







Języki
Strony
Prawny


Plan wykładów z genetyki ogólnej
01 Metody genetyki klasycznej
02 Metody analizy DNA
03 Metody analizy genomu
04 Genomy prokariontów
05 Genomy eukariontów
06 Zmienność genomów w populacjach
07 Genomy a cechy ilościowe
08 Transkrypcja i rola RNA w komórce
09 Translacja i proteom komórkowy
10 Replikacja i naprawa DNA
11 Mutacje i rekombinacje DNA
12 Regulacja ekspresji genów
13 Genetyka rozwoju
14 Genetyka nowotworów
15 Wykład uzupełniający

Metody genetyki klasycznej
1. Badania dziedziczenia cech przed Mendlem
2. Mendel: istnieją wydzielone jednostki dziedziczenia (geny)
3. Materiał genetyczny znajduje się w chromosomach
4. Crossing-over czyli rekombinacja cech
5. Mapowanie genetyczne
6. Test komplementacji i analiza szlaków metabolicznych
7. Test dziedziczenia cytoplazmatycznego

1. Badania dziedziczenia cech przed Mendlem
Nawet tak spektakularne
przypadki nieciągłości w
dziedziczeniu fenotypów nie były
interpretowane jako dowód na
nieciągłość informacji genetycznej.
Najczęściej uważano, że cechy się
zlewają a jeśli rozdzielają to tylko
przypadkowo i na krótko

1. Badania dziedziczenia cech przed Mendlem
Darwin nie dostrzegał ilościowych
stosunków wśród fenotypów …
… na przykład klasycznego
3:1
po skrzyżowaniu dwóch
heterozygot dominujących

2. Mendel: istnieją wydzielone jednostki dziedziczenia (geny)
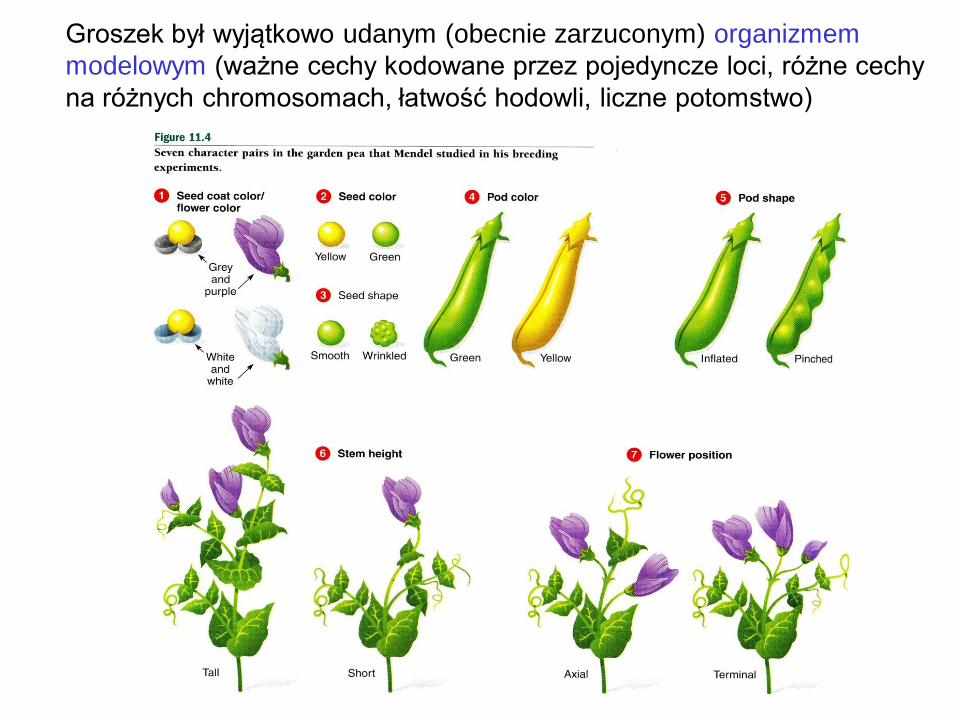
Groszek był wyjątkowo udanym (obecnie zarzuconym) organizmem
modelowym (ważne cechy kodowane przez pojedyncze loci, różne cechy
na różnych chromosomach, łatwość hodowli, liczne potomstwo)
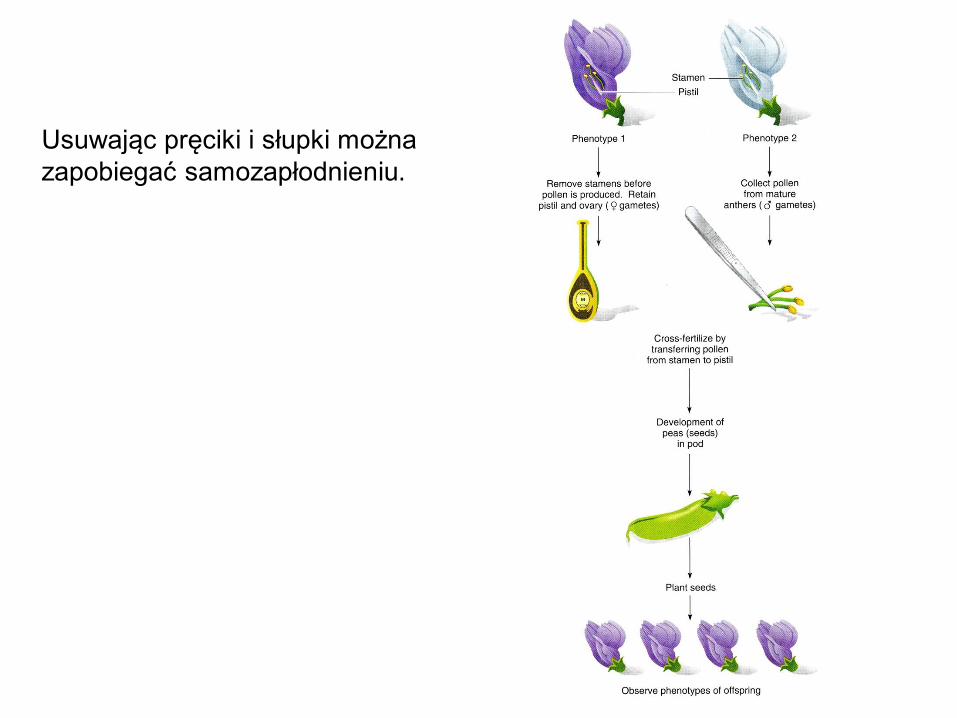
Usuwając pręciki i słupki można
zapobiegać samozapłodnieniu.
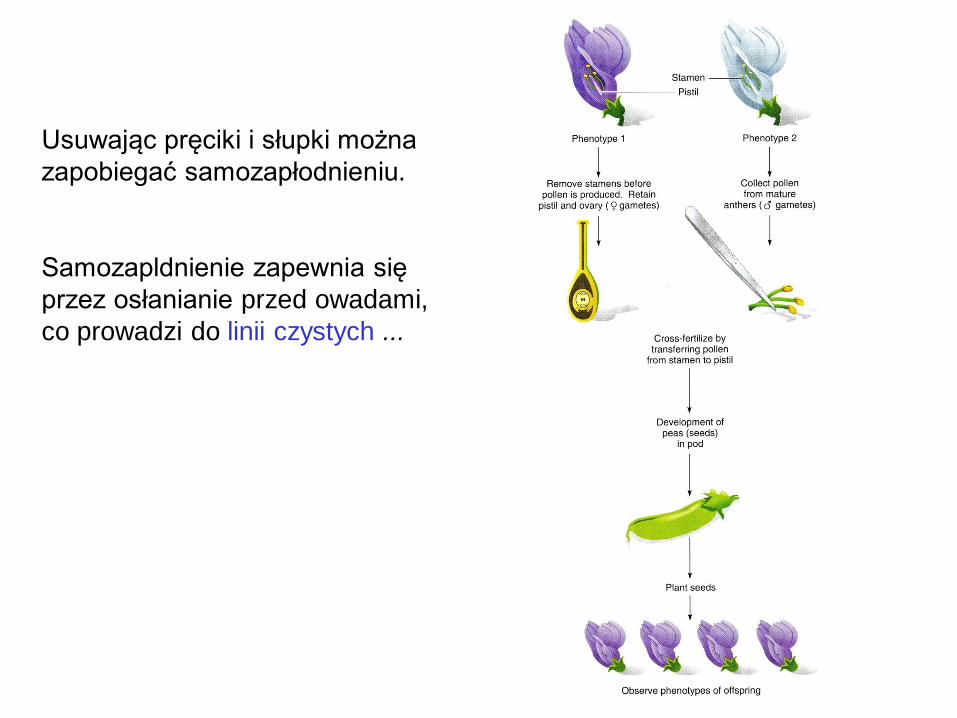
Usuwając pręciki i słupki można
zapobiegać samozapłodnieniu.
Samozapldnienie zapewnia się
przez osłanianie przed owadami,
co prowadzi do linii czystych ...
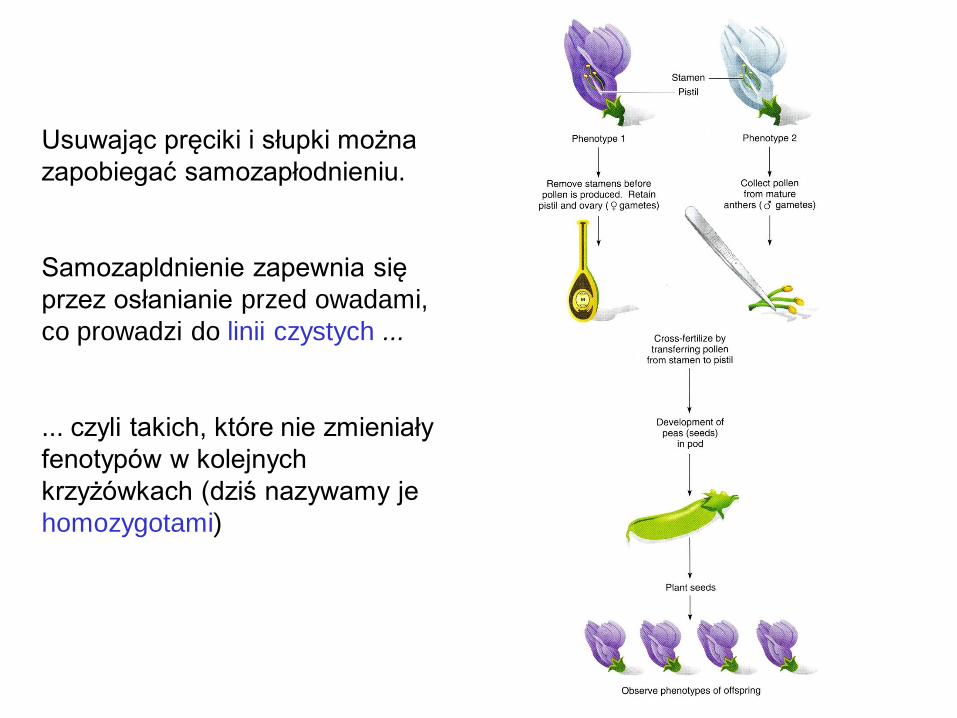
Usuwając pręciki i słupki można
zapobiegać samozapłodnieniu.
Samozapldnienie zapewnia się
przez osłanianie przed owadami,
co prowadzi do linii czystych ...
... czyli takich, które nie zmieniały
fenotypów w kolejnych
krzyżówkach (dziś nazywamy je
homozygotami)
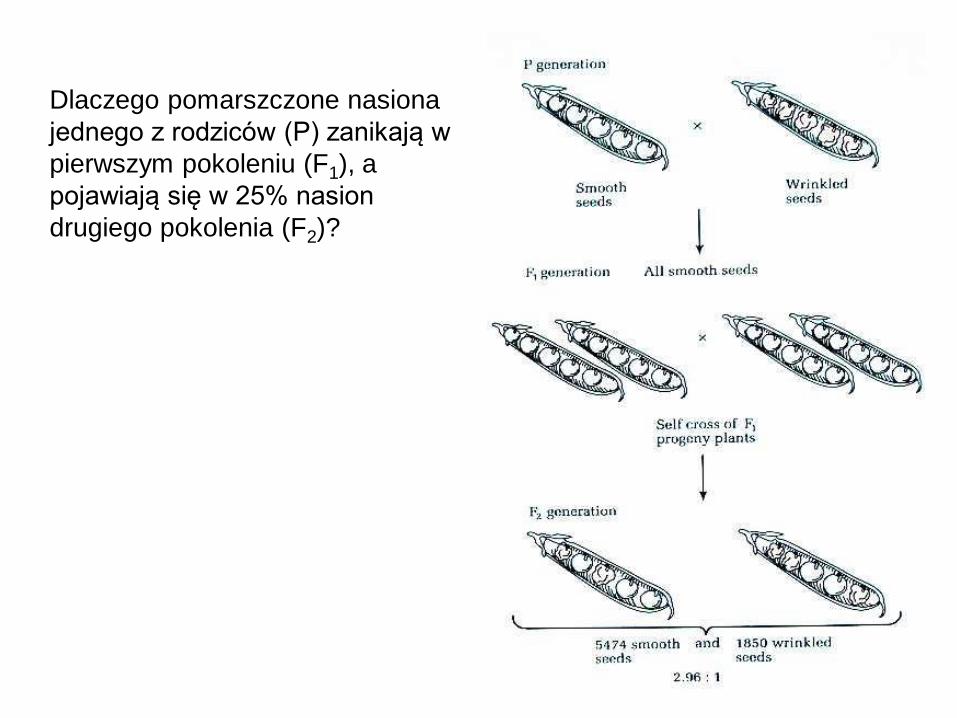
Dlaczego pomarszczone nasiona
jednego z rodziców (P) zanikają w
pierwszym pokoleniu (F1), a
pojawiają się w 25% nasion
drugiego pokolenia (F2)?
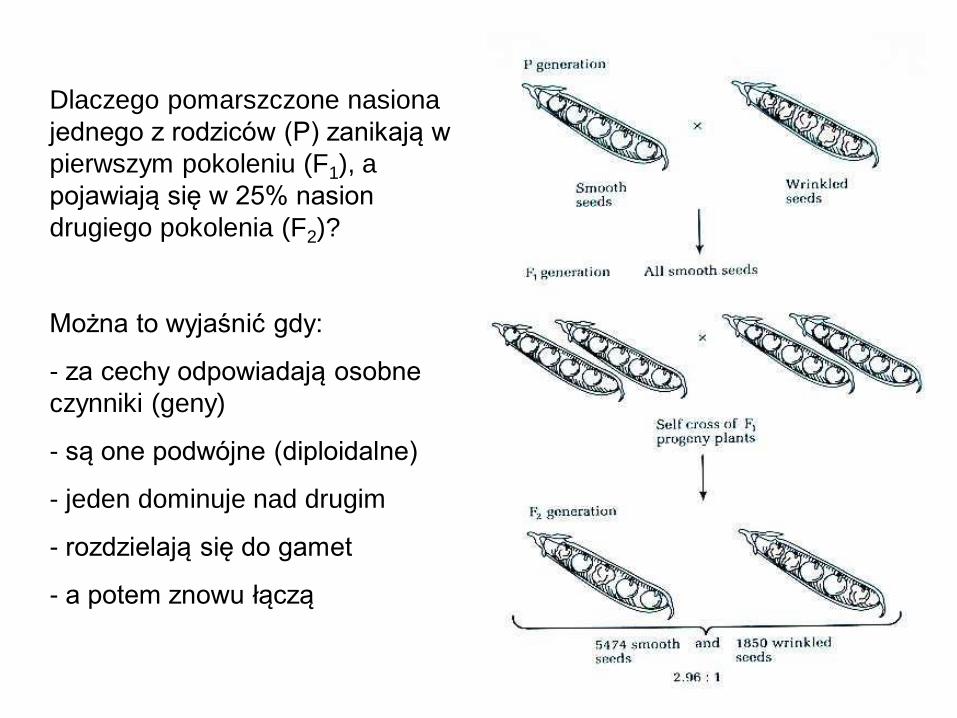
Dlaczego pomarszczone nasiona
jednego z rodziców (P) zanikają w
pierwszym pokoleniu (F1), a
pojawiają się w 25% nasion
drugiego pokolenia (F2)?
Można to wyjaśnić gdy:
- za cechy odpowiadają osobne
czynniki (geny)
- są one podwójne (diploidalne)
- jeden dominuje nad drugim
- rozdzielają się do gamet
- a potem znowu łączą
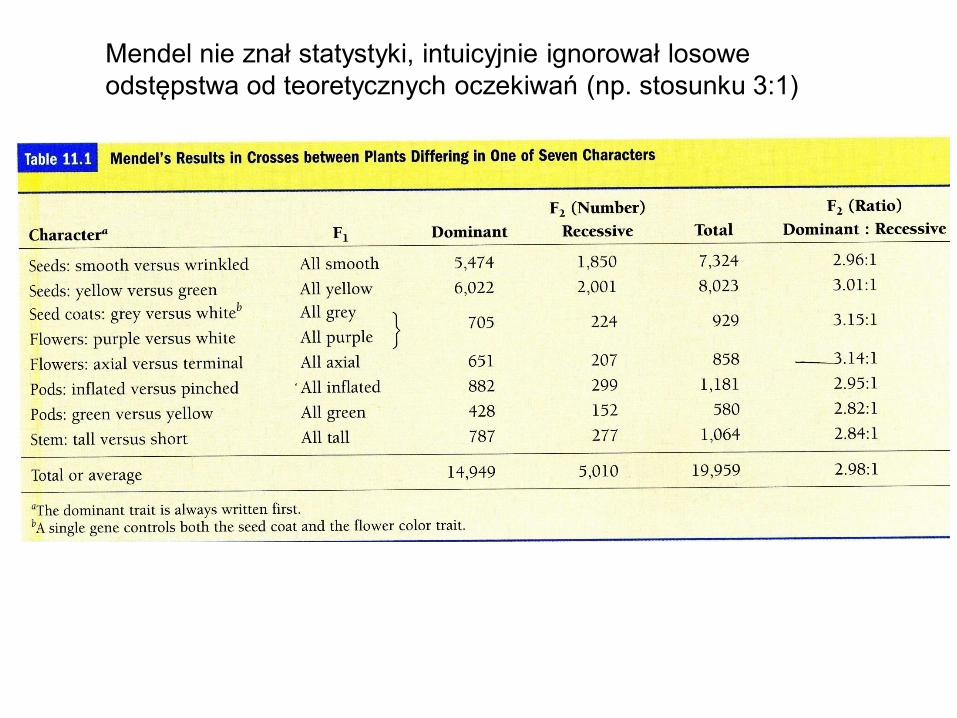
Mendel nie znał statystyki, intuicyjnie ignorował losowe
odstępstwa od teoretycznych oczekiwań (np. stosunku 3:1)
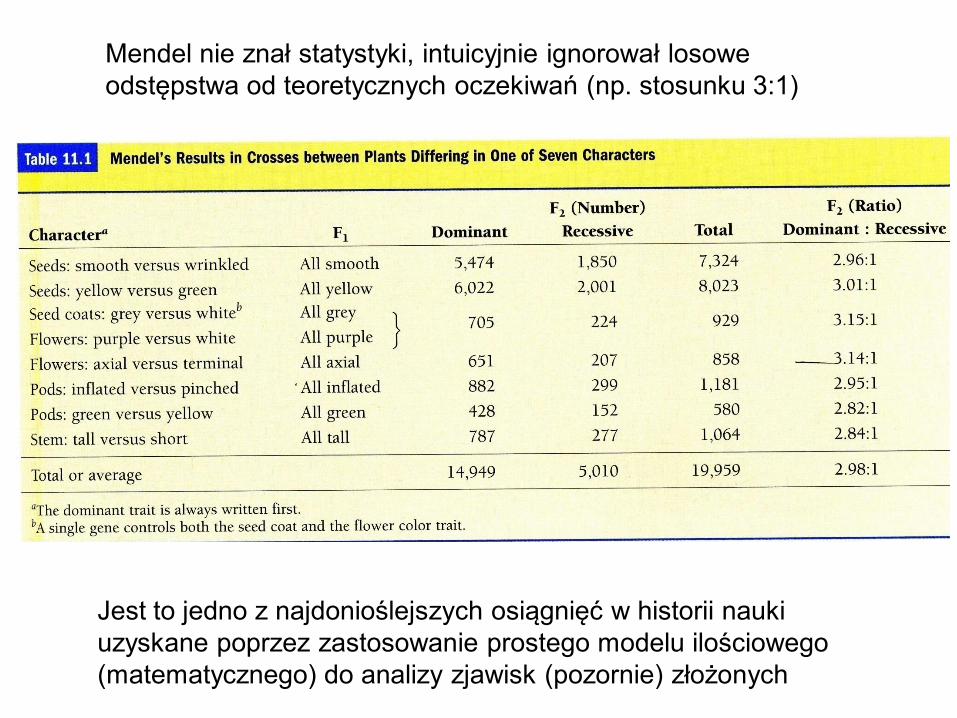
Mendel nie znał statystyki, intuicyjnie ignorował losowe
odstępstwa od teoretycznych oczekiwań (np. stosunku 3:1)
Jest to jedno z najdonioślejszych osiągnięć w historii nauki
uzyskane poprzez zastosowanie prostego modelu ilościowego
(matematycznego) do analizy zjawisk (pozornie) złożonych
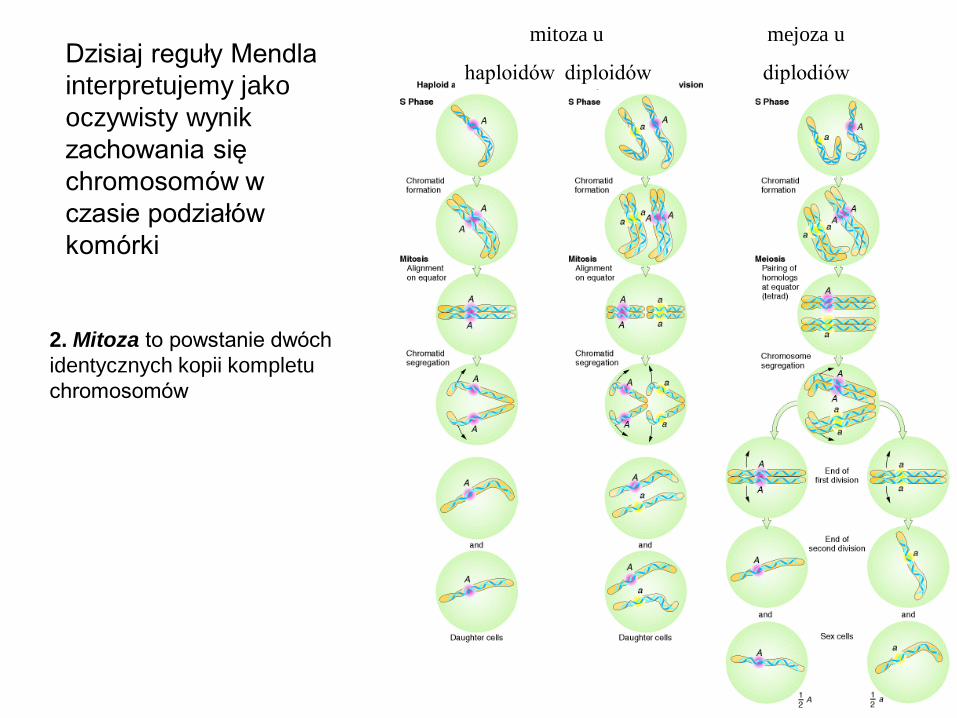
2. Mitoza to powstanie dwóch
identycznych kopii kompletu
chromosomów
mitoza u
haploidów diploidów
mejoza u
diplodiów Dzisiaj reguły Mendla
interpretujemy jako
oczywisty wynik
zachowania się
chromosomów w
czasie podziałów
komórki
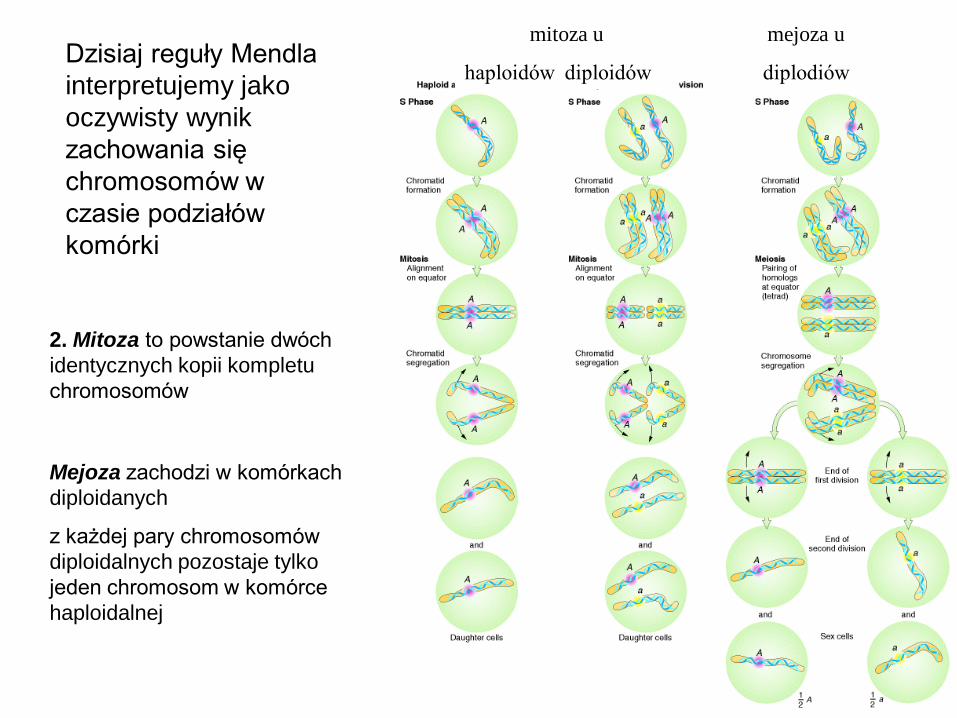
2. Mitoza to powstanie dwóch
identycznych kopii kompletu
chromosomów
Mejoza zachodzi w komórkach
diploidanych
z każdej pary chromosomów
diploidalnych pozostaje tylko
jeden chromosom w komórce
haploidalnej
mitoza u
haploidów diploidów
mejoza u
diplodiów Dzisiaj reguły Mendla
interpretujemy jako
oczywisty wynik
zachowania się
chromosomów w
czasie podziałów
komórki

Wiele par cech nie segregowało niezależnie, tak jak postulował
Mendel w swoim II prawie, wykazywały sprzężenie – było to
zagadnienie badane przez T. H. Morgana.
3. Materiał genetyczny znajduje się w chromosomach

Morgan zasugerował, że cechy
będą sprzężone gdy znajdują
się na jednym chromosomie
Rozwinął technikę krzyżówek
testowych, w której podwójną
heterozygotę Aa Bb krzyżował
z podwójną homozygotą
recesywną aa bb
Jeśli A i B znajdowały się na
różnych chromosomach to
segregowały tak jak
postulowało drugie prawo
Mendla
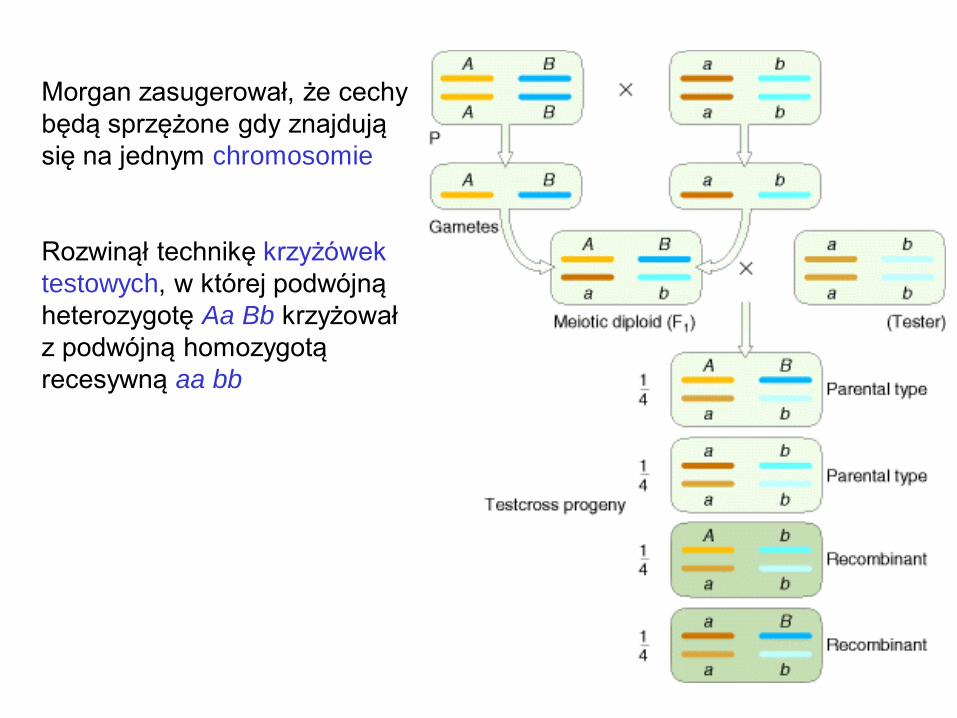
Morgan zasugerował, że cechy
będą sprzężone gdy znajdują
się na jednym chromosomie
Rozwinął technikę krzyżówek
testowych, w której podwójną
heterozygotę Aa Bb krzyżował
z podwójną homozygotą
recesywną aa bb
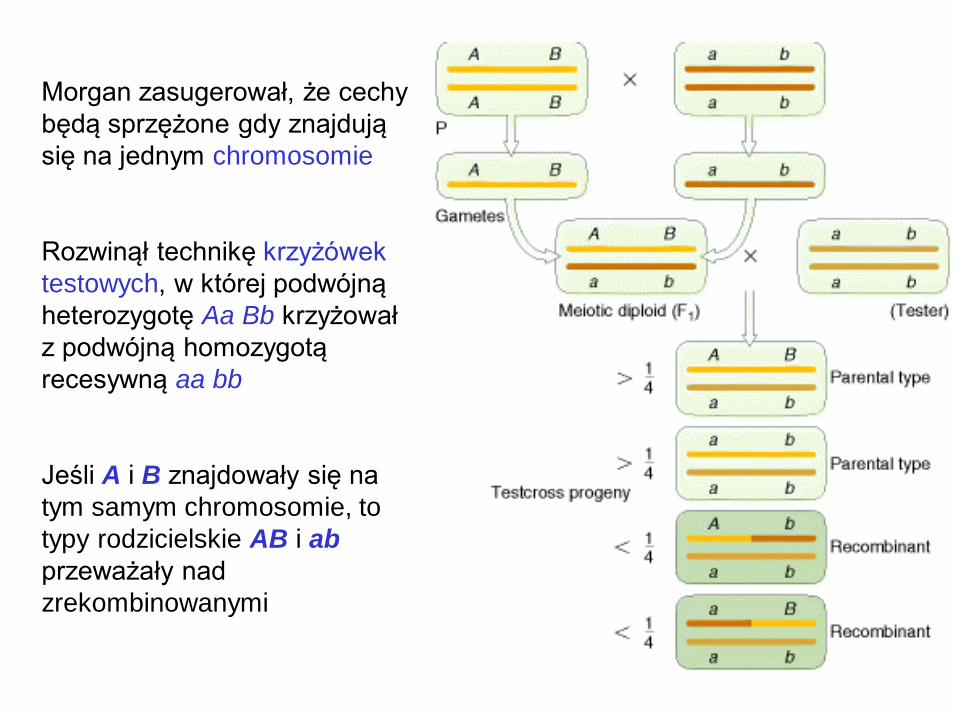
Morgan zasugerował, że cechy
będą sprzężone gdy znajdują
się na jednym chromosomie
Rozwinął technikę krzyżówek
testowych, w której podwójną
heterozygotę Aa Bb krzyżował
z podwójną homozygotą
recesywną aa bb
Jeśli A i B znajdowały się na
tym samym chromosomie, to
typy rodzicielskie AB i ab
przeważały nad
zrekombinowanymi
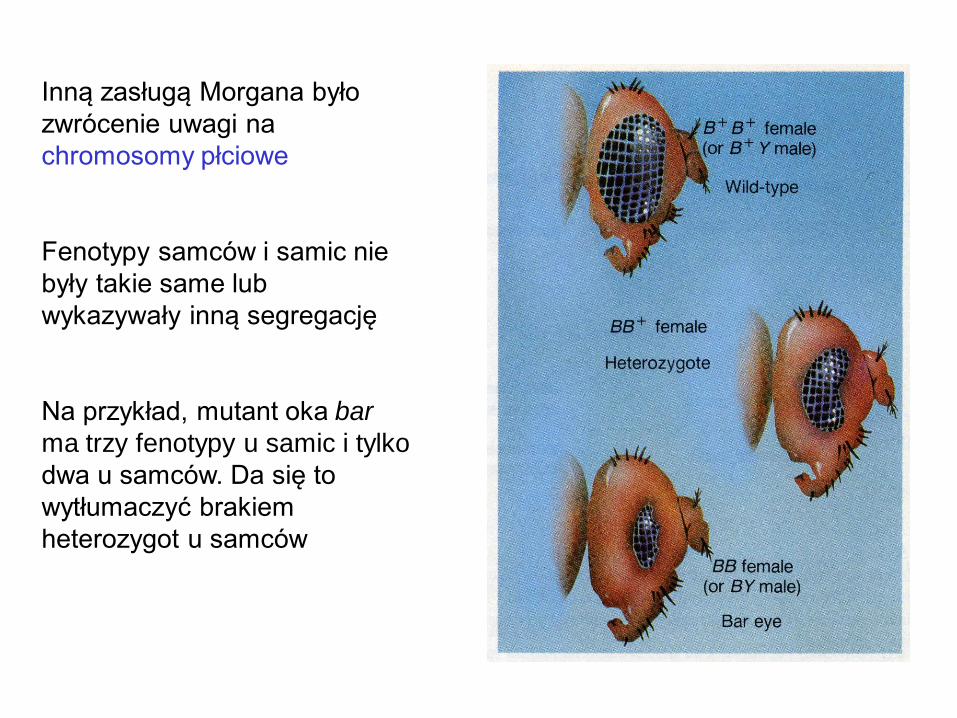
Inną zasługą Morgana było
zwrócenie uwagi na
chromosomy płciowe
Fenotypy samców i samic nie
były takie same lub
wykazywały inną segregację
Na przykład, mutant oka bar
ma trzy fenotypy u samic i tylko
dwa u samców. Da się to
wytłumaczyć brakiem
heterozygot u samców

Układ chromosomów płciowych nie tylko koreluje z płcią ale ją
często determinuje
a) mechanizm równowagowy: liczba chromosomów X względem
autosomalnych, gdy nX:nA jest równe lub większe 1 to mamy samicę, gdy
mniejsze lub równe ½ to samca
b) zwykły mechanizm chromosomowy: obecność Y jest wystarczająca dla
powstania samca
c) determinacja alleliczna, np. u grzybów: nie ma chromosomów płciowych a
płeć zależy od jednego locus (jak inne cechy autosomalne).
Mechanizm równowagi u muszki
XX AA 1:1 normalna samica
XY AA 1:2 normalny samiec
XXX AA 3:2 metasamica (bezpł)
XXY AA 1:1 samica
X AA 1:2 samiec (bezpł)
Mechanizm zwykły w człowieka
XX normalna kobieta
XY normalny mężczyzna
X z. Turnera kobieta)
XXX Triplo-X kobieta
XXXX Tetra-X kobieta
XXY z. Klinefeltera mężczyzna
XXXY z. Klinefeltera mężczyzna
XXYY z. Klinefeltera mężczyzna
XYY XYY mężczyzna
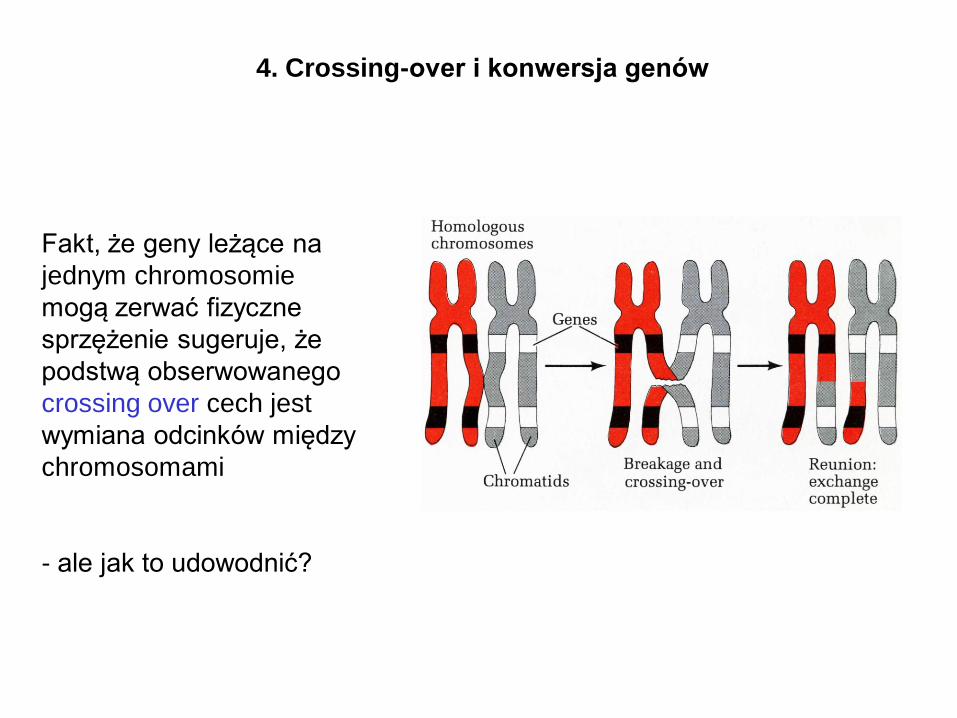
4. Crossing-over i konwersja genów
Fakt, że geny leżące na
jednym chromosomie
mogą zerwać fizyczne
sprzężenie sugeruje, że
podstwą obserwowanego
crossing over cech jest
wymiana odcinków między
chromosomami
- ale jak to udowodnić?
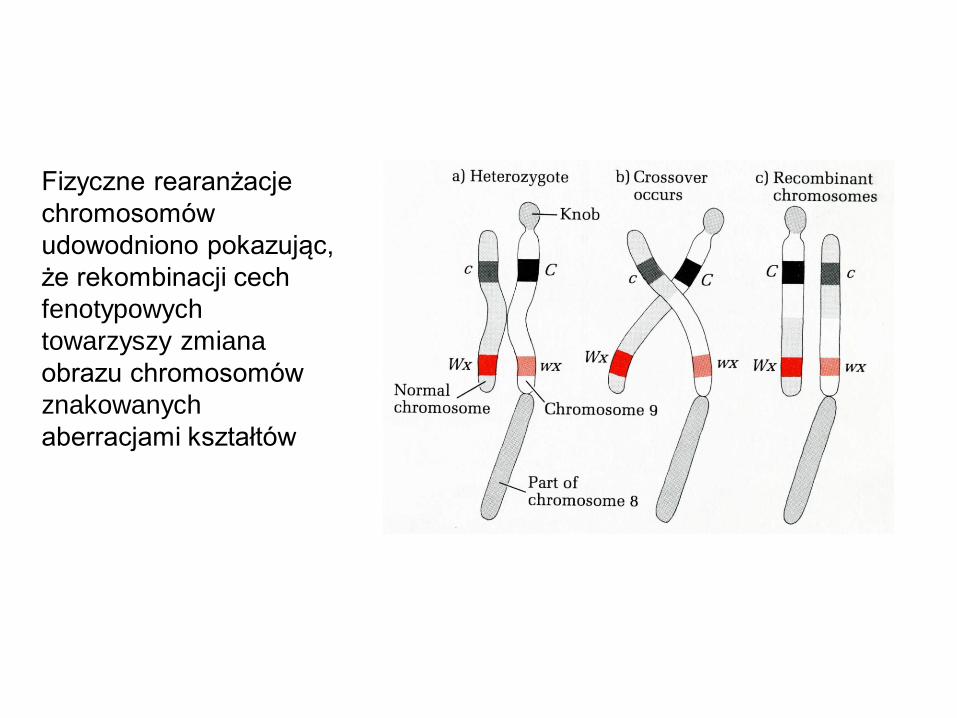
Fizyczne rearanżacje
chromosomów
udowodniono pokazując,
że rekombinacji cech
fenotypowych
towarzyszy zmiana
obrazu chromosomów
znakowanych
aberracjami kształtów
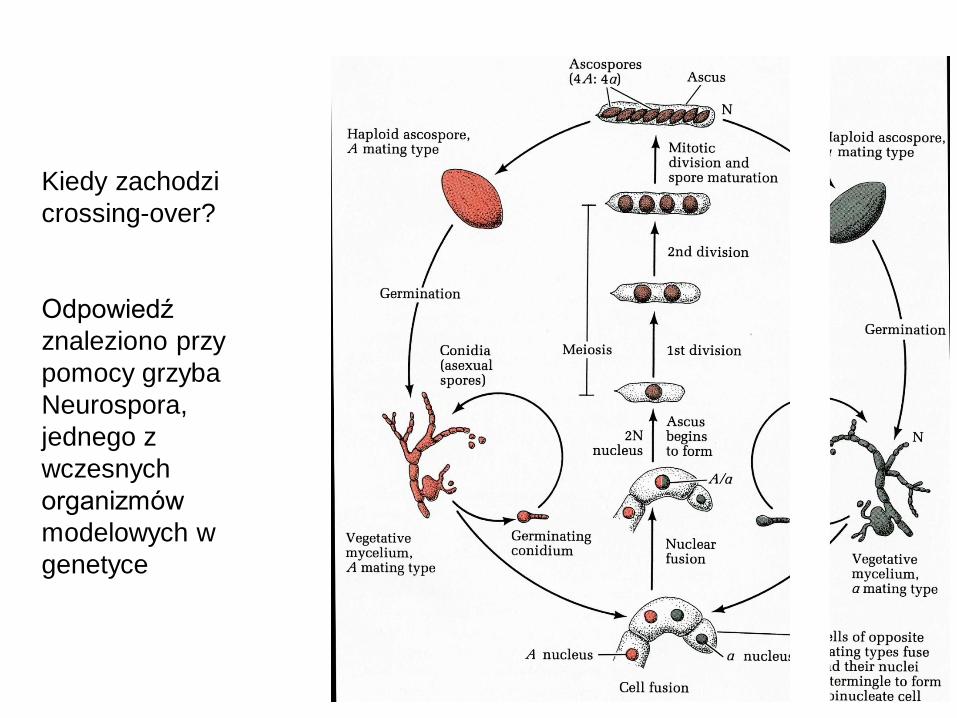
Kiedy zachodzi
crossing-over?
Odpowiedź
znaleziono przy
pomocy grzyba
Neurospora,
jednego z
wczesnych
organizmów
modelowych w
genetyce
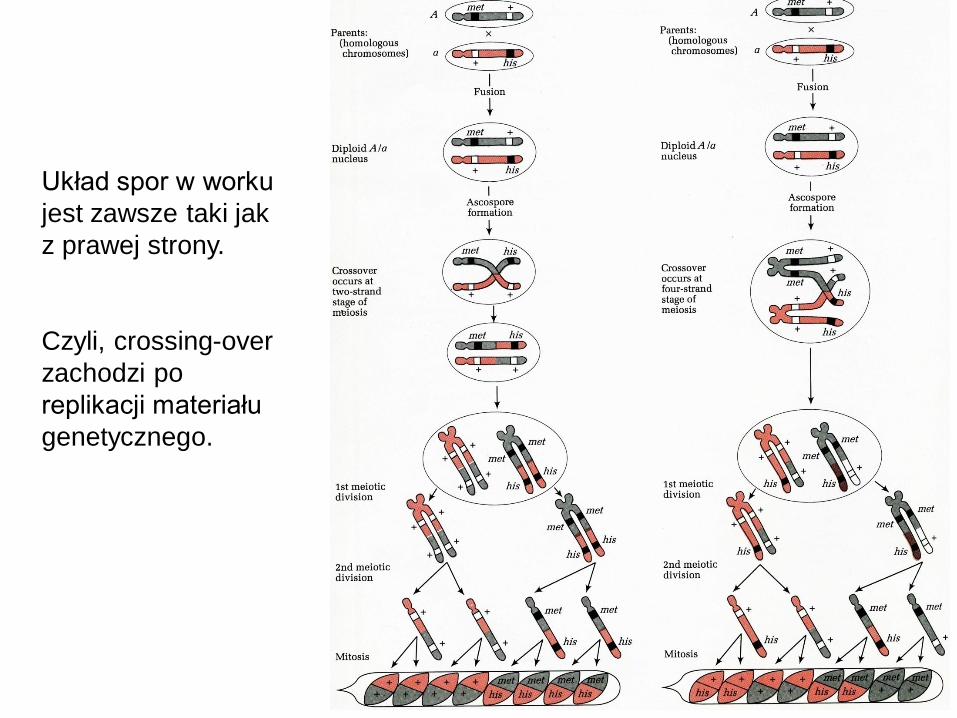
Układ spor w worku
jest zawsze taki jak
z prawej strony.
Czyli, crossing-over
zachodzi po
replikacji materiału
genetycznego.

5. Mapy genetyczne
Jeżeli geny leżą na
chromosomach, a
crossing-over może zajść
na całej ich długości, to ...
... im dalej od siebie geny
leżą tym częściej
zrekombinują.
A. Sturtevant
skonstruował w ten
sposób pierwszą mapę
genetyczną
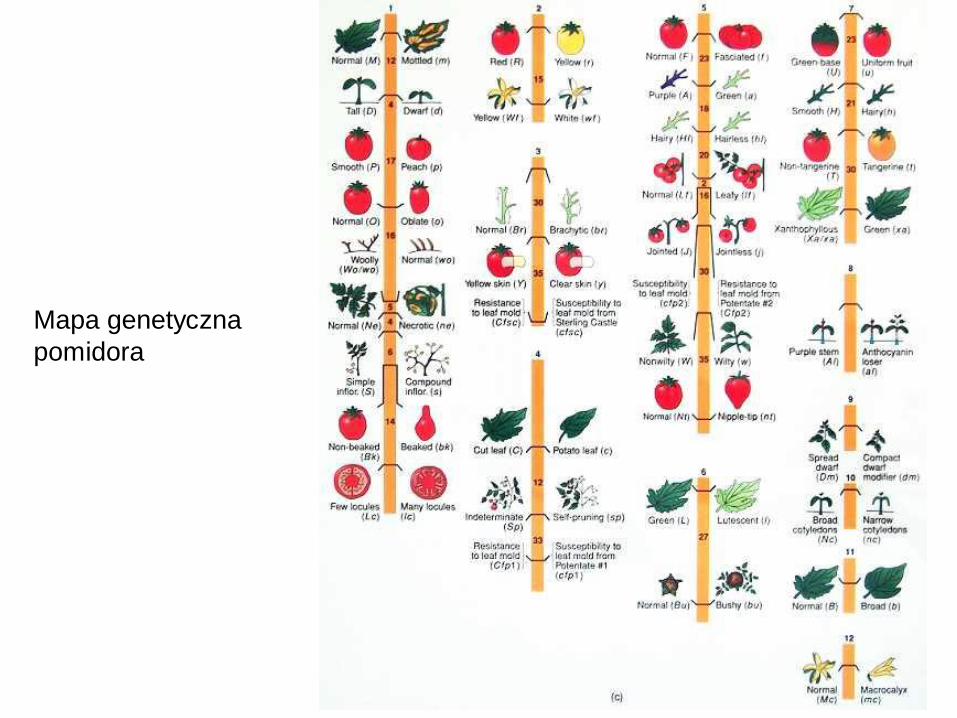
Mapa genetyczna
pomidora
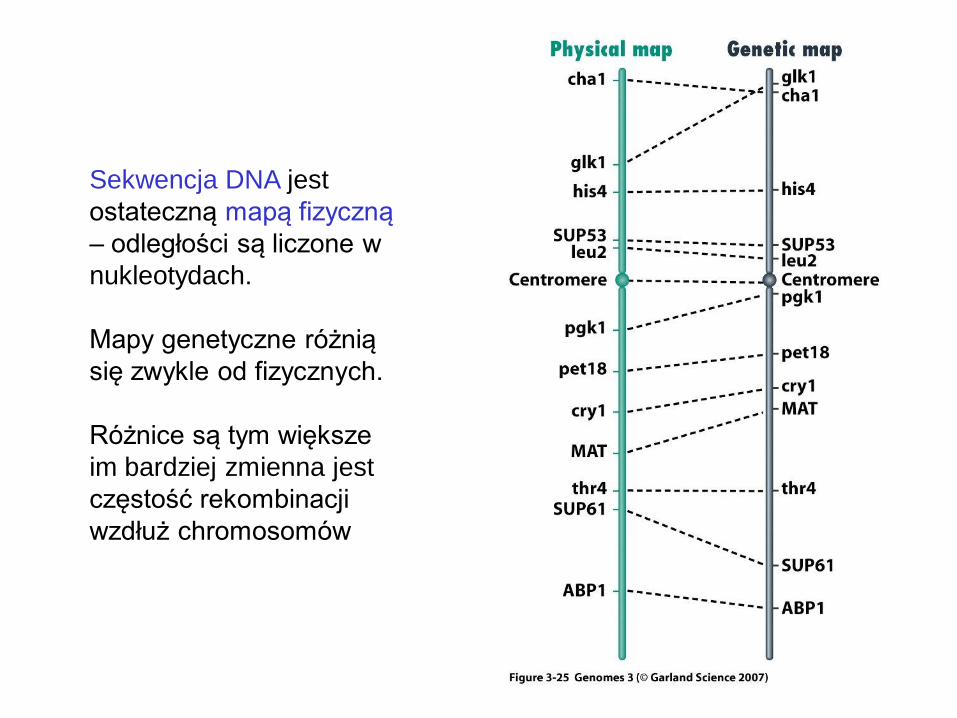
Sekwencja DNA jest
ostateczną mapą fizyczną
– odległości są liczone w
nukleotydach.
Mapy genetyczne różnią
się zwykle od fizycznych.
Różnice są tym większe
im bardziej zmienna jest
częstość rekombinacji
wzdłuż chromosomów
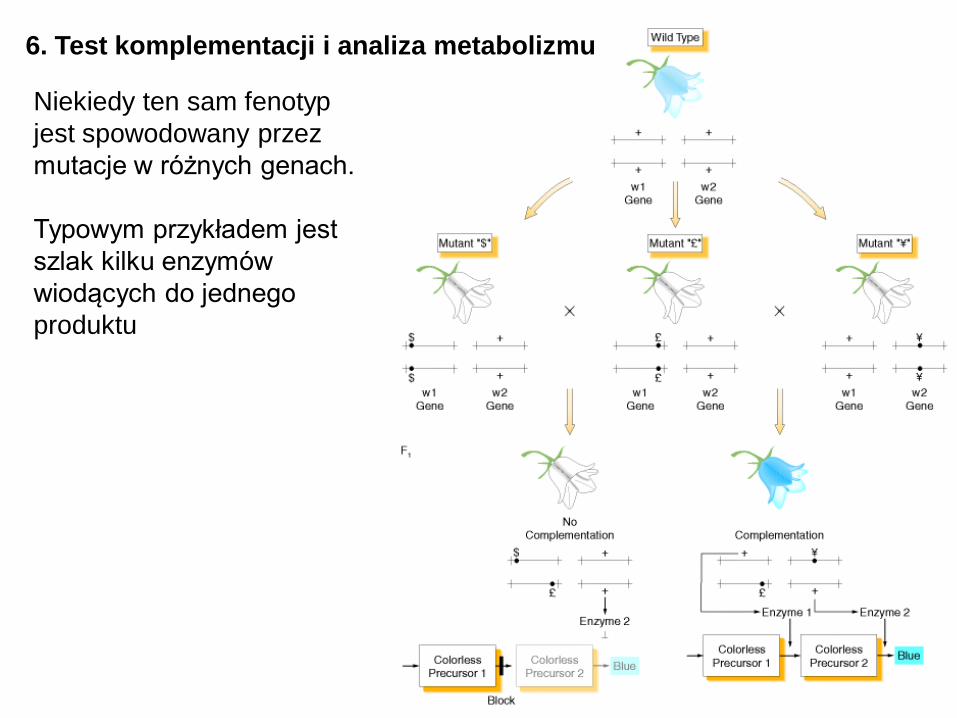
6. Test komplementacji i analiza metabolizmu
Niekiedy ten sam fenotyp
jest spowodowany przez
mutacje w różnych genach.
Typowym przykładem jest
szlak kilku enzymów
wiodących do jednego
produktu

6. Test komplementacji i analiza metabolizmu
Niekiedy ten sam fenotyp
jest spowodowany przez
mutacje w różnych genach.
Typowym przykładem jest
szlak kilku enzymów
wiodących do jednego
produktu
Test komplementacji
pozwala rozróżnić (a także
policzyć) geny wchodzące
w jeden szlak metaboliczny
Systematycznie stosowany,
doprowadził do połączenia
genetyki z biochemią:
„jeden gen – jeden enzym”

7. Test dziedziczenia cytoplazmatycznego
a) eukarionty miewają plazmidy i wirusy, ale poprzez „geny
cytoplazmatyczne” rozumie się najczęściej geny w DNA mitochondriów
lub chloroplastów
c) prostym testem czy dana cecha jest dziedziczona cytoplazmatycznie
jest test (braku) segregacji – czyli większa gameta dyktuje fenotyp
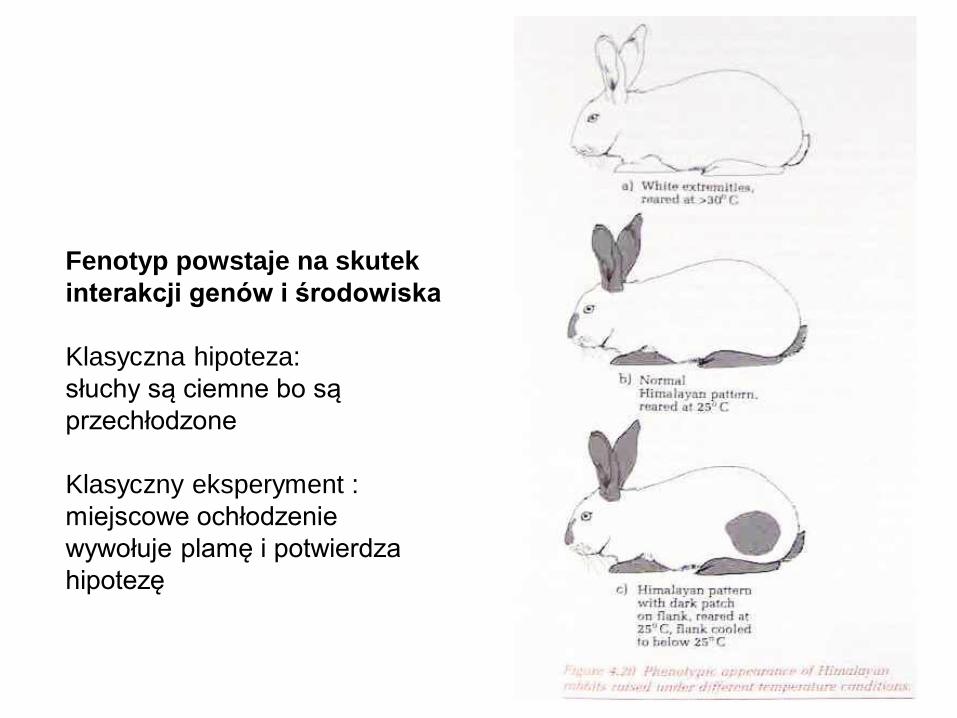
Fenotyp powstaje na skutek
interakcji genów i środowiska
Klasyczna hipoteza:
słuchy są ciemne bo są
przechłodzone
Klasyczny eksperyment :
miejscowe ochłodzenie
wywołuje plamę i potwierdza
hipotezę
Top Related