacji przemys-
łowych. Niektóre gatunki mikroglonów mogą wykorzystywać do wzrostu składniki
odżywcze znajdujące się w ściekach, lub innych odpadach organicznych. W niniejszej
pracy zaproponowano wykorzystanie odpadów poprodukcyjnych z biogazowi jako
taniego surowca do produkcji podłoży hodowlanych do uprawy mikroglonów.
2. Wstęp
Równowaga, właściwe proporcje i dostępność pierwiastków biogennych takich jak
azot czy fosfor mają bezpośredni wpływ na funkcjonowanie wszystkich ekosystemów.
Od czasów rewolucji przemysłowej człowiek w istotny sposób zmienił obieg tych
pierwiastków w przyrodzie. Zanieczyszczenie wód pierwiastkami biogennymi pocho-
dzącymi ze spływów i ścieków, aktywności rolniczej czy przemysłowej jest poważ-
nym problemem środowiskowym. Eutrofizacja wód powoduje szkody w ekosystemach
i gospodarce, prowadząc m. in. i do zakwitów wód, wyczerpania zasobów rozpusz-
czonego tlenu, śmierci organizmów wodnych takich jak ryby, ale również raf koralo-
wych [1-5]. Ponadto uniemożliwia wykorzystanie akwenów wodnych do celów rolni-
czych, rekreacyjnych, przemysłowych, a w dłuższym czasie prowadzi do zarastania
zbiorników. Szczególnym problemem jest zanieczyszczenie zbiorników stanowiących
źródło wody pitnej, której deficyt odnotowuje się coraz częściej na świecie. Rozwią-
zanie problemu wymaga znajomości technik oczyszczania, mających szerokie zastoso-
wanie w przywracaniu równowagi ekologicznej ekosystemów wodnych. W wielu
przypadkach dwustopniowe oczyszczanie nie usuwa wystarczającej ilości biogenów ze
ścieków i konieczne jest stosowanie dodatkowo trzeciego etapu [1]. Uprawa mikro-
1 [email protected], Pracownia Ekofizjologii Roślin, Wydział Biologii i Ochrony
Środowiska, Uniwersytet Łódzki, www.biol.uni.lodz.pl 2 [email protected], Pracownia Ekofizjologii Roślin, Wydział Biologii i Ochrony Środowiska,
Uniwersytet Łódzki, www.biol.uni.lodz.pl 3 [email protected], Pracownia Ekofizjologii Roślin, Wydział Biologii i Ochrony
Środowiska, Uniwersytet Łódzki, www.biol.uni.lodz.pl

Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp.
hodowanych na odciekach po fermentacji metanowej z biogazowni rolniczej
221
glonów może być alternatywnym sposobem doczyszczania ścieków komunalnych,
rolniczych, a nawet przemysłowych [6-8]. Technologia usuwania zanieczyszczeń
i związków biogennych przy pomocy glonów określana jest jako phykoremediacja.
Potencjalnie duży wpływ na rozkład zanieczyszczeń w ściekach może mieć wydziela-
nie przez mikroglony tlenu – produktu fotosyntezy, który stymuluje wzrost i metabo-
lizm mikroorganizmów aerobowych, produkujących enzymy zdolne do rozkładu
zanieczyszczeń organicznych. Zastosowanie mikroglonów pozwala ograniczyć
mechaniczne natlenianie ścieków, tym samym ograniczyć uwalnianie z nich związków
lotnych i zmniejszyć nakłady energii [9].
Glony są obecnie uważane za najważniejszy bio-surowiec przyszłości. Wiele
mikroglonów charakteryzują się wysokim zapotrzebowaniem na makroelementy azot
i fosfor. Ta cecha może być potencjalnie wykorzystana do oczyszczania ścieków i in-
nych bioodpadów z tych pierwiastków. Aktywność fotosyntetyczna wymaga dostar-
czenia komórkom dwutlenku węgla, który może pochodzić z atmosfery jak również
z gazów wylotowych. Niezależnie od źródła składników pokarmowych wytwarzanie
biomasy przez mikroglony związane jest z sekwestracją dwutlenku węgla. Jest to
pożądane zjawisko, ponieważ zwiększające się w wyniku działalności człowieka stę-
żenia dwutlenku węgla i innych gazów cieplarnianych w atmosferze, powoduje wzrost
temperatury na naszej planecie, a w konsekwencji zmiany w funkcjonowaniu global-
nego klimatu.
Chlorella to rodzaj jednokomórkowych zielenic, o kulistym kształcie, syntety-
zujących chlorofil a i b. Wiele gatunków charakteryzuje się wysoką efektywnością
fotosyntetyczną i produktywnością biomasy oraz możliwością wzrostu w trudnych
warunkach.
Biomasa glonów może zostać wykorzystana jako źródło energii odnawialnej, po-
przez m. in. przetworzenie na biopaliwa płynne, zgazowanie biomasy lub wytworzenie
z niej biogazu.
Zastosowania te są jednak przy obecnym stanie wiedzy i techniki w większości
przypadków nieopłacalne. Głównymi przyczynami jest wysoki koszt produkcji bio-
masy glonów i konieczność wysokich nakładów inwestycyjnych w infrastrukturę do
hodowli glonów w porównaniu do łatwiej dostępnych i tańszych odpadów organicz-
nych, komunalnych, z rolnictwa czy leśnictwa, które z powodzeniem mogą być wyko-
rzystane jako substrat do generowania energii elektrycznej i ciepła.
Połączenie oczyszczania ścieków i odpadów organicznych z sekwestracją dwutlen-
ku węgla oraz utylizacją związków azotu i siarki pochodzących z gazów wylotowych
wydaje się być obiecującą metodą znacznego obniżenia kosztów produkcji biomasy
mikroglonów. W efekcie można otrzymać biomasę o bardzo niskiej zawartości lignin
i celulozy, związków trudnych do hydrolizy w procesach biologicznych mającą
szerokie zastosowania [5].
Wiktor Pszczółkowski, Zdzisława Romanowska-Duda, Agata Pszczółkowska
222
Rysunek 2. Sposoby wykorzystania biomasy mikroglonów do produkcji biopaliw
[10 zmodyfikowany]

Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp.
hodowanych na odciekach po fermentacji metanowej z biogazowni rolniczej
223
Pomysł ten jest wprost zaczerpnięty ze zjawisk zachodzących na naszej planecie.
Zanieczyszczenie wód i podwyższona zawartość dwutlenku węgla i innych gazów
cieplarnianych prowadzi do powstawania zakwitów glonów czyli gwałtownego
namnażania się sinic i mikroglonów skutkującego wytworzeniem dużych ilości
biomasy [4].
3. Cel pracy
Celem badań było określenie możliwości wykorzystania odcieku pochodzącego
z produkcji biogazu jako źródła składników odżywczych dla dwóch szczepów mikro-
glonów z rodzaju Chlorella sp. hodowanych w skali laboratoryjnej.
4. Materiały i metody
4.1. Mikroglony
W badaniach wykorzystano dwa szczepy zielenic z rodzaju Chlorella sp.
o symbolach PER1 i PER2 pochodzące z kolekcji kultur Pracowni Ekofizjologii
Roślin, Wydziału Biologii i Ochrony Środowiska, Uniwersytetu Łódzkiego. Szczepy
zostały wyizolowane ze sztucznych zbiorników słodkowodnych o różnej trofii. Szczep
PER1 z Zalewu Sulejowskiego (woj. Łódzkie), a szczep PER2 z Zbiornika Przykona
(gmina Przykona, powiat turecki, woj. Wielkopolskie) utworzonego w ramach re
kultywacji wyrobisk Kopalni Węgla Brunatnego Adamów. Do przygotowania inoku-
lum oba szczepy uprawiano w aksenicznych kulturach in vitro w 250 ml kolbach
Erlenmeyera na podłożu Z8. Uprawę prowadzono w pomieszczeniu hodowlanym
z zachowaniem stałej temperatury 26±1°C oraz fotoperiodu: 16h światło 8h ciemności.
Gęstość komórek określano na podstawie wyników pomiaru gęstości optycznej (OD)
przy długościach fali λ=680 nm (absorpcja chlorofilu i rozproszenie światła) i λ=720
nm (rozproszenie światła) [11].
4.2. Odpad z biogazowni
W badaniach wykorzystano odpad pochodzący ze zintegrowanej instalacji do
produkcji etanolu, energii elektrycznej i cieplnej oraz nawozów organicznych zloka-
lizowanego w Piaszczynie w gminie Miastko w powiecie Bytowski, województwo:
Pomorskie (54°00'57.5"N 17°09'59.4"E). Z biogazowni pobrano jednorazowo repre-
zentatywną partię odcieku po fermentacji metanowej przeprowadzonej na odpadzie
z gorzelni. Badanym odciekiem była płynna frakcja odpadu pofermentacyjnego
otrzymana poprzez mechaniczną separację. Materiał do badań przechowywany był
w szczelnym zbiorniku z polipropylenu w temperaturze 4±1°C.
4.3. Warunki uprawy kultur in vitro
W doświadczeniu badano wzrost dwóch szczepów Chlorella sp. na podłożach
przygotowanych poprzez rozcieńczenie odcieku po fermentacji metanowej do stężenia
5, 10, 15, 20 i 30%, wodą destylowaną. Odciek nie był w żaden sposób modyfiko-
wany, ani sanityzowany/sterylizowany. W wariancie kontrolnym oraz do inokulacji
wykorzystano kultury in vitro uprawiane na podłożu Z8 zgodnie z Scandinavian

Wiktor Pszczółkowski, Zdzisława Romanowska-Duda, Agata Pszczółkowska
224
Culture Collection for Algae & Protozoa [12]. Hodowlę wyjściową porcjowano do
probówek wirowniczych i wirowano przy RFC=700 x g. Po usunięciu supernatantu,
osad (komórki) zawieszano w 100 ml podłoża zależnie od wariantu doświadczalnego.
Doświadczenie prowadzono w 250 ml kolbach Erlenmeyera w pomieszczeniu
hodowlanym z zachowaniem stałej temperatury 26±1°C oraz fotoperiodu: 16h światło
8h ciemności. Każdy wariant eksperymentalny był powtórzony pięciokrotnie
w osobnych kolbach.
Tabela 2 Skład medium Z8
Składnik Zawartość
[mg/l]
Składnik
(Gafron’s trace elements)
Zawartość
[µg/l]
Azot (N) 87,0 Na2WO4·H2O 3,3
Fosfor (P) 5,6 (NH4)6Mo7O24·4H2O 8,8
Potas (K) 14,0 KBr 12,0
Magnez (Mg) 2,4 KJ 8,3
Siarka (S) 3,2 ZnSO4·7H2O 28,7
Wapń (Ca) 14,0 Cd(NO3)2·4H2O 15,5
Sód (Na) 135,7 Co(NO3)2·6H2O 14,6
Chlor (Cl) 10,9 CuSO4·5H2O 12,5
Żelazo (Fe) 5,7 NiSO4(NH4)2SO4·6H2O 19,8
Cr(NO3)3·9H2O 4,1
V2O5 0,9
Al2(SO4)3K2SO4·24H2O 47,4
H3BO3 310,0
MnSO4·4H2O 223,0
4.4. Pomiar gęstości optycznej
Wzrost kultur monitorowano poprzez pomiar absorbancji za pomocą fluorymetru
AquaPen-C AP-C 100. Wszystkie warianty eksperymentalne były próbkowane w taki
sam sposób. Po dokładnym wymieszaniu ruchem okrężnym hodowli w kolbie,
pobrano 3 ml zawiesiny komórek. Przed każdym pomiarem urządzenie zerowano
wobec próby nie zawierającej mikroglonów. W celu wyeliminowania błędu wyni-
kającego z mętności podłoży hodowlanych próbę do zerowania przygotowano każdo-
razowo bezpośrednio przed pomiarem. Z pobranych 3 ml połowa objętości była wiro-
wana przy RFC=700 x g, a supernatant stosowano do zerowania fluorymentru. Na
pozostałych 1,5ml próby wykonywano po trzy pomiary. Za każdym razem mieszając
zawartość kuwety pomiarowej przez jej odwrócenie do góry dnem. Pomiary wyko-
nywano w dniu założenia doświadczenia (dzień „0”), w celu potwierdzenia braku róż-
nic w gęstości optycznej pomiędzy wszystkimi wariantami eksperymentalnymi.
Kolejne pomiary wykonano w 7, 14, 21 i 28 dniu hodowli.
4.5. Oznaczenie świeżej i suchej biomasy
Po zakończeniu hodowli określono suchą i świeżą masę oraz objętość pozostałych
po doświadczeniu kultur, które następnie przefiltrowano przy użyciu zestawu do

Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp.
hodowanych na odciekach po fermentacji metanowej z biogazowni rolniczej
225
filtracji próżniowej przez zważone filtry o średnicy porów 0,2 µm. Filtry zważono
ponownie i w ten sposób określono świeżą biomasę kultur. Suchą biomasę oznaczono
po inkubacji świeżej biomasy wraz z membraną filtracyjną pozostawioną w termo-
stacie przez 2 h w 50°C. Wyniki masy przeliczono na 1 litr hodowli.
4.6. Analiza statystyczna wyników
Analizę statystyczną wykonano w programie Statistica 13 (StatSoft). Po spraw-
dzeniu normalności rozkładów w poszczególnych grupach i jednorodności wariancji
pomiędzy porównywanymi grupami testem Levene’a wyniki poddano jednoczyn-
nikowej analizie wariancji ANOVA dla prób niezależnych. Jeżeli stwierdzono wystę-
powanie różnic pomiędzy średnimi wykonano test post hoc rozsądnej istotnej różnicy
(RIR) Tukey’a.
5. Wyniki i dyskusja
Funkcjonowanie biogazowni związane jest z generowaniem znacznych ilości
biomasy pofermentacyjnej. Dlatego istnieje pilna potrzeba zagospodarowanie tego
odpadu, który wśród społeczeństwa budzi wiele wątpliwości. Obawy te wynikają
w dużym stopniu z braku wiedzy m. in. w zakresie wpływu pofermentacyjnej biomasy
odpadowej na plonowanie roślin, a także właściwości fizyko-chemiczne i biologiczne
gleb. Istnieje konieczność wskazania poprzez badania naukowe oraz praktykę produ-
centów sposobu zagospodarowania tego substratu, którego podstawowym kierunkiem
powinno być wykorzystanie w celach nawozowych do uprawy roślin na glebach
słabych i zdegradowanych, jak również hodowli kultur in vitro, w tym glonów, co jest
przedmiotem niniejszej pracy. Glony w tym z rodzaju Chlorella są doskonałym
materiałem do pozyskania cennych substancji biotechnologicznych, energii i biopaliw
trzeciej generacji, a także mogą być zastosowane jako nawóz do uprawy roślin [13-15].
W prezentowanych badaniach wykazano pozytywny wpływ odcieku po fermentacji
metanowej na wzrost i rozwój dwóch szczepów Chlorella sp. (PER1 i PER2).
Badane szczepy Chlorella sp. wzrastały na podłożach zawierających od 5% do 30%
odcieku po fermentacji metanowej [Rysunek 3]. Na każdym podłożu zawierającym
odciek, mierzono wzrost kultur mikroglonów jako gęstość optyczną i był niższy niż na
podłożu kontrolnym Z8. Dla szczepu Chlorella sp. PER1 wzrastającego na podłożu
zawierającym 5% odcieku gęstość optyczna mierzona przy dł. fali λ=680 nm w 7 dniu
uprawy, była niższa o 8%, podczas gdy nie wykazano różnicy w gęstości optycznej
przy dł. fali 720 nm w stosunku do podłoża Z8. W kolejnych dniach wykonywania
pomiaru wartości dla obu gęstości optycznych były niższe (p<0,001). Odnotowano
w 14 dniu pomniejszone o 9% (dla 680 nm i 720 nm), w 21 dniu o 31% (680 nm)
i 24% (720 nm), a w 28 dniu o 47% i 41% odpowiednio dla fali 680 nm i 720nm.
Podobną zależność zaobserwowano w przypadku kultur szczepu Chlorella sp. PER2,
uprawianych na podłożu zawierającym 5% odcieku po fermentacji metanowej. W 7
dniu gęstość optyczna mierzona przy dł. fali 680 nm była niższa o 11% podczas gdy
przy dł. fali 720 nm nie wykazano różnicy gęstości w stosunku do kultur wzrastających
na podłożu kontrolnym Z8. W kolejnych dniach uprawy gęstość kultur była niższa

Wiktor Pszczółkowski, Zdzisława Romanowska-Duda, Agata Pszczółkowska
226
(p<0,001) odpowiednio w dniu 14 o 28% (680 nm) i 29% (720 nm), w dniu 21 o 50%
i 45%, a w dniu 28 o 65% i 62% odpowiednio dla 680 nm i 720 nm [Rysunek 3].
Rezultat ten może świadczyć o wyczerpaniu łatwo dostępnych składników odżyw-
czych i spowolnieniu wzrostu na podłożu zawierającym 5% odcieku w przypadku
obydwu szczepów.
Dla obu szczepów Chlorella sp. (PER1 i PER2) 7 dnia hodowli nie wykazano róż-
nic we wzroście na podłożach zawierających 10% i 15% odcieku. Dla szczepu PER2
14 dnia nie wykazano różnicy we wzroście na podłożach zawierających 5% i 15%
odcieku (pomiar przy 680 i 720nm), a 21 dnia na podłożach zawierających 10% i 15%
odcieku (pomiar przy 680 nm). Dla szczepu PER1 w 21 dniu nie wykazano różnicy
w gęstości optycznej pomiędzy kulturami uprawianymi na podłożach zawierających
5%, 10% i 15% odcieku (680 nm). W 28 dniu hodowli gęstość optyczna (dł. fali 680)
dla kultur szczepu PER1, na podłożach zawierających 10%, 15%, 20% i 30% była
niższa (p<0,001) odpowiednio o 31%, 22%, 61% i 81%, jak również dla dł. fali 720
nm uległa obniżeniu odpowiednio o 34%, 29%, 62% i 84% w stosunku do kultur
uprawianych na podłożu Z8. Dla kultur szczepu PER2 wzrastających na podłożach
zawierających 10%, 15%, 20% i 30% w 28 dniu hodowli gęstość optyczna (dł. fali
680) była niższa (p<0,001) odpowiednio o 33%, 25%, 58% i 92%, a dla dł. fali 720 nm
pomniejszona odpowiednio o 35%, 26%, 55% i 92% w stosunku do kultur upra-
wianych na podłożu Z8 [Rysunek 3].
Dla obu szczepów najbardziej efektywny wzrost kultur odnotowano na podłożu
zawierającym 15% odcieku po fermentacji metanowej, a najsłabszy w obecności 30%
tego odpadu. Przy czym Szczep PER1 wzrastał lepiej na podłożu zawierającym 10%
i 15% odcieku, a szczep PER2 na podłożach zawierających od 10-20%. Na podłożach
zaobserwowano spadek mętności medium w czasie, co najprawdopodobniej Przyspie-
szyło namnażanie się komórek w wyniku zwiększonej dostępności światła i spowodo-
wało wzrost aktywności fotosyntetycznej. Wysoka mętność podłoży zawierających
30% odcieku prawdopodobnie była przyczyną słabego wzrostu i rozwoju komórek obu
szczepów glonów. Argumentem za tą hipotezą jest fakt, że nie nastąpiła całkowita
inhibicja wzrostu jak to miało miejsce w przypadku innych badanych szczepów (dane
niepublikowane), a jedynie spowolnienie wzrostu w stosunku do niższych stężeń
odcieku. Wpływ mętności odcieku na wzrost kultur był istotny również ze względu na
statyczny charakter uprawy, gdzie część komórek osiadła na dnie kolb hodowlanych co
zmniejszyło dostępność promieniowania aktywnego fotosyntetycznego (PAR).
Analiza wyników pomiarów świeżej i suchej biomasy kultur szczepu PER1 wyka-
zała brak różnic pomiędzy kulturami uprawianymi na podłożu zawierającym 5 i 10%
odcieku. W przypadku suchej masy nie wykazano również różnic pomiędzy plonem na
podłożu zawierającym 15% odcieku, a plonem suchej masy na podłożu kontrolnym
Z8, podczas gdy na pozostałych podłożach zawierających 5%, 10%, 20% i 30%
odcieku plon suchej masy był niższy odpowiednio o 44%, 32%, 62%, 81%
w odniesieniu do podłoża Z8 [Rysunek 4].
Dla szczepu PER2 nie wykazano różnic w zawartości świeżej i suchej biomasy
pomiędzy kulturami uprawianymi na podłożu zawierającym 5% i 20% odcieku. Nie

Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp.
hodowanych na odciekach po fermentacji metanowej z biogazowni rolniczej
227
wykazano również różnic w zawartości świeżej biomasy pomiędzy kulturami upra-
wianymi na podłożu zawierającym 10% i 15% odcieku. Kultury szczepu PER2 upra-
wiane na podłożach zawierających 5%, 10%, 15%, 20% i 30% odcieku charakter-
ryzowały się niższym odpowiednio o 63%, 29%, 17%, 65% i 93% plonem suchej
masy w stosunku do kultur uprawianych na podłożu Z8 [Rysunek 5].
Rezultaty badań jednoznacznie wskazały, że dla obu szczepów Chlorella sp.
stężenie 15% odcieku w podłożu było optymalne i zapewniło najlepszy wzrost
i rozwój kultur oraz największy plon suchej masy. Szczególnie widoczne to było dla
szczepu PER1, gdzie nie wykazano różnicy w wartości suchej masy kultur hodowa-
nych na podłożu kontrolnym Z8, a medium zawierającym 15% odcieku po fermentacji
metanowej [Rysunek 4]. Może to oznaczać, że szczep ten jest w stanie wzrastać na
podłożu złożonym wyłącznie z odcieku i wody z taką samą wydajnością jak na
medium hodowlanym Z8 (kontrolnym). Tym samym możliwe jest prowadzenie pro-
cesu fykoremediacji odcieku w obecności szczepu PER1 z jednoczesną produkcją
wartościowej biomasy o szerokich możliwościach zastosowania takich jak produkcji
energii, barwników, pasz oraz nawozów[5, 6, 15-18]. Badania Zuliani i wsp. [11]
wykazały, że 3% dodatek odcieku pofermentacyjnego do standardowego podłoża
hodowlanego zmniejszał produktywność kultury mierzoną jako przyrost biomasy,
zwiększając jednocześnie produktywność białek, lipidów i karotenoidów. Autorzy [11]
wskazali również, że wyższe stężenia odcieku (10 i 20%) w medium hodowlanym
hamowały wzrost glonów z rodzaju Chlorella i Scenedesmus. Niezbędne są dalsze
badania w celu wykazania czy efektywność wzrostu szczepów PER1 i PER2 pozosta-
nie niezmieniona podczas uprawy w większej skali. Zastosowanie podłoża zawiera-
jącego odciek w hodowli w fotobioreaktorze, dałoby potencjalnie znacznie większy
przyrost biomasy w krótszym czasie [11, 19]. Nawet w przypadku uzyskania niższego
plonu suchej biomasy w stosunku do konwencjonalnych podłoży, proces może okazać
się opłacalny ekonomicznie ze względu na niemal zerowe koszty substratu. Åkerström
i wsp. [20] uzyskali wzrost mikroglonów z rodzaju Chlorella sp. w fotobioreaktorze
z suplementacji CO2 na odcieku w stężeniu do 100% efektywnie usuwając azot i fosfor
z podłoża. Największą produktywność biomasy uzyskali przy stężeniu 40-50%
odcieku rozcieńczonego wodą z oczyszczalni co sugeruje, że zastosowanie fotobio-
reaktora może doprowadzić w przyszłości do zwiększenia produktywności szczepów
PER1 i PER2 w stosunku do uprawy w warunkach in vitro.
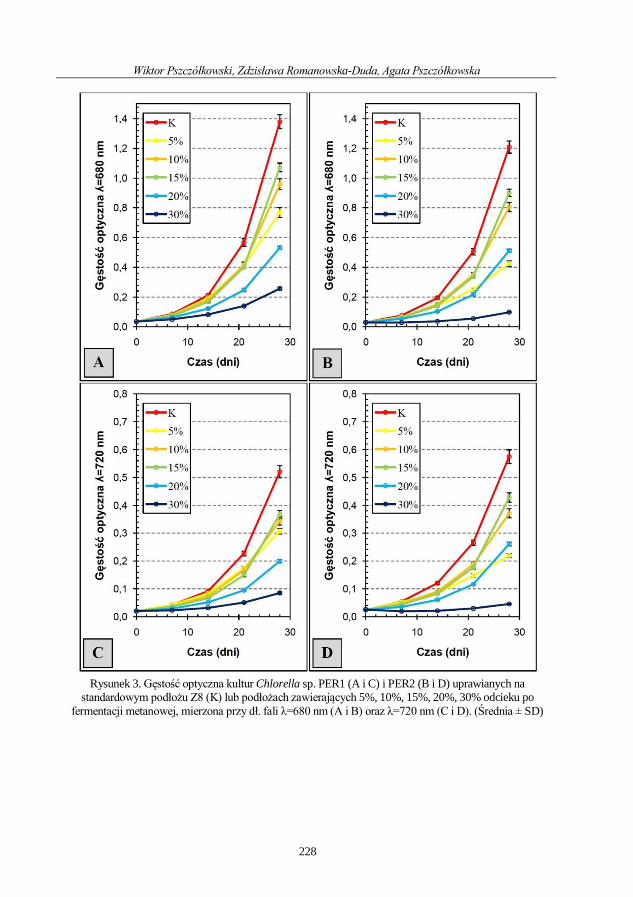
Wiktor Pszczółkowski, Zdzisława Romanowska-Duda, Agata Pszczółkowska
228
Rysunek 3. Gęstość optyczna kultur Chlorella sp. PER1 (A i C) i PER2 (B i D) uprawianych na standardowym podłożu Z8 (K) lub podłożach zawierających 5%, 10%, 15%, 20%, 30% odcieku po
fermentacji metanowej, mierzona przy dł. fali λ=680 nm (A i B) oraz λ=720 nm (C i D). (Średnia ± SD)
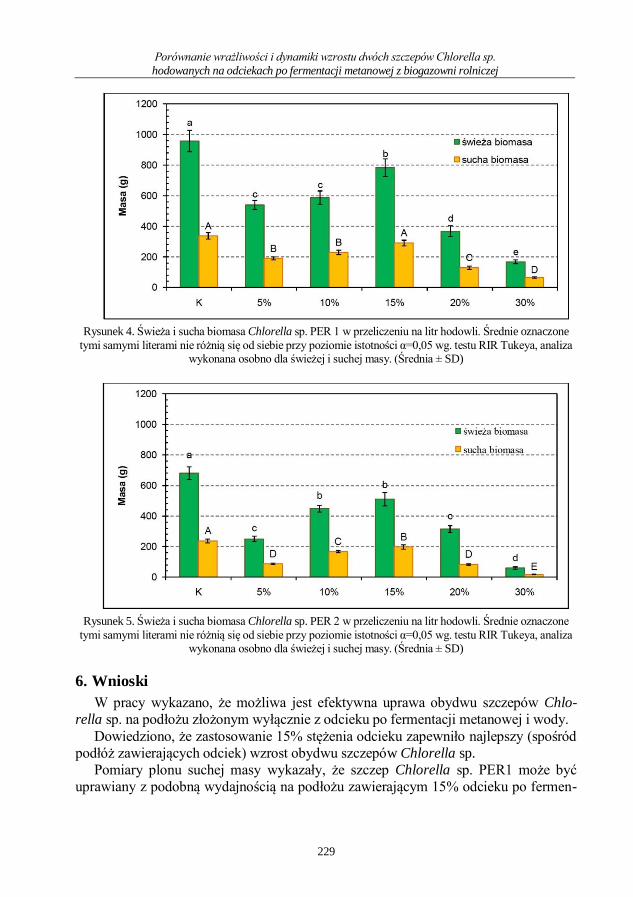
Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp.
hodowanych na odciekach po fermentacji metanowej z biogazowni rolniczej
229
Rysunek 4. Świeża i sucha biomasa Chlorella sp. PER 1 w przeliczeniu na litr hodowli. Średnie oznaczone
tymi samymi literami nie różnią się od siebie przy poziomie istotności α=0,05 wg. testu RIR Tukeya, analiza wykonana osobno dla świeżej i suchej masy. (Średnia ± SD)
Rysunek 5. Świeża i sucha biomasa Chlorella sp. PER 2 w przeliczeniu na litr hodowli. Średnie oznaczone
tymi samymi literami nie różnią się od siebie przy poziomie istotności α=0,05 wg. testu RIR Tukeya, analiza wykonana osobno dla świeżej i suchej masy. (Średnia ± SD)
6. Wnioski
W pracy wykazano, że możliwa jest efektywna uprawa obydwu szczepów Chlo-
rella sp. na podłożu złożonym wyłącznie z odcieku po fermentacji metanowej i wody.
Dowiedziono, że zastosowanie 15% stężenia odcieku zapewniło najlepszy (spośród
podłóż zawierających odciek) wzrost obydwu szczepów Chlorella sp.
Pomiary plonu suchej masy wykazały, że szczep Chlorella sp. PER1 może być
uprawiany z podobną wydajnością na podłożu zawierającym 15% odcieku po fermen-

Wiktor Pszczółkowski, Zdzisława Romanowska-Duda, Agata Pszczółkowska
230
tacji metanowej jak na standardowym podłożu Z8, co może znacznie obniżyć koszty
hodowli na skalę komercyjną.
Niezbędne są dalsze badania nad wykorzystaniem odcieku w uprawie glonów
w zwiększonej skali w celu zweryfikowania danych uzyskanych w doświadczeniach
laboratoryjnych.
Planowane są dalsze badania obejmujące kinetykę zmiany składu chemicznego
odcieku pofermentacyjnego w trakcie hodowli mikroglonów, a także charakterystykę
uzyskanej biomasy w zakresie zawartości barwników i innych wybranych składników
o potencjalnym zastosowaniu biotechnologiczym.
Wskazane są również dalsze prace badawcze w celu opracowania opłacalnych
i skutecznych systemów usuwania substancji odżywczych z biomasy odcieków
organicznych po fermentacji z biogazowni. Wyzwaniem będzie połączenie pewnej
podstawowej wiedzy z zakresu kinetyki wzrostu glonów, stosowanie niedrogich me-
diów hodowlanych, a także strategii pozyskiwania szczepów glonów o unikalnych ce-
chach w celu opracowania efektywnych biotechnologii.
Uwagi ogólne/Podziękowania
Badania sfinansowane przez Narodowe Centrum Badań i Rozwoju: BIOSTRATEG
2, Grant nr. 2/296369/5/NCBR/2016
Literatura
1. Abdel-Raouf N., Al-Homaidan A.A., Ibraheem I.B.M., Microalgae and wastewater
treatment. Saudi J. Biol. Sci., 19 (2012), s. 257-275.
2. Altieri A.H., Harrison S.B., Seemann J., Collin R., Diaz R.J., Knowlton N., Tropical dead
zones and mass mortalities on coral reefs. Proc. Natl. Acad. Sci., 114 (2017), s. 3660-3665.
3. Andersen J.H., Carstensen J., Conley D.J., Dromph K., Fleming-Lehtinen V., Gustafsson
B.G., Josefson A.B., Norkko A., Villnäs A., Murray C., Long-term temporal and spatial
trends in eutrophication status of the Baltic Sea, Eutrophication in the Baltic Sea. Biol.
Rev., 92 (2017), s. 135-149.
4. Bender-Champ D., Diaz-Pulido G., Dove S., Effects of elevated nutrients and CO 2
emission scenarios on three coral reef macroalgae. Harmful Algae, 65 (2017), s. 40-51.
5. Calicioglu O., Demirer G.N., Integrated nutrient removal and biogas production by
Chlorella vulgaris cultures. J. Renew. Sustain. Energy, 7 (2015), s. 033123.
6. Li C., Yang H., Li Y., Cheng L., Zhang M., Zhang L., Wang W., Novel bioconversions of
municipal effluent and CO2 into protein riched Chlorella vulgaris biomass. Bioresour.
Technol., 132 (2013), s. 171-177.
7. Wang L., Min M., Li Y., Chen P., Chen Y., Liu Y., Wang Y., Ruan R., Cultivation of
Green Algae Chlorella sp. in Different Wastewaters from Municipal Wastewater Treatment
Plant. Appl. Biochem. Biotechnol., 162 (2010), s. 1174-1186.
8. Wang L., Wang Y., Chen P., Ruan R., Semi-continuous Cultivation of Chlorella vulgaris
for Treating Undigested and Digested Dairy Manures. Appl. Biochem. Biotechnol., 162
(2010), s. 2324-2332.
9. Cakirlar B., Ogut G., Erguder T., Kaymak B., Demirer G., Uludag-Demirer S., Parallel
Nutrient Removal and Biomitigation of CO2 by Pure (Chlorella vulgaris) and Mixed Algal
Cultures. Int. Sci. J. Environ. Sci., 3 (2014), online.

Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp.
hodowanych na odciekach po fermentacji metanowej z biogazowni rolniczej
231
10. Alam F., Mobin S., Chowdhury H., Third generation biofuel from Algae Procedia Engineering 105 (2015) 763-768.
11. Zuliani L., Frison N., Jelic A., Fatone F., Bolzonella D., Ballottari M., Microalgae Cultivation on Anaerobic Digestate of Municipal Wastewater, Sewage Sludge and Agro-Waste. Int. J. Mol. Sci. 17, 1692 (2016), [online].
12. Scandinavian Culture Collection for Algae & Protozoa, Z8 [online]. http,//www.sccap.dk/media/freshwater/7.asp (16.6.2017),.
13. El-Sharony T.F., El-Gioushy S.F. and Amin O.A., Effect of Foliar Application with Algae and Plant Extracts on Growth, Yield and Fruit Quality of Fruitful Mango Trees Cv. Fagri Kalan. J Horticulture, 2, 4 (2015) [online].
14. Marks E.A.E., Minon J., Pascual A., Montero O., Navas L. M., Rad C., Application of a microalgal slury to soil stimulates heterotrophic activity and promotes bacterial growth. Science of the Total Environment 605-606 (2017) 610-617.
15. Grzesik M., Romanowska-Duda Z., Kalaji H.M., Effectiveness of cyanobacteria and green algae in enhancing the photosynthetic performance and growth of willow (Salix viminalis L.) plants under limited synthetic fertilizers application. Photosynthetica 55, 3 (2017) 510-521.
16. Salati S., D’Imporzano G., Menin B., Veronesi D., Scaglia B., Abbruscato P., Mariani P., Adani F., Mixotrophic cultivation of Chlorella for local protein production using agro-food by-products. Bioresour. Technol., 230 (2017), s. 82-89.
17. Feng Y., Li C., Zhang D., Lipid production of Chlorella vulgaris cultured in artificial wastewater medium. Bioresour. Technol., 102 (2011), s. 101-105.
18. Mahdy A., Ballesteros M., Gonzalez-Fernandez C., Enzymatic pretreatment of Chlorella vulgaris for biogas production, Influence of urban wastewater as a sole nutrient source on macromolecular profile and biocatalyst efficiency. Bioresour. Technol., 199 (2016), s. 319-325.
19. Benavente-Valdes J.R., Mendez-Zavala A., Morales-Oyervides L., Chisti Y., Montanez J., Effects of shear rate, photoautotrophy and photoheterotrophy on production of biomass and pigments by Chlorella vulgaris, Shear rate effect on Chlorella vulgaris cultures. J. Chem. Technol. Biotechnol., (2017), [online].
20. Åkerström, A.M., Mortensen, L.M., Rusten, B., Gislerød, H.R. Biomass production and nutrient removal by Chlorella sp. As affected by sludge liquor concentration. J. Environ. Manag. (2014), 144, 118-124.
Porównanie wrażliwości i dynamiki wzrostu dwóch szczepów Chlorella sp. hodowanych na odciekach po fermentacji metanowej z biogazowni rolniczej
Streszczenie: Na świecie prowadzone są intensywne prace nad rozwojem technologii produkcji biomasy jednokomór-kowych glonów na skalę przemysłową. Działania te mają na celu maksymalne wykorzystanie potencjału mikroglonów, jako źródła cennych składników chemicznych, biopaliw i pożywienia. W niniejszej pracy zaproponowano wykorzystanie odpadów poprodukcyjnych z biogazowni jako taniego surowca do pro-dukcji podłoży hodowlanych do uprawy mikroglonów. Celem badań było określenie możliwości wyko-rzystania odcieku po produkcji biogazu jako źródła składników odżywczych dla mikroglonów. Wyko-rzystano dwa szczepy z rodzaju Chlorella sp.. Hodowlę prowadzono w kolbach Erlenmeyera w temp. 26±1°C z zastosowaniem fotoperiodu 16 h światła, 8 h ciemności. Jako podłoża posłużyły rozcieńczony odciek po fermentacji metanowej (5, 10 15, 20 i 30%) oraz podłoże mineralne Z8 (kontrola). W trakcie hodowli wykonano pomiary gęstości optycznej kultur i wyznaczono krzywe dynamiki wzrostu. Na zakończenie hodowli oznaczono świeżą i suchą biomasę. Rezultaty badań wykazały znaczące różnice w dynamice wzrostu pomiędzy badanymi wariantami stężeń odcieku, na których hodowane były mikro-glony. Największy wzrost mikroglonów, porównywalny ze wzrostem na podłożu kontrolnym, odnotowano na 15% odcieku. Odciek może zostać wykorzystany jako substrat w kulturach iv vitro. Niezbędne są dalsze badania by zastosować odciek po fermentacji metanowej do uprawy mikroglonów w większej skali. Słowa kluczowe: Chlorella, odciek po produkcji biogazu

Wiktor Pszczółkowski, Zdzisława Romanowska-Duda, Agata Pszczółkowska
232
Comparison of sensitivity and growth dynamics of two Chlorella sp. strains
cultured in liquid fraction of post-fermentation wastes from agricultural biogas
plant
Abstract
Intensive work is being carried out to develop an industrial-scale unicellular algae biomass production technology. These actions aims for maximal utilization of microalgae, as a source of valuable chemical
components, biofuels and food. This paper proposes to use post-production waste from biogas plant as an
inexpensive raw material for the production of culture media for growing microalgae. The purpose of the
study was to determine the potential of utilization of liquid fraction of wastes from the biogas plant as a source of nutrients for microalgae. Two strains of the genus Chlorella sp. were used. The culture was
carried out in Erlenmeyer flasks at 26±1°C with photoperiod 16 h light, 8 h dark. The diluted wastes from
methane fermentation (5, 10 15, 20 and 30%) and a full standard Z8 mineral medium (control) were used
as a substrates. Optical density of cultures were used to determine growth dynamics. At the end of the culture the fresh and dry biomass was measured. The results of the study showed significant differences in
growth dynamics between wastes concentrations on which microalgae were grown. The highest growth
rate of microalgae, comparable to the growth on control medium, was recorded on medium with 15% of
wastes from methane fermentation. The liquid fraction of wastes form biogas plant can be used as a growth substrate in iv vitro cultures. More research is needed to use wastes from methane fermentation in large
scale microalgae production.
Keywords: Chlorella, biogas post-fermentation wastes

233
Agata Pszczółkowska1, Zdzisława Romanowska-Duda
2, Wiktor Pszczółkowski
3
Potencjał wykorzystania odpadu pofermentacyjnego
z biogazowni rolniczej jako substratu w produkcji
podłoża hodowlanego do uprawy cyanobacteria
1. Wprowadzenie
Rozwój gospodarczy wywiera duży wpływ na środowisko poprzez m. in. eksplo-
atację zasobów naturalnych, generowanie zanieczyszczeń i odpadów przemysłowych.
Problem dotyczy całego świata, jednak dewastacja środowiska naturalnego jest
charakterystyczna przede wszystkim dla krajów rozwijających się. W krajach wysoko
rozwiniętych od wielu lat próbuje się znaleźć rozwiązania prowadzące do zrówno-
ważonego rozwoju i przeciwdziałające negatywnym skutkom dla środowiska [1].
Produkcja biomasy mikroglonów jest jedną z intensywniej rozwijanych technologii
ze względu na potencjalne korzyści ekonomiczne związane z substancjami natural-
nymi, produkowanymi przez te organizmy oraz korzyści dla środowiska, wynikające
z redukcji emisji gazów cieplarnianych. Mniej uwagi poświęca się grupie fotoautotro-
ficznych mikroorganizmów prokariotycznych. Wśród przedstawicieli sinic wiele
gatunków (nietoksycznych) posiada zdolność do produkcji użytecznych związków
chemicznych, które z powodzeniem mogą być wykorzystywane m. in. w produkcji
rolniczej. Sinice ze względu na bardzo wydajną fotosyntezę mogą wiązać duże ilości
dwutlenku węgla z atmosfery [2]. Aby produkcja biomasy jednokomórkowych glonów
miała dodatni bilans finansowy i energetyczny, poszukuje się tanich i łatwo dostępnych
surowców będących źródłem substancji odżywczych.
Celem badań było zaadaptowanie dwóch nietoksycznych szczepów cyanobacteria
z rodzaju Anabaena sp. do podłoża hodowlanego przygotowanego na bazie odpadu
pofermentacyjnego, pochodzącego z biogazowni w Piaszczynie.
2. Wstęp
Mianem glony początkowo określano grupę organizmów zawierających chlorofil
i bytujących w środowisku wodnym. Termin ten był używany jako nazwa taksono-
miczna. Wraz z rozwojem metod identyfikacji i postępem wiedzy stracił taką rangę
i stał się nazwą zwyczajową odnoszącą się do organizmów pochodzących z wielu
różnych linii ewolucyjnych, które łączy szereg cech morfologicznych i ekologicznych [3].
1 [email protected], Pracownia Ekofizjologii Roślin, Wydział Biologii i Ochrony
Środowiska, Uniwersytet Łódzki, www.biol.uni.lodz.pl 2 [email protected], Pracownia Ekofizjologii Roślin, Wydział Biologii i Ochrony Środowiska,
Uniwersytet Łódzki, www.biol.uni.lodz.pl 3 [email protected], Pracownia Ekofizjologii Roślin, Wydział Biologii i Ochrony
Środowiska, Uniwersytet Łódzki, www.biol.uni.lodz.pl

Agata Pszczółkowska, Zdzisława Romanowska-Duda, Wiktor Pszczółkowski
234
Glony, nazywane również algami, od anglojęzycznej nazwy algae łączy foto-
autotrofizm i wodne środowisko życia, choć i od tych głównych cech bywają wyjątki.
Grupa ta zawiera organizmy eukariotyczne (Alveolata, Chlorarachniophyta, Chloro-
phyta, Cryptophyta, Euglenophyta, Glaucocystophyta, Haptophyta, Heterokonta,
Rhodophyta) i prokariotyczne (Cyanobacteriaceae) [4]. W jej skład wchodzą mikro-
skopijne organizmy występujące w postaci pojedynczych komórek, luźnych lub
przejściowych konglomeratów, nitkowatych lub rozgałęzionych kolonii, mogących
tworzyć maty, lub bardziej złożone formy przypominające liście u roślin. Ponad to do
grupy należą organizmy makroskopowe przypominające swym wyglądem rośliny,
choć w rzeczywistości ich ciała tworzy plecha [3]
Prokariotycznymi przedstawicielami glonów są sinice zwane również cyjanobakte-
riami (ang. Blue-Green Algae). Są to organizmy jednokomórkowe, mimo to biomasa
wielu przedstawicieli sinic może być obserwowana makroskopowo ze względu na
zdolność do tworzenia kolonii i zakwitów w wodach słodkich i słonych [3]. Niektóre
gatunki tworzące kolonie są w stanie wyspecjalizować komórki do tzw. heterocyst –
o grubych ścianach, w których wytwarzana jest nitrogenaza, enzym katalizujący
reakcję asymilacji azotu cząsteczkowego (N2) i przekształcenia do jonów amonowych
(NH3+
). Struktura heterocyst umożliwia utrzymanie warunków anaerobowych i chroni
tym samym nitrogenezę przed inhibicją ze strony tlenu cząsteczkowego. Ta wyjątkowa
cecha pozwala na wiązanie azotu atmosferycznego niezależnie od warunków środo-
wiska i oddziaływań symbiotycznych. W heterocystach fotosynteza została drastycznie
ograniczona by wewnątrz panowały warunki beztlenowe, niezbędne do zachodzenia
diazotrofii. Przedstawicielem sinic diazotroficznych są gatunki z rodzaju Anabaena [5].
Wykorzystanie sinic do oczyszczania ścieków jest stosunkowo nowym pomysłem.
W latach 40-tych i 50-tych XX wieku pojawiła się koncepcja stosowania organizmów
fotoautotroficznych w tym sinic do przetwarzania ścieków [6, 7]. Praktyczne wyko-
rzystanie koncepcji miało miejsce w latach 80-tych [8]. Cyanobacteria są zdolne do
usuwania ze ścieków: amoniaku, azotanów, ortofosforanów i niektórych metali cięż-
kich. Dzięki tym właściwością są przydatne w trzeciorzędowym oczyszczaniu ścieków
komunalnych, rolniczych i przemysłowych, ograniczając eutrofizację zbiorników
naturalnych i akumulowanie metali ciężkich w organizmach wodnych i osadach
dennych.
Cyanobacteria pełnią istotne funkcje w naturalnych ekosystemach. Są to organizmy
kosmopolityczne występujące niemal w każdej szerokości geograficznej. Ze względu
na specyficzne wymagania i adaptację poszczególnych gatunków oraz ich rozmiesz-
czenie, skład jakościowy i ilościowy, ekologia czy okresowość występowania
znacząco się różnią.
Jednym z największych problemów związanych z wykorzystaniem biomasy
mikroorganizmów w oczyszczaniu ścieków jest oddzielenie biomasy po zakończeniu
procesu. Jednym z rozwiązań jest stosowanie biomasy unieruchomionej. Gatunki
drobnoustrojów obecne w odpadach są zdolne do wzrostu i efektywnego usuwania
z nich zanieczyszczeń. Zarówno sinice jak i inne drobnoustroje w obecności odpadu

Potencjał wykorzystania odpadu pofermentacyjnego z biogazowni rolniczej jako substratu w produkcji podłoża hodowlanego do uprawy cyanobacteria
235
narażone są na stres środowiskowy. Wynika on między innymi z zaburzenia równo-
wagi mikro i makroelementów, obecności czynników toksycznych, niekorzystnych
właściwości fizyko-chemicznych (pH, natlenienie, zawartość dwutlenku węgla,
temperatura, potencjał oksydoredukcyjny itp.), a w przypadku organizmów foto-
autotroficznych również mętność i absorpcja promieniowania fotosyntetycznego przez
składniki odpadu. Jedną z dróg poszukiwania gatunków sinic mogących wzrastać
w ściekach są badania koncentrujące się na bioróżnorodności miejsc zanieczysz-
czonych lub samych ściekach, które będą poddane procesowi oczyszczania. Wielu
autorów w swoich badaniach poszukuje odpowiednich gatunków sinic, wykorzystując
ścieki z rafinerii, fabryki nawozów, piwowarstwie, gorzelnictwie, produkcji barwni-
ków, papieru lub zakładach farmaceutycznych [9, 10].
Nieoczyszczone lub niewystarczająco oczyszczone ścieki, zawierające znaczną
ilości pierwiastków biogennych, stanowią zagrożenie dla ekosystemów wodnych [11].
Zastosowanie sinic jako bioremediatorów jest technologią przyjazną dla środowiska
i ekonomicznie opłacalną w stosunku do powszechnie stosowanych w oczyszczalniach
aktywnych osadów czynnych. Sinice posiadają szereg przydatnych do tego celu cech [12]:
mają niewielkie wymagania pokarmowe, dzięki czemu ścieki bogate w azot
i fosfor oraz podstawowe makro i mikroelementy mogą służyć jako podłoże do
wzrostu,
są organizmami fotoautotroficznymi, wiążą dwutlenek węgla z atmosfery, lub
innych źródeł takich jak gazy wylotowe, przez co nie potrzebują innych źródeł
węgla, a przy tym mogą ograniczać emisje gazów cieplarnianych,
prowadząc fotosyntezę wydzielają tlen, który może stymulować wzrost mikro-
organizmów heterotroficznych, mających zdolność degradacji substancji orga-
nicznych, tym samym pośrednio prowadzić do rozkładu związków, których same
sinice nie są w stanie metabolizować;
mogą spełniać podwójną rolę w bioremediacji ścieków oraz generując biomasę
o szerokim, spektrum potencjalnych zastosowań: jako pasza, bionawóz, substrat
do produkcji biopaliw czy źródło cennych składników chemicznych;
niektóre gatunki wiążą azot atmosferyczny co może mieć kluczowe znaczenie
w oczyszczaniu ścieków o wysokiej zawartości fosforu w stosunku do azotu.
3. Materiał i metody
3.1. Cyanobacteria
Anabaena PER 1 i Anabaena PER 2 to dwa szczepy diazotroficznych sinic
pochodzące z kolekcji kultur Pracowni Ekofizjologii Roślin, UŁ. Obydwa szczepy są
nietoksyczne i mogą być zastosowane jako bionawóz. Szczepy stanowiące inokulum
do badań uprawiano w 250 ml kolbach Erlenmeyera na podłożu BG-110 w pomiesz-
czeniu hodowlanym z zachowaniem stałej temperatury 25±1°C oraz fotoperiodu: 16h
światło 8h ciemności. Do przygotowania kultur o założonej gęstości komórkowej

Agata Pszczółkowska, Zdzisława Romanowska-Duda, Wiktor Pszczółkowski
236
wykorzystywano pomiar gęstości optycznej przy dwóch długościach fali λ=680
i λ=720.
3.2. Materiał odpadowy z przemysłu
Do badań użyto odpad pochodzący z biogazowi w Piaszczynie. Wykorzystano
odciek po fermentacji metanowej czyli płynną frakcję otrzymaną po separacji masy
pofermentacyjnej.
3.3. Warianty kontrolne
Kontrolne warianty doświadczalne uprawiano na podłożu BG-110 lub 5, 10 i 15%
roztworach odcieku pofermentacyjnego w wodzie destylowanej Rys. 1.
Rysunek 6 Warianty kontrolne. Schemat przedstawia procentową zawartość trzech wariantów badanego podłoża
3.4. Warunki hodowli
Doświadczenia prowadzono w pomieszczeniu hodowlanym z zachowaniem stałej
temperatury 25±1°C oraz stałym natężeniu światła i fotoperiodzie: 16h światło 8h
ciemności. Jako podłoża stosowano mieszaninę podłoża BG-110, odcieku oraz wody
destylowanej. Badania prowadzono w 100 ml kolbach Erlenmeyera.
3.5. Przebieg doświadczenia
Doświadczenie prowadzono w czerech etapach, stopniowo zwiększając stężenie
odpadu pofermentacyjnego w podłożu hodowlanym od 0% (kultura wyjściowa) do
15%, jednocześnie redukując objętość standardowego medium BG-110 od 100%
(kultura wyjściowa) do 0% (Tab.1). Każdorazowo w celu uzyskania jednolitej gęstości
komórkowej, pobierano hodowlę o znanej liczbę komórek (obliczaną na podstawie
gęstości optycznej) potrzebną w doświadczeniu. Następnie wirowano hodowlę przy
RFC=700 x g, supernatant usuwano a zwirowane komórki zawieszano w nowej porcji
podłoża. Po uzyskaniu homogenicznej zawiesiny kulturę porcjowano do kolb
o pojemności 100 ml. Każda kolba stanowiła jedno powtórzenie wariantu ekspery-
mentalnego. W ten sposób uzyskano szereg powtórzeń o takiej samej wyjściowej
wartości gęstości optycznej (bez istotnych różnic statystycznych).

Potencjał wykorzystania odpadu pofermentacyjnego z biogazowni rolniczej jako substratu w produkcji podłoża hodowlanego do uprawy cyanobacteria
237
Tabela 1. Procentowa zawartość trzech komponentów podłoża w czterech etapach doświadczenia.
BG-110 odciek H2Odest.
I etap 60% 5% 35%
II etap 30% 10% 60%
III etap 10% 15% 75%
IV etap 0% 15% 85%
Źródło: Opracowanie własne
3.6. Pomiar gęstości optycznej
Do oceny kinetyki wzrostu cyanobacteria zastosowano metodę pomiaru absorbancji
za pomocą fluorymetru AquaPen-C AP-C 100. Każdorazowo pomiary były wyko-
nywane w dniu t0 w celu potwierdzenia jednolitej gęstości optycznej we wszystkich
wariantach eksperymentalnych, a następnie po 7, 14, 21 i 28 dniach hodowli. W celu
wyeliminowania błędu pomiarowego wynikającego z mętności podłoży hodowlanych,
przed pomiarami z każdego wariantu pobierano próbę i wirowano ją przy
RFC=700 x g. Supernatant uzyskany z poszczególnych wariantów eksperymentalnych
służył to zerowania fluorymentru przed pomiarem gęstości optycznej hodowli.
3.7. Analiza statystyczna
Wyniki zostały poddane analizie statystycznej w programie Statistica wersja 13
(StatSoft). Zastosowano jednoczynnikową analizę wariancji ANOVA dla prób nieza-
leżnych. Dla grup wyników, w których stwierdzono różnice pomiędzy średnimi wyko-
nano analizę post hoc rozsądnej istotnej różnicy (RIR) Tukey’a.
4. Wyniki i dyskusja
Kultury cyanobacteria pasażowane z podłoża BG-110 bezpośrednio na podłoża
składające się z 5, 10 lub 15% odpadu po fermentacji metanowej oraz wody desty-
lowanej nie rozwijały się i następowało ich stopniowe zamieranie (Wyk.1A, Wyk. 2A).
Od 7 dnia wszystkie kultury obydwu szczepów Anabaena sp. hodowane na odcieku
z wodą destylowaną miały niższą gęstość optyczną w porównaniu do kultury kontrol-
nej hodowanej na medium BG-110 (p<0,001).
W przypadku obu szczepów cyanobacteria odnotowano wzrost na podłożu za-
wierającym 60% BG-110 i 5% odcieku po fermentacji metanowej. Po 28 dniach
pierwszego pasażu na podłożu zawierającym 60% BG-110 i 5% odcieku kultury
Anabaena sp. PER1 i PER2 miały o 49% i 41% odpowiednio niższą gęstość optyczną
od kultur kontrolnych (p<0,001). Przez cały okres wzrostu gęstość optyczna była
istotnie (p<0,001) niższa niż wariantów kontrolnych. Po drugim pasażu do 7 dnia ho-
dowli nie wykazano różnic w gęstości optycznej kultur Anabaena sp. PER2 wzrasta-
jących na podłożu 60% BG-110 i 5% odcieku, a kulturami na podłożu BG-110. Po 28
dniach drugiego pasażu kultury Anabaena sp. PER1 i PER2 miały o 20% i 9% niższą
gęstość od kultur kontrolnych (p>0,001). Do 14 dnia po trzecim pasażu na podłoże
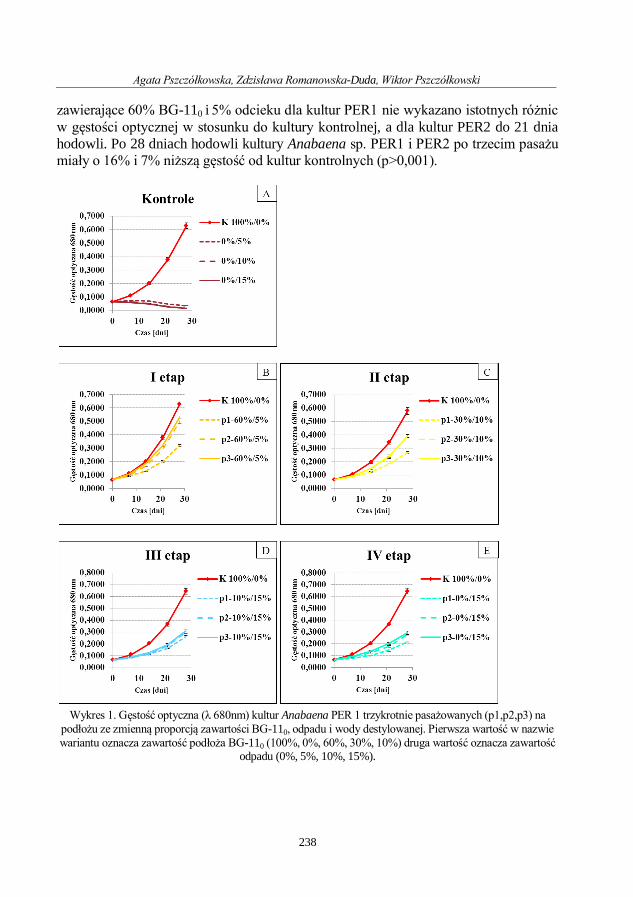
Agata Pszczółkowska, Zdzisława Romanowska-Duda, Wiktor Pszczółkowski
238
zawierające 60% BG-110 i 5% odcieku dla kultur PER1 nie wykazano istotnych różnic
w gęstości optycznej w stosunku do kultury kontrolnej, a dla kultur PER2 do 21 dnia
hodowli. Po 28 dniach hodowli kultury Anabaena sp. PER1 i PER2 po trzecim pasażu
miały o 16% i 7% niższą gęstość od kultur kontrolnych (p>0,001).
Wykres 1. Gęstość optyczna (λ 680nm) kultur Anabaena PER 1 trzykrotnie pasażowanych (p1,p2,p3) na
podłożu ze zmienną proporcją zawartości BG-110, odpadu i wody destylowanej. Pierwsza wartość w nazwie
wariantu oznacza zawartość podłoża BG-110 (100%, 0%, 60%, 30%, 10%) druga wartość oznacza zawartość odpadu (0%, 5%, 10%, 15%).
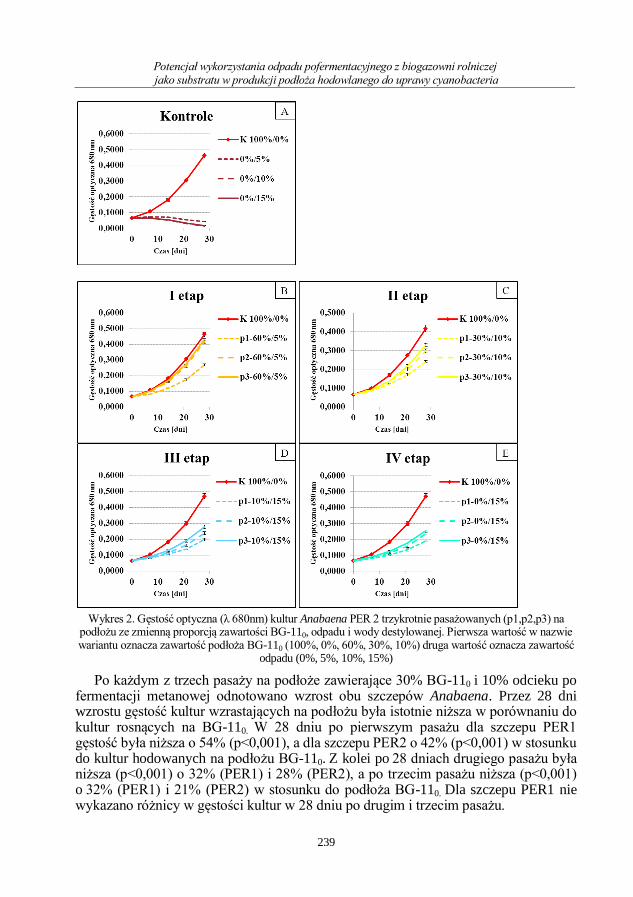
Potencjał wykorzystania odpadu pofermentacyjnego z biogazowni rolniczej jako substratu w produkcji podłoża hodowlanego do uprawy cyanobacteria
239
Wykres 2. Gęstość optyczna (λ 680nm) kultur Anabaena PER 2 trzykrotnie pasażowanych (p1,p2,p3) na
podłożu ze zmienną proporcją zawartości BG-110, odpadu i wody destylowanej. Pierwsza wartość w nazwie wariantu oznacza zawartość podłoża BG-110 (100%, 0%, 60%, 30%, 10%) druga wartość oznacza zawartość
odpadu (0%, 5%, 10%, 15%)
Po każdym z trzech pasaży na podłoże zawierające 30% BG-110 i 10% odcieku po fermentacji metanowej odnotowano wzrost obu szczepów Anabaena. Przez 28 dni wzrostu gęstość kultur wzrastających na podłożu była istotnie niższa w porównaniu do kultur rosnących na BG-110. W 28 dniu po pierwszym pasażu dla szczepu PER1 gęstość była niższa o 54% (p<0,001), a dla szczepu PER2 o 42% (p<0,001) w stosunku do kultur hodowanych na podłożu BG-110. Z kolei po 28 dniach drugiego pasażu była niższa (p<0,001) o 32% (PER1) i 28% (PER2), a po trzecim pasażu niższa (p<0,001) o 32% (PER1) i 21% (PER2) w stosunku do podłoża BG-110. Dla szczepu PER1 nie wykazano różnicy w gęstości kultur w 28 dniu po drugim i trzecim pasażu.

Agata Pszczółkowska, Zdzisława Romanowska-Duda, Wiktor Pszczółkowski
240
Po każdym z trzech pasaży na podłoże zawierające 10% BG-110 i 15% odcieku po fermentacji metanowej odnotowano wzrost obu szczepów Anabaena sp. i w każdym z przypadków był on wolniejszy przez cały okres prowadzenia hodowli w porównaniu do kultur hodowanych na podłożu BG-110. Po 28 dniach od pierwszego pasażu gęstość optyczna kultur szczepów PER1 i PER2 była niższa (p<0,001) odpowiednio o 59% i 58%, po 28 dniach po drugim pasażu niższa (p<0,001) o 54% i 49%, a po 28 dniach po trzecim pasażu niższa (p<0,001) o 51% i 41%.
W kolejnych pasażach na podłożu zawierającym 15% odcieku i wodę destylowaną oba szczepy wzrastały w podobnym tempie jak na podłożu zawierającym 10% BG-110
i 15% odcieku. W każdym przypadku gęstość optyczna była istotnie (p<0,001) niższa niż kultur na podłożu BG-110. Po 28 dniach hodowli w kolejnych pasażach gęstość optyczna kultur szczepu PER1 była niższa o 67% (p1), 57% (p2), 55% (p3), a szczepu PER2 niższa o 60% (p1), 50% (p2), 46% (p3) w stosunku do kultur wzrastających na podłożu kontrolnym.
W doświadczeniach zastosowano najprostsze statyczne warunki wzrostu kultur bez jakiegokolwiek mieszania (za wyjątkiem dni w których pobierano próby), zapewniając jednocześnie dużą stabilność warunków środowiska (temperatura, oświetlenie). W ten sposób wzrost cyanobacteria był jednolity i stosunkowo powolny zarówno w przy-padku wariantów zawierających odciek jak i kulturę hodowaną na podłożu BG-110. Zastosowane podłoże nie zawierające azotu również miało wpływ na wolniejszy wzrost i rozwój kultur. W doświadczeniu wykazano, że w tak podstawowych warun-kach możliwa jest hodowla szczepów Anabaena sp. na podłożu złożonym jedynie z 15% odcieku po fermentacji metanowej i wody destylowanej z wydajnością 46-55% w porównaniu do serii kontrolnej hodowanej na podłożu BG-110. Oba wykorzystane w doświadczeniach szczepy są nietoksyczne i mogą być stosowane jako bionawóz [13-15]. Tym samym przy minimalnych inwestycjach można przetworzyć odpad jakim jest odciek po fermentacji metanowej na wartościowy substrat do uprawy roślin. Niezbęd-ne są dalsze badania , które będą miały na celu poprawę wzrostu szczepów Anabaena sp, na podłożu zawierającym jedynie odciek i wodę. Jednym z podstawowych czyn-ników spowalniających wzrost na podłożu z odciekiem jest barwa i mętność odcieku ograniczająca dostęp promieniowania fotosyntetycznie czynnego. Planowane są dalsze badania w celu zapewnienia równomiernej ekspozycji komórek cyanobacteria na światło, co znacznie zwiększy dynamikę wymiany gazów: usuwanie powstałego w wyniku fotosyntezy tlenu i rozpuszczanie dwutlenku węgla i azotu z atmosfery oraz dynamikę wykorzystania substancji odżywczych [16]. W przypadku uprawy przemy-słowej kulturę można suplementować dwutlenkiem węgla.
5. Wnioski
Kolejne pasażowania kultur Anabena sp. PER1 i PER2 na podłoża o stopniowo zwiększanym stężeniu odpadu od 0% do 15% i obniżając stężenia BG-110 ze 100% do 0% doprowadziło do aklimacji komórek sinic do nowych warunków wzrostowych,
Kolejne pasaże pozwoliły na wzrost kultur na podłożach składających się wyłącznie z odpadu pofermentacyjnego i wody,

Potencjał wykorzystania odpadu pofermentacyjnego z biogazowni rolniczej jako substratu w produkcji podłoża hodowlanego do uprawy cyanobacteria
241
Możliwe jest wykorzystania odcieku po fermentacji metanowej jako podłoża w hodowli komórek sinic in vitro,
Zastosowanie odpadu z produkcji biogazu jako składnika podłoży hodowlanych, może być opłacalne ekonomicznie biorąc pod uwagę również korzyści płynące z produktów, uzyskiwanych z sinic np. bionawozów,
Niezbędne są dalsze prace nad globalnym zdefiniowaniem warunków wzrostu sinic z wykorzystaniem odpadów pofermetacyjnych w celu hodowli w warunkach skali ćwierć-technicznej.
Uwagi ogólne:
Badania sfinansowane przez Narodowe Centrum Badań i Rozwoju w ramach programu – Biostrateg II; Grant nr 2/296369/5/NCBR/2016.
Literatura
1. Kaspersen B.S., Christensen T.B., Fredenslund A.M., Møller H.B., Butts M.B., Jensen N.H., Kjaer T. Linking climate change mitigation and coastal utrophication management through biogas technology: Evidence from a new Danish bioenergy concept, Science of The Total Environment., 541:15 (2016), s. 1124-1131.
2. Schuurmans R.M., Alphen P., Schuurmans J.M., Matthijs H.C.P., Hellingwerf K.J. Comparison of the photosynthetic yield of cyanobacteria and green algae: different methods give different answers, PLoS One. 10:9 (2015) online.
3. Barsanti L., Gualtieri P. Algae : Anatomy, Biochemistry, And Biotechnology, Second Edition, Boca Raton, FL : CRC Press, (2014), ISBN: 9781439867327.
4. Brasil B.S.A.F., Siqueira F.G., Salum T.F.C., Zanette C.M., Spier M.R. Microalgae and cyanobacteria as enzyme biofactories, Algal Research., 25 (2017), s. 76-89.
5. Whitton B.A. Ecology of Cyanobacteria II : Their Diversity in Space and Time. Cz. 2, ed. Dordrecht: Springer, (2012), ISBN: 9789400755987.
6. Caldwell D.H. Sewage Oxidation Ponds: Performance, Operation and Design, Sewage Works Journal., 18 (1946), s. 433-458.
7. Oswald W.J., Asce A.M., Gotaas H.B., Asce M. Photosynthesis in sewage treatment, Transactions of the American Society of Civil Engineers., 122 (1957), s. 73-105.
8. Noue J.D.L., Proulx D. Biological tertiary treatment of urban waste Water with chitosan- immobilized Phormidium. Applied Microbiology and Biotechnology., 29 (1988), s. 292-297.
9. Manoharan C., Subramanian G. Sewage-cyanobacterial interaction-A case study, Indian Journal of Environmental Protection., 12 (1992), s. 254-258.
10. Vijayakumar S. Potential Applications of Cyanobacteria in Industrial Effluents-A Review, Journal of Bioremediation and Biodegradation., 6 (2012).
11. Kim J., Hwang M.J., Lee S.J., Noh W., Kwon J.M., Choi J.S., Kang C.M., Efficient recovery of nitrate and phosphate from wastewater by an amine-grafted adsorbent for cyanobacterial biomass production, Bioresource Technology, 205 (2016), s. 269-273.
12. Sood A., Prasanna R., Renuka N., Ahluwalia A.S. Cyanobacteria as Potential Options for Wastewater Treatment,Springer International Publishing (2015), s. 83-93.
13. Grzesik M., Romanowska-Duda Z., Ability of cyanobacteria and green algae to improve metabolic activity and development of willow plants, Polish Journal of Environmental Studies., 24:3 (2015), s. 1003-1012.

Agata Pszczółkowska, Zdzisława Romanowska-Duda, Wiktor Pszczółkowski
242
14. Singh J.S., Kumar A., Rai A.N., Singh D.P., Cyanobacteria: a precious bio-resource in agriculture, ecosystem, and environmental sustainability, Frontiers of Microbiology., 7:529 2016.
15. Grzesik M, Romanowska-Duda Z., Kalaji H.M. Effectiveness of cyanobacteria and green algae in enhancing the photosynthetic performance and growth of willow (Salix viminalis L.) plants under limited synthetic fertilizers application, Photosynthetica., 55:3 (2017), s. 510-521.
16. Santos A.M., Vieira K.R., Sartori R.B., Santos A.M., Queiroz M.I., Zepka L.Q., Lopes E.J. Heterotrophic cultivation of cyanobacteria: study of effect of exogenous sources of organic carbon, absolute amount of nutrients, and stirring speed on biomass and lipid productivity, Frontiers in Bioengineering and Biotechnology., 5:12 (2017), (online).
Potencjał wykorzystania odpadu pofermentacyjnego z biogazowni rolniczej jako substratu w produkcji podłoża hodowlanego do uprawy cyanobacteria
Streszczenie Produkcja biomasy mikroglonów jest intensywnie rozwijana ze względu na potencjał ekonomiczny substancji produkowanych przez te organizmy. Mniej uwagi poświęcane jest grupie mikroorganizmów fotoautotroficznych – cyanobacteria, wśród których występują gatunki posiadające zdolność do produkcji związków chemicznych, użytecznych m. in. w produkcji rolniczej. Celem badań było zaadaptowanie dwóch nietoksycznych szczepów cyanobacteria z rodzaju Anabaena sp. do podłoża hodowlanego przy-gotowanego z odpadu pofermentacyjnego, pochodzącego z biogazowni w Piaszczynie. Doświadczenia prowadzono z zachowaniem stałej temperatury 25±1°C oraz fotoperiodu: 16 h oświetlenia i 8 h ciemności. Stopniowo zwiększano stężenie odpadu pofermentacyjnego w podłożu hodowlanym jednocześnie redu-kując objętość standardowego medium BG-110. Do oceny kinetyki wzrostu cyanobacteria zastosowano metodę pomiaru fluorescencji za pomocą fluorymetru AquaPen-C AP-C 100. Rezultaty badań wykazały możliwość wykorzystania odpadu pofermentacyjnego w hodowli komórek cyanobacteria in vitro. Kolejne pasaże pozwoliły na wzrost kultur na podłożach składających się wyłącznie z odpadu pofermentacyjnego i wody. Wydajność wzrostu kultur Anabaena sp. na podłożu złożonym z odcieku pofermentacyjnego i wody wynosiła 46-55% wzrostu otrzymanego na standardowym medium BG-110. Słowa kluczowe: Anabaena, odciek pofermentacyjny
Digestate from agricultural biogas plant as substrate for production of culture medium for cyanobacteria
Abstract The production of microalgal biomass is intensively developed due to the economic potential of the substances produced by these organisms. Less attention is paid to the group of photoautotrophic microorganisms – cyanobacteria, among which there are species with the ability to produce chemical compounds useful in i.e. agriculture. The aim of the study was to adapt two non-toxic cyanobacteria strains of the genus Anabaena sp. to a culture medium prepared from digestate from a biogas plant in Piaszczyna. Experiments were conducted at a constant temperature of 25 ± 1°C and photoperiod: 16 h of lighting and 8 h of darkness. Gradually the concentration of digestate in the culture medium was increased and the volume of standard BG11 medium was reduced. The fluorescence measurement using the AquaPen-C AP-C 100 fluorimeter was used to evaluate the cyanobacterial growth kinetics. Results of the study showed the possibility of using digestate in cyanobacterial in vitro cell culture. Subsequent passages allowed for the cultivation of cultures on substrates consisting solely of digestate waste and water. The efficiency of Anabaena sp. culture cultivation on the substrate composed of digestate and water was 46-55% of the growth on standard BG-110 medium. Key words: Anabaena, digestate,

243
Indeks Autorów
Bossowska-Nowicka M. .............. 7, 64
Brągoszewska E. ............................. 182
Dobrzyńska D. .................................. 45
Dołhańczuk-Śródka A. ..................... 71
Dubisz A. ........................................... 45
Duda S. M. ............................. 190, 200
Gajek A. ........................................... 148
Garbacz M. ............................. 190, 200
Gospodarek-Komkowska E. . 112, 148
Grabiński J. ...................................... 210
Hałgas M. .......................................... 36
Iwańczyk B. ..................................... 126
Jarzynka S. ....................................... 126
Jurkowski A....................................... 28
Kochan E. .................................... 80, 92
Kozieł M. ........................................... 14
Kozioł J. J. ......................................... 28
Kozioł M. M. ..................................... 36
Kręcidło M. ..................................... 171
Krzyśko-Łupicka T. ........................ 171
Kuś R. ................................................ 36
Kwiatkowski P. ................................. 92
Kwiecińska-Piróg J. ............... 112, 148
Ludwiczak A. .................................... 54
Lue G. ................................................ 36
Majewska E. .................................... 160
Małajowicz J. .................................. 160
Marchwińska K. ................................ 45
Mielcarska M. B.................................. 7
Mikos-Szymańska M...................... 210
Minkiewicz A. ................................. 126
Nabrdalik M. ..................................... 71
Nowak D.......................................... 160
Obuch-Woszczatyński P. ............... 105
Olender A. ......................................... 36
Olędzka O. ....................................... 126
Olszewski J. ....................................... 92
Pacia A. .............................................. 71
Pastuszka J. S................................... 182
Piotrowski M. .................................. 105
Pituch H. .......................................... 105
Przekwas J. ...................................... 112
Pszczółkowska A. .................. 220, 233
Pszczółkowski W. .................. 220, 233
Rajtor M. .......................................... 182
Romanowska-Duda Z............ 220, 233
Sienkiewicz M. ............................80, 92
Strom K............................................ 126
Szczepańska I. ................................... 28
Szulc-Dąbrowska L. ......................... 64
Szułczyńska J. ................................. 138
Toka F. N. .......................................... 64
Vizcarra F. G. .................................... 36
Włodarczyk K. .................................. 14
Wojton D. .......................................... 14
Wychowański P. ............................. 126
Wyzińska . ....................................... 210
Zielińska-Bliźniewska H. ...........80, 92


![Biochemia i mikrobiologia środowiska 3wch.pwr.edu.pl/fcp/YGBUKOQtTKlQhbx08SlkTUAJQX2o8... · [2] Mikrobiologia materiałów- pod red. B. Zyski i Z. Żakowskiej- Łódź 2005 [3]](https://static.fdocuments.pl/doc/165x107/5e32c16078321c5fa50dd727/biochemia-i-mikrobiologia-rodowiska-3wchpwreduplfcpygbukoqttklqhbx08slktuajqx2o8.jpg)















