Two rare AKAP9 variants are associated with Alzheimer's disease … · 2020-01-13 · Two rare...
21
Two rare AKAP9 variants are associated with Alzheimer’s disease in African Americans Mark W. Logue a,b,1 , Matthew Schu a,1 , Badri N. Vardarajan a , John Farrell a , David A. Bennett c , Joseph D. Buxbaum d , Goldie S. Byrd e , Nilufer Ertekin-Taner f , Denis Evans g , Tatiana Foroud h , Alison Goate i , Neill R. Graff-Radford f , M. Ilyas Kamboh j , Walter A. Kukull k , Jennifer J. Manly l , Alzheimer Disease Genetics Consortium 2 , Jonathan L. Haines m , Richard Mayeux l , Margaret A. Pericak-Vance n , Gerard D. Schellenberg o , Kathryn L. Lunetta b , Clinton T. Baldwin a , M. Daniele Fallin p , Lindsay A. Farrer a,b,q,r,s, * a Department of Medicine (Biomedical Genetics), Boston University School of Medicine, Boston, MA, USA b Department of Biostatistics, Boston University School of Public Health, Boston, MA, USA c Rush Alzheimer’s Disease Center, Rush University Medical Center, Chicago, IL, USA d Departments of Neuroscience and Genetics & Genomic Sciences, Mount Sinai School of Medicine, New York, NY, USA e Department of Biology, North Carolina A & T State University, Greensboro, NC, USA f Departments of Neuroscience and Neurology, Mayo Clinic, Jacksonville, FL, USA g Rush Institute for Healthy Aging, Department of Internal Medicine, Rush University Medical Center, Chicago, IL, USA h Department of Medical and Molecular Genetics, Indiana University, Indianapolis, IN, USA i Department of Psychiatry and Hope Center Program on Protein Aggregation and Neurodegeneration, Washington University School of Medicine, St. Louis, MI, USA j Department of Human Genetics and Alzheimer’s Disease Research Center, University of Pittsburgh, Pittsburgh, PA, USA k National Alzheimer’s Coordinating Centerand Department of Epidemiology, University of Washington, Seattle, WA, USA l Department of Neurology and the Taub Institute, Columbia University, New York, NY, USA Abstract Background: Less is known about the genetic basis of Alzheimer’s disease (AD) in African Amer- icans (AAs) than in non-Hispanic whites. Methods: Whole exome sequencing (WES) was performed on seven AA AD cases. Disease associ- ation with potentially AD-related variants from WES was assessed in an AA discovery cohort of 422 cases and 394 controls. Replication was sought in an AA sample of 1037 cases and 1869 controls from the Alzheimer Disease Genetics Consortium (ADGC). Results: Forty-four single nucleotide polymorphisms (SNPs) from WES passed filtering criteria and were successfully genotyped. Nominally significant (P , .05) association to AD was observed with two African-descent specific AKAP9 SNPs in tight linkage disequilibrium: rs144662445 (P 5 .014) and rs149979685 (P 5 .037). These associations were replicated in the ADGC sample (rs144662445: P 5 .0022, odds ratio [OR] 5 2.75; rs149979685: P 5 .0022, OR 5 3.61). m Department of Epidemiology and Biostatistics, Case Western Reserve University, Cleveland, OH, USA n The John P. Hussman Institute for Human Genomics, University of Miami, Miami, FL, USA o Department of Pathology and Laboratory Medicine, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA p Department of Epidemiology, Johns Hopkins University School of Public Health, Baltimore, MD, USA q Department of Neurology, Boston University School of Medicine, Boston, MA, USA r Department of Ophthalmology, Boston University School of Medicine, Boston, MA, USA s Department of Epidemiology, Boston University School of Public Health, Boston, MA, USA 1 These two authors contributed equally to this work. 2 See Acknowledgments for additional ADGC members who contributed to this study. *Corresponding author. E-mail address: [email protected] 1552-5260/$ - see front matter Ó 2014 The Alzheimer’s Association. All rights reserved. http://dx.doi.org/10.1016/j.jalz.2014.06.010 Alzheimer’s & Dementia 10 (2014) 609-618
Transcript of Two rare AKAP9 variants are associated with Alzheimer's disease … · 2020-01-13 · Two rare...

Alzheimer’s & Dementia 10 (2014) 609-618
Two rare AKAP9 variants are associated with Alzheimer’s diseasein African Americans
Mark W. Loguea,b,1, Matthew Schua,1, Badri N. Vardarajana, John Farrella, David A. Bennettc,Joseph D. Buxbaumd, Goldie S. Byrde, Nilufer Ertekin-Tanerf, Denis Evansg, Tatiana Foroudh,Alison Goatei, Neill R. Graff-Radfordf, M. Ilyas Kambohj, Walter A. Kukullk, Jennifer J. Manlyl,
Alzheimer Disease Genetics Consortium2, Jonathan L. Hainesm, Richard Mayeuxl,Margaret A. Pericak-Vancen, Gerard D. Schellenbergo, Kathryn L. Lunettab, Clinton T. Baldwina,
M. Daniele Fallinp, Lindsay A. Farrera,b,q,r,s,*aDepartment of Medicine (Biomedical Genetics), Boston University School of Medicine, Boston, MA, USA
bDepartment of Biostatistics, Boston University School of Public Health, Boston, MA, USAcRush Alzheimer’s Disease Center, Rush University Medical Center, Chicago, IL, USA
dDepartments of Neuroscience and Genetics & Genomic Sciences, Mount Sinai School of Medicine, New York, NY, USAeDepartment of Biology, North Carolina A & T State University, Greensboro, NC, USA
fDepartments of Neuroscience and Neurology, Mayo Clinic, Jacksonville, FL, USAgRush Institute for Healthy Aging, Department of Internal Medicine, Rush University Medical Center, Chicago, IL, USA
hDepartment of Medical and Molecular Genetics, Indiana University, Indianapolis, IN, USAiDepartment of Psychiatry and Hope Center Program on Protein Aggregation and Neurodegeneration, Washington University School of Medicine, St. Louis,
MI, USAjDepartment of Human Genetics and Alzheimer’s Disease Research Center, University of Pittsburgh, Pittsburgh, PA, USAkNational Alzheimer’s Coordinating Center and Department of Epidemiology, University of Washington, Seattle, WA, USA
lDepartment of Neurology and the Taub Institute, Columbia University, New York, NY, USA
mDepartment of Epidemiology and Biostatistics, Case Western Reserve University, Cleveland, OH, USAnThe John P. Hussman Institute for Human Genomics, University of Miami, Miami, FL, USAoDepartment of Pathology and Laboratory Medicine, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA
pDepartment of Epidemiology, Johns Hopkins University School of Public Health, Baltimore, MD, USAqDepartment of Neurology, Boston University School of Medicine, Boston, MA, USA
rDepartment of Ophthalmology, Boston University School of Medicine, Boston, MA, USAsDepartment of Epidemiology, Boston University School of Public Health, Boston, MA, USA
Abstract Background: Less is known about the genetic basis of Alzheimer’s disease (AD) in African Amer-
1These two authors co2See Acknowledgmen
this study.
1552-5260/$ - see fro
http://dx.doi.org/10.10
icans (AAs) than in non-Hispanic whites.Methods: Whole exome sequencing (WES) was performed on seven AA AD cases. Disease associ-ation with potentially AD-related variants fromWES was assessed in an AA discovery cohort of 422cases and 394 controls. Replication was sought in an AA sample of 1037 cases and 1869 controlsfrom the Alzheimer Disease Genetics Consortium (ADGC).Results: Forty-four single nucleotide polymorphisms (SNPs) fromWES passed filtering criteria andwere successfully genotyped. Nominally significant (P , .05) association to AD was observed withtwo African-descent specific AKAP9 SNPs in tight linkage disequilibrium: rs144662445 (P 5 .014)and rs149979685 (P5 .037). These associations were replicated in the ADGC sample (rs144662445:P 5 .0022, odds ratio [OR] 5 2.75; rs149979685: P 5 .0022, OR 5 3.61).
ntributed equally to this work.
ts for additional ADGC members who contributed to
*Corresponding author.
E-mail address: [email protected]
nt matter � 2014 The Alzheimer’s Association. All rights reserved.
16/j.jalz.2014.06.010

M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618610
Conclusions: Because AKAP9 was not previously linked to AD risk, this study indicates a potentialnew disease mechanism.� 2014 The Alzheimer’s Association. All rights reserved.
Keywords: Whole-exome sequencing; Late-onset Alzheimer’s disease; Rare variant; Genetic association; African American;
AKAP9
1. Background
Late onset Alzheimer’s disease (AD) is the most commonform of dementia in the elderly. It has a substantial geneticcomponent, with heritability as high as 75% [1]. The apoli-poprotein E (APOE) gene ε4 variant has been shown to be amajor component of this risk [2]; among non-Hispanicswhites (NHW), ε4/ε4 homozygotes are estimated to haveup to 15-fold increased odds of AD compared with thosewith the most common ε3/ε3 genotype [3]. Genome-wideassociation studies (GWAS) including very large NHW sam-ples have identified genes harboring additional modest-effect AD risk susceptibility loci (odds ratios between 1.1and 1.3) [4–8].
Interestingly, even though the proportion of risk due togenetic influences is similar in African-Americans (AA)and NHWs [9], there is evidence that the genetic architectureof the disease differs between these two populations. Forexample, the effect of APOE ε4 on AD risk is considerablyweaker in AAs than NHWs [3,10,11], although thisdifference may be accounted for in part by the age-dependent effect of ε4 [11]. Several genes associated withAD and AD-related hippocampal atrophy measured by mag-netic resonance imaging in NHWs also show evidence of as-sociation in AAs, however, most SNPs associated withNHWs have not been replicated in AAs [12–14]. While itmay be that the AA studies are underpowered comparedwith studies with NHWs, it also may be due to differencesin the linkage disequilibrium (LD) structure between AAsand NHWs or population-specific causal variants underlyingthe associations. Interestingly, two GWAS replicated the as-sociation with ABCA7 in AAs [12,14] and the risk associatedwith ABCA7 (odds ratio [OR] 5 1.79) approaches that ofAPOE ε4 in AAs (OR 5 2.31) [14]. Nevertheless, a largeproportion of heritability in AAs remains unexplained.Much of the heritability may be ascribed to uncommon(frequency ,5%) or rare (frequency ,1%) variants whichwould be undetectable in GWAS except in very large sam-ples [15]. Causal high-penetrant rare variants in APP,PSEN1, and PSEN2 have been identified primarily in theearly onset familial form of the disease [16]. However, todate no causal variants in these genes have been reportedin AAs.
Next-generation sequencing methodologies have thepotential to identify rare or private risk variants manyof which would be expected to exert moderate to high ef-fects on disease risk compared with most commondisease-associated variants [15,17]. One efficient study
design to detect highly penetrant disease variantsentails sequencing individuals from families withmultiple affected members who are more likely thansporadic cases to possess such variants. Potentiallydisease-causing variants identified in familial cases canthen be genotyped in other family members or in a largersample of unrelated individuals to evaluate associationwith disease. This study design was recently used inCaucasian samples to identify rare variants in the phos-pholipase D3 (PLD3) gene [18]. Resequencing PLD3exons in a larger number of cases and controls identifiedadditional AD-associated PLD3 variants includingAla442Ala, which was subsequently shown to be nomi-nally significantly associated (P 5 .03) with AD in amodest AA sample (130 cases, 172 controls). In thisstudy, we performed whole-exome sequencing (WES)of seven familial AA late-onset AD cases and subse-quently tested association of AD risk with 44 rare vari-ants selected on the basis of frequency, sequencingquality, and relationship to previously implicated ADgenes in 422 AA cases and 394 cognitively normal AAcontrols. We identified two African-descent specific ge-netic variants in the A kinase anchor protein 9(AKAP9) gene which are significantly enriched in AAAD cases versus controls, and subsequently replicatedthis association in a large independent AA cohort. Thestudy design is summarized in Figure 1.
2. Methods
2.1. Study samples
The discovery sample included AA participants of theMulti-Institutional Research on Alzheimer’s Genetic Epide-miology (MIRAGE) Study and the Genetic and Environ-mental Risk Factors for Alzheimer’s Disease amongAfrican Americans Study (GenerAAtions). Ascertainmentmethods, inclusion criteria, and genome-wide genotypingin the MIRAGE and GenerAAtions cohorts are describedin detail elsewhere [9,12]. The MIRAGE cohort consists ofnuclear families most of which are discordant sib pairs.There are also several affected sib pairs and largersibships. Seven MIRAGE-Study AD cases with multipleaffected family members, and thus more likely to be en-riched with rare AD-related variants, were selected forWES. Descriptive statistics for these subjects are summa-rized in Supplementary Table 1. Four of these subjectswere affected sib-pairs. The genotypes of single nucleotide

Fig. 1. The overall study design including a whole-exome sequencing (WES) of seven African-American (AA) late-onset AD cases, followed by variant
prioritization and filtering, genotyping in a larger AA discovery sample, and replication in a large AA Alzheimer’s Disease Genetics Consortium
(ADGC) cohort.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 611
polymorphisms (SNPs) identified as potentially AD relatedin WES were genotyped in 218 cases (including the sevensequenced subjects) and 237 controls from the MIRAGEcohort and 223 cases and 190 controls from the GenerAA-tions cohort. Analyses were performed on a discovery cohortmade up of the genotyped subjects excluding the sevensequenced AD cases and subjects less than 60 years at thetime of assessment. The discovery cohort includes 199 casesand 209 controls from the MIRAGE cohort and 223 casesand 185 controls from the GenerAAtions cohort. Character-istics of the discovery cohort are summarized inSupplementary Table 1. The most significant variants weregenotyped in a replication cohort comprising a subset ofthe Alzheimer’s Disease Genetics Consortium (ADGC)AA sample, excluding the MIRAGE and GenerAAtions co-horts, which contains 1037 cases and 1869 controls(Supplementary Table 2). The ADGC AA cohort isdescribed in detail elsewhere [14]. Studies were performedwith appropriate institutional review board approval andinformed consent was obtained from all study participants.
2.2. Sequencing and genotyping
A detailed description of WES, SNP genotypingmethods and quality control, and Sanger-sequencing usedto confirm genotypes of the most significant SNPs fromthe analysis of the discovery data is provided inSupplementary Methods.
2.3. Variant filtering
Variants identified by WES and passing quality control(QC) were filtered for novelty (absence in the dbSNPdatabase v. 132, using time since discovery as a proxyfor low frequency because rare SNPs are more likely tobe included in later dbSNP revisions) and potential fordeleterious effect (must be a nonsynonymous base pairchange). To filter out possible sequencing artifacts, weadditionally required that variants be observed in morethan one case, whether the variants occurred in siblingsor unrelated subjects. The novel non-synonymous variants

M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618612
were selected for genotyping if they were in genes whichhad been either previously-identified as associated withAD—that is, genes that are listed in the AlzGene data-base—or in genes belonging to networks with establishedAD genes. We constructed gene networks using the IN-GENUITY Pathway Analysis (IPA) software. IPA ana-lyses were seeded with genes emerging from WES andestablished risk genes for early or late onset AD(APOE, BIN1, CLU, CR1, PICALM, MS4A6E, CD2AP,CD33, ABCA7, EPHA1, SORL1, ACE, PSEN1, PSEN2,and APP) [2,4–8,19–25] to identify genes and thevariants contained therein that might be relatedphysiologically to AD-related processes or are related togenes with a known connection to AD. These networksdo not exclusively consist of genes involved in a singleAD-related biological function, but instead include genesthat interact, either directly or indirectly, with known ADgenes and hence may implicate new disease-related path-ways.
Table 1
The most significant associations between AD and genotyped SNPs in the
discovery cohort
Gene Position SNP ID Freq cases Freq controls P
AKAP9* 7:91709085 rs144662445 0.011 0.0013 .014*
AKAP9* 7:91732110 rs149979685 0.0060 0.000 .037*
ATP2B4 1:203672867 rs145963279 0.023 0.013 .092
LRP4 11:46911956 rs138878258 0.0072 0.0026 .17
ITGA9 3:37774225 rs142726080 0.032 0.024 .20
Abbreviations: AD, Alzheimer’s disease; Freq, frequency; SNP, single-
nucleotide polymorphism.
*Genotypes for AKAP9 SNPs confirmed via Sanger sequencing.
2.4. Statistical analysis
Tests of association between genotyped SNPs and ADwere performed using R [26]. Significance was determinedusing a one-sided Fisher’s exact test because rare SNPscan cause numerical stability issues in logistic models, andbecause we specified a priori that our search was focusedon deleterious rare variants. The test of association withinthe discovery cohort may underestimate the strength of atrue association in the MIRAGE data set because many ofthe controls in this sample are unaffected siblings of ADcases who would presumably be carriers of risk alleles at ahigher rate than unrelated controls. Although the MIRAGEstudy ascertained primarily discordant sib-pairs, the samplecontained three affected sib-pairs including the two affectedsib-pairs in the WES sample. Both members of the sib-pairnot included in theWES sample were included in the discov-ery cohort. The possibility of inflated significance due tonon-independence of the samples is unlikely (see the gener-alized estimating equation [GEE] analysis in theSupplementary Results). Principal components (PC) anal-ysis with EIGENSTRAT [27] based on all minor allele fre-quency (MAF) .5% markers from a whole-genomegenotyping panel (described in [12]) found no evidence ofAD-associated population substructure within the discoverysample (all P . .05 on tests of association between AD andeach of the first 10 PCs). In the replication sample, SNPswere analyzed for association with AD using a logisticregression model adjusting for sex and study site. Methodsfor the haplotype, phylogenetic dendogram, and bio-informatic analyses (using PolyPhen-2 [28], Sorts IntolerantFrom Tolerant substitutions [SIFT] [29], and MutPred [30]software) are described in Supplementary Methods. Haplo-type analyses were performed with PLINK [31] using bothWES variant data and tag SNPs from whole-genome geno-typing.
3. Results
A total of 88,867 variants were identified by WES. Nocoding APP, PSEN1, PSEN2, or PLD3 variants were identi-fied in the seven AA AD cases. After filtering, 63 SNPs from58 genes were selected for further study and 44 of theseSNPs were successfully genotyped in the entire sample.Detailed results of sequencing and variant filtering are pre-sented in the supplementary materials, including a completelist of variants that were genotyped (SupplementaryTable 4). Most genotyped SNPs were rare (29 SNPs had aMAF ,1%). Nominally significant results were obtainedwith AKAP9 SNPs rs144662445 (P 5 .014, OR 5 8.4) andrs149979685 (P 5 .037; the OR for rs149979685 could notbe estimated because the rare allele was not observed inany controls) (Table 1, Supplementary Table 5). The geno-types for these AKAP9 SNPs were confirmed by Sangersequencing. Five AD cases, in addition to the two casesincluded in WES, were observed with rare alleles for bothrs144662445 and rs149979685 (Supplementary Table 6).Five other subjects (four AD cases and one control) hadthe minor allele for rs144662445 but not rs149979685. Sub-jects having at least one of these two rare variants are desig-nated AKAP91. No subjects were observed having theminor allele for rs149979685 but not rs144662445. Norare-allele homozygotes were observed for either SNP.One control subject who was genotyped but not includedin the discovery sample due to age—a 54-year-old unaf-fected sibling of an AD proband (whose age at onset was64)—had rare alleles for both rs144662445 andrs149979685. The AKAP91 subjects are all unrelated ac-cording to their self-reported data and identity by descent(IBD) estimates calculated by PLINK [31] using genome-wide SNP data. A PC analysis in EIGENSTRAT [27] indi-cated that these subjects were not members of a clearlydefined population subgroup within AAs (SupplementaryFigure 1). AKAP9 genotypes were available for relatives offour AKAP91AD cases. Two of these cases each had an un-affected sibling who was AKAP9-. Each of the other twoAKAP91 cases who were included in the WES discoverysample had an AKAP9-affected sibling.
Next, we sought replication of the association onlywith the two AKAP9 SNPs in the ADGC AA sample.

M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 613
Valid genotypes for both SNPs were obtained in 1037 ADcases and 1869 controls. In this sample, the average ageat onset among AD cases was 80.1 and the average exam-ination age for controls was 74.5. Average ages weresimilar among AKAP91 (79.7) and AKAP2 (80.1) cases(P 5 .83) and among AKAP91 (75.9) and AKAP2(74.5) controls (P 5 .47). The APOE ε4 allele frequencywas 33.7% in cases and 18.5% in controls in the replica-tion sample. This does not differ between AKAP91(38.8%) and AKAP92 (33.6%) cases (P 5 .59) or be-tween AKAP91 (12.5%) and AKAP92 (18.6%) controls(P 5 .50). A logistic regression model adjusting for ageand study site demonstrated significant association ofAD risk with the minor alleles of both rs144662445(P 5 .0022, OR 5 2.75) and rs149979685 (P 5 .0022,OR 5 3.61) (Table 2). Hence, the association observedin the discovery data was replicated in a larger indepen-dent sample. A logistic regression analysis of the subsetof ADGC AA replication subjects who had bothAKAP9 SNP genotypes and genome-wide genotype data(877 cases and 1641 controls) yielded nearly identical re-sults for both SNPs when PCs were included(rs144662445 P 5 .0034, OR 5 2.72 versus P 5 .0035,OR 5 2.72 with PCs; 149979685 P 5 .0064,OR 5 3.17 versus P 5 .0063, OR 5 3.19 with PCs), indi-cating that population substructure did not contribute tothe confirmation of the AKAP9 association in the replica-tion sample.
To examine the possibility that an untested AD causalvariant in AKAP9 or surrounding region could account forthe observed association, we compared haplotypes ofAKAP91 and AKAP92 individuals in the discovery sample.The 50 end of AKAP9 overlaps a large LD block which in-cludes the neighboring genes CYP51A1, KRIT1, ANKIB1,and LRRD1 (see Supplementary Figure 2). Haplotype anal-ysis of the 10 common (MAF .0.05) AKAP9 SNPs in theGWAS-genotyping array revealed that all of the AKAP91subjects harbored the same haplotype (designated HAP0).That is, the minor alleles (either as a singleton or together)are present on the haplotype GGAAGGAAGC as definedby rs1859037, rs2299233, rs2282973, rs6465347,rs2158138, rs733957, rs2079082, rs13239875, rs4265, andrs1063243 (Supplementary Table 7). The frequency of thishaplotype is greater in AD cases (6.1%) compared with un-related controls (4.1%), but this difference is not quite signif-
Table 2
The association between AKAP9 variants and AD using a logistic regression adju
SNP ID
Frequency (proportion) of minor alleles
Cases Controls
rs144662445 0.011 (22y/2052) 0.0043 (16/372
rs149979685 0.0072 (15/1059) 0.0027 (10/372
Abbreviations: AD, Alzheimer’s disease; CI, confidence interval; OR, odds rat
*Effect size estimates, CI, and P values were derived from a logistic regressionyIncludes one AD case who was homozygous for the minor allele.
icant (P5 .071). No other coding variants in this region wereobserved in the two AKAP91 subjects with WES datathrough whom this association was ascertained.
Next, we performed phylogenetic dendogram analysis ofthe SNP information in the haplotype region among 11 of the246 African-descent subjects in 1000 Genomes for whomgenotypes are available (including 88 Yorubans fromNigeria, 97 Luhyans from Kenya, and 61 AAs from theAmerican Southwest) and who had the HAP0 haplotype asdefined by the common SNPs. Three of these 11 individuals(NA18499, NA19448, and NA20126) had minor alleles atboth rs144662445 and rs149979685, and one individual(NA20298) had only the rs144662445 variant. TheseAKAP9 variants were observed in each of the threeAfrican-descent groups suggesting that the AKAP9 variantsare present in multiple African ancestral populations. Clus-ter analysis of the phased alleles for the 22 haplotypesfurther discriminated the HAP01 haplotypes from theHAP02 haplotypes (Figure 2A). There is also a branch pointseparating the HAP01 haplotype containing one AKAP9variant (designated HAP011) from the haplotype contain-ing both AKAP9 variants (HAP012).
Based on these results, we examined the 1000 Ge-nomes phased haplotypes for plausible exonic candidateAD-risk SNPs on the same haplotypes as rs144662445and rs149979685 by comparing the high-risk haplotypes(HAP012 and HAP011) to the low-risk haplotype(HAP02). Within the region from 91.3 Mb to 92.1Mb—which extends well beyond the regions sharedacross those harboring AKAP9 mutations (Figure 2B)—the three subjects with HAP012 haplotypes shared 69variants with MAF ,10% that were not observed in sub-jects with the HAP02 haplotype. Rs144662445 andrs149979685 are the only coding variants in this groupof SNPs. Next, we compared the single copy ofHAP011 from 1000 Genomes (NA20298A) to HAP02.In the same region, NA20298A contains 49 variantswith MAF ,10%, which were not observed on the(low-risk) HAP02 haplotypes, including three codingvariants: rs144662445 and two additional nonsynonymouscoding AKAP9 SNPs rs186619641 and rs201858518.Rs186619641 and rs201858518 are unique to individualNA20298 in the 1000 Genomes database. Using only1000 Genomes data we cannot determine whether thesetwo additional nonsynonymous variants are shared by
sted for sex and study site in the replication cohort
OR* 95% CI P
2) 2.75 1.38–5.61 .0022
8) 3.61 1.51–9.00 .0022
io; SNP, single nucleotide polymorphism.
model adjusted for sex and study site.

M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618614

=
Table 3
Bioinformatic Examination of the AD-associated nonsynonymous AKAP9 variants
SNP
Polyphen-2 SIFT MutPred
Score* Prediction Scorey Prediction Probability deleteriousz Actionable hypothesesz
rs144662445 0.008 Benign 0.11 Tolerated 21% NA
rs149979685 0.99 Probably damaging 0.01 Damaging 57% Loss of helix (P 5 .0033)
Gain of loop (P 5 .024)
Loss of phosphorylation (P 5 .029)
Abbreviations: AD, Alzheimer’s disease; NA, not applicable; SNP, single nucleotide polymorphism; SIFT, Sorts Intolerant from Tolerant substitutions.
*In Polyphen-2 higher scores indicate that a SNP is more likely to be damaging.yIn SIFT, lower scores indicate that a SNP is more likely to be deleterious.zIn MutPred, a higher probability indicates that a SNP is more likely to be damaging. Actionable hypotheses are predicted functional changes for which the
probability deleterious is ,50% and for which the structural change P , .05.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 615
all subjects in our study with HAP011, present in only aportion of subjects with HAP011 haplotypes, or uniqueto 1000 Genomes individual NA20298 who is the onlyAA subject in 1000 Genomes with one HAP011 haplo-type. We sought clarification in the ESP database which,based on allele counts of the AD-associated SNPsrs144662445 and rs149979685, contains 3 copies ofHAP011. These two exonic SNPs (rs186619641 andrs201858518) were absent from the ESP database sug-gesting that the two AD-associated variants definingHAP011 and HAP012 are not surrogate markers forrs186619641 and rs201858518. Therefore, based on ourexamination of HAP011 and HAP012 in public data-bases, we conclude that rs144662445 and rs149979685are the only plausible AD-related coding variants on thesebackground haplotypes within AKAP9 or the surroundingregion.
Bioinformatic analysis using several methods suggeststhat rs149979685 (S3771L transcript Q99996-1) is likelyto be functional whereas rs144662445 (I2558M transcriptQ99996-1) is not (Table 3). The MutPred analysis also indi-cated that the most likely impact of the rs149979685 variantis loss of a helix (P5 .0033), but gain of a loop (P5 .024) orloss of phosphorylation (P5 .029) are also possible. Howev-er, because MutPred makes these predictions based on ho-mology to known functional variants and the 3D structureof AKAP9 is yet unknown, it is unclear whether these struc-tures are present near rs149979685 or critical to the functionof AKAP9. The two NA20298-specific non-synonymousSNPs (rs186619641 and rs201858518) were also predictedto be benign according to Polyphen-2.
4. Discussion
In a small WES study of seven familial AD cases, weidentified two AKAP9 SNPs which co-occur on a single un-
Fig. 2. Examination of the 1000 Genomes subjects with the haplotype containing th
haplotypes and (B) positions of infrequent single nucleotide polymorphisms (SNPs
Key: HAP0: The background haplotype which contains the two identified AKAP9
tatively Alzheimer’s disease (AD)-associated variants; HAP02: the background h
haplotype with one of the AD-associated AKAP9 variants; HAP012 the backgrou
common background haplotype (HAP0) spanning AKAP9and the surrounding area. These SNPs were significantly en-riched in AD cases compared with cognitively normal con-trols in a large African-American sample, and theassociations were replicated in a larger independent AAcohort. These missense variants are rare in African-descentpopulations (ESP AA sample: MAFrs144662445 5 0.43%and MAFrs149979685 5 0.36%; 1000 Genomes AFR sample:MAFrs144662445 5 0.81% and MAFrs149979685 5 0.61%).They are not present in approximately 4000 Caucasians inthe ESP database or in currently available data for 379 Euro-pean and 286 East-Asian subjects in the 1000 Genomes data-base. Although the estimated OR’s are lower in thereplication sample compared with the discovery sample(as expected due to ‘winner’s curse’), the magnitude of ef-fect of these variants on AD risk in the replication sample(estimated ORs of 2.75 and 3.61) is slightly larger thanthat reported for APOE ε4 in AAs (OR5 2.31) [14]. Hence,as expected given their frequency, the identified AKAP9 riskvariants not likely to explain a great proportion of AD risk inAAs. However, those inheriting at least one risk allele have asubstantially increased risk of AD. Bioinformatic analysisindicated that one of these SNPs (rs149979685) likely hasan adverse impact on the structure or post-translationalmodification of the AKAP protein.
We observed many differences in the 1000 Genomes data-base between the HAP0 haplotype containing the AD-associated AKAP9 SNPs and HAP0 haplotypes lacking bothmutations, suggesting that the instances of the rarers149979685 and rs144662445 variants are likely identicalby descent, although derived from a very oldmutational event.This conclusion is supported by the observation of these vari-ants in subjects of diverse African ancestry and by evidence inour study that subjects sharing these alleles do not appear to berelated or from a particular population subgroup based ongenome-wide measures of IBD sharing and PC analysis.
e two identified AKAP9 variants including (A) a phylogenetic dendogram of
; 5% minor allele frequency) shared by haplotypes with 0, 1, and 2 variants.
variants. HAP01: the background variant with one or more of the two pu-
aplotype without either AD-associated variants; HAP011 the background
nd haplotype with both AD-associated AKAP9 variants.

M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618616
AKAP9, located on chromosome 7q21-22, encodes amember of the A kinase anchoring protein (AKAP) fam-ily. AKAPs bind or tether protein kinase A (PKA) andother signaling molecules to relevant targets [32].AKAP9 is differentially spliced into isoforms whichlocalize to different cellular compartments [32] and areinvolved in distinct biological processes. AKAP9 geneproducts include a short isoform called Yotiao and longisoforms (AKAP350, AKAP450, and CG-NAP) oftenreferred to collectively as AKAP450 based on their mo-lecular weight.
AKAP9 was identified in a cDNA library of human brainwhere the Yotiao isoform is expressed in the hippocampus,cerebellum, and cerebral cortex [33]. A SNP in Yotiao hasbeen identified as a cause of long-QT syndrome [34], acongenital heart abnormality characterized by long-QT in-tervals and arrhythmias. Disruption of the binding of PKAand AKAPs (including AKAP5 and Yotiao) at nerve termi-nals in the hippocampus has been shown to interfere withcellular mechanisms associated with spatial memory [35].The locations of rs144662445 and rs149979685 are approx-imately 38 kb 30 and 61 kb 30, respectively, from the bound-aries of the Yotiao coding region. Therefore, these variantsprobably do not affect the Yotiao isoform, unless they areregulatory or simply in LD with unidentified non-codingrisk variants. Hence it is more likely that the AD-associated AKAP9 variants affect the structure or functionof AKAP450.
AKAP450 localizes to centrosomes of cells [36]. It is ex-pressed in most tissues including brain. Disrupted binding ofAKAP450 to the centrosome interferes with centriole dupli-
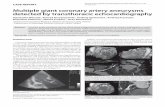
Fig. 3. Annotated protein domain of AKAP9 canonical transcript Q99996-1 inclu
centrosomal targeting (PACT) domain; Long QT syndrome amino-acid change; an
rs144662445 (I2558M) and rs149979685 (S3771L).
cation and cell cycle progression [37]. Studies have alsoshown that AKAP450 is involved in microtubule anchoringand organization at the centrosome [38] and necessary forthe initiation of new microtubule formation on the cis-sideof the golgi [39]. The non-synonymous change induced byrs144662445 (I2558M transcript Q99996-1) is 5 amino acidsfrom the R2 binding site of AKAP450 (Figure 3).Rs149979685 (S3771L transcript Q99996-1) is locatedwithin the pericentrin/AKAP450 centrosomal targetingdomain which is a highly conserved region near the 30 endof AKAP450 and is the region responsible for the localiza-tion of AKAP450 to the centrosome [40] (Figure 3). Therole of AKAP450 in AD is unclear, however, it has func-tional similarity with tau protein which is involved in micro-tubule stability and assembly and is a key constituent ofneurofibrillary tangles which accumulate in AD brain [41–44]. Reports also indicate that phosphorylated tau does notaccount for all of the microtubule loss and shorteningobserved in neurons of those with AD [45]. Thus, it ispossible that functional defects in AKAP450 may contributeto AD pathogenesis.
These results should be interpreted cautiously. The bio-informatics programs have predicted that rs149979685 hasa possible function while rs144662445 does not. However,without functional data and supporting molecular work, wecannot definitively demonstrate that rs149979685 is thecausal locus or whether rs144662445 is potentially causaldue to some other mechanism not accounted for in the soft-ware used, or even whether another non-coding regulatoryvariant in LD with both rs144662445 and rs149979685 isresponsible for the observed association with AD.
ding Yotiao; AKAP450; the PKA binding sites; the pericentrin/AKAP450
d the conservation of the two Alzheimer’s disease (AD)-associated variants

M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 617
However, examination of the region in 1000 Genomes andthe ESP database indicates that such a variant is not likelyto be a non-synonymous coding change. Resolution of thisquestion will require functional experimentation. Finally,we acknowledge that our findings do not represent thebreadth of possible AD-related rare variation in AfricanAmericans because most such variants would not be pre-sent in the exons of seven individuals. Thus, for example,it is not surprising that we did not observe previously re-ported rare AD-risk variants in PLD3 [18]. It is alsopossible that several of the non-AKAP9 variants identifiedby WES and selected for genotyping are actually true riskloci which the discovery sample was underpowered toconfirm by association. Targeted resequencing in both Af-rican- and European-descent populations will be necessaryto identify the full complement of genetic variants whichwill provide greater understanding of the role of AKAP9in AD pathogenesis.
Acknowledgments
We gratefully acknowledge the assistance of Madison Steeleand Alison Peltz, participants in Boston University’sResearch Internship in Science & Engineering (RISE) Pro-gram.Additional Members of the Alzheimer’s Disease GeneticsConsortiumWho Contributed to This Study: Lisa L. Barnes,PhD, Guiqing Cai, PhD, Laura B. Cantwell, MPH, Philip L.De Jager, MD, PhD, Rodney C. P. Go, PhD, Patrick Griffith,MD, Rosalyn Lang, PhD, Oscar L. Lopez, MD, Thomas O.Obisesan, MD, Towfique Raj, PhD.Funding Support: This work supported by NIA/NIH grantsR01-AG09029, R01-AG025259, P30-AG13846 (Dr. Farrer);U01-AG032984, RC2-AG036528, U01-AG016976 (Dr. Ku-kull); U24-AG026395, U24-AG026390; R01-AG037212,R37-AG015473 (Dr. Mayeux); U24-AG021886 (Dr. Foroud);R01-AG20688 (Dr. Fallin); P50-AG005133, AG041718,AG030653 (Dr. Kamboh); R01-AG019085 (Dr. Haines);R01-AG1101, R01-AG030146, RC2-AG036650 (Dr. Evans);P30-AG10161, R01-AG15819, R01-AG17917 (Dr. Bennett);R01AG028786 (Dr. Manly); R01-AG22018, P30-AG10161(Dr. Barnes); P50AG-16574, R01-032990, KL2-RR024151(Dr. Ertekin-Taner); R01-AG027944, R01-AG028786 (Dr.Pericak-Vance); P20-MD000546, R01-AG28786 (Dr. Byrd);AG005138 (Dr. Buxbaum); P50-AG05681, P01-AG03991,P01-AG026276 (Dr. Goate); and UO1-AG032984 (Dr. Schel-lenberg).Conflicts of Interest: The authors of this paper have no con-flicts of interest to report.Web Resources: AlzGene database: http://www.alzgene.org;INGENUITY Pathway Analysis: http://www.ingenuity.com; Applied Biosystems’ Sequence Scanner Softwarev1.0: https://products.appliedbiosystems.com/ab/en/US/adirect/ab?cmd5catNavigate2&catID5600583; TheR project for stastistical computing: http://www.r-project.org; 1000 Genomes Project: www.1000genomes.org;
PLINK:http://pngu.mgh.harvard.edu/wpurcell/plink/; NHLBI Grand Opportunity Exome Sequencing Project (ESP):https://esp.gs.washington.edu/drupal/.
RESEARCH IN CONTEXT
1. Systematic review: As members of the Alzheimer’sDisease Genetics Consortium, the authors havebeen privy to much of the relevant literature as it de-velops. PubMed and Google searches were used toensure that other relevant publications were not over-looked.
2. Interpretation: This is the third report of rare Alz-heimer’s disease (AD) risk variants identified bywhole-exome sequencing (WES) and the firstreport in African Americans (AAs). Moreover, theestimated magnitude of effect of these variants isslightly larger than estimated risk due to APOE ε4 inAAs. This study indicates a potential new mecha-nism for AD.
3. Future directions: One of the identified rare variantsis predicted to be deleterious, but whether this variantis causal will need to be confirmed in functionalstudies. Additionally, Sequencing in AA and otherpopulations will be necessary to find the completeset of AD-associated AKAP9 variants.
References
[1] Bienvenu OJ, Davydow DS, Kendler KS. Psychiatric ’diseases’ versus
behavioral disorders and degree of genetic influence. Psychol Med
2011;41:33–40.
[2] Saunders AM, Strittmatter WJ, Schmechel D, George-Hyslop PH,
Pericak-Vance MA, Joo SH, et al. Association of apolipoprotein E
allele epsilon 4 with late-onset familial and sporadic Alzheimer’s dis-
ease. Neurology 1993;43:1467–72.
[3] Farrer LA, Cupples LA, Haines JL, Hyman B, Kukull WA, Mayeux R,
et al. Effects of age, sex, and ethnicity on the association between
apolipoprotein E genotype and Alzheimer disease. A meta-analysis.
APOE and Alzheimer Disease Meta Analysis Consortium. JAMA
1997;278:1349–56.
[4] Lambert JC, Heath S, Even G, Campion D, Sleegers K, Hiltunen M,
et al. Genome-wide association study identifies variants at CLU and
CR1 associated with Alzheimer’s disease. Nat Genet 2009;41:1094–9.
[5] Seshadri S, Fitzpatrick AL, Ikram MA, DeStefano AL, Gudnason V,
Boada M, et al. Genome-wide analysis of genetic loci associated
with Alzheimer disease. JAMA 2010;303:1832–40.
[6] Harold D, Abraham R, Hollingworth P, Sims R, Gerrish A,
Hamshere ML, et al. Genome-wide association study identifies vari-
ants at CLU and PICALM associated with Alzheimer’s disease. Nat
Genet 2009;41:1088–93.
[7] Naj AC, Jun G, BeechamGW,Wang LS, Vardarajan BN, Buros J, et al.
Common variants at MS4A4/MS4A6E, CD2AP, CD33 and EPHA1

M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618618
are associated with late-onset Alzheimer’s disease. Nat Genet 2011;
43:436–41.
[8] Hollingworth P, Harold D, Sims R, Gerrish A, Lambert JC,
Carrasquillo MM, et al. Common variants at ABCA7, MS4A6A/
MS4A4E, EPHA1, CD33 and CD2AP are associated with Alzheimer’s
disease. Nat Genet 2011;43:429–35.
[9] Green RC, Cupples LA, Go R, Benke KS, Edeki T, Griffith PA, et al.
Risk of dementia among white and African American relatives of pa-
tients with Alzheimer disease. JAMA 2002;287:329–36.
[10] Tang MX, Stern Y, Marder K, Bell K, Gurland B, Lantigua R, et al.
The APOE-epsilon4 allele and the risk of Alzheimer disease
among African Americans, whites, and Hispanics. JAMA 1998;
279:751–5.
[11] Graff-Radford NR, Green RC, Go RC, Hutton ML, Edeki T,
Bachman D, et al. Association between apolipoprotein E genotype
and Alzheimer disease in African American subjects. Arch Neurol
2002;59:594–600.
[12] LogueMW, SchuM, Vardarajan BN, Buros J, Green RC, Go RC, et al.
A comprehensive genetic association study of Alzheimer disease in
African Americans. Arch Neurol 2011;68:1569–79.
[13] Melville SA, Buros J, Parrado AR, Vardarajan B, Logue MW, Shen L,
et al. Multiple loci influencing hippocampal degeneration identified by
genome scan. Ann Neurol 2012;72:65–75.
[14] Reitz C, Jun G, Naj A, Rajbhandary R, Vardarajan BN,Wang LS, et al.
Variants in the ATP-binding cassette transporter (ABCA7), apolipo-
protein E 4,and the risk of late-onset Alzheimer disease in African
Americans. JAMA 2013;309:1483–92.
[15] Cirulli ET, Goldstein DB. Uncovering the roles of rare variants in com-
mon disease through whole-genome sequencing. Nat Rev Genet 2010;
11:415–25.
[16] Bird TD. Genetic aspects of Alzheimer disease. Genet Med 2008;
10:231–9.
[17] Schork NJ, Murray SS, Frazer KA, Topol EJ. Common vs. rare allele
hypotheses for complex diseases. Curr Opin Genet Dev 2009;
19:212–9.
[18] Cruchaga C, Karch CM, Jin SC, Benitez BA, Cai Y, Guerreiro R, et al.
Rare coding variants in the phospholipase D3 gene confer risk for Alz-
heimer’s disease. Nature 2014;505:550–4.
[19] Rogaeva E, Meng Y, Lee JH, Gu Y, Kawarai T, Zou F, et al. The
neuronal sortilin-related receptor SORL1 is genetically associated
with Alzheimer disease. Nat Genet 2007;39:168–77.
[20] Reitz C, Cheng R, Rogaeva E, Lee JH, Tokuhiro S, Zou F, et al. Meta-
analysis of the association between variants in SORL1 and Alzheimer
disease. Arch Neurol 2011;68:99–106.
[21] Lehmann DJ, Cortina-Borja M, Warden DR, Smith AD, Sleegers K,
Prince JA, et al. Large meta-analysis establishes the ACE insertion-
deletion polymorphism as a marker of Alzheimer’s disease. Am J Epi-
demiol 2005;162:305–17.
[22] Meng Y, Baldwin CT, Bowirrat A, Waraska K, Inzelberg R,
Friedland RP, et al. Association of polymorphisms in the
Angiotensin-converting enzyme gene with Alzheimer disease in an Is-
raeli Arab community. Am J Hum Genet 2006;78:871–7.
[23] Rogaev EI, Sherrington R, Rogaeva EA, Levesque G, Ikeda M,
Liang Y, et al. Familial Alzheimer’s disease in kindreds with missense
mutations in a gene on chromosome 1 related to the Alzheimer’s dis-
ease type 3 gene. Nature 1995;376:775–8.
[24] Sherrington R, Rogaev EI, Liang Y, Rogaeva EA, Levesque G,
Ikeda M, et al. Cloning of a gene bearing missense mutations
in early-onset familial Alzheimer’s disease. Nature 1995;
375:754–60.
[25] Goate A, Chartier-Harlin MC, Mullan M, Brown J, Crawford F,
Fidani L, et al. Segregation of a missense mutation in the amyloid pre-
cursor protein gene with familial Alzheimer’s disease. Nature 1991;
349:704–6.
[26] R Development Core Team. R: A language and environment for statis-
tical computing. The R Foundation for Statistical Computing; Vienna,
Austria; 2011.
[27] Price AL, Patterson NJ, Plenge RM, Weinblatt ME, Shadick NA,
Reich D. Principal components analysis corrects for stratification in
genome-wide association studies. Nat Genet 2006;38:904–9.
[28] Adzhubei IA, Schmidt S, Peshkin L, Ramensky VE, Gerasimova A,
Bork P, et al. A method and server for predicting damaging missense
mutations. Nat Methods 2010;7:248–9.
[29] Ng PC, Henikoff S. Predicting deleterious amino acid substitutions.
Genome Res 2001;11:863–74.
[30] Li B, Krishnan VG, Mort ME, Xin F, Kamati KK, Cooper DN, et al.
Automated inference of molecular mechanisms of disease from amino
acid substitutions. Bioinformatics 2009;25:2744–50.
[31] Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D,
et al. PLINK: a tool set for whole-genome association and population-
based linkage analyses. Am J Hum Genet 2007;81:559–75.
[32] Wong W, Scott JD. AKAP signalling complexes: focal points in space
and time. Nat Rev Mol Cell Biol 2004;5:959–70.
[33] Lin JW, Wyszynski M, Madhavan R, Sealock R, Kim JU, Sheng M.
Yotiao, a novel protein of neuromuscular junction and brain that inter-
acts with specific splice variants of NMDA receptor subunit NR1. J
Neurosci 1998;18:2017–27.
[34] Chen L, Marquardt ML, Tester DJ, Sampson KJ, Ackerman MJ,
Kass RS. Mutation of an A-kinase-anchoring protein causes long-
QT syndrome. Proc Natl Acad Sci U S A 2007;104:20990–5.
[35] Nie T, McDonough CB, Huang T, Nguyen PV, Abel T. Genetic disrup-
tion of protein kinase A anchoring reveals a role for compartmental-
ized kinase signaling in theta-burst long-term potentiation and
spatial memory. J Neurosci 2007;27:10278–88.
[36] Witczak O, Skalhegg BS, Keryer G, Bornens M, Tasken K, Jahnsen T,
et al. Cloning and characterization of a cDNA encoding an A-kinase
anchoring protein located in the centrosome, AKAP450. Embo J
1999;18:1858–68.
[37] Keryer G, Witczak O, Delouvee A, Kemmner WA, Rouillard D,
Tasken K, et al. Dissociating the centrosomal matrix protein
AKAP450 from centrioles impairs centriole duplication and cell cycle
progression. Mol Biol Cell 2003;14:2436–46.
[38] Keryer G, Di Fiore B, Celati C, Lechtreck KF, Mogensen M,
Delouvee A, et al. Part of Ran is associated with AKAP450 at the
centrosome: involvement in microtubule-organizing activity. Mol
Biol Cell 2003;14:4260–71.
[39] Rivero S, Cardenas J, Bornens M, Rios RM.Microtubule nucleation at
the cis-side of the Golgi apparatus requires AKAP450 and GM130.
Embo J 2009;28:1016–28.
[40] Gillingham AK, Munro S. The PACT domain, a conserved centroso-
mal targeting motif in the coiled-coil proteins AKAP450 and pericen-
trin. EMBO Rep 2000;1:524–9.
[41] Grundke-Iqbal I, Iqbal K, Quinlan M, Tung YC, Zaidi MS,
Wisniewski HM. Microtubule-associated protein tau. A component
of Alzheimer paired helical filaments. J Biol Chem 1986;261:6084–9.
[42] Grundke-Iqbal I, Iqbal K, Tung YC, Quinlan M, Wisniewski HM,
Binder LI. Abnormal phosphorylation of the microtubule-associated
protein tau (tau) in Alzheimer cytoskeletal pathology. Proc Natl
Acad Sci U S A 1986;83:4913–7.
[43] Iqbal K, Grundke-Iqbal I, Zaidi T, Merz PA,Wen GY, Shaikh SS, et al.
Defective brain microtubule assembly in Alzheimer’s disease. Lancet
1986;2:421–6.
[44] Yoshida H, Ihara Y. Tau in paired helical filaments is functionally
distinct from fetal tau: assembly incompetence of paired helical fila-
ment-tau. J Neurochem 1993;61:1183–6.
[45] Cash AD, Aliev G, Siedlak SL, Nunomura A, Fujioka H, Zhu X, et al.
Microtubule reduction in Alzheimer’s disease and aging is indepen-
dent of tau filament formation. Am J Pathol 2003;162:1623–7.

Supplementary online content
Two rare AKAP9 missense variants are associated withAlzheimer disease in African Americans
Supplementary methods
Whole-exome sequencing and quality controlWhole-exome capture was performed using the Agilent
SureSelect Human All Exon kit. Sequencing was performedusing the Illumina GA IIx platform. The alignment of the80 bp single-end reads was completed with the Burrows-Wheeler Aligner algorithm [1,2]. Single nucleotidepolymorphism (SNP) discovery and quality filtering wascompleted using a pipeline based on the Genome AnalysisToolkit [3,4]. Single nucleotide variants (SNVs) meetinginitial quality control (QC) criteria had a minimum depth of10 reads, mapping quality scores of at least 30, and depthquality (QD) scores of at least 5. While these parametersare somewhat liberal, only SNVs observed in more than onesequenced Alzheimer’s disease (AD)-case (whether theyoccurred within a sibling pair) were considered for furtheranalysis to filter out additional possible sequencing artifacts.This strategy likely increased the selection of AD-related var-iants because sequencing was performed in cases only.
GenotypingSNPs from whole-exome sequencing that were selected
for association testing were genotyped using KASP allele-specific PCR-based assays from LGC Genomics (Hoddes-don UK, Beverly MA) and an Applied Biosystems 7900Real-Time PCR System with 384-Well Block Module. Per-formance of the assays was validated by comparing geno-types derived from sequencing and SNP genotyping forthe seven whole-exome sequenced individuals. Subjectswith a call rate of less than 90% were excluded from furtheranalysis, except for analyses using genotypes that wereconfirmed using Sanger sequencing. Methods used togenerate genome-wide tag SNP data used for haplotypeanalysis of the Multi-Institutional Research on AlzheimerGenetic Epidemiology Study (MIRAGE) and Genetic andEnvironmental Risk Factors for Alzheimer Disease amongAfrican Americans Study (GenerAAtions) cohorts inPLINK and PC analysis in EIGENSTRAT are described inLogue et al. 2011 [5].
Sanger sequencingPCRwas performed in a three step cycle repeated 35 times
using 100 mg of DNA and amplification primers using theQiagen Hot Start Tag polymerase. Custom primers were de-signed using PrimerQuest software and supplied by IntegratedDNA technology (see Supplementary Table 3 for details). Ge-notypes for which only the reverse or forward read was avail-able were excluded. The same primers were used for both thePCR and Sanger sequencing reactions. The results of theexperiment were examined using Applied Biosystems’Sequence Scanner Software v1.0. Genotypes from Sanger
sequencing were determined blind to the initial genotype callsand to AD status. Sanger sequencing was performed for allMIRAGE and GenerAAtions subjects which had been identi-fied as having the minor allele at one of these SNPs (n5 24).This included subjects whose genotypes had been excludedfrom other analyses because of a high genotype-missingnessrate. Additionally we genotyped any subject who was pre-dicted to have the background haplotype H0 but for whom ge-notypes were missing or had been excluded due to highmissing rate (n 5 13) and several other control subjects (4)who were identified as having H0 but whose genotype wasthe homozygous reference at each. In total, 41 sampleswere identified to be genotyped using Sanger sequencing.Two subjects had Sanger sequencing which failed for one orboth SNPs and were excluded from further analysis.
Haplotype and bioinformatic analysesHaplotype inference, haplotype frequency estimation, and
haplotype association tests were performed using PLINK (v.1.07, October 2009) [6]. Haplotypes were generated usingboth rare-SNP genotypes determined in this study and geno-types for common SNPs obtained previously for the same in-dividuals [5]. Haplotypes were also generated for subjectsincluded in release 1 of the 1000 Genomes Project usingPLINK [7]. Regional LD was assessed using Haploview [8]employing publically available HapMap2 reference panelsfor the Yoruban (YRI) sample. Phylogenetic dendogramswere generated from of 1000 Genomes Project phased haplo-types using R [9], first by measuring the similarity of the hap-lotypes using the dist function and then by performinghierarchical clustering of the distances using the hclust func-tion. Genomic positions were fixed according to the hg19(February 2009) human genome assembly. The functionalityof associated SNPs was assessed using bioinformatic ap-proaches implemented in software packages includingPolyPhen-2 [10] which is a Bayesian probability functiondeveloped via a machine learning algorithm, SIFTwhich ex-amines conservation of elements across similar sequences toidentify whether mutations are potentially deleterious [11],andMutPred which models the change induced by a mutationby comparing it to the original sequence and produces a prob-ability of a change of structure or function [12].
Replication analysesThe replication data consisted of a subset of the Alz-
heimer Disease Genetics Consortium African-American(AA) sample [13] excluding those subjects from theMIRAGE/GenerAAtions cohort. Specifically, genotypeswere generated for rs144662445 and rs149979685 in avail-able AA samples from the Chicago Health and Aging Project[14,15], the four different sites of the African AmericanAlzheimer’s Disease Genetics Study (Columbia University,University of Miami, North Carolina A&T StateUniversity, and Vanderbilt) [16], a subset of unrelated indi-viduals from the National Institute on Aging Late-OnsetAlzheimer’s Disease study [17], the NIA Alzheimer’s Dis-ease Centers [18], the Mayo Clinic [19], The Mount Sinai
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 618.e1

School of Medicine Brain Bank [20], the Rush UniversityAlzheimer’s disease Center [21–23], the University ofMiami/Vanderbilt University study [24–26], the Universityof Pittsburgh [27], the Washington Heights InwoodColumbia Aging Project [28], and Washington University[29–32]. Genotyping of rs144662445 and rs149979685was performed using TaqMan assays and an AppliedBiosystems 7900 Real-Time PCR System with 384-WellBlockModule. Only subjects for whom both SNPs were suc-cessfully genotyped were included in the analysis. To beconsistent with the discovery analysis, subjects wereexcluded if their age was less than 60 years at diagnosis(cases) or at last examination (controls). Subjects whowere diagnosed with other forms of dementia or mild cogni-tive impairment were also excluded. The final sample sizewas 1037 cases and 1869 controls (Supplementary Table 2).
Supplementary results
Exome sequencing and variant filteringThe success rate for aligning the 80 bp sequences from
the seven subjects was 96.3% with 17.5x mean coveragewithin target regions. Before filtering, 88,867 SNPs wereidentified and the transition:transversion (TI/TV) ratio forthese SNPs was 2.8. A total of 81,335 SNPs after filteringfor quality (read depth .10, QD Score .5, Map qualityscore .30). This number was reduced to 50,904 afterfiltering for those present in at least two subjects. The TI/TV ratio for these SNPs was 2.9, which approximates the ex-pected value for exonic SNPs. Filtering SNPs that were ab-sent in dbSNP build 132 yielded 1364 SNPs. Of these, 431were also non-synonymous. Only one of these SNPs is in agene with replicated association to AD: rs149704584 inCD33. Of the 431 that passed cleaning and filtering criteria,a total of 63 SNPs from 58 genes deemed most likely to befunctionally related to AD were genotyped and tested for as-sociation. Twelve of these SNPs were in genes listed in Alz-Gene. The remainder were from three Ingenuity PathwayAnalysis (IPA)-generated gene networks having genesinvolved in a variety of biological functions includingcellular assembly and organization, nervous system develop-ment and function, carbohydrate metabolism, cell-to-cellsignaling and interaction, hematological system develop-ment and function, renal and urological system developmentand function, and cellular compromise (SupplementaryFigure 3). Forty-six of these variants were identified previ-ously (i.e., had assigned rsIDs in dbSNP build 135 eventhough they were absent according to build 132) and hadpopulation frequencies reported in the NHLBI Grand Op-portunity Exome Sequencing Project. Forty-four of the 63SNPs were successfully genotyped in the cohort of 441 casesand 426 controls.
Generalized Estimating Equations (GEE) modeling of theAKAP9 variants
Because of the numerical instability induced by examina-tion of rare variants in the estimation of logistic regression
models, our initial screening analysis was limited to a Fisher’sexact test for association. However, after this initial screening,we explored the possibility of other sources of confoundingbias. Because the more frequent AKAP9 variantrs144662445 was observed in at least one control in theMIRAGE/GenerAAtions data set, it was numerically possibleto fit logistic GEE models [33,34] for this SNP. GEE allowsfor the inclusion of singletons and family data, is robust tomisspecification of the correlation model, and was sufficientto prevent inflation on a genome-wide investigation of ADin these samples [5]. In a GEE model which included covari-ates adjusting for age, sex, and study (MIRAGE/GenerAA-tions), the P-value for the rs144662445 covariateapproximated the P-value reported in Table 1 which wasnot adjusted for covariates (P 5 .022) and the OR estimatewas nearly identical (OR 5 8.3). Thus, age, non-independence of samples, sex, and study of origin do notappear to be confounding the association with rs144662445.These factors are also not likely confounding the results forrs149979685 because these SNPs are in high LD.
Supplementary References
[1] Li H, Durbin R. Fast and accurate short read alignment with Burrows-
Wheeler transform. Bioinformatics 2009;25:1754–60.
[2] Li H, Durbin R. Fast and accurate long-read alignment with Burrows-
Wheeler transform. Bioinformatics 2010;26:589–95.
[3] McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K,
Kernytsky A, et al. The Genome Analysis Toolkit: a MapReduce
framework for analyzing next-generation DNA sequencing data.
Genome Res 2010;20:1297–303.
[4] DePristo MA, Banks E, Poplin R, Garimella KV, Maguire JR, Hartl C,
et al. A framework for variation discovery and genotyping using next-
generation DNA sequencing data. Nat Genet 2011;43:491–8.
[5] LogueMW, SchuM, Vardarajan BN, Buros J, Green RC, Go RC, et al.
A comprehensive genetic association study of Alzheimer disease in
African Americans. Arch Neurol 2011;68:1569–79.
[6] Purcell S, Neale B, Todd-Brown K, Thomas L, Ferreira MA, Bender D,
et al. PLINK: a tool set for whole-genome association and population-
based linkage analyses. Am J Hum Genet 2007;81:559–75.
[7] The 1000 Genomes Project Consortium. A map of human genome
variation from population-scale sequencing. Nature 2010;
467:1061–73.
[8] Barrett JC, Fry B, Maller J, Daly MJ. Haploview: analysis and visual-
ization of LD and haplotype maps. Bioinformatics 2005;21:263–5.
[9] R Development Core Team. R: A language and environment for statis-
tical computing. The R Foundation for Statistical Computing; Vienna,
Austria; 2011.
[10] Adzhubei IA, Schmidt S, Peshkin L, Ramensky VE, Gerasimova A,
Bork P, et al. A method and server for predicting damaging missense
mutations. Nat Methods 2010;7:248–9.
[11] Ng PC, Henikoff S. Predicting deleterious amino acid substitutions.
Genome Res 2001;11:863–74.
[12] Li B, Krishnan VG, Mort ME, Xin F, Kamati KK, Cooper DN, et al.
Automated inference of molecular mechanisms of disease from amino
acid substitutions. Bioinformatics 2009;25:2744–50.
[13] Reitz C, Jun G, Naj A, Rajbhandary R, Vardarajan BN,Wang LS, et al.
Variants in the ATP-binding cassette transporter (ABCA7), apolipo-
protein E 4,and the risk of late-onset Alzheimer disease in African
Americans. JAMA 2013;309:1483–92.
[14] Bienias JL, Beckett LA, Bennett DA,Wilson RS, Evans DA. Design of
the Chicago Health and Aging Project (CHAP). J Alzheimers Dis
2003;5:349–55.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618618.e2

[15] Evans DA, Bennett DA, Wilson RS, Bienias JL, Morris MC,
Scherr PA, et al. Incidence of Alzheimer disease in a biracial urban
community: relation to apolipoprotein E allele status. Arch Neurol
2003;60:185–9.
[16] Meier IB, Manly JJ, Provenzano FA, Louie KS, Wasserman BT,
Griffith EY, et al. White matter predictors of cognitive functioning
in older adults. J Int Neuropsychol Soc 2012;18:414–27.
[17] Lee JH, Cheng R, Graff-Radford N, Foroud T, Mayeux R. Analyses of
the National Institute on Aging Late-Onset Alzheimer’s Disease Fam-
ily Study: implication of additional loci. Arch Neurol 2008;
65:1518–26.
[18] Jun G, Naj AC, Beecham GW, Wang LS, Buros J, Gallins PJ, et al.
Meta-analysis confirms CR1, CLU, and PICALM as Alzheimer dis-
ease risk loci and reveals interactions with APOE genotypes. Arch
Neurol 2010;67:1473–84.
[19] Carrasquillo MM, Zou F, Pankratz VS, Wilcox SL, Ma L, Walker LP,
et al. Genetic variation in PCDH11X is associated with susceptibility
to late-onset Alzheimer’s disease. Nat Genet 2009;41:192–8.
[20] Haroutunian V, Perl DP, Purohit DP, Marin D, Khan K, Lantz M, et al.
Regional distribution of neuritic plaques in the nondemented elderly
and subjects with very mild Alzheimer disease. Arch Neurol 1998;
55:1185–91.
[21] Bennett DA, Schneider JA, Bienias JL, Evans DA, Wilson RS. Mild
cognitive impairment is related to Alzheimer disease pathology and
cerebral infarctions. Neurology 2005;64:834–41.
[22] Bennett DA, Schneider JA, Buchman AS, Mendes de Leon C,
Bienias JL, Wilson RS. The Rush Memory and Aging Project: study
design and baseline characteristics of the study cohort. Neuroepidemi-
ology 2005;25:163–75.
[23] Bennett DA, Wilson RS, Schneider JA, Evans DA, Beckett LA,
Aggarwal NT, et al. Natural history of mild cognitive impairment in
older persons. Neurology 2002;59:198–205.
[24] Beecham GW, Martin ER, Li YJ, Slifer MA, Gilbert JR, Haines JL,
et al. Genome-wide association study implicates a chromosome 12
risk locus for late-onset Alzheimer disease. Am J Hum Genet 2009;
84:35–43.
[25] Edwards TL, Scott WK, Almonte C, Burt A, Powell EH,
Beecham GW, et al. Genome-wide association study confirms SNPs
in SNCA and the MAPT region as common risk factors for Parkinson
disease. Ann Hum Genet 2010;74:97–109.
[26] Scott WK, Nance MA, Watts RL, Hubble JP, Koller WC, Lyons K,
et al. Complete genomic screen in Parkinson disease: evidence for
multiple genes. JAMA 2001;286:2239–44.
[27] Kamboh MI, Minster RL, Demirci FY, Ganguli M, Dekosky ST,
Lopez OL, et al. Association of CLU and PICALM variants with Alz-
heimer’s disease. Neurobiol Aging 2012;33:518–21.
[28] TangMX, Stern Y,Marder K, Bell K, Gurland B, Lantigua R, et al. The
APOE-epsilon4 allele and the risk of Alzheimer disease among Afri-
can Americans, whites, and Hispanics. JAMA 1998;279:751–5.
[29] Berg L, McKeel DW Jr, Miller JP, Storandt M, Rubin EH, Morris JC,
et al. Clinicopathologic studies in cognitively healthy aging and Alz-
heimer’s disease: relation of histologic markers to dementia severity,
age, sex, and apolipoprotein E genotype. Arch Neurol 1998;
55:326–35.
[30] Morris JC, Roe CM, Xiong C, Fagan AM, Goate AM, Holtzman DM,
et al. APOE predicts amyloid-beta but not tau Alzheimer pathology in
cognitively normal aging. Ann Neurol 2010;67:122–31.
[31] Morris JC, Weintraub S, Chui HC, Cummings J, Decarli C, Ferris S,
et al. The Uniform Data Set (UDS): clinical and cognitive variables
and descriptive data from Alzheimer Disease Centers. Alzheimer
Dis Assoc Disord 2006;20:210–6.
[32] Storandt M, Grant EA, Miller JP, Morris JC. Longitudinal course and
neuropathologic outcomes in original vs revised MCI and in pre-MCI.
Neurology 2006;67:467–73.
[33] Liang KY, Zeger SL. Longitudinal data analysis using linear models.
Biometrika 1986;73:13–22.
[34] Zeger SL, Liang KY. Longitudinal data analysis for discrete and
continuous outcomes. Biometrics 1986;42:121–30.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 618.e3

Supplementary Figure 1. Plots of the first 10 principal components of population substructure in the African American (AA) sample of 436 Alzheimer’s disease
(AD) cases and 424 controls derived by EIGENSTRAT based on all MAF .5% markers from a whole-genome genotyping panel. The color of the plotting
symbols indicates AKAP9 genotype: Black 5 subject lacks rs144662445 and rs149989685 variants; Red 5 subject has rs144662445 variant;
Green 5 subject has both rs144662445 and rs149989685 variants.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618618.e4

Supplementary Figure 2. Linkage disequilibrium (R2) plot of AKAP9 and surrounding region based on HapMap YRI reference panel of SNPs.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 618.e5

Supplementary Figure 3. Networks derived by INGENUITY software containing previously-known Alzheimer’s disease (AD) genes (green) and genes con-
taining non-synonymous SNPs identified through whole-exome sequencing (WES) (red). These networks are characterized by genes with biological functions
related to: (A) cellular assembly and organization, nervous system development and function, and carbohydrate metabolism; (B) cell-to-cell signaling and inter-
action, cellular assembly and organization, and nervous system development and function; (C) hematological system development and function, renal and uro-
logical system development and function, and cellular compromise.
Supplementary Table 1
Characteristics of subjects from the discovery samples included in whole-exome sequencing (WES) and association studies
Group N AD cases Controls Males Females Mean age (SD) APOE ε4 freq
WES subjects 7 7 (100%) 0 (0%) 3 (43%) 4 (57%) 79.4 (5.2) 21%
Analyzed MIRAGE subjects 408 199 (49%) 209 (51%) 121 (30%) 287 (70%) 75.1 (8.1) 34%
Analyzed GenerAAtions subjects 408 223 (55%) 185 (45%) 173 (42%) 235 (58%) 79.5 (6.7) 29%
Total discovery 816 422 (52%) 394 (48%) 294 (36%) 522 (64%) 77.3 (7.5) 32%
Abbreviations: AD, Alzheimer disease; MIRAGE, Multi-Institutional Research on Alzheimer Genetic Epidemiology Study; GenerAAtions, Genetic and
Environmental Risk Factors for Alzheimer Disease among African Americans Study.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618618.e6

Supplementary Table 2
Characteristics of genotyped subjects from the ADGC AA replication cohort by study of origin
Group N AD cases Controls Males Females Mean age (SD)* APOE ε4 freq
ADC 24 4 (17%) 20 (83%) 8 (33%) 16 (67%) 73.6 (5.8) 14%
CHAP 328 177 (54%) 151 (46%) 105 (32%) 223 (68%) 81.7 (6.6) 18%
U. Miami 246 82 (33%) 164 (67%) 63 (26%) 183 (74%) 72.7 (8.4) 26%
Washington U. 165 125 (76%) 40 (24%) 39 (24%) 126 (76%) 75.5 (7.6) 29%
U. Pittsburgh 201 109 (54%) 92 (46%) 55 (27%) 146 (73%) 76.7 (7.4) 30%
AAG - Columbia 242 20 (8%) 222 (92%) 44 (18%) 198 (82%) 69.8 (7.6) 21%
AAG – Miami 199 40 (20%) 159 (80%) 46 (23%) 153 (77%) 71.7 (7.8) 25%
AAG - NC A&T 226 61 (27%) 165 (73%) 68 (30%) 157 (69%) 71.3 (8.6) 27%
AAG – Vandy 124 37 (30%) 87 (70%) 25 (20%) 99 (80%) 72.1 (8.8) 29%
Mayo Clinic 308 80 (26%) 228 (74%) 68 (22%) 240 (78%) 78.2 (7.2) 24%
MSSM 43 29 (67%) 14 (33%) 17 (40%) 26 (60%) 82.7 (10.0) 28%
NIA-LOAD 15 6 (40%) 9 (60%) 1 (7%) 14 (93%) 72.5 (7.0) 25%
ROS/MAP 130 28 (22%) 102 (78%) 54 (42%) 76 (58%) 81.0 (5.3) 18%
Vanderbilt U. 34 9 (26%) 25 (74%) 7 (21%) 27 (79%) 70.4 (6.4) 24%
WHICAP 621 230 (37%) 391 (63%) 159 (26%) 462 (74%) 80.8 (6.4) 19%
Total 2906 1037 (36%) 1869 (64%) 759 (26%) 2146 (74%) 75.3 (8.5) 23%
Abbreviations: AA, African-Americans; AD, Alzheimer disease; ADGC, Alzheimer Disease Genetics Consortium.
*Age at evaluation or, alternately, age at diagnosis/death for samples and entire cohorts (MSSM) where age at evaluation was not available.
Supplementary Table 3
Primers used for Sanger Sequencing of the two AD-associated AKAP9 SNPs
SNP Forward primer Reverse primer
Rs144662445 5-GAG CAC TGTAAA GGA C-3 5-TGACCCCAC CTCATC TTC TCT TAG-3
Rs149979685 5-GTTACT GGG GTT CCA GG-3 5-GAC ATG CTT CAG TGATGG CAC-3
Abbreviation: SNP, single-nucleotide polymorphism.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 618.e7

Supplementary Table 4
Characteristics of the SNPs identified for genotyping
Gene CHR POS rsID
ESPMAF:EA%/AA
%*
Mapping
quality
(MQ)
Read
depth
(DP)
Depth
quality
(QD)
#WES
subjects
w. Alt.
Allele
In
AlzGene Network
Successfully
genotyped
NoC2L 1 888563 rs150127608 0.0116/0.7036 35.55 45 13.45 2 No N1 Yes
NoC2L 1 892380 rs150615968 0.0/0.6809 36.91 212 11.93 2 No N1 No
ISG15 1 949491 rs148041041 0.0116/0.1816 36.84 64 12.58 2 No N3 Yes
AGRN 1 987191 rs139415524 0.0/0.4085 37 37 16.33 2 No N2 Yes
FCRLA 1 161681237 rs144403707 0.0/0.1362 36.84 62 12.01 2 No N3 No
FCRLA 1 161681238 rs144830049 0.0/0.1362 36.84 63 7.35 3 No N3 Yes
ATP2B4 1 203672867 rs145963279 0.0/2.1108 37 141 13.52 2 No N2 Yes
PLX02 1 208215558 rs144005934 0.0/0.0454 37 66 12.07 2 No N2 Yes
AGT 1 230838903 rs143479528 0.0/0.522 37 90 15.6 2 Yes N1 Yes
IFIH1 2 163137881 rs147000317 0.0/0.8171 37 180 14.52 2 No N3 Yes
LRP2 2 170134388 rs145094511 0.0/0.0681 36.96 252 13.03 2 Yes N2 Yes
ITGA9 3 37774225 rs142726080 0.0116/3.2683 37 46 16.67 2 No N2 Yes
SCN5A 3 38646398 NA 0.0/0.0753 37 17 12.72 2 No N2 Yes
NKTR 3 42678946 rs146815110 0.0/2.1335 36.88 340 12.91 2 No N11N3 Yes
C3orf15 3 119456282 NA NA 37 219 18.39 2 No N1 No
LRAT 4 155665876 rs147855559 0.0/0.0681 37 91 14.75 2 Yes N1 Yes
HAND2 4 174448432 rs140861359 0.0/0.2724 37 88 18.8 2 No N1 Yes
RIPK1 6 3105849 NA NA 37 164 16.92 2 No N3 No
GPLD1 6 24436908 rs140754732 0.0/0.1362 37 123 12.76 2 No N1 Yes
TAP2 6 32798457 rs150253319 0.1661/2.184 36.89 91 13.93 2 Yes – Yes
TAP2 6 32802938 rs140654840 0.8859/2.1869 37 71 16.44 2 Yes – Yes
FRK 6 116289888 rs143351471 0.0/0.0681 36.91 214 12.32 2 No N1 No
TNFAIP3 6 138195991 rs146534657 0.1163/0.0908 36.91 109 18.32 2 No N3 Yes
GRM1 6 146755454 rs144944927 0.0/2.5658 36.88 81 13.15 2 No N1 Yes
GLI3 7 42007460 rs146130351 0.0/1.4299 37 177 15.72 2 No N1 Yes
BLVRA 7 43832373 rs142897365 0.0116/0.7717 37 77 14.2 2 No N1 No
SEMA3A 7 83823846 rs139951141 0.0/0.3404 36.89 283 12.47 2 No N2 No
AKAP9 7 91709085 rs144662445 0.0/0.4312 36.92 131 13.06 2 No N1 Yes
AKAP9 7 91732110 rs149979685 0.0/0.3631 37 166 12.71 2 No N1 Yes
TRIP6 7 100466396 NA 0.0122/1.5259 37 21 15.15 2 No N3 Yes
TRIB1 8 126448615 rs142512119 0.0/0.1362 37 176 14.05 2 No N2 No
IF07 9 21202136 rs145794215 0.0116/2.6555 36.27 165 17.12 2 No N3 No
MUSK 9 113530236 NA 0.0/0.1791 36.93 151 14.9 2 No N2 Yes
RALGDS 9 135985773 rs140586035 0.0116/1.793 36.85 66 10.9 2 No N1 Yes
ALOX5 10 45938883 NA NA 37 13 8.77 2 Yes – No
TACR2 10 71167027 rs142415572 0.0116/5.9691 36.82 56 19.81 2 Yes – Yes
SORCS3 10 106974224 NA 0.0/0.4539 36.94 177 12.15 2 Yes – Yes
KNDC1 10 135015178 rs141463252 0.0/0.3404 36.68 32 11.99 2 No N1 Yes
LRP4 11 46911956 rs138878258 0.1279/0.4771 37 57 12.14 2 No N2 Yes
STX3 11 59562950 rs142968661 0.0466/2.3171 36.84 63 9.34 2 No N1 No
BAD 11 64051762 NA NA 37 115 11.17 2 No N2 Yes
SPTBN2 11 66475201 NA NA 37 67 7 2 No N1 Yes
CEP290 12 88512485 rs147371999 0.0/1.8904 37 102 12.8 2 No N2 Yes
ACACB 12 109660694 rs146002202 0.0116/0.8171 37 19 27.8 2 No N2 No
ACACB 12 109683516 rs145398869 0.0/0.0908 36.91 107 13.17 2 No N2 Yes
ATP2A2 12 110784163 NA 0.0/0.0227 37 64 22.08 2 No N2 No
KL 13 33591385 NA 0.0/0.1235 37 13 22.79 2 No N2 Yes
KIF26A 14 104642981 NA 0.0/0.5185 37 14 27.38 2 No N1 No
HERC2 15 28518046 rs150363648 NA 35.25 51 7.8 5 No N2 No
MEF2A 15 100211761 NA NA 36.81 52 5.49 2 Yes N2 No
UBN1 16 4924905 rs146192777 0.0465/0.751 37 175 11.65 2 No N1 No
PELP1 17 4579700 rs147186006 0.012/1.5853 37 59 17.67 2 No N1 Yes
MYH13 17 10247309 rs144732640 0.0116/0.8852 35.61 165 13.12 2 Yes – Yes
NoS2 17 26110055 rs149623743 0.0/0.0681 36.9 96 18.23 2 No N3 Yes
IGFBP4 17 38612790 NA NA 37 64 13.24 2 No N2 No
STXBP2 19 7712054 rs150174842 0.0117/0.7976 37 39 23.82 2 No N1 Yes
MIA 19 41282931 rs144785745 0.0/0.227 36.79 49 18.95 2 No N2 No
XRCC1 19 44047554 NA 0.0349/0.0 37 48 10.24 2 Yes – Yes
CD33 19 51742799 rs149704584 0.0/0.0908 36.73 111 13.69 2 Yes N11N3 Yes
(Continued )
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618618.e8

Supplementary Table 4
Characteristics of the SNPs identified for genotyping (Continued )
Gene CHR POS rsID
ESPMAF:EA%/AA
%*
Mapping
quality
(MQ)
Read
depth
(DP)
Depth
quality
(QD)
#WES
subjects
w. Alt.
Allele
In
AlzGene Network
Successfully
genotyped
APOBEC3B 22 39382414 rs146055882 0.0467/13.1256 33.28 222 13.66 3 No N2 No
CBX7 22 39530663 NA NA 37 10 12.27 2 No N1 No
SBF1 22 50903029 rs149528827 0.0/2.1199 37 50 13.21 2 No N1 No
CNGA2 X 150912171 rs150539917 0.0297/9.2568 37 116 14.1 2 No N1 Yes
Abbreviations: AA, African-Americans; SNP, single-nucleotide polymorphism; CHR, chromosome; POS, position; MAF, minor allele frequency; EA, Eu-
ropean American.
*Allele frequency from NHLBI Grand Opportunity Exome Sequencing Project; (ESP) ESP6500 release/percentage in European Americans/African Amer-
icans in the ESP database; Mapping Quality (MQ), Read Depth (DP), and Depth Quality (QD) as reported by the Genome Analysis Toolkit (GATK); NA: SNP
not present in dbSNP v. 135 or in ESP.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 618.e9

Supplementary Table 5
Association results for all genotyped SNPs including allele counts comparing all cases to all controls
Gene CHR POS rsID
HWE
P-value
# Case min.
alleles
# Case maj.
alleles
# Control min.
alleles
# Control maj
alleles
Fisher’s
exact P
NoC2L 1 888563 rs150127608 1 5 819 3 773 .396
ISG15 1 949491 rs148041041 1 0 832 0 784 1
AGRN 1 987191 rs139415524 1 2 824 4 776 .904
FCRLA 1 161681238 rs144830049 1 5 819 3 777 .393
ATP2B4 1 203672867 rs145963279 1 19 811 10 768 .092
PLX02 1 208215558 rs144005934 1 1 831 0 782 .5154
AGT 1 230838903 rs143479528 1 3 827 3 781 .683
IFIH1 2 163137881 rs147000317 1 7 819 4 778 .305
LRP2 2 170134388 rs145094511 1 0 830 0 784 1
ITGA9 3 37774225 rs142726080 1 26 804 18 760 .197
SCN5A 3 38646398 NA 1 0 828 2 782 1
NKTR 3 42678946 rs146815110 .26 9 823 21 761 .995
LRAT 4 155665876 rs147855559 1 0 832 0 782 1
HAND2 4 174448432 rs140861359 1 2 828 1 783 .521
GPLD1 6 24436908 rs140754732 1 1 831 1 777 .767
TAP2 6 32798457 rs150253319 1 19 811 14 770 .296
TAP2 6 32802938 rs140654840 .33 15 817 18 764 .812
TNFAIP3 6 138195991 rs146534657 1 0 832 0 782 1
GRM1 6 146755454 rs144944927 1 14 816 21 761 .939
GLI3 7 42007460 rs146130351 1 14 818 18 760 .861
AKAP9 7 91709085 rs144662445 1 9 835 1 785 .014
AKAP9 7 91732110 rs149979685 1 5 837 0 788 .0366
TRIP6 7 100466396 NA 1 18 810 15 767 .427
MUSK 9 113530236 NA 1 2 830 0 784 .265
RALGDS 9 135985773 rs140586035 1 15 807 13 763 .486
TACR2 10 71167027 rs142415572 .43 61 771 51 731 .294
SORCS3 10 106974224 NA 1 3 825 2 780 .527
KNDC1 10 135015178 rs141463252 1 6 824 3 777 .285
LRP4 11 46911956 rs138878258 1 6 826 2 778 .166
STX3 11 59562950 rs142968661 .67 18 818 17 711 .661
BAD 11 64051762 NA 1 0 832 0 784 1
SPTBN2 11 66475201 NA 1 0 832 0 780 1
CEP290 12 88512485 rs147371999 1 13 819 16 768 .819
ACACB 12 109683516 rs145398869 1 0 832 1 783 1
KL 13 33591385 NA 1 4 824 5 779 .773
KIF26A 14 104642981 NA .85 3 745 7 653 .965
PELP1 17 4579700 rs147186006 1 12 820 19 763 .948
MYH13 17 10247309 rs144732640 1 7 825 10 774 .864
NoS2 17 26110055 rs149623743 1 3 829 2 782 .528
IGFBP4 17 38612790 NA 1 0 838 0 722 1
STXBP2 19 7712054 rs150174842 1 6 824 6 776 .653
XRCC1 19 44047554 NA 1 0 832 0 782 1
CD33 19 51742799 rs149704584 1 1 827 0 782 .514
CNGA2 X 150912171 rs150539917 .21 74 756 67 709 .456
Abbreviations: SNP, single-nucleotide polymorphism; CHR, chromosome; POS, position; HWE, Hardy-Weinberg Equilibrium.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618618.e10

Supplementary Table 6
Characteristics of 13 AA Subjects in the MIRAGE/GenerAAtions sample with at least one AKAP9 variant in rs144662445 and rs149989685
Study Whole-exome sequenced Sex APOE Age* AD status rs144662445 rs149979685
GenerAAtions No Female 34 75 Control AG CC
MIRAGE No Female 44 55y Control AG CT
MIRAGE No Female 33 86 Case AG CC
MIRAGE No Male 23 74 Case AG CC
MIRAGE No Female 34 82 Case AG CC
GenerAAtions No Female 33 88 Case AG CC
MIRAGE No Female 33 81 Case AG CT
MIRAGE Yes Female 34 82 Case AG CT
MIRAGE Yes Female 44 84 Case AG CT
MIRAGE No Male 33 78 Case AG CT
GenerAAtions No Male 33 64 Case AG CT
GenerAAtions No Female 33 71 Case AG CT
GenerAAtions No Female 23 91 Case AG CT
Abbreviations: AA, African-Americans; MIRAGE, Multi-Institutional Research on Alzheimer Genetic Epidemiology Study; GenerAAtions, Genetic and
Environmental Risk Factors for Alzheimer Disease among African Americans Study; SNP, single-nucleotide polymorphism.
*Age at assessment.yExcluded from analysis due to age ,60 at time of assessment.
Supplementary Table 7
Estimated frequencies of .1% MAFAKAP9 haplotypes based on SNPs
from GWAS genotyping
Haplotype* Frequency
AGAGGAGAAC 0.425
GAAAAGAGGA 0.218
GGCAGGAGGA 0.129
GGAAGGAGGA 0.118
GGAAGGAAGCy 0.052
GGAGGAGAAC 0.045
Abbreviations: MAF, minor allele frequency; GWAS, Genome-wide as-
sociation study; SNP, single-nucleotide polymorphism.
*Alleles for SNPs rs1859037, rs2299233, rs2282973, rs6465347,
rs2158138, rs733957, rs2079082, rs13239875, rs4265, and rs1063243,
respectively.yHaplotype HAP0 containing rare variants at rs144662445 and
rs149989685.
M.W. Logue et al. / Alzheimer’s & Dementia 10 (2014) 609-618 618.e11