
1. i Nazwisko: z podaniem nazwy, miejsca - ibb.waw.pl w j.polskim_2.pdf · angielskim: Comment...
38
1 Autoreferat 1. Imię i Nazwisko: Robert Szoszkiewicz 2. Posiadane dyplomy, stopnie naukowe/ artystyczne – z podaniem nazwy, miejsca i roku ich uzyskania oraz tytułu rozprawy doktorskiej. dyplom magistra fizyki, Uniwersytet Jagielloński (UJ), Kraków, 1999. “Spectroscopia Dielektryczna w badaniu dynamiki szkła organicznego ortoterfenylu (OTP)“ Promotor: Prof. Józef K. Mościcki, Wydział Matematyki i Fizyki, Uniwersytet Jagielloński, Kraków. Praca to była rezultatem pięcioletnich studiów magisterskich na kierunku “Studia Matematyczno-Przyrodnicze”. Po pierwszych dwóch latach studiów, studenci byli zobowiązani do wyboru tradycyjnego kierunku studiów magisterskich. W rezultacie uzyskałem dyplom magisterski fizyki na Wydziale Matematyki i Fizyki UJ, a także zaliczyłem osiem semestrów z dziesięcio-semestralnego programu studiów magisterskich z chemii biologicznej na Wydziale Chemii UJ. dyplom doktora fizyki, Szwajarska Politechnika Rządowa w Lozannie (Ecole Polytechnique Federale de Lausanne, EPFL), Lozanna, Szwajcaria, paździenik 2003. Tytuł rozprawy doktorskiej: “Histereza adhezji i tarcie w skalach lokalnych” (Tytuł oryginalny w języku angielskim: “Adhesion hysteresis and friction at local scales”). Celem mojej pracy doktorskiej było zrozumienie związków pomiędzy histerezą adhezji a tarciem w mikro- i nano- skalach, określanych dalej jako “skale lokalne”. Studiowane materiały były materiałami inżynierskimi, polimerami i białkami. Na podstawie wyników eksperymentalnych i symulacji komputerowych stworzyłem model fizyczny opisujący zbadane zjawiska. W trakcie tej pracy rozwinąłem także zaawansowane techniki badawcze związane z ultradźwiękową mikroskopią sił (ang. ultrasonic force microscopy) i nanoindentacją. Promotor: Prof. Gerald Gremaud, Wydział Fizyki, EPFL. Recenzenci: Prof. Walter Arnold, Fraunhofer Institute for Non-Destructive Testing, Saarbrücken, Republika Federalna Niemiec; Prof. Bernard Cretin, Université de Franche- Comté, Besançon, Francja; Prof. Giovanni Dietler, Université de Lausanne (UNIL), Szwajcaria; Dr. Andrzej Kulik, EPFL, Szwajcaria.
Transcript of 1. i Nazwisko: z podaniem nazwy, miejsca - ibb.waw.pl w j.polskim_2.pdf · angielskim: Comment...

1
Autoreferat
1. Imię i Nazwisko: Robert Szoszkiewicz
2. Posiadane dyplomy, stopnie naukowe/ artystyczne – z podaniem nazwy, miejsca
i roku ich uzyskania oraz tytułu rozprawy doktorskiej.
dyplom magistra fizyki, Uniwersytet Jagielloński (UJ), Kraków, 1999.
“Spectroscopia Dielektryczna w badaniu dynamiki szkła organicznego ortoterfenylu (OTP)“
Promotor: Prof. Józef K. Mościcki, Wydział Matematyki i Fizyki, Uniwersytet Jagielloński,
Kraków.
Praca to była rezultatem pięcioletnich studiów magisterskich na kierunku “Studia
Matematyczno-Przyrodnicze”. Po pierwszych dwóch latach studiów, studenci byli
zobowiązani do wyboru tradycyjnego kierunku studiów magisterskich. W rezultacie
uzyskałem dyplom magisterski fizyki na Wydziale Matematyki i Fizyki UJ, a także
zaliczyłem osiem semestrów z dziesięcio-semestralnego programu studiów magisterskich z
chemii biologicznej na Wydziale Chemii UJ.
dyplom doktora fizyki, Szwajarska Politechnika Rządowa w Lozannie (Ecole
Polytechnique Federale de Lausanne, EPFL), Lozanna, Szwajcaria, paździenik 2003.
Tytuł rozprawy doktorskiej: “Histereza adhezji i tarcie w skalach lokalnych” (Tytuł
oryginalny w języku angielskim: “Adhesion hysteresis and friction at local scales”).
Celem mojej pracy doktorskiej było zrozumienie związków pomiędzy histerezą adhezji a
tarciem w mikro- i nano- skalach, określanych dalej jako “skale lokalne”. Studiowane
materiały były materiałami inżynierskimi, polimerami i białkami. Na podstawie wyników
eksperymentalnych i symulacji komputerowych stworzyłem model fizyczny opisujący
zbadane zjawiska. W trakcie tej pracy rozwinąłem także zaawansowane techniki badawcze
związane z ultradźwiękową mikroskopią sił (ang. ultrasonic force microscopy) i
nanoindentacją.
Promotor: Prof. Gerald Gremaud, Wydział Fizyki, EPFL.
Recenzenci: Prof. Walter Arnold, Fraunhofer Institute for Non-Destructive Testing,
Saarbrücken, Republika Federalna Niemiec; Prof. Bernard Cretin, Université de Franche-
Comté, Besançon, Francja; Prof. Giovanni Dietler, Université de Lausanne (UNIL),
Szwajcaria; Dr. Andrzej Kulik, EPFL, Szwajcaria.

2
3. Informacje o dotychczasowym zatrudnieniu w jednostkach naukowych/ artystycznych.
Politechnika Warszawska, Wydział Inżynierii Materiałowej, Warszawa, Polska
2015.04 - obecnie: Adiunkt naukowy
Kansas State University (KSU), Department of Physics, Manhattan, KS, USA
2014.03 - 2015.02: Tenured Associate Professor (Profesor nadzwyczajny)
2008.07 - 2014.03: Tenure-track Assistant Professor
Columbia University in the City of New York, Department of Biological
Sciences, New York, NY, USA
2007.01 - 2008.07: Staż podoktorski w pracowniach Prof. Julio Fernandeza
Georgia Institute of Technology, Department of Physics, Atlanta, GA, USA
2004.01 - 2006.12: Staż podoktorski w pracowniach Prof. Elisy Riedo
Swiss Federal Institute of Technology in Lausanne, Department of Physics,
Switzerland
1999.10 - 2003.11: Doktorant
4. Wskazanie osiągnięcia wynikającego z art. 16 ust. 2 ustawy z dnia 14 marca 2003 r. o stopniach naukowych i tytule naukowym oraz o stopniach i tytule w zakresie sztuki (Dz. U. nr 65, poz. 595 ze zm.):
a) tytuł osiągnięcia naukowego/artystycznego:
Badania mechanicznie indukowanych zmian konformacyjnych w wybranych białkach
na poziomie pojedynczych cząsteczek.
b) (autor/autorzy, tytuł/tytuły publikacji, rok wydania, nazwa wydawnictwa, recenzenci wydawniczy),
Poniżej przedstawiam, w porządku chronologicznym, publikacje wchodzące w skład cyklu
habilitacyjnego:
N. Ploscariu, K. Kuczera, K. E. Malek, M. Wawrzyniuk, A. Dey, and R. Szoszkiewicz,
Studia mechanicznie-indukowanej ekspozycji miejsca S2 w domenie NRR białka Notch na
poziomie pojedynczej cząsteczki (Tytuł oryginalny w języku angielskim: Single Molecule
Studies of Force-Induced S2 Site Exposure in the Mammalian Notch Negative Regulatory
Domain), Journal of Physical Chemistry B, 118(18), 4761-4770 (2014).
Impact Factor: 3.377 (2013), 3.527 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 30
Cytowania: Web of Science: 1 Google Scholar: 1
Wkład pracy dr Roberta Szoszkiewicza: Wymyśliłem ten kierunek badawczy, pomagałem przy
badaniach AFM, a także przy zbieraniu pozostałych danych eksperymentalnych, wytyczyłem szlaki
analizy wyników i uczestniczyłem w tej analizie, połączyłem wyniki eksperymentalne z symulacjami
komputerowymi, i wreszcie napisałem ten manuskrypt.

3
J. Cruz, P. H. Pfromm, R. Szoszkiewicz, and M. E. Rezac, Hydrolazy na powierzchniach
krzemionkowych: związki między pokryciem powierzchni, aktywnością i własnościami
cząsteczkowymi (Tytuł oryginalny w języku angielskim: Hydrolases on Silica Surfaces:
Coverage-Activity-Molecular Property Relationships Revealed, Process Biochemistry
49(5), 830-839 (2014).
Impact Factor: 2.524 (2013), 2.922 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 30
Cytowania: Web of Science: 0 Google Scholar: 1
Wkład pracy dr Roberta Szoszkiewicza: Moją rolą było zorganizowanie i rozpoczęcie badań przy
pomocy AFM-u, a także pomoc w analizie danych z AFM-u.
K. E. Malek and R. Szoszkiewicz, Zmiany sztywności cząsteczki białka w wykrywaniu
stanów przejściowych w trakcie procesu fałdowania białek (Tytuł oryginalny w języku
angielskim: Changes of protein stiffness during folding detect protein folding
intermediates), Journal of Biological Physics 40(1), 15-23 (2014).
Impact Factor: 1.152 (2013), 1.346 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 15
Cytowania: Web of Science: 2 Google Scholar: 3
Wkład pracy dr Roberta Szoszkiewicza: Wymyśliłem ten kierunek badawczy, pomagałem przy
badaniach AFM i analizie uzyskanych danych, wytyczyłem szlaki analizy wyników i uczestniczyłem
w tej analizie, i wreszcie napisałem ten manuskrypt.
N. Ploscariu, R. Szoszkiewicz, Metoda pomiarów własności nanomechanicznych
obiektów biologicznych (Tytuł oryginalny w języku angielskim: A method to measure
nanomechanical properties of biological objects), Applied Physics Letters 103, 263702
(2013).
Impact Factor: 3.515 (2013), 3.739 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 40
Cytowania: Web of Science: 3 Google Scholar: 5
Wkład pracy dr Roberta Szoszkiewicza: Wymyśliłem ten kierunek badawczy, pomagałem przy
badaniach AFM i analizie uzyskanych danych, wytyczyłem szlaki analizy wyników, uczestniczyłem
w analizie wyników, a w szczególności w modelowaniu analitycznym jak i obliczeniach przy pomocy
komputera, i wreszcie napisałem ten manuskrypt.
R. Szoszkiewicz, Badania schematów redukcji pojedynczych wiązań dwusiarczkowych
przez tioredoksynę ludzką (Tytuł oryginalny w języku angielskim: Single-molecule studies
of disulfide bond reduction pathways used by human thioredoxin), Biophysical
Chemistry, 173, 31-38 (2013).
Impact Factor: 2.319 (2013), 2.145 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 20
Cytowania: Web of Science: 0 Google Scholar: 0
Wkład pracy dr Roberta Szoszkiewicza: Jako jedyny autor, wytyczyłem ten kierunek badawczy,
zebrałem i opracowałem wszystkie dane eksperymentalne, opracowałem wyniki pomiarowe, i
napisałem tą publikację.

4
Dey, R. Szoszkiewicz, Dokładna analiza szumów eksperymentalnych w spektrometrze
AFM i jego aplikacja do badania nanomechaniki białka ludzkiego Notch1 (Tytuł
oryginalny w języku angielskim: Complete noise analysis of a force spectroscopy AFM
setup and its applications to study nanomechanics of human Notch 1 protein),
Nanotechnology 23, 175101, 12 pages (2012).
Impact Factor: 3.842 (2012), 3.867 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 9 Google Scholar: 17
Wkład pracy dr Roberta Szoszkiewicza: Wytyczyłem ten kierunek badawczy, uczestniczyłem w
konstrukcji opisanego aparatu badawczego, uczestniczyłem w analizie wszystkich danych, i
napisałem tą publikację.
R. Szoszkiewicz, Komentarz do publikacji “Print your AFM” (Tytuł oryginalny w języku
angielskim: Comment about “print your AFM” paper), Review Scientific Instruments,
83, 037101, 3 pages, (2012). This is a peer-reviewed paper.
Impact Factor: 1.602 (2012), 1.556 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 30
Cytowania: Web of Science: 3 Google Scholar: 4
Wkład pracy dr Roberta Szoszkiewicza: Jako jedyny autor, wymyśliłem ten kierunek badawczy,
zebrałem i opracowałem wszystkie dane eksperymentalne, opracowałem wyniki pomiarowe, i
napisałem tą publikację.
R. Szoszkiewicz, Spektroskopia AFM w modzie krzywych siłowych (FX) i w modzie
ustalonej siły (FC) w badaniach reakcji nanomechanicznych i własności mechanicznych
pojedynczych cząstek biologicznych (Tytuł oryginalny w języku angielskim: Force-
extension (FX) and force-clamp (FC) AFM spectroscopies in investigating
mechanochemical reactions and mechanical properties of single biomolecules), rodział
opublikowany na zaproszenie w książce „Skaningowa mikroskopia w Naukach „Nano” i
w Nanotechnologii (Tytuł oryginalny w języku angielskim: Scanning Probe Microscopy
in Nanoscience and Nanotechnology), p. 395-423, edited by B. Bhushan, Springer-
Verlag, Heidelberg (2010).
Impact Factor i punkty ministerialne nie są dostępne.
Cytowania: Web of Science: 4 Google Scholar: 2
Wkład pracy dr Roberta Szoszkiewicza: Jako jedyny autor, wymyśliłem temat tego rozdziału i
napisałem tą publikację.
R. Szoszkiewicz, S. R. K. Ainavarapu, A. P. Wiita, R. Perez-Jimenez, J. Fernandez,
Analiza typu dwell-time dla reakcji mechanochemicznych na poziomie pojedynczej
cząsteczki (Tytuł oryginalny w języku angielskim: Dwell time analysis of a single
molecule mechanochemical reaction), Langmuir, 24, 1356 (2008).
Impact Factor: 4.097 (2008), 4.489 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 25 Google Scholar: 28

5
Wkład pracy dr Roberta Szoszkiewicza: Uczestniczyłem w wytyczaniu tego kierunku badawczego,
zebrałem większość danych eksperymentalnych i przewodziłem ich analizie, i wreszcie napisałem tą
publikację.
c) omówienie celu naukowego/artystycznego ww. pracy/prac i osiągniętych wyników
wraz z omówieniem ich ewentualnego wykorzystania.
Białka uczestniczą w większość istotnych procesów komórkowych związanych z rozwojem i
homeostazą. Używając obecnie dostępnych narzędzi do badań procesów biofizycznych na
poziomie pojedynczych cząsteczek jesteśmy w stanie nie tylko podglądać, ale i zrozumieć
szczegóły zmian konformacyjnych uczestniczących cząsteczek białek i to zarówno wewnątrz
pojedynczej komórki jak i na poziomie oddziaływujących ze sobą komórek. Przykładami
takich procesów wewnątrzkomórkowych są: replikacja cząsteczek DNA, transkrypcja DNA,
procesy rekombinacji cząsteczek DNA, translacja białek, procesy fałdowania się białek,
procesy związane z translokacją i degradacją białek, i wreszcie badania aktywności
enzymatycznej pojedynczych cząsteczek enzymów. Podobnie, przykładami procesów
związanych z oddziaływaniami międzykomórkowymi są badania procesów adhezji
międzykomórkowej, a także blisko-zasięgowej komunikacji międzykomórkowej. Z wyników
badań tych procesów wynika, że nie tylko oddziaływania biochemiczne, ale również siła
mechaniczna są istotnymi elementami biochemii i biofizyki życia na poziomie
molekularnym, gdyż większość procesów rozwojowych jak i podtrzymujących życie jest
realizowana dzięki maszynom molekularnym. Każda z takich maszyn molekularnych zawiera
co najmniej parę cząsteczek białek, która pracują wspólnie i oddziaływają za sobą zarówno
biochemicznie jak i mechanicznie. Oddziaływania mechaniczne zachodzą częstokroć dzięki
transdukcji siły poprzez specyficzne oddziaływania białko-białko oraz wymuszaniu zmian
konformacyjnych białek wskutek stresu mechanicznego. Stąd też, aby pomóc w zrozumieniu
podstawowych procesów wpływających na rozwój jak i podtrzymywania życia, w
zaprezentowanym poniżej cyklu habilitacyjnym skupiłem się na badaniu mechanicznie
indukowanych zmian konformacyjnych w wybranych białkach na poziomie pojedynczych
cząsteczek. Aby zajrzeć w głąb obserwowanych procesów rozwinąłem nowe narzędzia
biofizyczne w formie zarówno nowego metod analizy dostępnych danych eksperymentalnych
jak i nowego sprzętu badawczego. Badania z użyciem tych narzędzi pozwoliły zrozumieć
szereg mechanochemicznie indukowanych zmiany konformacyjne wybranych białek. W
szczególności, badaliśmy zmiany konformacyjne pojedynczych wiązań dwusiarczkowych w
białkach i związane z nimi procesy katalizy enzymatycznej, a także procesy fałdowania i
rozfałdowywania się pewnych modelowych białek.
Zmiany konformacyjne pojedynczych wiązań i związane z nimi
procesy katalizy enzymatycznej Na wstępie skupiłem się na badaniach wpływu siły mechanicznej na redukcję pojedynczego
wiązania dwusiarczkowego. Wiązanie to zostało wprowadzone do struktury prostej
modelowej proteiny używając narzędzi inżynierii białkowej.

6
Wiązania dwusiarczkowe to najsilniejsze oddziaływania stabilizujące struktury
trzeciorzędowej białek. Mają one różnorakie znaczenia, ale miedzy innymi kontrolują
kinetykę fałdowania się białek poprzez kontrolę populacji stanów pośrednich i
termodynamiczną oraz mechaniczną stabilizację cząsteczek białek w ich stanach natywnych.
Opracowanie danych dostępnych w bazie Protein Data Bank i opublikowane przez Schmidta
i Hogga w 2007 w czasopiśmie BMC Structural Biology wskazało na 42960 unikalnych
wiązań dwusiarczkowych obecnych w 31611 cząsteczek białek, których struktura została
określona dzięki krystalografii rentgenowskiej. Wskazuje to wszechobecność wiązań
dwusiarczkowych w cząsteczkach białek. W szczególności, wiele protein zawiera wiązania
dwusiarczkowe, które są wystawione na działanie siły mechanicznej in vivo. Dobrymi
przykładami są tutaj proteiny uczestniczące w procesach adhezji komórkowej takie jak
cadhedryny, selektyny, czy IgCAMs. Inne przykłady to białka biorące udział w utrzymaniu
struktury macierzy międzykomórkowej takie jak fibronektyna, a także białka wpływające na
elastyczność tkanek, a wiec takie jak fibrilina czy tytyna.
Regulacja stanu redoks wiązań dwusiarczkowych wskutek stresu mechanicznego wskazuje
na duże znaczenie siły mechanicznej w procesach redukcji pojedynczego wiązania
dwusiarczkowego w białkach. Redukcja wiązań dwusiarczkowych w białkach, a wiec
rozerwanie wiązania kowalencyjnego pomiędzy grupami tiolowymi cystein w obecności
czynnika redukującego, zachodzi według następującego schematu:
-Cys - S1 – S2 – Cys- + R-S-H <-> -Cys-S1-S-R + H- S2 – Cys- Schemat 1
Najbardziej prawdopodobnym mechanizmem tej reakcji jest dwucząsteczkowa substytucja
nukleofilowa, SN2. Według tego mechanizmu, cząsteczka R – SH używa wolnej pary
elektronowej zlokalizowanej na atomie siarki do przeprowadzenia ataku nukleofilowego na
jeden z atomów siarki w wiązaniu dwusiarczkowym (tutaj wybrany został atom S1). Udany
atak skutkuje rozerwaniem wiązania dwusiarczkowego miedzy cysteinami. Siarka nie jest
jedynym atomem, który może pełnić funkcję czynnika nukleofilowego. Związki fosforu, a w
szczególności tris(2-carboxyethyl)fosfina (TCEP), okazały się działać znacznie wydajniej w
białkach.
R. Szoszkiewicz, S. R. K. Ainavarapu, A. P. Wiita, R. Perez-Jimenez, J. Fernandez,
Dwell time analysis of a single molecule mechanochemical reaction, Langmuir, 24, 1356
(2008).
Wcześniejsze dane z grupy Prof. Fernandeza z Uniwersytetu Columbia pokazały, że redukcja
pojedynczych wiązań dwusiarczkowych w cząsteczka białek może zostać zmierzona
ilościowo przy pomocy mikroskopu sił atomowych (AFM) działającego w modzie stałej siły
ciągnięcia (force-clamp AFM). Metoda ta została zobrazowana na Rysunku 1.

7
Rysunek 1. Aplikacja mikroskopii sił atomowych w modzie stałej siły ciągnięcia (force-clamp
atomic force microscopy, FC AFM) do detekcji procesu redukcji pojedynczych wiązań
dwusiarczkowych. A) Cząsteczka rekombinowanego białka składającego się z identycznych modułów
połączonych szeregowo w ten sam sposób jest umiejscowiona pomiędzy dźwignią/belką AFMu (żółty
trójkąt na górze rysunku) a próbką przytwierdzoną do skanera AFM (nie pokazano, na dole rysunku).
Każdy moduł białka, tutaj: 27 moduł tytyny z ludzkiego mięśnia sercowego, oznaczony dalej I27,
zawiera dokładnie jedno wiązanie dwusiarczkowe wprowadzone metodami inżynierii białkowej i
zaznaczone żółtymi sferami. Każde z takich wiązań dwusiarczkowych staje się dostępne dla procesu
redukcji, np. za pomocą cząstek TCEP, po uprzednim częściowym rozfałdowaniu się fragmentu białka
zaznaczonego kolorem czerwonym. W wyniku procesu redukcji następuje dalsze, całkowite
rozwiniecie się każdego z modułów białka. B) „Length” oznacza długość cząsteczki białka pomiędzy
belka AFM a próbka, i jest to parametr mierzony przez FC AFM w funkcji czasu. Każdy zdarzenie
częściowego rozwinięcia się I27 w celu ekspozycji wiązania dwusiarczkowego odpowiada skokowi
rozwiniętej długości białka o 10.8 nm ± 1.1 nm przy użytej sile rozciągającej 180 pN. Analogicznie,
każdy wzrost długości białka o 14.1 nm ± 1.4 nm odpowiada pojedynczemu procesowi redukcji
wiązania dwusiarczkowego. Czas każdego zdarzenia redukcji –S-S- nazywamy później „dwell time”.
Czasy te liczone są od początku fazy ekspozycji wszystkich wiązań dwusiarczkowych na redukcję i
oznaczone dalej od t1 do t6.
Kinetyka procesu redukcji wiązań dwusiarczkowych pokazana na Rysunku 1 została
zmierzona poprzez dopasowania modelu mono-eksponencjalnego do krzywych zmian
długości białka w funkcji czasu zobrazowanych na Rys. 1.B i uśrednionych dla wielu cząstek
przy danej sile ciągnięcia. Takie przykładowe dopasowania zostały zobrazowane na Rysunku
2. Jednakże, gdy mamy do czynienia z reakcjami chemicznymi zachodzącymi według
różnych schematów to dopasowania modeli eksponencjalnych okazują się częstokroć
niewystarczające i nieodpowiednie, patrz Rysunek 2.A.
Częściowe
Rozwijanie Redukcja
A) B)

8
Rysunek 2. Wyniki kinetyki postępu redukcji wiązań dwusiarczkowych otrzymane metoda FC
AFM. Strona Lewa: Uśrednienie wielu krzywych zmian długości białka w funkcji czasu podczas
procesów redukcji dla wybranych czterech wartości sil ciągnięcia, patrz Rys. 1.B. Dopasowania
modelu mono-eksponencjalnego do każdej z tych krzywych pozwalają na otrzymanie stałych szybkości
reakcji, r, dla każdej z użytych sił ciągnięcia, F. Strona Prawa: Stale szybkości reakcji zależą w
sposób eksponencjalny od siły F, co zostało zobrazowane przez dopasowanie na niebiesko. Dane
pomiarowe zostały uzyskane dla białka rekombinowanego I278 i używając 12.5 mM roztworu
ditiotretoilu (DTT).
W publikacji napisanej przez Szoszkiewicza et al. opublikowanej w czasopiśmie Langmuir w
roku 2008, rozwinęliśmy metodę analizy „dwell time” do detekcji procesów redukcji wiązań
dwusiarczkowych. W kontraście do dopasowań modeli eksponencjalnych do kinetyki
redukcji wiązań –S-S- zobrazowanej na Rys. 2, metoda „dwell time” pozwala na
bezpośrednie rozróżnienie ilości równocześnie przebiegających reakcji chemicznych.
Technika ta polega na zebraniu mierzonych czasów „dwell time”, patrz Rys. 1.B, a następnie
skonstruowaniu histogramu obrazującego pierwiastek kwadratowy ze zliczeń w funkcji
logarytmu naturalnego czasu. Rysunek 3 obrazuje takie przykładowe histogramy.
Dopasowywując następnie odpowiednie funkcje rozkładu gęstości prawdopodobieństwa
otrzymuje się stałe szybkości zachodzących reakcji chemicznych. Co najważniejsze, taka
metoda wizualizacji wyników pozwala na jednoznaczne i szybkie rozróżnienie czy mamy do
czynienia z jednym czy wieloma schematami reakcji chemicznych.
Uzywajac metody „dwell time” zbadaliśmy kinetykę redukcji wiązan dwusiarczkowych dla
dwóch reduktorow: prostej cząsteczki nieorganiczej, TCEP, a także enzymu E. coli
tioredoksyny (TRX). W ten sposób pokazaliśmy również porównanie miedzy kinetyka
redukcji wiazan –S-S- za pomocą prostej cząsteczki nieorganiczej i skomplikowanego
enzymu. Tioredoksyna to wszechobecna reduktaza wiązań dwusiarczkowych w organizmach
żywych odpowiedzialna miedzy innymi za utrzymanie homeostazy redox w komórkach. Co
więcej, tioredoksyna wykazuje również aktywność anty-apoptyczna i bierze udział w
wybranych procesach komunikacji międzykomórkowej. Wcześniejsze publikacje z użyciem
modeli matematycznych do opisu kinetyki redukcji wiązań dwusiarczkowych wskazywały na
istnienie jednego schematu redukcji za pomocą prostych cząstek nieorganicznych jak TCEP i
dwóch schematów redukcji tych wiązań za pomocą TRX. Nasze rezultaty pokazały w sposób
bezpośredni, że w granicach naszej rozdzielczości eksperymentalnej jesteśmy w stanie
wykryć rzeczywiście jeden mechanizm redukcji –S-S- za pomocą TCEPu i dwa mechanizmy

9
gdy reduktorem jest TRX. Stąd też otrzymane przez nas dane stanowiły potwierdzenie
pewnych zmian konformacyjnych zachodzących w TRX w trakcie tej reakcji, które to
zmiany zostały opisane nieco wcześniej przez grupe Prof. Fernandeza w magazynie Nature z
roku 2007. Co jest również istotne, to, że nasze rezultaty pokazały, ze metoda histogramów
„dwell time” może z powodzeniem być zastosowana do opisu reakcji chemicznych badanych
metodami FC AFM. Jak pokazano nieco później przez X. Xie et al. w czasopiśmie Physics
Review B w roku 2008, metoda ta daje nie takie same, a komplementarne informacje
względem metody dopasowań krzywych eksponencjanych do opisu kinetyki reakcji
chemicznych.
Rysunek 3. Metoda analizy “dwell time” zastosowana do badania kinetyki reakcji redukcji wiązań
dwusiarczkowych za pomocą FC AFM. Logarytmiczne histogramy czasów „dwell time” są
otrzymane dla procesów redukcji wiązań dwusiarczkowych w I278 za pomocą cząsteczek TCEP i TRX.
Każdy z czasów „dwell time” został otrzymany jak wyjaśniono na Rys. 1.B. Dane wskazują na
istnienie jednego schematu reakcji dla TCEP-u i dwóch dla TRX.
R. Szoszkiewicz, Single-molecule studies of disulfide bond reduction pathways used by
human thioredoxin, Biophysical Chemistry, 173-174, 31-38 (2013).
Zakładając istnienie tylko jednej bariery energetycznej, stała szybkości reakcji redukcji
wiązań dwusiarczkowych, k, która jest mierzalna przy pomocy metody FC AFM może zostać
opisana ilościowo przy użyciu modelu Bella, patrz Rys. 4, w następujący sposób:
k = [czynnika redukującego] A exp ((FΔx-Ea) / kBT) Równanie 1
gdzie: [czynnika redukującego] to stężenie czynnika redukującego, A jest to tzw. form-faktor
i ma znacznie częstotliwości natarcia na barierę energetyczną, F to siła rozciągająca, Ea to
energia aktywacji, kB to stała Boltzmanna, Δx to średnie wydłużenie wiązania do stanu
przejściowego, a T to temperatura w skali bezwzględnej Kelvina.
Warto zauważyć, ze znajomość parametrów Ea oraz Δx pozwala na opisanie stanu
przejściowego reakcji, co stanowi unikalną informacje fizyko-chemiczną, która to jest
niesłychanie trudno osiągalna innymi zaawansowanymi metodami badawczymi, jak np.
spektroskopiami typu femto- i atto-sekundowymi czy spektroskopiami typu NMR.

10
Rysunek 4. Aplikacja modelu Bella do opisu redukcji
pojedynczego wiązania dwusiarczkowego przez DTT
(na podstawie danych otrzymanych przez Wiita et al.,
PNAS, 2007). Model Bella opisuje stałą szybkości
reakcji zerwania się wiązania ligand-receptor na skutek
siły mechanicznej. Rolą siły jest rozciąganie wiązania, a
wiec obniżanie energii aktywacji procesu jego zerwania.
Według tego modelu siła obniża energie aktywacji
proporcjonalnie do wydłużania się wiązania wzdłuż
kierunku ciągnięcia, xr, który pełni również rolę
parametru postępu reakcji. Takie zachowanie skutkuje
eksponencjalną zależnością stałej szybkości reakcji
względem siły mechanicznej, patrz Równanie 1. Stosując
model Bella do danych FC AFM uzyskanych dla DTT
jako czynnika redukującego, patrz Rys. 2, Wiita et al.
uzyskali energię aktywacji Ea = 65 kJ/mol i wartość
rozciągnięcia się wiązania dwusiarczkowego do stanu
przejściowego, Δx = 0.34 Å. Dane te okazały się bardzo
zbliżonego do danych przewidzianych metodami
symulacji komputerowych.
Dzięki zrozumieniu molekularnych aspektów redukcji wiązań dwusiarczkowych przy
pomocy prostych cząsteczek nieorganicznych zdecydowałem się na aplikację histogramów
typu „dwell time” do bardziej złożonych studiów, a mianowicie redukcji tychże wiązań przy
udziale enzymów. Jak poprzednio wybrałem tioredoksynę, ale tym razem skupiłem się na
tioredoksynie ludzkiej (hTrx). Z wcześniejszych studiów kinetyki redukcji pojedynczych
wiązań dwusiarczkowych przez Escherichia coli tioredoksyne otrzymano, że stała szybkości
redukcji tych wiązań początkowo malała a następnie rosła wraz ze wzrostem siły ciągnącej
(ang. clamping force) w badaniach FC AFM. Niemniej jednak, w przypadku hTrx, która jest
strukturalnie bardzo podobna do E. coli TRX, stale szybkości redukcji malały monotonicznie
wraz ze wzrostem siły, aż do stałego poziomu przy silach 300 pN i większych. Stąd też,
pojawiła się hipoteza badawcza, że w przeciwieństwie do E. coli TRX, hTrx używa
najprawdopodobniej jednego schematu redukcji wiązań dwusiarczkowych. Niemniej jednak,
powstają pytania skąd tak duża zmiana w przypadku hTrx przy bardzo dużej analogii
strukturalnej między E. coli TRX a hTrx, i czy może w takim przypadku nie powinno się
zacząć rozważać procesów redukcji wiązania dwusiarczkowego z wykorzystaniem zjawisk
tunelowania elektronów z rodników siarkowych z katalitycznych cystein hTrx? Tym
bardziej, ze procesy tunelowania nie powinny mocno zależeć od siły rozciągającej. Aby
zweryfikować te hipotezy i uzyskać bezpośrednią informacje eksperymentalna na temat ilości
schematów redukcji wiązań dwusiarczkowych dla hTrx, zdecydowałem się na analizę typu
dwell time bardzo dużej ilości danych otrzymanych metoda FC AFM.
Co ciekawe, po otrzymaniu i opracowaniu dużej ilości danych udało mi się wyodrębnić nie
jeden, ale dwa schematy redukcji wiązań dwusiarczkowych przez hTrx, patrz Rys. 7. W
omawianej publikacji przedyskutowałem bardzo dokładnie otrzymane wyniki ze
szczegółowym uwzględnieniem wcześniejszych doniesień literaturowych. W rezultacie,

11
jeden ze schematów redukcji przypisałem schematowi katalizy typu Michaelisa-Mentena, w
wariancie, który jest zależny od siły mechanicznej. Kataliza zachodząca według schematu
Michaelisa-Mentena jest zjawiskiem częstym dla enzymów biologicznych. Pierwszy etap
takiej reakcji polega na wiązaniu się enzymu do białka. Właściwa redukcja, czyli wymiana
elektronu pomiędzy katalitycznymi cysteinami w tioredoksynie a wiązaniem
dwusiarczkowym, zachodzi natomiast w etapie drugim, i w omawianym przypadku jest
zależna od siły mechanicznej rozciągającej wiązanie dwusiarczkowe uwięzione w białku.
Odkryty tutaj drugi schemat redukcji rzeczywiście okazał się bardzo słabo zależny od siły
napinającej i rozciągającej wiązanie dwusiarczkowe. Tym bardziej więc prawdopodobną
okazała się hipoteza redukcji wiązania –S-S- przy dzięki tunelowania elektronów (ET), którą
rozważyłem bardzo szczegółowo, co pokrótce przedstawiam poniżej.
Tunelowanie nie powoduje rozerwania żadnych wiązań i stąd też proces ten nie
charakteryzuje się żadną energią aktywacji, ani istnieniem stanów przejściowych. Niemniej
jednak system musi się przygotować poprzez utworzenie reaktywnych rodników, co skutkuje
istnieniem bariery energetycznej w postaci energii reorganizacji. W rozpatrywanym
przypadku ET wydaje się zachodzić z rodnika tiylowego (-S) z jednej z katalitycznych
cystein wewnątrz centrum aktywnego tioredoksyny bezpośrednio na jeden z atomów siarki z
wiązania dwusiarczkowego w badanej modelowej proteinie (I27SS)8. W wyniku tunelowania
wiązanie –S-S- ulega przekształceniu bez żadnej bariery energetycznej w tiolan (-S-) i rodnik
tiylowy, które automatycznie są separowane dzięki sile rozciągającej to wiązanie.
Stała szybkości procesu ET, kET, jest zazwyczaj wyrażana poprzez pół-klasyczną teorię nie-
adiabatycznego transferu elektronu bazującą na tzn. złotej regule Fermiego i zobrazowanej w
Równaniu 2:
kET = (4/h)* (4kBT)-1/2
*V*exp[-(ΔG+)
2/(4kBT)]. Równanie 2
W równaniu 2: h to stała Planka, kB to stała Boltzmana, T to temperature, to energia
reorganizacji, V to element macierzy hybrydyzacyjnej odpowiadający nakładaniu się
potencjałów elektronowych donora i akceptora, zaś ΔG to różnica potencjałów redoks między
cysteiną katalityczną, z której będzie następować tunelowanie, a cysteiną w wiązaniu
dwusiarczkowym, na którą będzie następować tunelowanie.
Ze względu na dużą trudność w znalezieniu wartości potencjału V dla białek w roztworach,
wartość kET jest częstokroć przybliżana za pomocą fenomenologicznego równania Mosera-
Dattona, przedstawionego w Równaniu 3:
log(kET) 13 – 0.6(RDA – 3.6) – 3.1[(ΔG +)2/] Równanie 3
gdzie: RDA to średnia odległość miedzy donorem a akceptorem elektronu.
Bazując na danych eksperymentalnych oraz dostępnych danych literaturowych oszacowałem
wszystkie zmienne i parametry w równaniu 3. Otrzymałem wartość stałej kET pomiędzy 2,000
s-1
a 7,000 s-1
co jest cztery rzędy wielkości więcej niż wartości stałych k otrzymanych
doświadczalnie dla schematu drugiego, patrz Rys. 5. Stąd też, mimo, że nie można w

12
omawianym przypadku wykluczyć zjawiska ET, zaproponowałem inny mechanizm drugiego
schematu redukcji wiązania dwusiarczkowego przez hTrx.
Ze względu na często występujące współzawodnictwo procesów SN2 z procesami
dwucząsteczkowej eliminacji E2, zaproponowany drugi schemat redukcji –S-S- zawiera dwa
etapy. W pierwszym etapie następuje eliminacja E2 na atomie węgla w najbliższym
otoczeniu wiązania dwusiarczkowego, z utworzeniem jonu wodorkowego H-. W etapie
drugim następuje rozpad wiązania dwusiarczkowego w wyniku substytucji nukleofilowej z
udziałem anionu H-. Szczególy zaproponowanego mechanizmu są dokładnie
przedyskutowane w omawianym artykule.
Reasumując, stosując metodę histogramów „dwell time” do analizy danych FC AFM byłem
w stanie zagłębić się w molekularne mechanizm redukcji wiązania –S-S- przez hTrx i
zaproponować nowy mechanizm do nieznanego wcześniej schematu enzymatycznej redukcji
wiązań dwusiarczkowych.
Rysunek 5. Dwa schematy redukcji
wiązania dwusiarczkowego przez
tioredoksynę ludzką. Schemat
oznaczony „Path A” został
przyporządkowany znanemu
wcześniej w literaturze schematowi
Michaelisa-Mentena. Schemat
oznaczony „Path B” okazał się
nieznanym wcześniej schematem,
które został szczegółowo omówiony w
omawianej publikacji.
J. Cruz, P. H. Pfromm, R. Szoszkiewicz, and M. E. Rezac, Hydrolases on Silica Surfaces:
Coverage-Activity-Molecular Property Relationships Revealed, Process Biochemistry
49(5), 830-839 (2014).
W kolejnym z omawianych artykułów również studiowaliśmy zależności między strukturą i
właściwościami mechanicznymi, a aktywnością enzymatyczną wybranych białek-enzymów.
Tym razem jednak przeszliśmy do studiów aktywności enzymatycznej paru typów hydrolaz
w fukcji ich całościowej konformacji. Wybrane hydrolazy to: Candida antarctica lipaza B
(CALB), subtilisin Carlsberg, oraz lipaza Thermomyces lanuginosus (TLL).
Wcześniejsze badania opisały nową platformę katalityczną dla reakcji w rozworach, która
oparta została na unieruchomieniu pojedynczych cząsteczek enzymów na rozdrobnionych
nanocząstkach krzemionki. Cząsteczki enzymów zostały fizycznie zaadsorbowane na
nanocząstkach i zliofilizowane w celu otrzymania nanobiokatalizatorów. Dla przykładu, S.
Carlsberg i CALB zostały osadzone na krzemionce koloidalnej, zliofilizowane, a następnie
testowane w środowisku heksanu poprzez pomiar stałych szybkości reakcji dla pewnych
typów katalizy o znaczeniu komercyjnym. Zaobserwowane aktywności katalityczne zależały
od warunków reakcji i pokrycia powierzchni przez cząsteczki enzymów, niemniej jednak

13
związki pomiędzy uśrednioną konformacją cząsteczek enzymów i wzajemnym
oddziaływaniem cząsteczek enzymów a ich aktywnością katalityczną nie zostały wcześniej
szczegółowo poznane i wyjaśnione. Stąd też w opisywanej publikacji skoncentrowaliśmy się
na dostarczeniu odpowiedzi na te pytania, a więc znalezieniu związków między morfologią
zaadsorbowanych cząsteczek enzymów CALB, S. Carlsberg, i TLL na modelowej
powierzchni krzemionkowej a ich aktywnością katalityczną. W tym celu dokonaliśmy
szczegółowej analizy konformacji zaadsorbowanych enzymów dzięki wysoko-rozdzielczemu
obrazowaniu metodami mikroskopii sił atomowych (AFM). Dane AFM zostały
przenalizowane w celu wyznaczenia: wielkości, rozkładów gęstości powierzchniowej i
objętości zaadsorbowanych agregatów cząsteczek enzymatycznych. Próbkowaliśmy szeroki
wachlarz stężeń powierzchniowych enzymów: od agregatów składających się z pojedynczych
cząstek w bardzo małym stężeniu powierzchniowym do powierzchni całkowicie pokrytych
cząsteczkami enzymów. Dane te zostały przyporządkowane do badań aktywności
katalitycznej, a także sztywności agregatów enzymatycznych w celu rozróżnienia między
enzymatycznie aktywnymi i nie-aktywnymi konformacjami enzymów, patrz Rysunek 6. W
rezultacie, udało nam się wyznaczyć szereg reguł, które pozwoliły na zrozumienie i
znalezienie związków pomiędzy zmierzonymi parametrami topografii zaadsorbowanych
cząstek enzymów z ich sztywnością cząsteczkową i aktywnością katalityczną.
Rysunek 6. Schematyczny model
oddziaływania badanych cząstek enzymów
CALB, s. Carlsberg, i TLL z powierzchnią
w celu wyjaśnienia ich aktywności
katalitycznej z procentowym pokryciem
powierzchni (% SC). Nasz model powstał
na bazie obserwacji topograficznych z
użyciem wysokorozdzielczej mikroskopii
AFM, a także obserwacji sztywności
enzymów rozumianej jako ich podatności
na deformację mechaniczną. Strzałki
skierowane w górę lub dół oznaczają
odpowiednio wzrost lub spadek aktywności
katalitycznej. Zwiększająca się sztywność
od CALB to TLL została oznaczona
poprzez zwiększającą się grubość konturów
dla narysowanych reprezentacji cząsteczek
enzymów.
Rozwój Metod Badawczych w Celu Badania Zmian Konformacyjnych
Białek w Procesach ich Fałdowania i Rozfałdowywania Indukowanych
przez Siłę Mechaniczną. W toku dalszej swojej kariery badawczej skupiłem się na badaniach in vitro zmian
konformacyjnych pojedynczych cząsteczek modelowych białek o funkcjach mechanicznych
in vivo w trakcie procesów ich fałdowania i rozfałdowywania z użyciem siły mechanicznej.

14
Celem tych badań było zrozumienie tych procesów, a także dostarczenie nowych metod
badawczych do ich poznania.
Dey, R. Szoszkiewicz, Complete noise analysis of a force spectroscopy AFM setup and
its applications to study nanomechanics of human Notch 1 protein, Nanotechnology 23,
175101, 12 pages (2012).
W początkowym etapie rozwoju pracowni Szoszlab na Uniwersytecie Stanowym w Kansas,
skupiłem się na konstrukcji i charakteryzacji nowego spectrometru FX, FC AFM w celu
dokładniejszego niż dotychczas badania zmian konformacyjnych białek w wyniku ich
zwijania i rozwijania przy użyciu siły mechanicznej, patrz Rysunek 7.
Rysunek 7. Podstawy spektroskopii krzywych siłowych (ang. force-extension AFM, FX AFM) i
nowy spektrometr FX, FC AFM skonstruowany w pracowni Szosz-lab na Uniwersytecie Stanowym
w Kansas. A) W badaniach rozwijania pojedynczych cząsteczek białek za pomocą siły mechanicznej
badana cząsteczka białka jest sflankowane przez kowalencyjnie związane inne nisko-cząsteczkowe
białko, jak np. I27, o znanej odpowiedzi mechanicznej na mechaniczne rozciąganie/zwijanie. Celem
takiego przygotowywania eksperymentu jest dostarczenie tzw. „odcisku palca”, a więc pewności, że
rozciągane będzie interesujące nas białko w ściśle określonej konfiguracji i to w formie natywnej. W
przypadku spektroskopii AFM w modzie krzywych siłowych (FX AFM) badana próbka z cząsteczkami
białka zaabsorbowanymi na modelowych powierzchniach jest przybliżana i następnie oddalana
do/od belki AFMu (AFM Cantilever) przez skaner AFM (AFM Piezo) ze stałą szybkością. W
przypadku związania się cząsteczki badanego białka z ostrzem belki AFMu następuje szereg ugięć
tejże belki w funkcji zmieniającej się dlugości cząsteczki białka. Ugięcia te są monitorowane przez
światło lasera odbijające się od tylnej powierzchni ostrza i zbieranego przez fotodetektor. Po
kalibracji, sygnał ugięcia belki jest konwertowany na siłę mechaniczną. Wówczas zmiany
konformacyjne cząsteczek białka mierzone są poprzez zmiany siły ciągnięcia w funkcji długości
rozwijanej cząsteczki białka, co nie zostało tu pokazane. B) schemat spektrometru FX, FC AFM
zbudowanego w pracowni Szosz-lab. C) Zdjęcie elektroniki użytej w tym spektrometrze. D)
Skonstruowany zestaw FX, FC AFM pozwolił na uzyskanie konfiguracji otwartej na dodatkową
wiązkę lasera oświetlającą zestaw eksperymentalny od tylnej strony belki AFM. Dzięki temu możliwe
stało się powiązanie badań spektroskopii sił AFM z użyciem tego zestawu, z dodatkowymi
badaniami, jak np. zaprezentowana tu interferometria heterodynowa, dane jeszcze nieopublikowane.
Skonstruowaliśmy nasz zestaw FX, FC AFM według najnowszego stanu wiedzy na lata
2008-2009, w których następowała jego budowa w pracowni Szosz-lab. Użyliśmy
dostępnych komercyjnie komponentów elektronicznych, standardowych adaptacji
C) D) D)

15
mechanicznych i własnoręcznie napisanego oprogramowania opartego o oprogramowanie
dostępne w pracowni Prof. Julio Fernandeza na Uniwersytecie Columbia w Nowym Jorku, w
Stanach Zjednoczonych Ameryki Północnej. Niewątpliwe zalety naszego systemu leżą
zarówno w jego otwartej konfiguracji sprzętowej jak i otwartej platformie oprogramowania.
Podobna platforma oprogramowania jest wykorzystywana od niedawna również przez
czołowych producentów systemów komercyjnych, jak Asylum Research, USA. Nasz system
został dokładnie opisany w omawianej publikacji, a więc jego transfer do innych pracowni i
sprzęganie z innymi technikami badawczymi wydaje się jak najbardziej możliwe.
Scharakteryzowaliśmy nasz spektrometer AFM głównie poprzez pomiary jego mechaniczno-
elektronicznego szumu własnego i szybkości jego odpowiedzi na zmiany konformacyjne w
cząsteczkach modelowego białka dla typowych belek AFM stosowanych w spektroskopii sił.
Po uzyskaniu pomiarów okazało się, że parametry naszego spektrometru przewyższają
parametry najlepszych zestawów komercyjnych, jak np. mikroskop Cypher produkowany
przez firmę Asylum Research, USA. Charakterystyka szumów własnych naszego
spektrometru jest zdominowana przez termiczne szumy Browna belki pomiarowej AFM, a
więc limit eksperymentalny. Dla przykładu, nie-brownowska komponenta szumów
przecałkowana dla pierwszego termicznie wzbudzonego modu fleksyjnego przykładowej
belki AFM wynosiła mniej niż 2% całkowitej mocy szumu. Czas odpowiedzi spektrometru
dla przykładowej belki AFMu okazał się dwa razy krótszy niż dla prezentowanych wówczas
w literaturze spektrometrów i jedynie około dwa razy dłuższy niż graniczna wartość
teoretyczna. Również i inne parametry oraz właściwości naszego spektrometru zostały
opisane w prezentowanej publikacji.
W następnym kroku zaaplikowaliśmy nasz spektrometr do badania wymuszonego przez siłę
mechaniczną rozfałdowywania się konstruktu białkowego zawierającego domenę NRR (ang.
Negative Regulatory Domain) z ludzkiej proteiny Notch1. Celem tych badań było
zrozumienie nieznanych jeszcze aspektów molekularnego mechanizmu funkcjonowania
szlaku Notch-a. Na podstawie zebranych danych FX AFM wyodrębniliśmy conajmniej pięć
istotnych zmian konformacyjnych w procesie rozwijania się białka NRR. Rezultaty te
stanowiły solidne podwaliny do dalszych studiów indukowanych siła mechaniczną procesów
rozwijania sie domeny NRR. Procesy te stanowić będą temat jednej z kolejnych publikacji w
przedstawianym cyklu habilitacyjnym.
R. Szoszkiewicz, Comment about “print your AFM” paper, Review Scientific
Instruments, 83, 037101, 3 pages, (2012). To również była publikacja recenzowana w
systemie peer-review.
W tej publikacji skupiłem się na wyjaśnieniu pewnych istotnych cech spektroskopii sił przy
użyciu AFMu, aby zwrócić uwagę na pewne często popełniane błędy w interpretacji szumu
eksperymentalnego w spektrometrach AFMu. Wytłumaczyłem między innymi jak powinno
się obliczać szum w drganiach belki AFMu. Następnie rozróżniłem przepustowość (ang.
bandwidth) elektroniczną i mechaniczną systemów AFM, wyłuszczyłem ich typowe
ograniczenia, i wytłumaczyłem jak używać tych informacji w celu charakterystyki systemów
spektroskopii sił. W szczególności pokazałem, że przepustowość elektroniczna ma wpływ na

16
amplitudę odpowiedzi belki AFMu, zaś przepustowość mechaniczna definije szybkość
odpowiedzi belki AFMu.
Zmiany Konformacyjne Białek Związane z Procesami ich Rozwijania N. Ploscariu, K. Kuczera, K. E. Malek, M. Wawrzyniuk, A. Dey, and R. Szoszkiewicz,
Single Molecule Studies of Force-Induced S2 Site Exposure in the Mammalian Notch
Negative Regulatory Domain, Journal of Physical Chemistry B, 118(18), 4761-4770
(2014).
Dzięki konstrukcji nowego spektrometru AFM w pracowni Szosz-lab, mogłem skupić się na
studiach mechanicznie indukowanych zmian konformacyjnych w białkach, które mają
znaczenie w transdukcji sygnałów mechanicznych. Przykładem takiego białka jest białko
Notch uczestniczące w blisko-zasięgowej komunikacji międzykomórkowej.
Rysunek 8. Badania procesów aktywacji białka Notch z użyciem spektroskopii sił AFM. STRONA
LEWA: Białko transmembranowe Notch składa się z ektodomeny w skład której wchodzą domeny
EGF-podobne i domena NRR. Część wewnątrzkomórkowa zawiera domenę ICN. Kanoniczny proces
sygnalizacji z użyciem tego białka jest aktywowany gdy którykolwiek z ligandów z rodziny Delta-
Serrate-Lag2 (DLS) zwiąże się do ektodomeny receptorów Notch. Oddziaływania ligand-receptor
powodują zmiany konformacyjne w strukturze domeny NRR, które umożliwiają odsłanianie i cięcie
wiązania peptydowego w miejscu S2 zlokalizowanego w okolicach C-końca domeny NRR. Cięcie to
następuje z użyciem jednej z proteaz rodziny disintegrin i metaloproteaz (ADAM). Następnie,
następuje kolejne cięcie już w części śródmembranowej białka, w miejscu S3, przez γ-sekretazę. W
wyniku cięcia w miejscu S3 uwalniania jest domena ICN, która jest translokowana do jądra
komórkowego, gdzie przyłącza się do niej białko CLS w celu aktywacji ekspresji stosownych genów.
ŚRODEK: Schemat ekperymentu FX AFM w celu badania indukowanego siłą rozciągania białka
NRR. STRONA PRAWA: histogram otrzymanych danych z FX AFM pokazujący cztery klasy
głównych zmian konformacyjnych w proteinie NRR zaobserwowane na skutek jej mechanicznego
rozciągania. Każda z klas odpowiada pewnemu zakresowi długości rozciąganego białka NRR, przy
których zaobserwowano zrywanie stosownych oddziaływań wewnątrz rozfałdowywującej się domeny
NRR. Jak wywnioskowaliśmy z badań struktury krystalicznej białka NRR, a także symulacji
komputerowych, klasa trzecia zmian konformacyjnych zmierzona w trakcie naszych eksperymentów
odpowiada otworzeniu się miejsca S2 dla ataku proteaz.
Komunikacja komórkowa szlakiem Notch-a to bardzo ważny i wysoce konserwatywny szlak
sygnalizacyjny. Kanoniczny mechanizm sygnalizacji za pomocą białka Notch to piękny,
wielostopniowy proces, który został zobrazowany na Rysunku 8. Szlak Notch-a kontroluje
proliferację pewnych typów komórek, śmierć komórkową, a także programy różnicowania

17
się komórek w całym podkrólestwie zwierząt wielokomórkowych. Procesy regulacji
transkrypcji genowej za pomocą szlaku białka Notch są szeroko opisane w literaturze
fachowej przez liczne doniesienia strukturalne i biochemiczne. Jednakże, wiele aspektów
mechanicznych tego szlaku nie jest jeszcze poznanych. W szczególności, niewiele wiadomo
o mechaniźmie odsłaniania i cięcia wiązania peptydowego w miejscu S2 w domenie NRR,
które to cięcie jest kluczowym elementem kaskady sygnalizacyjnej białka Notch i w co
najprawdopodobniej zaangażowana jest również siła mechaniczna.
Używając metod FX AFM dla domeny NRR1 ze ssaczego analogu białka Notch1 (mNRR1)
wraz z odpowiednimi danymi symulacjami dynamiki molekularnej typu „steered molecular
dynamics” dostarczyliśmy zrozumienia procesu odsłaniania miejsca S2 na skutek siły
mechanicznej. Aby przeanalizować odsłanianie się miejsca S2 w ściśle określonej geometrii
rozciągającej zsytetyzowaliśmy zrekombinowane białka I272−mNRR1−I272, gdzie domena
mNRR1 została sflankowana przez dobrze poznane standardy mechaniczne, a mianowicie
białka I27, patrz Rysunek 8. Następnie dostarczyliśmy testów biofizycznych i
biochemicznych pozwalających ocenić aktywność biochemiczną otrzymanych konstruktów
mNRR1. Tutaj między innymi przeprowadziliśmy testy cięcia I272−mNRR1−I272 przez
stosowne proteazy z rodziny ADAM-ów. Następnie, używając spektroskopii sił FX AFM
zebraliśmy dane zmian konformacyjnych w białku I272−mNRR1−I272 na skutek jego
rozciągania mechanicznego przy pomocy belki AFM. W szczególności wyznaczyliśmy
przedziały długości rozciąganego białka (N-to-C termini lengths) przy których następują
poważniejsze zmiany konformacyjne wewnątrz domeny mNRR1. Dzięki dostępnym danym
literaturowym jak i symulacjom dynamiki molekularnej zgrupowaliśmy otrzymane zmiany
konformacyjne w cztery klasy, odpowiadające czterem najbardziej prawdopodobnym krokom
procesu rozwijania się domeny mNRR1. Dokładnie przedyskutowaliśmy zarówno możliwe
zmiany konformacyjne w każdej z klas jak i zakres i znaczenie średnich sił potrzebnych do
realizacji każdej z tych klas. Następnie przeprowadziliśmy analizę prawdopodobieństwa
warunkowego, która poparła istniejącą w literaturze hipotezę o sekwencyjnym rozwijaniu się
domeny mNRR1, tzn. rozfałdowywaniu się według schematu klasa 1 -> klasa 2 -> klasa 3 ->
klasa 4. Na podstawie naszych symulacji komputerowych przypisaliśmy pierwsze trzy klasy
rozwijania się mNRR1 procesom uwieńczonym całkowitym odsłonieniem miejsca S2.
Zmierzone średnie siły potrzebne do realizacji zdarzeń w każdym z pierwszych trzech klas
zdarzeń były pomiędzy 70 pN a 90 pN i przy szybkościach ciągnięcia 400 nm/s. Pokazaliśmy
następnie, że te bardzo znaczące siły stanowią efektywną barierę przed odsłonięciem miejsca
S2 in vivo, i wymagają skoordynowanej, a nie przypadkowej, aplikacji siły i to za pomocą
paru kroków. Nasze rezultaty badawcze otworzyły drogę do studiów aktywacji receptorów
białek typu Notch na poziomie pojedynczej cząsteczki w warunkach fizjologicznych.
Zmiany Konformacyjne Związane z Fałdowaniem się Wybranych
Białek. Oprócz studiowania procesów mechanicznie indukowanego rozfałdowywania się wybranych
białek, skupiłem się również na nowatorskich badaniach biofizycznych procesu fałdowania
się białek na poziomie pojedynczej cząsteczki.

18
Proces fałdowania się białek został uznany za jedno z największych wyzwań naukowych
następnego ćwierćwiecza w roku 2005 przez Science Magazine, i to pomimo ponad 40 lat
badań na ten temat. Jest tak dlatego, że liczba dostępnych schematów procesu zwijania białek
i to nawet dla małych białek jest astronomicznie duża a środowisko komórkowe, w którym
odbywa się ten proces in vivo jest zatłoczone przez wielu innych cząsteczek, które mogą
pomóc, ale też utrudnić, lub nawet odwrócić ten proces. Co więcej, proces zwijania się białek
ma związek z wieloma chorobami, takich jak choroba Creutzfelda-Jakoba, choroba
Alzheimera, choroba Huntingtona, i choroba Parkinsona. Błędy w procesie zwijania się
białek lub ich nadmierna degradacja prowadzą do chorób takich jak mukowiscydoza. Stąd też
studia procesów zwijania się białek stały się jednym z centralnych pytań nauki na pograniczu
fizyki, chemii i biologii.
Eksperymenty z wykorzystaniem spektrometrii FX FC AFM są w stanie wywierać na
pojedyncze cząsteczki siły rzędu pikoniutonów, które to siły są siłami fizjologicznymi i stąd
są aplikowalne do studiów rozfałdowywania i fałdowania się pojedynczych cząsteczek
białek. Dodatkowo, wydaje się, że to właśnie szczegółowe badania zmian konformacyjne w
czasie trajektorii fałdowania się prostych białek są w stanie przynieść znaczący postęp w
dogłębnym zrozumieniu procesów fałdowania się białek. Niemniej jednak zarówno
rozdzielczość w detekcji zmian długości rozciąganego i zwijanego białka jak i czasów tych
zmian są wciąż niewystarczające do badania szybkich zmian konformacyjnych w
niskocząsteczkowych białkach. Eksperymentalnie osiągalna kontrola pozycji belki AFM jest
obecnie rzędu paru nanometrów, i co za tym idzie śledzenie zmian konformacyjnych białek w
tym zakresie, nie jest obecnie możliwe dla giętkich belek AFM zoptymalizowanych w celu
wywierania na białka sił w zakresie paru pN. Obecne możliwości detekcji zmian pozycji
belek AFMu z rozdzielczością milisekundową są również niewystarczające dla badań
szybkich przejść konformacyjnych w prostych białkach. Stąd też, postanowiłem skupić się na
detekcji procesów zwijania się prostych białek poprzez detekcję własciwości mechanicznych
badanych cząsteczek.
K. E. Malek and R. Szoszkiewicz, Changes of protein stiffness during folding detect
protein folding intermediates, Journal of Biological Physics 40(1), 15-23 (2014).
W omawianej publikacji pokazaliśmy jak zmierzyć zmiany sztywności mechanicznej prostej
cząsteczki białka I27 w trakcie jej fałdowania się in situ w konfiguracji eksperymentalnej
typu force-clamp AFM, a więc podobnie jak na Rysunku 1. Wybraliśmy cząsteczkę I27
bowiem jest ona przykładem prostej cząsteczki białka z jednym typem struktury
drugorzędowej, tutaj: struktury typu beta kartka. Następnie pokazaliśmy, patrz Rysunek 9,
jak wywnioskować ze zmiany sztywności mechanicznej obecność zbioru tranzytywnych
stanów przejściowych w procesie fałdowania się białka I27. Stany te składają się
najprawdopodobniej ze skolapsowanych, niesfałdowanych jeszcze struktur I27. Wedle
opublikowanych wcześniej symulacji dynamiki Langevina przez grupę Prof. Thirumalai-a z
Uniwersytetu College Park w stanie Maryland, USA, struktury te pojawiają się w trakcie
fałdowania się cząsteczek I27 zarówno ze stanów rozciągniętych przez siłę mechaniczną jak i
ze stanów uprzednio zdenaturowanych termicznie.

19
Rysunek 9. Tranzytywne sztywności fałdującej się cząsteczki I27. Panel lewy: Używając
eksperymentów typu force-quench AFM dla cząsteczek I274 byliśmy w stanie rozfałdowywać, a
następnie pozwolić cząsteczkom I27 sfałdować się z powrotem. Podobnie jak na Rysunku 1, dla modu
FC AFM, w początkowym stadium cząsteczki I27 są rozciągane, a proces rozwinięcia się każdego z
modułów I27 powoduje skokową zmianę długości rozciąganego białka, co jest zaznaczone za pomocą
gwiazdek. Po rozwinięciu się całej struktury I274, siła rozciągająca zostaje powoli zmniejszana i
molekuła I274 zaczynała się zwijać, co jest widoczne w zmniejszaniu się długości rozciągniętego
białka. Po pozostawieniu cząsteczki białka przez parę sekund przy bardzo małej sile rozciągającej
(tutaj ~15 pN), siła rozciągająca jest powtórnie zwiększana. W zaprezentowanym przykładzie
cząsteczka I27 nie zdołała się poprawnie sfałdować co widać brakiem skokowej zmiany długości
białka. Panel prawy: Dzieląc przez siebie cząstkowe zmiany długości (DL) rozciąganej cząsterczki
I274 przez zmiany siły rozciągającej (DF) policzyliśmy cząsteczkową giętkość (ang. compliance), Cm,
podczas trajektori fałdowania się cząsteczek I27. Następnie, obliczyliśmy średnie kwadratowe
fluktuacje długości <(L)2> fałdującej się proteiny względem wartości średnich zmian długości
białka. Następnie otrzymaliśmy zależność <(L)2> vs. Cm, której tu nie pokazano. I wreszcie,
obliczyliśmy autokorelację zależności <(L)2> vs. Cm. Otrzymane maksima lokalne tak
przedstawionej tu autokorelacji przyporządkowaliśmy zbiorowi tranzytywnych „sztywności”
cząsteczki białka. Obliczeń dokonano wewnątrz zacieniowanego na panelu górnym (ang. „region of
interest”) i w interwałach czasowych 1 ms. Pokazane dane to rezultaty uśrednień dla sześciu
trajektorii z udziałem czterech domen I27, 25 trajektorii z udziałem trzech domen I27, 20 trajektorii z
udziałem dwóch domen I27, i 10ciu trajektorii z udziałem jednej domeny I27.
N. Ploscariu, R. Szoszkiewicz, A method to measure nanomechanical properties of
biological objects, Applied Physics Letters 103, 263702 (2013).
Jak zauważyliśmy w omówionej powyżej pracy, znajmość „giętkości” bądź „sztywności”
cząsteczkowych nie jest wystarczająca do rozróżnienia pomiędzy różnymi konformacjami
przejściowymi fałdującej się cząsteczki białka. Jednakże, jeśli moglibyśmy zmierzyć więcej
parametrów mechanicznych, wówczas dokładniejsza charakteryzacja fałdującej się cząsteczki
byłaby możliwa. Stąd też zaadoptowaliśmy model reologiczny Kelvina-Voighta, aby w

20
przyszłości wyznaczyć nie tylko molekularną sztywność fałdującej się cząsteczki wzdłuż
kierunku działania siły mechanicznej, ale tzw. sygnaturę mechaniczną fałdującej się
cząsteczki. Zaadoptowana przez nas sygnatura mechaniczna, patrz Rysunek 10, składa się z
czterech parametrów: dwóch wartości sztywności wzdłuż dwóch ortogonalnych kierunków, a
także związanych z nimi molekularnych czynników tłumienia. Następnie opracowaliśmy
model analityczny w celu otrzymania sygnatury mechanicznej fałdujących się białek z
pomiarów zachowania się częstostotliwości rezonansowych belek AFM w kontakcie z
rozwijającymi się białkami. Punktem wyjścia jest znajomość geometrycznych i
mechanicznych właściwości belki AFMu oraz lepkości i gęstości roztworu. Model ten został
opracowany dla typowych giętkich belek AFM o małym ilorazie ich długości do szerokości i
biorąc pod uwagę opór hydrodynamiczny roztworu. Używając tego modelu obliczyliśmy
częstotliwości rezonansowe studiowanych belek AFM w powietrzu, a następnie zmiany tych
częstotliwości dla belek w roztworze typowego buforu i wreszcie belek w buforze i w
kontakcie z natywnymi prostymi białkami, jak I27. Założyliśmy przy tym parametry
sygnatury mechanicznej I27 wywnioskowane z dostępnych danych literaturowych. W
przyszłości nasz model powinien mieć zastosowanie do detekcji zmian mechanicznych w
strukturze białek w różnych stadiach ich zwijania się, co jest celem moich przyszłych prac w
tym kierunku badawczym.
Rysunek 10. Sygnatura mechaniczna proteiny w kontakcie z belką AFM. Panel lewy: Model
reologiczny obrazujący cząsteczkę białka jako dwie sprężynki dyssypatywne o uśrednionej sztywności
(k) i czynnikach dyssypacji energii () odpowiednio w kierunkach normalnym i prostopadłym do
powierzchni próbki. Panel prawy:„Experimental data”, dane w kolorze czerwonym; wykres ten
obrazuje transformaty Fouriera z detekcji drgań belki AFM obrazujące pierwszych pięć modów
fleksyjnych tejże belki w roztworze typowego buforu (tutaj: PBS) stosowanego do badań zwijania i
rozciągania białek za pomocą spektroskopii sił AFM. „Generalized hydrodynamic function”, dane w
kolorze niebieskim: wykres ten przedstawia częstotliwości rezonansowe badanej belki AFM w
roztworze PBS wyznaczone za pomocą naszego modelu.”Expected shift in frequency….”, dane w
kolorze zielonym, linia przerywana: wykres przedstawia wyznaczone z naszego modelu zmiany
częstotliwości rezonansowych drgań fleksyjnych belki AFM w kontakcie z białkiem opisanym
sygnaturą mechaniczną z panelu lewego przy następujących parametrach dla sfałdowanej cząsteczki
prostego białka kn = klat = 10 pN/nm and n = lat = 10-8
kg/s w PBSie.
R. Szoszkiewicz, Force-extension (FX) and force-clamp (FC) AFM spectroscopies in
investigating mechanochemical reactions and mechanical properties of single

21
biomolecules in Scanning Probe Microscopy in Nanoscience and Nanotechnology, p.
395-424, edited by B. Bhushan, Springer-Verlag, Heidelberg (2010).
W tym rozdziale książkowym dokonałem przeglądu kluczowych technik eksperymentalnych
używanych obecnie do otrzymania informacji na temat zmian konformacyjnych w
biomolekułach poddanych kontrolowanemu rozciąganiu i kompresji. W szczególności,
przedyskutowałem zastosowania metod typu FX AFM i FC AFM do próbkowania stabilności
mechaniczej pojedynczych cząsteczek białek i polisacharydów, a także studiów procesów
redukcji pojedynczych wiązań dwusiarczkowych w białkach. Dodatkowo, przedstawiłem
swój punkt widzenia na perspektywy rozwoju studiów reakcji mechanochemicznych z
użyciem białek i własności mechanicznych pojedynczych biomolekuł jak białka, cząsteczki
RNA i cząsteczki DNA.
5. Publikacje niemogące wejść w skład osiągnięcia naukowego.
Publikacja opisana poniżej nie mogła wejść w skład opisanego powyżej osiągnięcia gdyż nie
udało mi się dotrzeć do jednego ze współautorów, Dr. Tzu-Ling Kuo. Niemniej jednak dla
kompletu opisuję poniżej tą publikację.
S. Garcia-Manyes, J. Liang, R. Szoszkiewicz, T.-L. Kuo, J. M. Fernandez, Force
activated reactivity switch in a bimolecular chemical reaction, Nature Chemistry, 1, 236
(2009);
Impact Factor: 17.927 (2010) – czasopismo zapoczątkowało istnienie w roku 2009, stąd też pierwszy
opublikowany impact factor pochodzi z roku 2010; 24.537 (5 year).
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 45
Cytowania: Web of Science: 50 Google Scholar: 58
Wkład pracy dr Roberta Szoszkiewicza: 10%. Mój wkład pracy polegał na dostarczeniu danych FC
AFM dla jednego z reduktorów wiązań dwusiarczkowych: molekuły TCEP.
Publikacja ta powstała na bazie naszej studiów w celu poszukiwania odpowiedzi na
podstawowe pytania dotyczące rozciągania się wiązania dwusiarczkowego do stanu
przejściowego w procesie jego redukcji indukowanej siła mechaniczną. W szczególności
nurtowała nas odpowiedź na następujące pytania: jaki potencjał fizyczny opisuje rozciąganie
się i zrywanie wiązań dwusiarczkowych, i czy model Bell-a opisany równaniem nr 1
rzeczywiście stosuje się w całym eksperymentalnie osiągalnym zakresie sił rozciągających
wiązanie –S-S-.
W roku 2006 Dudko et al., PRL, otrzymali, że zależność stałej szybkości rozrywania się
pojedynczych wiązań chemicznych powinna być opisana następującym równaniem:
1
11
0 11exp1)(G
xF
Tk
G
G
xFkFk
B
Równanie 4
W powyższym równianiu wartość v zależy od wyboru właściwego potencjału opisującego ten
proces: v =1 w przypadku modelu Bell-a, v = 2/3 w przypadku potencjału liniowego z

22
rozciąganiem, który transformuje się w zależność kubiczną w bliskiej okolicy stanu
przejściowego, i v = 0.5 w przypadku potencjału harmonicznego dla początkowego stadium
rozciągania wiązania, który to przechodzi przechodzi gwałtownie do zera w wyniku
rozerwania się wiązania i szybkiej separacji uczestniczących w nim atomów.
Aby sprawdzić, które z proponowanych przez równanie 4 rozwiązań jest najbliższe opisu
rzeczywistej sytuacji eksperymentalnej, zaczeliśmy studia zależności stałej k w zakresie
“dużych” sił. Nasze początkowe rezultaty, patrz rysunek 11, potwierdziły znaczące
odchylenia od liniowości ln(k) vs. F. Jednakże, zakres użytych sił nie pozwolił nam na
rozróżnienie, który z opisanych modeli aplikuje się najlepiej do naszych danych. Stąd też
potrzebowaliśmy danych dla większej siły rozciągającej. Używając typowych związków
redukcji jak DTT lub TCEP nie pozwoliło nam na otrzymanie większej ilości danych dla sił
rozciągających powyżej 700 pN, patrz rysunek 11.
Rysunek 11. Wyniki naszych
początkowych studiów FC AFM
pokazujące logarytm naturalny ze stałej
szybkości redukcji pojedynczego
wiązania siarkowego w cząsteczce I278
przez TCEP (oś rzędnych po lewej
stronie) w funkcji siły rozciągającej.
Przedstawiono dwa zbiory niezależnie
otrzymanych danych oznaczone
odpowiednio kolorami niebieskim i
czerwonym. Dla kompletu oś rzędnych po
prowej stronie i dane w kolorze zielonym
przedstawiają logarytm naturalny ze
stałej szybkości procesu rozwijania się
białek I278 do momentu ekspozycji
wiązań dwusiarczkowych, patrz Rysunek
1. Przedstawione dane pokazują
odstępstwa od modelu Bela dla procesów
redukcji wiązań dwusiarczkowych przy
siłach rozciągających powyżej 400 pN.
Dopiero, kiedy zdecydowaliśmy się użyć innego reduktora wiązań dwusiarczkowych, tj.
jonów hydroksylowych, OH-, byliśmy w stanie zebrać dane pomiarowej dla znacząco
większego reżimu sił rozciągających niż wcześniej. Zebraliśmy dane przy różnych
wartościach pH, lub też stężenia jonów OH-, które to pokazujemy na rysunku 12. Dane dla
jonów OH- nakładają się na dane dla TCEPu, co pokazuje najprawdopodobniej ten sam
mechanizm reakcyjny w obu przypadkach.
Jednakże wyniki naszych badań przedstawione na Rysunku 12 przyniosły zaskakujące
rezultaty. Zamiast oczekiwanych odchyleń od modelu Bell-a otrzymaliśmy przejście między
dwoma reżimami reakcyjnymi określonymi odpowiednio parametrami Δx = 0.50 Å i Δx =
0.11 Å. Dane te wytłumaczyliśmy na parę możliwych sposobów, patrz Rysunek 12.
Następnie używając paru różnych mutantów z wiązaniami dwusiarczkowymi w różnych
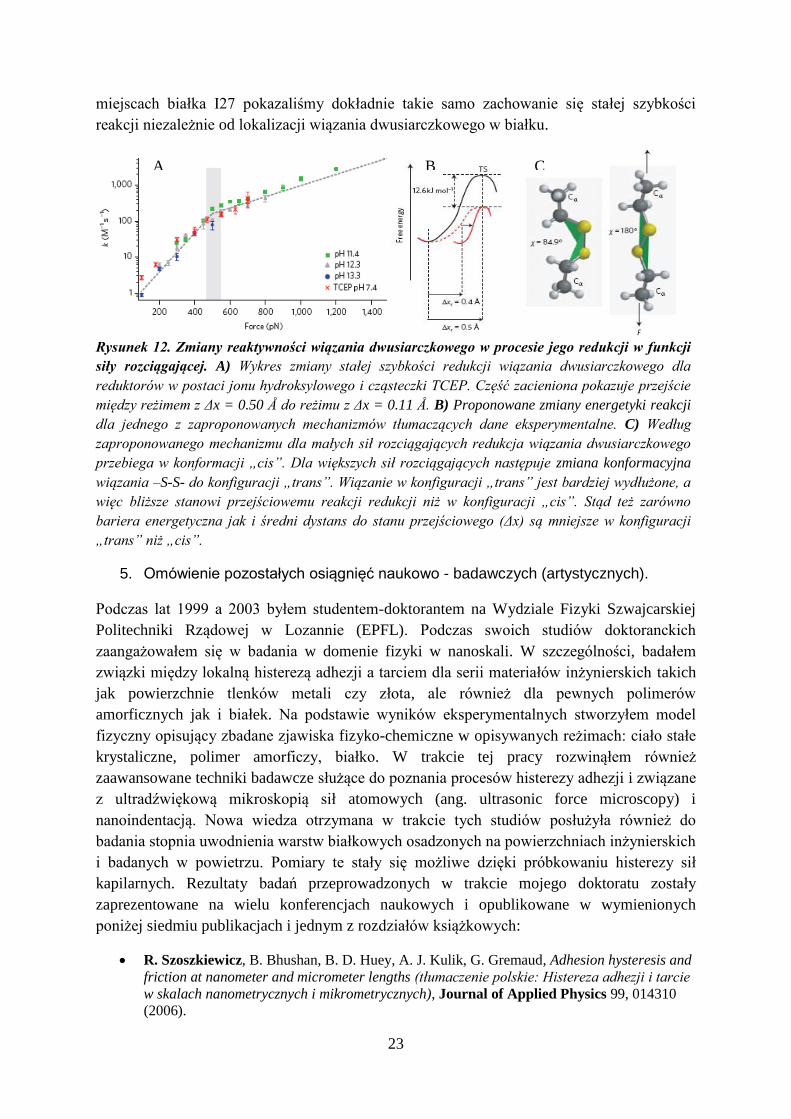
23
miejscach białka I27 pokazaliśmy dokładnie takie samo zachowanie się stałej szybkości
reakcji niezależnie od lokalizacji wiązania dwusiarczkowego w białku.
Rysunek 12. Zmiany reaktywności wiązania dwusiarczkowego w procesie jego redukcji w funkcji
siły rozciągającej. A) Wykres zmiany stałej szybkości redukcji wiązania dwusiarczkowego dla
reduktorów w postaci jonu hydroksylowego i cząsteczki TCEP. Część zacieniona pokazuje przejście
między reżimem z Δx = 0.50 Å do reżimu z Δx = 0.11 Å. B) Proponowane zmiany energetyki reakcji
dla jednego z zaproponowanych mechanizmów tłumaczących dane eksperymentalne. C) Według
zaproponowanego mechanizmu dla małych sił rozciągających redukcja wiązania dwusiarczkowego
przebiega w konformacji „cis”. Dla większych sił rozciągających następuje zmiana konformacyjna
wiązania –S-S- do konfiguracji „trans”. Wiązanie w konfiguracji „trans” jest bardziej wydłużone, a
więc bliższe stanowi przejściowemu reakcji redukcji niż w konfiguracji „cis”. Stąd też zarówno
bariera energetyczna jak i średni dystans do stanu przejściowego (Δx) są mniejsze w konfiguracji
„trans” niż „cis”.
5. Omówienie pozostałych osiągnięć naukowo - badawczych (artystycznych). Podczas lat 1999 a 2003 byłem studentem-doktorantem na Wydziale Fizyki Szwajcarskiej
Politechniki Rządowej w Lozannie (EPFL). Podczas swoich studiów doktoranckich
zaangażowałem się w badania w domenie fizyki w nanoskali. W szczególności, badałem
związki między lokalną histerezą adhezji a tarciem dla serii materiałów inżynierskich takich
jak powierzchnie tlenków metali czy złota, ale również dla pewnych polimerów
amorficznych jak i białek. Na podstawie wyników eksperymentalnych stworzyłem model
fizyczny opisujący zbadane zjawiska fizyko-chemiczne w opisywanych reżimach: ciało stałe
krystaliczne, polimer amorficzy, białko. W trakcie tej pracy rozwinąłem również
zaawansowane techniki badawcze służące do poznania procesów histerezy adhezji i związane
z ultradźwiękową mikroskopią sił atomowych (ang. ultrasonic force microscopy) i
nanoindentacją. Nowa wiedza otrzymana w trakcie tych studiów posłużyła również do
badania stopnia uwodnienia warstw białkowych osadzonych na powierzchniach inżynierskich
i badanych w powietrzu. Pomiary te stały się możliwe dzięki próbkowaniu histerezy sił
kapilarnych. Rezultaty badań przeprowadzonych w trakcie mojego doktoratu zostały
zaprezentowane na wielu konferencjach naukowych i opublikowane w wymienionych
poniżej siedmiu publikacjach i jednym z rozdziałów książkowych:
R. Szoszkiewicz, B. Bhushan, B. D. Huey, A. J. Kulik, G. Gremaud, Adhesion hysteresis and
friction at nanometer and micrometer lengths (tłumaczenie polskie: Histereza adhezji i tarcie
w skalach nanometrycznych i mikrometrycznych), Journal of Applied Physics 99, 014310
(2006).
A B C

24
Impact Factor: 2.316 (2006), 2.259 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 30
Cytowania: Web of Science: 8 Google Scholar: 14
Wkład pracy dr Roberta Szoszkiewicza: 75%; Uczestniczyłem w wytyczaniu tego kierunku
badawczego, zebrałem wszystkie dane eksperymentalnych i zrobiłem ich analizę, uczestniczyłem w
stworzeniu modelu fizycznego opisującego badane korelacje, i wreszcie napisałem tą publikację.
R. Szoszkiewicz, B. Bhushan, B. D. Huey, A. J. Kulik, G. Gremaud, Correlations between
adhesion hysteresis and friction at molecular scales (tłumaczenie polskie: Związki między
histerezą adhezji i tarciem w skalach molekularnych), Journal of Chemical Physics 122, 144708
(2005).
Impact Factor: 3.138 (2005), 3.142 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 20 Google Scholar: 30
Wkład pracy dr Roberta Szoszkiewicza: 75%; Zebrałem wszystkie dane eksperymentalne i zrobiłem
ich analizę, uczestniczyłem w stworzeniu modelu fizycznego, i wreszcie napisałem tą publikację.
R. Szoszkiewicz, A.J. Kulik, G. Gremaud, Quantitative measure of nanoscale adhesion hysteresis
by ultrasonic force microscopy (tłumaczenie polskie: Kwantytatywne pomiary adhezji histerezy w
w nanoskali z użyciem ultradźwiękowej mikroskopii sił atomowych), Journal of Chemical
Physics 122, 134706 (2005).
Impact Factor: 3.138 (2005), 3.142 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 5 Google Scholar: 8
Wkład pracy dr Roberta Szoszkiewicza: 80%; Zebrałem wszystkie dane eksperymentalnych i
zrobiłem ich analizę, uczestniczyłem w stworzeniu modelu fizycznego, i wreszcie napisałem tą
publikację.
R. Szoszkiewicz, A. J. Kulik, G. Gremaud, M. Lekka, Probing local water contents of in vitro
protein films by ultrasonic force microscopy (tłumaczenie polskie: Pomiary lokalnej zawartości
wody w filmach białkowych in vitro z użyciem ultradźwiękowej mikroskopii sił atomowych),
Applied Physics Letters 86, 123901 (2005)
Impact Factor: 4.127 (2005), 3.739 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 40
Cytowania: Web of Science: 18 Google Scholar: 22
Wkład pracy dr Roberta Szoszkiewicza: 80%; Wytyczyłem ten kierunek badawczy, zebrałem
wszystkie dane eksperymentalnych i zrobiłem ich analizę, stworzyłem modelu fizyczny tłumaczący
badane wyniki, i wreszcie napisałem tą publikację.
B. D. Huey, C. Ramanujan, M. Bobji, J. Blendell, G. White, R. Szoszkiewicz, A. J. Kulik, The
importance of distributed loading and cantilever angle in piezo-force microscopy (tłumaczenie
polskie: Znaczenie zdystrybuowanej siły i kąta natarcia belki AFM względem powierzchni próbki
w mikroskopii piezoelektrycznej), Journal of Electroceramics 13, 287-291 (2004).
Impact Factor: 1.124 (2004), 1.227 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 30
Cytowania: Web of Science: 27 Google Scholar: 29

25
Wkład pracy dr Roberta Szoszkiewicza: 5%; Uczestniczyłem w projektowaniu symulacji
komputerowych do tego artykułu i miałem mały wkład w napisanie tego manuskryptu.
R. Szoszkiewicz, G. Gremaud, B. D. Huey, A. J. Kulik, How ultrasound can help with connecting
friction and adhesion hysteresis at local scales (tłumaczenie polskie: Jak ultradźwięki mogą
pomóc w znalezieniu relacji pomiędzy tarciem a histerezą adhezji w skalach lokalnych), rozdział
w książce “Acoustical Imaging 27”, 741-748 (2004).
Impact Factor: none published
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: not on the list
Cytowania: Web of Science: not on the list Google Scholar: not on the list
Wkład pracy dr Roberta Szoszkiewicza: 70%; Zebrałem wszystkie dane eksperymentalne i zrobiłem
ich analizę, a także napisałem tą publikację.
G. Rochat, Y. Leterrier, C. J. G. Plummer, J. A. E. Manson, R. Szoszkiewicz, A. J. Kulik, P.
Fayet, Effect of substrate crystalline morphology on the adhesion of plasma enhanced chemical
vapor deposited thin silicon oxide coatings on polyamide (tłumaczenie polskie: Wpływ
krystalicznej morfologii substratu na adhezję cienkich warstw krzemionki naniesionej metodą
chemicznego osadzania z fazy gazowej na powierzchniach poliamidowych), Journal of Applied
Physics, 95, 5429 (2004).
Impact Factor: 2.255 (2004), 2.259 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 30
Cytowania: Web of Science: 9 Google Scholar: 11
Wkład pracy dr Roberta Szoszkiewicza: 15%; Wykonałem badania lokalnej sztywności powłok
krzemionkowych za pomocą nowatorskiego podejścia z wykorzystaniem techniki SLAM (ang.
scanning local acceleration microscopy), a także dokonałem interpretacji i opisu uzyskanych
wyników.
R. Szoszkiewicz, B. D. Huey, O. V. Kolosov, G. A. D. Briggs, G. Gremaud, A. J. Kulik,
Tribology and ultrasonic hysteresis at local scales (tłumaczenie polskie: Trybologia i histereza
ultradźwiękowa w skalach lokalnych), Applied Surface Science 210, 54 (2003).
Impact Factor: 1.284 (2003), 2.469 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 9 Google Scholar: 13
Wkład pracy dr Roberta Szoszkiewicza: 50%; Zebrałem wszystkie dane eksperymentalne i zrobiłem
ich analizę, a także uczestniczyłem w pisaniu tej publikacji.
Po ukończeniu doktoratu w październiku 2003 zaakceptowałem pozycję naukowca na stażu
podoktorskim na Wydziale Fizyki na Uniwersytecie Technologicznym w Stanie Georgia
(Georgia Tech) poczynając od stycznia 2004 w grupie Prof. Elizy Riedo. W Georgia Tech-u
zajmowałem się badaniami podstawowymi z dziedziny fizyki w nanoskali. Początkowo
zająłem się badaniami kondensacji wody w skalach lokalnych. Badałem procesy formowania
i rozrywania mostków wodnych pomiędzy nanowymiarowymi chropowatościami na
powierzchniach materiałów inżynierskich takich jak szkło czy mika, a atomowo-ostrymi
ostrzami belek AFM. Następnie studiowałem lepkość i procesy dyssypacji energii
mechanicznej pomiędzy warstwami cząsteczek wody usieciowanymi na arbitralnych
powierzchniach hydrofilowych i hydrofobowych. Rezultaty tych badań zostały zaaplikowane

26
póżniej przez innych naukowców do studiów organizacji warstw wody na powierzchniach
żywych komórek. Wyniki tych badań były prezentowane na wielu prezentacjach naukowych
i opublikowane w następujących czterech publikacjach:
R. Szoszkiewicz, E. Riedo, Nanoscopic friction as a probe of local phase transitions
(tłumaczenie polskie: Badania tarcia w nanoskali jak metoda wykrywania lokalnych przejść
fazowych), Applied Physics Letters 87, 033105 (2005).
Impact Factor: 4.127 (2005), 3.739 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 40
Cytowania: Web of Science: 18 Google Scholar: 29
Wkład pracy dr Roberta Szoszkiewicza: 60%; Wykonałem i opracowałem wszystkie dane
eksperymantalne. Pomagałem w pisaniu tej publikacji.
R. Szoszkiewicz, E. Riedo, Nucleation time of nanoscale water bridges (tłumaczenie polskie:
Czas nukleacji mostków wodnych w nanoskali), Physical Review Letters 95, 135502 (2005).
Impact Factor: 7.489 (2005), 7.411 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 45
Cytowania: Web of Science: 57 Google Scholar: 85
Wkład pracy dr Roberta Szoszkiewicza: 80%. Wykonałem i zanalizowałem wszystkie dane
eksperymantalne. Napisałem tą publikację.
T.-D. Li, J. Gao, R. Szoszkiewicz, U. Landman, E. Riedo, Structured and viscous water in
sub-nanometer gaps (tłumaczenie polskie: Ustrukturyzowana i lepka woda w przestrzeniach
sub-nanometrycznych), Physical Review B 75, 115415 (2007)
Impact Factor: 3.172 (2007), 3.564 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 137 Google Scholar: 186
Wkład pracy dr Roberta Szoszkiewicza: 15%; Pomagałem zbierać dane eksperymentalne,
dostarczyłem niektórych obliczeń koniecznych do tej publikacji, pomagałem pisać tą publikację.
L. Sirghi, R. Szoszkiewicz, E. Riedo, Volume of a nanoscale water bridge (tłumaczenie polskie:
Objętość mostku wodnego w nanoskali), Langmuir 22, 1093 (2006)
Impact Factor: 3.902 (2006), 4.489 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 58 Google Scholar: 84
Wkład pracy dr Roberta Szoszkiewicza: 25%; Pomagałem w analizie danych eksperymentalnych i
pisaniu tej publikacji.
Również w Georgia Tech-u, w grupie Prof. Riedo, wynalazłem termochemiczną nanolitografię
ciepłem (ang. thermochemical nanolithography, TCNL). W metodzie tej, gorąca belka AFM
wywołuje sterowaną ciepłem reakcję chemiczną na arbitralnej powierzchni. Koncept tej metody
wymyśliłem i pokazałem dla powierzchni polimerowych. Wyniki tych badań zostały opublikowane w
dwóch publikacjach i dwóch zgłoszeniach patentowych:
R. Szoszkiewicz, T. Okada, S. C. Jones, T.-D. Li, W. P. King, S. R. Marder, E. Riedo, High-
speed, thermochemical nanolithography with sub-15 nm feature size (tłumaczenie polskie:
Szybka nanolitografia termomechaniczna o rozdzielczości sub-15 nanometrowej), Nano
Letters, 7(4), 1064 (2007)

27
Impact Factor: 9.627 (2007), 14.452 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 45
Cytowania: Web of Science: 101 Google Scholar: 124
Wkład pracy dr Roberta Szoszkiewicza: 50%; Wynalazlem tą metodę i dostarczyłem pierwszych
wyników eksperymentalnych i obliczeń. Wniosłem też znaczący wkład w napisanie tej publikacji.
D. Wang, R. Szoszkiewicz, T. Okada, S. C. Jones, M. Lucas, W. P. King, S. R. Marder, E.
Riedo, Local wettability modification by thermochemical nanolithography with write-read-
overwrite capability (tłumaczenie polskie: Lokalne zmiany zwilżania powierzchni
polimerowych z użyciem termolitografi ciepłem z modzie zapisu-ścierania-powtórnego
zapisu), Applied Physics Letters, 91, 243104 (2007).
Impact Factor: 3.596 (2007), 3.739 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 40
Cytowania: Web of Science: 16 Google Scholar: 29
Wkład pracy dr Roberta Szoszkiewicza: 25%. Byłem pierwszym, który zaobserwował potencjał i
możliwości metody TCNL do użycia w modzie zapisu – ścierania – powtórnego zapisu. Zebrałem
pierwsze dane eksperymentalne na ten temat.
E. Riedo, S. Marder, D. Wang, J. Curtis, S. Jones, T. Okada, R. Szoszkiewicz, V. Kodali, C.
Henderson, Y. Hua, W. de Heer, Thermochemical nanolithography: components, systems and
methods (tłumaczenie polskie: Termochemiczna nanolitografia ciepłem: komponenty, systemy
i metody), Dokonano dwóch zgłoszeń patentowych: zgłoszenia w Stanach Zjednoczonych
Ameryki Północnej, zgłoszenie numer 12/791,466 i zgłoszenia w Międzynarodowym
Urzędzie Patentowym w Genewie, w Szwajcarii do patentu o zasięgu ogólno-światowym,
zgłoszenie numer PCT/US10/36871. Obydwa patenty zostały przyznane, odpowiednio w
latach 2013 i 2012. Technologia ta została zlicencjonowana w Stanach Zjednoczonych,
niemniej jednak firma, która wykupiła tą technologię upadła.
Cytowania: Web of Science: 0 Google Scholar: 2
Wkład pracy dr Roberta Szoszkiewicza: 15%. Wynalazłem ten pomysł i dostarczyłem pierwszych
wyników eksperymentalnych i obliczeń potwierdzających możliwości tej metody.
Na dzień dzisiejszy metoda TCNL została bardzo szeroko rozwinięta przez wiele grup badawczych, w
szczególności w Europie i Stanach Zjednoczonych, w aplikacjach wywoływania kontrolowanych
ciepłem reakcji na powierzchniach materiałów półprzewodnikowych, polimerów
elektroluminescencyjnych, do redukcji mono-atomowych warstw tlenku grafenu, a także do
deprotekcji pewnych grup chemicznych na powierzchniach polimerów organicznych w celu
utworzenia matryc do adsorbcji białek i cząsteczek DNA.
Dodatkowo, w trakcie mojej pracy na Georgia Tech-u, dostarczałem wielu analiz za pomocą
mikroskopii AFM, jak np. analizy nanostruktury powierzchni organicznych ogniw fotowoltaicznych
opartych o fulereny i polimery organiczne. Badania te zostały opublikowane w następującej
publikacji:
S. Yoo, W. J. Potscavage Jr., B. Domercq, S.-H. Han, T.-D. Li, S. Jones, R. Szoszkiewicz, D.
Levi, E. Riedo, S. R. Marder, B. Kippelen, Analysis of improved photovoltaic properties of
pentacene/C60 organic solar cells: Effects of exciton blocking layer thickness and thermal
annealing (tłumaczenie polskie: Analiza ogniw słonecznych o udoskonalonych
właściwościach fotowoltaicznych: efekty grubości warstwy blokującej ekscytony a także
wygrzewania), Solid-State Electronics, 51, 1367 (2007).

28
Impact Factor: 1.259 (2007), 1.508 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 25
Cytowania: Web of Science: 79 Google Scholar: 98
Wkład pracy dr Roberta Szoszkiewicza: 5%; Zebrałem i zinterpretowałem część danych AFM do
charakteryzacji morfologicznej powierzchni otrzymanych ogniw fotowoltaicznych.
I wreszcie, również na Georgia Tech-u włożyłem znaczący wkład pracy w napisanie trzech
następujących rozdziałów książkowych:
D. Wang, R. Szoszkiewicz, Vamsi Kodali, Jennifer Curtis, Seth Marder, E. Riedo, A new
AFM based lithography method: Thermochemical Nanolithography (tłumaczenie polskie:
Nowa metoda litograficzna oparta o AFM: Termochemiczna nanolitografia ciepłem), rozdział
w książce: Scanning Probe Microscopy in Nanoscience and Nanotechnology, 795-812,
edytor: Prof. Bharat Bhushan, wydawnictwo: Springer-Verlag, Heidelberg, 2010.
Wkład pracy dr Roberta Szoszkiewicza: 10%; Napisałem małą część tego rozdziału książkowego.
L. Merchan, R. Szoszkiewicz, E. Riedo, NanoMechanics: elasticity in nano-objects
(tłumaczenie polskie: Nanomechanika: elastyczność nano-obiektów), rozdział w książce:
Fundamentals of friction and wear - Springer Book, 219-254, edytorzy: Prof. E. Meyer,
Dr. E. Gnecco, wydawnictwo: Springer-Verlag, Heidelberg, 2007.
Wkład pracy dr Roberta Szoszkiewicza: 40%; Napisałem znaczącą część tego rozdziału książkowego.
R. Szoszkiewicz, E. Riedo, New AFM developments to study elasticity and adhesion at the
nanoscale (tłumaczenie polskie: Nanomechanika: elastyczność nano-obiektów), rozdział w
książce: Applied Scanning Probe Methods V, 269-286, edytorzy: Prof. Bharat Bhushan,
Prof. H.Fuchs, Prof. S. Kawata, wydawnictwo: Springer-Verlag, Heidelberg, 2007.
Cytowania: Web of Science: 1
Wkład pracy dr Roberta Szoszkiewicza: 70%; Napisałem większość tego rozdziału książkowego.
Od stycznia 2007 zaakceptowałem pozycję naukowca na stażu podoktorskim w grupie Prof.
Julio Fernandeza, na Wydziale Nauk Biologicznych Uniwersytetu Columbia w Nowym
Jorku, w Stanach Zjednoczonych Ameryki Północnej. W trakcie tego stażu skupiłem się na
badaniach biofizyki procesów zmian konformacyjnych w białkach indukowanych siłą
mechaniczną. Prace z tego okresu są opisane w przedstawionym tutaj cyklu habilitacyjnym.
Od czerwca 2008 zaakceptowałem samodzielną pozycję typu „Assistant Profesor” na
wydziale Fizyki Uniwersytetu Stanowego w Kansas (KSU), w Stanach Zjednoczonych
Ameryki Północnej. Po rygorystycznej ocenie, tzw. tenure review, w marcu 2014 otrzymałem
awans na stanowisko Associate Professor i tzw. Tenure, a więc stałą posadę na uniwersytecie.
W KSU kontynuowałem swoje prace badawcze z zakresu biofizyki i fizyki materiałowej z
wykorzystaniem nanolitografii TCNL. Angażowałem się również w pomoc innym
naukowcom w badaniach pokrewnych, jak np. badaniach właściwości adsorpcyjnych i
morfologicznych kompleksów pewnych peptydów z cząsteczkami DNA:
Avila-Flores, L. Aps, P. Sukthankar, N. Ploscariu, S. Gudlur, L. Simo, R. Szoszkiewicz, Y.
Park, S. Lee, T. Iwamoto, and J. Tomich, Branched amphiphilic cationic oligo-peptides form
peptiplexes with DNA: A study of their biophysical properties and transfection efficiency
(tłumaczenie polskie: Rozgałęzione amfifilowe polipeptydy kationowe tworzące kompleksy z

29
cząsteczkami DNA: studia ich właściwości biofizycznych i wydajności transfekcji), Molecular
Pharmaceutics 12, 706-715 (2015).
Impact Factor: 4.787 (2013), 5.603 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 45
Cytowania: Web of Science: 0 Google Scholar: 0
Wkład pracy dr Roberta Szoszkiewicza: 5%; Zebrałem pierwsze dane AFM i pomogłem w
interpretacji wyników pomiarowych.
W trakcie moich badań w dziedzinie fizyki materiałowej na KSU skoncentrowałem się
głównie na zrozumieniu procesów powstawania zmarszczek powierzchniowych na
wybranych powierzchniach polimerów i kopolimerów. Badania te dostarczyły zrozumienia
powiązań między właściwościami wiskoelastycznymi, a procesami erozji i mikro-transportu
materiału polimerowego na powierzchni polimerów wskutek lokalnego grzania tychże
powierzchni. Rezultaty tych badań zostały przedstwione na wielu wystąpieniach naukowych i
opisane w pięciu zrecenzowanych publikacjach:
R. Szoszkiewicz and E. Riedo, Sliding charges (tłumaczenie polskie: Przesuwające się ładunki),
Nature Materials (News and Views), 13, 666-668 (2014).
Impact Factor: 36.425 (2013), 41.775 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 50
Cytowania: Web of Science: 0 Google Scholar: 0
Wkład pracy dr Roberta Szoszkiewicza: 80%; Dostaczyłem pomysłu i napisałem na tą publikację.
R. H. Rice, E. Gnecco, W. P. King, and R. Szoszkiewicz, Heterogeneity of spiral wear patterns
produced by local heating on amorphous polymers (tłumaczenie polskie: Heterogeniczność
spiralnych struktrur topograficznych wytworzonych na powierzchniach polimerów amorficznych
wskutek lokalnego grzania), Materials Chemistry and Physics 141(1), 477-481 (2013).
Impact Factor: 2.129 (2013), 2.427 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 0 Google Scholar: 0
Wkład pracy dr Roberta Szoszkiewicza: 50%; Wymyśliłem ten kierunek badawczy, dostarczyłem
kierunków analizy danych, i napisałem tą publikację.
R. H. Rice, E. Gnecco, R. Wannemacher, and R. Szoszkiewicz, Velocity dependence of nano-
abrasive wear obtained using a spiral scan pattern (tłumaczenie polskie: Zależność procesu
wyżłabiania spiralnych struktur w wartswach polimerowych od szybkości tego procesu), Polymer
54(14), 3620-3623 (2013).
Impact Factor: 3.766 (2013), 4.224 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 40
Cytowania: Web of Science: 0 Google Scholar: 0
Wkład pracy dr Roberta Szoszkiewicza: 50%; Wymyśliłem ten kierunek badawczy, dostarczyłem
kierunków analizy danych, i napisałem tą publikację.
R. H. Rice, P. Mokarian-Tabari, W. P. King, and R. Szoszkiewicz, Local thermo-mechanical
analysis of a microphase separated thin lamellar PS-b-PEO film (tłumaczenie polskie: lokalna
termo-mechaniczna analiza mikrofazowo rozseperowanych cienkich filmów kopolimerowych z
PS-b-PEO), Langmuir 28, 13503 - 13511 (2012).

30
Impact Factor: 4.187 (2012), 4.489 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 6 Google Scholar: 8
Wkład pracy dr Roberta Szoszkiewicza: 60%; Wymyśliłem ten kierunek badawczy, dostarczyłem
kierunków analizy danych, dostarczyłem modelu analitycznego do opisu otrzymanych danych i
napisałem tą publikację.
E. Gnecco, E. Riedo, W. P. King, S. R. Marder, R. Szoszkiewicz, Linear ripples and traveling
circular ripples produced on polymers by thermal AFM probes (tłumaczenie polskie: zmarszczki
liniowe i podróżące zmarszczki kołowe wytworzone na polimerach poprzez belki termiczne AFM),
Physical Review B, 79, 235421 (2009)
Impact Factor: 3.475 (2009), 3.564 (5 year)
Punkty według Listy Ministerialnej z dnia 31 grudnia 2014: 35
Cytowania: Web of Science: 11 Google Scholar: 20
Wkład pracy dr Roberta Szoszkiewicza: 45%; I designed this research, provided directions for
analysis of the experimental data, and I wrote this paper.
W roku 2015 opuściłem Uniwersytet Stanowy w Kansas i powróciłem do Polski, gdzie od
kwietnia 2015 zaakceptowałem pozycję „adiunkta naukowego” na Wydziale Inżynierii
Materiałowej Politechniki Warszawskiej. Na Politechnice planuję kontynuować swój rozwój
naukowy w dziedzinach biofizyki i inżynierii materiałowej.
Sumaryczny impact factor publikacji naukowych według listy Journal Citation
Reports (JCR), zgodnie z rokiem opublikowania:
IF = 145,678
Suma cytacji według bazy Web of Science z dnia 02.06.2015:
675 (619 bez samocytowań)
Indeks Hirscha według bazy danych Web of Science: 12
Indeks Hirscha według bazy danych Google Scholar: 14
Kierowanie projektami badawczymi:
Fundusz na start pracowni Szosz-lab na Wydziale Fizyki Uniwersytetu Stanowego w
Kansas (ang. Start-Up Funds)
R. Szoszkiewicz, Kierownik Projektu
Okres: 15/07/08 - 31/12/11

31
Sponsor: Kansas State University, Manhattan, Kansas, USA
Innowacyjny grant-nagroda w badaniach podstawowych nad rakiem w edycji jesień 2008
R. Szoszkiewicz, Kierownik Projektu
Okres: 01/04/09 –01/04/12
Sponsor: Centrum Badań Podstawowych na Rakiem w Stanie Kansas, im. Terrego C.
Johnsona, Stany Zjednoczone Ameryki Północnej (ang. T. Johnson Center for Basic Cancer
Research, Kansas, USA.)
Tytuł: Badania procesu aktywacji białka Notch w skali pojedynczej cząsteczki.
Tzw. First Award przyznawana przez Krajową Fundację Nauki Amerykańskiej (ang.
National Science Foundation, NSF)
R. Szoszkiewicz, Kierownik Projektu
Okres: 01/05/11 - 31/07/12
Tytuł: Nanomechaniczne studia nad formowaniem się struktury drugorzędowej z użyciem
mikroskopu sił atomowych i intereferometru (ang. Nanomechanical Studies of the Secondary
Structure Folding using AFM and interferometry).
Innowacyjny grant-nagroda w badaniach podstawowych nad rakiem w edycji jesień 2013
R. Szoszkiewicz, Kierownik Projektu
Okres: 01/04/14 – 30/03/16
Sponsor: Centrum Badań Podstawowych na Rakiem w Stanie Kansas, im. Terrego C.
Johnsona, Stany Zjednoczone Ameryki Północnej (ang. T. Johnson Center for Basic Cancer
Research, Kansas, USA.)
Tytuł: W stronę szybkiej i ilościowej detekcji biomarkerów rakowych.
NSF, Dyrektorat Inżynierii (ang. Directorate for Engineering), Jednostka Inżynierii
Cywilnej, Mechanicznej i Innowacyjnych Procesów Wytwarzania (ang. Civil,
Mechanical and Manufacturing Innovation, CMMI, Division), grant number #1434787
Grant ten został otrzymany w konsorcjum między Uniwersytetem Stanu Illinois w Urbana-
Champain (ang. University of Illinois in Urbana-Champain), Instytutem Technologii w
Atlancie w Stanie Georgia (ang. Georgia Institute of Technology) i KSU. Robert
Szoszkiewicz był kierownikiem projektu w KSU.
Okres: 01/09/14 – 30/08/17. Zakończyłem swoją część w styczniu 2015, ze względu na
powrót do Polski.
Tytuł: Badania we współpracy: Kontrola Chemii w Nanoskali: Paralelizacja, Powtarzalność i
Dokładność (ang. Collaborative Research: Controlling the chemistry at the nanoscale:
Parallelization, Robustness, and Registration).
Parę grantów na zakup sprzętu badawczego finansowanych przez Centrum Badań
Podstawowych na Rakiem w Stanie Kansas, im. Terrego C. Johnsona, Stany Zjednoczone
Ameryki Północnej. Okres: lata 2009 – 2013.

32
Nagrody:
Stypendium Ministra Edukacji Narodowej 10/1997 - 7/1998
Stypendium – nagroda na Uniwersytecie Boston College Bostonie, USA 1/1998 - 6/1998
Stypendium Ministra Edukacji Narodowej 10/1996 - 7/1997
Prezentacje na krajowych i zagranicznych konferencjach naukowych:
Jedna zaproszona prezentacja konferencyjna: 1. VI Seminarium AFM/STM: Badania prowadzone metodami skaningowej mikroskopii bliskich
oddziaływań, Zakopane, Poland, "Pulling power of proteins: force activated reactivity switch at the
single bond level and mechanical properties of human Notch 1" (tłumaczenie polskie: “Aktywowany
siłą sposób zmiany reaktywaności pojedynczego wiązania dwusiarczkowego a także właściwości
mechaniczne białka ludzkiego Notch1”) (12/2010).
Dwadzieścia jeden (21) ustnych prezentacji konferencyjnych: 1. Annual APS Prairie Section Meeting, Lawrence, KS, USA (tłumaczenie polskie: Coroczne
Spotkanie Sekcji Prerii Amerykańskiego Towarzystwa Fizycznego, Lawrence, Stan Kansas, Stany
Zjednoczone Ameryki Północnej), “Protein folding intermediates probed by ensemble of their
transient stiffnesses in single-molecule force-quenched AFM“ (tłumaczenie polskie: Struktury
przejściowe w trakcie procesu fałdowania się białek próbkowane przez zmiany sztywności w
eksperymentów typu force-quench AFM”), 11/2012.
2. “Mechanics in Biology” workshop at UCLA (tłumaczenie polskie: Warsztaty „Mechanika w
Biologii” zorganizowane w Kalifornijskim Uniwersytecie Stanowym w Los Angeles), “Mechanical
unfolding of NRR domain from human Notch 1“(tłumaczenie polskie: Mechaniczne rozfałdowywanie
się domeny NRR z ludziego białka Notch1), 1/2012.
3. Gordon Research Conference on protein folding in Ventura, CA, USA (tłumaczenie polskie:
Konferencja na temat zwijania się białek zorganizowana przez Towarzystwo Gordon Research w
miejscowości Ventura, w Stanie Kalifornia, USA) “Mechanical unfolding of NRR domain from human
Notch 1“(tłumaczenie polskie: Mechaniczne rozfałdowywanie się domeny NRR z ludziego białka
Notch1), 1/2012.
4. XIII Annual Linz Winter Workshop on Advances in Single-Molecule Research for Biology &
Nanoscience, Linz, Austria, (tłumaczenie polskie: XIII Coroczne Warsztaty Zimowe w Linz na temat
Postępów w badaniach pojedynczych cząsteczek w Biologii i Nanonaukach) “Mechanical properties
of NRR domain from human Notch 1 studied by single molecule AFM force spectroscopy”
(tłumaczenie polskie: Własności mechaniczne domeny NRR z ludziego białka Notch1 badane za
pomocą spektroskopii sił AFM na poziomie pojedynczej cząsteczki), 2/2011.
5. APS March Meeting, Dallas, TX, (tłumaczenie polskie: Spotkanie Marcowe Amerykańskiego
Towarzystwa Fizycznego, Dallas, Stan Teksas, Stany Zjednoczone Ameryki Północnej) “Mechanical
properties of NRR domain from human Notch 1 studied by single molecule AFM force spectroscopy”
(tłumaczenie polskie: Własności mechaniczne domeny NRR z ludziego białka Notch1 badane za
pomocą spektroskopii sił AFM na poziomie pojedynczej cząsteczki), 3/2011.
8. “Nanomeasure 2010” – konferencja zorganizowana przez firmę Agilent, Stany Zjednoczone
Ameryki Północnej, i Uniwersytet Jagielloński, Kraków, “The force-activated reactivity switch in a
bimolecular chemical reaction at the single-molecule level” (tłumaczenie polskie: Zmiana reaktywności w dwucząsteczkowej reakcji chemicznej aktywowana siłą mechaniczną), 5/2010.

33
9. Annual Meeting of Four Corners Section of APS, Ogden, UT, (tłumaczenie polskie: Coroczne
Spotkanie Sekcji Four Corners Amerykańskiego Towarzystwa Fizycznego, Ogden, Stan Utah, Stany
Zjednoczone Ameryki Północnej) "Mechanical properties of NRR domain from human Notch 1
studied by single molecule AFM force spectroscopy and steered molecular dynamics" (tłumaczenie
polskie: Własności mechaniczne domeny NRR z ludziego białka Notch1 badane za pomocą spektroskopii sił AFM na poziomie pojedynczej cząsteczki), 10/2010.
10. Annual Meeting of Four Corners Section of APS, Colorado School of Mines, Golden, CO,
(tłumaczenie polskie: Coroczne Spotkanie Sekcji Four Corners Amerykańskiego Towarzystwa
Fizycznego, Golden, Stan Kolorado, Stany Zjednoczone Ameryki Północnej) “Force-activated
reactivity switch in a bimolecular chemical reaction at the single molecule level” (tłumaczenie
polskie: Zmiana reaktywności w dwucząsteczkowej reakcji chemicznej aktywowana siłą mechaniczną), 10/2009.
11. Kansas Physical Chemistry Symposium, University of Kansas, Lawrence, KS, (tłumaczenie
polskie: Sympozjium Chemii Fizycznej w Kansas, Lawrence, Stany Zjednoczone Ameryki Północnej)
“Force-activated reactivity switch in a bimolecular chemical reaction at the single molecule level”
(tłumaczenie polskie: Zmiana reaktywności w dwucząsteczkowej reakcji chemicznej aktywowana siłą
mechaniczną), 2009.
12. APS March Meeting, Baltimore, MD, (tłumaczenie polskie: Spotkanie Marcowe Amerykańskiego
Towarzystwa Fizycznego, Baltimore, Stany Zjednoczone Ameryki Północnej) “Polymer dynamics at
local scales: origin of ripples formation” (tłumaczenie polskie: Dynamika polimerów w skalach
lokalnych: geneza formowania się zmarszczek na powierzchniach polimerów), 2006.
13. APS March Meeting, Baltimore, MD, (tłumaczenie polskie: Spotkanie Marcowe Amerykańskiego
Towarzystwa Fizycznego, Baltimore, Stany Zjednoczone Ameryki Północnej) “Formation of
nanoscale water bridges” (tłumaczenie polskie: Formowanie się mostków kapilarnych w nanoskali),
2006.
14. MRS Fall Meeting, Boston, MA, USA (tłumaczenie polskie: Spotkanie Jesienne Amerykańskiego
Towarzystwa Badań Materiałów, Boston, Stany Zjednoczone Ameryki Północnej) “Friction force
microscopy as a tool to probe local phase transitions” (tłumaczenie polskie: Mikroskopia pomiaru sił
tarcia jako narzędzie do badania lokalnych przejść fazowych), 2005.
15. 23rd European Conference on Surface Science, Berlin, Republika Federalna Niemiec, (tłumaczenie
polskie: 23 Konferencja Europejska z Nauk o Powierzchni) “Friction force microscopy as a tool to
probe local phase transitions” (tłumaczenie polskie: Mikroskopia pomiaru sił tarcia jako narzędzie do
badania lokalnych przejść fazowych), 2005.
16. International Non-contact Atomic Force Microscopy Conference, Seattle, WA, USA (tłumaczenie
polskie: Międzynarodowa Konferencja z Niekontaktowej Mikroskopii Sił Atomowych, Seattle, Stan
Washington, Stany Zjednoczone Ameryki Północnej) “Adhesion hysteresis and its correlations with
friction” (tłumaczenie polskie: Histereza adhezji i jej związki z tarciem), 2004.
17. 27th International Symposium on Acoustical Imaging, Saarbrucken, Germany, (tłumaczenie
polskie: 27 Międzynarodowe Sympozjum z Obrazowania Ultradźwiękami, Saaebrucken, Republika
Federalna Niemiec) “How can ultrasounds help with connecting friction and adhesion at local scales”
(tłumaczenie polskie: Jak ultradźwięki mogą pomóc w badaniu związków między tarciem a histerezą
adhezji w skalach lokalnych), 2003.
18. Swiss Physical Society Annual Meeting, Basel, Switzerland, (tłumaczenie polskie: Coroczne
Spotakanie Szwajcarskiego Towarzystwa Fizycznego, Bazylea, Szwajcaria) “How can ultrasounds
help with connecting friction and adhesion at local scales” (tłumaczenie polskie: Jak ultradźwięki
mogą pomóc w badaniu związków między tarciem a histerezą adhezji w skalach lokalnych), 2003.
19. 3rd International Symposium on Tribochemistry, Cracow, Poland, (tłumaczenie polskie: 3e
Międzynarodowe Sympozjum z Trybochemii) “Adhesion hysteresis and ultrasonic force microscopy”
(tłumaczenie polskie: Histereza adhezji i ultradźwiękowa mikroskopia sił), 2001.

34
20. Mini-Symposium „Son et Lumiere”, Grasmere, Wielka Brytania, (tłumaczenie polskie: Mini-
sympozjum o Dźwięku i Świetle) “Jumping off, jumping on - principles of ultrasonic force
microscopy” (tłumaczenie polskie: Podskakiwanie w górę i w dół – koncepty ultradźwiękowej
mikroskopii sił), 2000.
21. 12th Międzynarodowa Konferencja Studentów Fizyki, Wiedeń, Austria, “Viscosity of discotic
liquid crystals” (tłumaczenie polskie: lepkość ciekłych kryształów o strukturach dyskotycznych), 1997.
Osiemnaście (18) Zaproszonych Seminariów i Wykładów: 1. SEMINARIUM Z FIZYKI BIOLOGICZNEJ I BIOINFORMATYKI, Instytutu Biochemii i
Biofizyki PAN, Instytutu Fizyki PAN, i Zakladu Biofizyki UW, IBB, Warszawa, Stiffness
and internal friction of single protein molecules: AFM studies (tłumaczenie polskie:
Sztywność i tarcie wewnętrzne pojedynczych cząsteczek białek: studia AFM), 3/2015
2. Seminarium Fizyki Miękkich Faz, Instytut Chemii Fizycznej PAN, Exerting mechanical force
on single proteins and polymer surfaces (tłumaczenie polskie: Wywieranie siły mechanicznej
na pojedyncze cząsteczki białek i substancje polimerowe), 12/2014.
3. Imperial College London, Wielka Brytania, Departament Biomateriałów, Using single
molecule AFM methods to learn about proteins (tłumaczenie polskie: Używanie metod AFM
w badaniach pojedynczych cząsteczek w celu zdobycia wiedzy o białkach), 11/2013.
4. Wydział Fizyki, Matematyki i Informatyki, Politechnika Krakowska, Using single molecule
AFM methods to find and characterize folding intermediates of a simple model protein
(tłumaczenie polskie: Używanie metod AFM w celu znalezienia i charakteryzacji struktur
pośrednich w trakcie zwijania się prostych cząsteczek białek), 11/2013.
5. Centrum NANOSAM, Uniwersytet Jagielloński, Kraków, Using single molecule AFM
methods to find and characterize folding intermediates of a simple model protein
(tłumaczenie polskie: Używanie metod AFM w celu znalezienia i charakteryzacji struktur
pośrednich w trakcie zwijania się prostych cząsteczek białek), 11/2013.
6. Międzywydziałowe Seminarium z Fizyki/Chemii/Biochemii, Georgia Tech, Stany
Zjednoczone Ameryki Północnej, Exerting force on biomolecules (tłumaczenie polskie:
Wywieranie siły mechanicznej na biomolekuły), 8/2013.
7. Wydział Fizyki, Laboratorium Narodowe Argonne, Stan Illinois, Stany Zjednoczone Ameryki
Północnej, Exerting force on proteins (tłumaczenie polskie: Wywieranie siły mechanicznej na
białka), 3/2013.
8. Wydział Fizyki, Uniwersytet Purdue, Stan Indiana, Stany Zjednoczone Ameryki Północnej,
Exerting force on proteins (tłumaczenie polskie: Wywieranie siły mechanicznej na białka),
3/2013.
9. Exerting Mechanical Force on Single Proteins (tłumaczenie polskie: Wywieranie siły
mechanicznej na pojedyncze cząsteczki białka), Wydział Nauk o Życiu, Ecole Polytechnique
Federale de Lausanne, Switzerland, 3/2012.
10. Chemical nad topographical nano-patterning of polymers using thermochemical
nanolithography (tłumaczenie polskie: Chemiczne i topograficzne nanostrukturyzowanie
polimerów z użyciem termochemicznej nanolitografii), Centrum NANOSAM, Uniwersytet
Jagielloński, Kraków, 3/2012.
11. Centrum NANOSAM, Uniwersytet Jagielloński, Kraków, Some applications and basic
research in bio-nano-technology: from pulling single protein molecules to thermochemical
nanolithography” (tłumaczenie polskie: Aplikacje i badania podstawowe w bio-nano-
technologii: od rozwijania pojedynczych cząsteczek białek do nanolitografii
termochemicznej), 2010.
12. Wykład gościnny dla studentów Studiów Matematyczno-Przyrodniczych, Uniwersytet
Jagielloński, Kraków, "Pomiędzy fizyką, chemią i biologią: jak siła mechaniczna może być
aplikowana do pojedynczych cząsteczek i wiązań chemicznych", 2010
13. Wykład gościnny dla przedmiotu “Nowe Materiały w Nanotechnologii”, Uniwersytet
Jagielloński, Kraków, 2010.
14. Wydział Mechaniczny Politechniki Radomskiej,“Some applications and basic research in
bio-nano-technology: from pulling single protein molecules to thermochemical

35
nanolithography” (tłumaczenie polskie: Aplikacje i badania podstawowe w bio-nano-
technologii: od rozwijania pojedynczych cząsteczek białek do nanolitografii
termochemicznej), 2010.
15. Międzynarodowy Instytut Biologii Molekularnej i Komórkowej, Warszawa,“Some
applications and basic research in bio-nano-technology: from pulling single protein
molecules to thermochemical nanolithography” (tłumaczenie polskie: Aplikacje i badania
podstawowe w bio-nano-technologii: od rozwijania pojedynczych cząsteczek białek do
nanolitografii termochemicznej), 2010.
16. Wydział Chemii, Uniwersytet Łódzki, “Some applications and basic research in bio-nano-
technology: from pulling single protein molecules to thermochemical nanolithography”
(tłumaczenie polskie: Aplikacje i badania podstawowe w bio-nano-technologii: od rozwijania
pojedynczych cząsteczek białek do nanolitografii termochemicznej), 2010.
17. Narodowy Instytut Standartów i Technologii (NIST), Gaithersburg, Stan Maryland, Stany
Zjednoczone Ameryki Północnej, “Local capillaries of water and water confined at sub-
nanometer gaps: a prelude to bioinspired surfaces“ (tłumaczenie polskie: Lokalne mostki
wodne i woda związana w przestrzeniach sub-nanometrowych), 2007.
18. Fraunhofer Society, IZFP, i Uniwersytet w Saarbrucken, Republika Federalna Niemiec,
“Correlations between adhesion hysteresis and friction at local scales” (tłumaczenie polskie:
Korelacje między histerezą adhezji a tarciem w skalach lokalnych), 2003.
Dwanaście (12) wykładów w formie kolokwiów:
1. Wydział Biochemii KSU, Exerting force on biomolecules (tłumaczenie polskie: Wywieranie
siły mechanicznej na biomolekuły), 11/2013
2. Wydział Fizyki KSU, Biological and Soft-Matter Physics in the Szosz-lab (tłumaczenie
polskie: Fizyka biologiczna i fizyka miękkich faz w pracowni Szosz-lab), 9/2013.
3. Wydział Fizyki Uniwersytetu Stanowego w Wichita, Kansas, Exerting force on biomolecules,
(tłumaczenie polskie: Wywieranie siły mechanicznej na biomolekuły), 5/2013.
4. Wydział Chemii Uniwersytetu w Kansas, Exerting force on single proteins (tłumaczenie
polskie: Wywieranie siły mechanicznej na pojedyncze cząsteczki białek), 3/2013.
5. Wydział Fizyki KSU, "Szosz-lab: Molecular Biophysics and Soft-Matter Materials Physics"
(tłumaczenie polskie: Biofizyka cząsteczkowa i fizyka miękkich faz w pracowni Szosz-lab),
1/2011.
6. Wydział Fizyki Uniwersytetu Stanu Teksas w Dallas, “Single-Molecule Studies of
Mechanical (un)Folding of Proteins, Enzymatic Catalysis and Water Structure in Cell
Membranes” (tłumaczenie polskie: Studia na poziomie pojedynczych cząsteczek i
mechanicznego rozciągania i zwijania się białek, katalizy enzymatycznej i struktury wody w
membranach biologicznych), 2008.
7. Wydział Biochemii KSU, “Single-molecule studies of mechanical (un)folding of proteins and
enzymatic catalysis” (tłumaczenie polskie: Studia na poziomie pojedynczych cząsteczek i
mechanicznego rozciągania i zwijania się białek, a także katalizy enzymatycznej) , 2008.
8. Wydział Fizyki KSU, “Single-molecule and local studies of mechanical (un)folding of
proteins, enzymatic catalysis and water structure at solid surfaces” (tłumaczenie polskie:
Studia na poziomie pojedynczych cząsteczek i mechanicznego rozciągania i zwijania się
białek, katalizy enzymatycznej i struktury wody na powierzchniach ciał stałych), 2008.
9. Wydział Fizyki Uniwersytetu Stanowego w Północnej Karolinie, USA “Single-molecule and
local studies of mechanical (un)folding of proteins, enzymatic catalysis and water structure at
solid surfaces” (tłumaczenie polskie: Studia na poziomie pojedynczych cząsteczek i
mechanicznego rozciągania i zwijania się białek, katalizy enzymatycznej i struktury wody na
powierzchniach ciał stałych), 2008.
10. Wydział Fizyki Uniwersytetu Stanowego na Florydzie, USA “Single-molecule and local
studies of mechanical (un)folding of proteins, enzymatic catalysis and water structure at solid
surfaces” (tłumaczenie polskie: Studia na poziomie pojedynczych cząsteczek i mechanicznego

36
rozciągania i zwijania się białek, katalizy enzymatycznej i struktury wody na powierzchniach
ciał stałych), 2008.
11. Wydział Fizyki Uniwersytetu Stanowego “Wayne State”, Stan Michigan, USA, “Local
capillaries of water and water confined at sub-nanometer gaps: a prelude to bioinspired
surfaces” (tłumaczenie polskie: Lokalne mostki wodne i woda związana w przestrzeniach sub-
nanometrowych: preludium do bioinspirowanych powierzchni), 2007.
12. Narodowe Laboratorium w Brookhaven w Stanie Nowy Jork, USA, “Water at local scales”
(tłumaczenie polskie: Woda w skalach lokalnych), 2006.
Trzynaście (13) wykładów w formie seminariów wydziałowych: 1. Zakład Projektowania Materiałów Wydziału Inżynierii Materiałowej Politechniki
Warszawskiej, Thermochemical nanolithography on polymers (tłumaczenie polskie:
termochemiczna nanoligrafia na polimerach), 3/2015.
2. Biogrupa Wydziału Inżynierii Materiałowej Politechniki Warszawskiej, Stiffness and internal
friction of single protein molecules: AFM studies (tłumaczenie polskie: Sztywność i tarcie
wewnętrzne pojedynczych cząsteczek białek: studia AFM), 4/2015.
3. Wydział Fizyki KSU, Thermochemical nanolithography on polymers (tłumaczenie polskie:
termochemiczna nanoligrafia na polimerach), 9/2013.
4. Wydział Fizyki KSU, “Nano-abrasion of polymers” (tłumaczenie polskie: nanoabrazja
polimerów), 9/2012.
5. Wydział Fizyki KSU, “Nanomechanics of Proteins at the Single Molecule Level”
(tłumaczenie polskie: nanomechanika pojedynczych cząsteczek białek), 9/2011.
6. Wydział Inżynierii Chemicznej KSU, "The Szosz-lab at KSU: Investigating the effects of
mechanical force on single chemical bonds and proteins, as well as some results with
thermochemical nanolithography on polymers" (tłumaczenie polskie: Studia nad efektami siły
mechanicznej na pojedyncze wiązania chemiczne i pojedyczne cząsteczki białka, a także
rezultaty badań z użyciem termochemicznej nanolitografii na polimerach w pracowni Szosz-
lab w KSU), 2010.
7. Wydział Fizyki KSU, "2D assemblies of nanoparticles by thermochemical nanolithography"
(tłumaczenie polskie: Dwuwymiarowe struktury z nanocząsteczek utworzone za pomocą
termochemicznej nanolitografii ciepłem), 2010.
8. Wydział Fizyki KSU, "Current research overview: stretching proteins and patterning
polymers" (tłumaczenie polskie: Przegląd bieżących badań: rozciąganie białek i
nanostrukturyracja polimerów), 2009.
9. Wydział Chemii KSU, “The pulling power of proteins: investigating single-molecule disulfide
reductions one by one” (tłumaczenie polskie: Rozciąganie białek: badania procesów redukcji
pojedynczych wiązań dwusiarczkowych), 2009.
10. Wydział Fizyki KSU, "Confined water, water capillaries and control of hydrophilicity at
nanoscopic lengths" (tłumaczenie polskie: Zaadsorbowane woda, mostki kapilarne i konrola
hydrofilowości w skalach nanoskopowych), 2008.
11. Wydział Chemii KSU, “Confined water, water capillaries and control of hydrophilicity at
nanoscopic lengths” (tłumaczenie polskie: Zaadsorbowane woda, mostki kapilarne i konrola
hydrofilowości w skalach nanoskopowych), 2008.
12. Wydział Fizyki Georgia Tech-u, Stany Zjednoczone Ameryki Północnej “Water at local
scales” (tłumaczenie polskie: Woda w skalach lokalnych), 2005.
13. Wydział Fizyki Georgia Tech-u, Stany Zjednoczone Ameryki Północnej “Nanoscale
capillary bridges investigated by friction force microscopy” (tłumaczenie polskie:
Nanoskopowe mostki kapilarne badane za pomocą mikroskopii pomiaru tarcia w nanoskali),
2005.

37
Zajęcia Dydaktyczne i Opieka nad Studentami
Podczas mojej 6.5-letniej pracy na KSU opracowałem własne materiały dydaktyczne do
wielu zajęć ze studentami, które prowadziłem na poziomie licencjackim. W szczególności
opracowałem zajęcia a także prowadziłem wykłady i ćwiczenia z Mechaniki Klasycznej
(numer katalogowy w KSU: PHYS 522: Mechanics) a także Termodynamiki z Elementami
Fizyki Statystycznej (numer katalogowy w KSU: PHYS 664: Thermal Physics).
W lutym 2015 opracowałem, wygłosiłem i prowadziłem ćwiczenia z zaproszonego wykładu
autorskiego „Single molecule biophysics and nanotechnology” (tłum. polskie: Badania
pojedynczych cząsteczek w biofizyce i nanotechnologii) na Wydziale Inżynierii Materiałowej
Politechniki Warszawskiej. Kurs ten składał się z sześciu godzin wykładów i sześciu godzin
pracowni laboratoryjnej z udziałem mikroskopii sił atomowych i symulacji komputerowych.
Byłem opiekunem naukowym i wypromowałem czterech magistrantów: Nicoleta Ploscariu,
Magisterium z Fizyki, KSU, 12/2014; Reginald Rice, Magisterium z Fizyki, KSU, 12/2012;
Magisterium z Fizyki, 12/2014; Ashim Dey, Magisterium z Fizyki, KSU, 12/2010; Katarzyna
Małek, Magisterium z Fizyki otrzymane na Wydziale Fizyki UJ, 07/2012, gdzie pełniłem rolę
promotora pomocniczego.
Pełniłem rolę opiekuna dwóch naukowców zatrudnionych w pracowni Szosz-lab w KSU w
ramach ich stażów po-doktorskich: Dr. Neelam Khan, KSU, w latach 2009/2010: obecnie
zatrudniona jako Assistant Professor w Georgia Gwinnett College, GA, USA; Dr. Hui Li,
KSU, w roku 2009, obecnie: zatrudniona jako naukowiec na Wydziale Biochemii KSU. W
przypadku dr. Li pełniłem rolę opiekuna pomocniczego wraz z innym opiekunem naukowym,
z Wydziału Biochemii KSU.
Na przestrzeni mojej kariery w KSU opiekowałem się naukowo jedynastoma (11) studentami
studiów licencjackich, którzy pracowali w pracowni Szosz-lab: Brandin Davis, Jeremy
Pogue, Aaron Morgan, Magdalena Wawrzyniuk, Kushan Weerasinghe, Michael H. Cochran,
Anthony Lang, Katarzyna Malek, Jacek Szczerbinski, Heidi Martin, Vera Okuneva.
Działania Synergiczne:
Popularyzcja nauki w społeczeństwie:
W lipcu 2007 zaprojektowałem i przeprowadziłem dwugodzinny tutorial z AFMu dla
uczestników programu Excite workshop. Program ten został stworzony w KSU z myślą o
popularyzacji Nauk Ścisłych i Technicznych dla dziewcząt z liceów. W zajęciach w
pracowni Szosz-lab wzięło udział około 25 uczestniczek.
W paździeniku 2010 zaprojektowałem i przeprowadziłem około półdniowe aktywności
dla ok. 15u dzieci z lokalnego przedszkola. Zajęcia te miały na celu zainteresowanie
dzieci nauką.



















