
MORPHOMETRY OF THE URSUS SPELAEUSREMAINS FROM … · Geo.Alp,Vol.2,S.115–126,2005 MORPHOMETRY OF...
12
Geo.Alp, Vol. 2, S. 115–126, 2005 MORPHOMETRY OF THE URSUS SPELAEUS REMAINS FROM VALSTRONA (NORTHERN ITALY) Alessandro de Carlis 1 , Enrico Alluvione 1 , Alessandro Fonte 1 , Mario Rossi 2 & Giuseppe Santi 1 With 14 figures and 2 plates Abstract Morphometric analyses on cave bear fossils of the Valstrona-Valsesia region (Piedmont, Northern Italy) (in particular from the Delle Streghe Cave), allow the distinction of at least two or three populations of Ursus spelaeus Rosenmüller, 1794, of different sizes. Elements of smaller size are likely to be found in the Buco dell’Orso Cave (Laglio, Como province, Lombardy), and in the older strata of the Grotta Sopra Fontana Marella –GSFM- (Varese province). Differences in size can be linked to the chronological position of the fossils stud- ied: in fact, fossils of smaller dimensions should be chronologically older, but can also be linked to climatic and thermoregulation factors. The increase of size could represent a response to a cooler climate. An alter- native hypothesis associates this reduction of size to the cooler climatic shift. Observations on the Delle Streghe Cave fossils indicate that they are very similar to those inferred for the GSFM population, linking this trend to climatic variation. The reason for an increase in size could also be linked to the rapid evolution of the cave bears and the Delle Streghe fossils should cover a chronological range approximately similar to the fossils from GSFM. 115 1 Dipartimento di Scienze della Terra, Via Ferrata 1, I-27100 Pavia (Italy); e-mail: [email protected] (corresponding author) 2 Museo Civico di Storia Naturale, Lungadige Porta Vittoria 9, I-37129 Verona (Italy) Introduction During the Late Pleistocene Ursus spelaeus Rosenmüller, 1794 (Rosendhal & Kempe, 2004) col- onized a large part of Europe, migrating to the south (central Italy), towards the west (Spain), and towards the north-west (Great Britain) exhibiting the most numerous morphological and morphome- trical varieties. Several studies concerning the cave bear group, Ursus spelaeus and its ancestor Ursus deningeri Von Reichenau, 1906 and U. arctos species coeval to U. spelaeus (Rabeder, 1999; Weinstock, 1999; Perego et al., 2001; Rossi & Santi, 2001 a, b; Santi & Rossi, 2001; Santi et al., 2003 and others) have allowed several hypotheses to be advanced about the relationships between cave bears, U. deningeri and coeval species, and about possible phyletic lines indicated by the fossiliferous record and by recent mtDNA examination (Hofreiter et al., 2002, 2004). Currently, radiometric data is only available for a few caves in Northern Italy: the best known example is the Grotta Sopra Fontana Marella (Varese Province, Lombardy). For the other caves (i.e. the Caverna Generosa, Varese Province) the data are still incomplete (Bona, 2004). Radiometric data and aminoacid racemization of the bear bones from Grotta Sopra Fontana Marella provide the following ages: sample FM4 over 26000 years BP, FM2, 22310 ±200 years BP and FM1 21810±200 years BP (Perego et al., 2001). The wealth of discoveries in various caves in Northern Italy have shed new light on the distribution of the
Transcript of MORPHOMETRY OF THE URSUS SPELAEUSREMAINS FROM … · Geo.Alp,Vol.2,S.115–126,2005 MORPHOMETRY OF...

Geo.Alp, Vol. 2, S. 115–126, 2005
MMOORRPPHHOOMMEETTRRYY OOFF TTHHEE UURRSSUUSS SSPPEELLAAEEUUSS RREEMMAAIINNSS FFRROOMM VVAALLSSTTRROONNAA ((NNOORRTTHHEERRNN IITTAALLYY))
Alessandro de Carlis1, Enrico Alluvione1, Alessandro Fonte1, Mario Rossi2 & Giuseppe Santi1
With 14 figures and 2 plates
AAbbssttrraaccttMorphometric analyses on cave bear fossils of the Valstrona-Valsesia region (Piedmont, Northern Italy) (in
particular from the Delle Streghe Cave), allow the distinction of at least two or three populations of Ursusspelaeus Rosenmüller, 1794, of different sizes. Elements of smaller size are likely to be found in the Bucodell’Orso Cave (Laglio, Como province, Lombardy), and in the older strata of the Grotta Sopra Fontana Marella–GSFM- (Varese province). Differences in size can be linked to the chronological position of the fossils stud-ied: in fact, fossils of smaller dimensions should be chronologically older, but can also be linked to climaticand thermoregulation factors. The increase of size could represent a response to a cooler climate. An alter-native hypothesis associates this reduction of size to the cooler climatic shift. Observations on the DelleStreghe Cave fossils indicate that they are very similar to those inferred for the GSFM population, linkingthis trend to climatic variation. The reason for an increase in size could also be linked to the rapid evolutionof the cave bears and the Delle Streghe fossils should cover a chronological range approximately similar tothe fossils from GSFM.
115
1 Dipartimento di Scienze della Terra, Via Ferrata 1, I-27100 Pavia (Italy); e-mail: [email protected] (corresponding author)2 Museo Civico di Storia Naturale, Lungadige Porta Vittoria 9, I-37129 Verona (Italy)
IInnttrroodduuccttiioonn
During the Late Pleistocene Ursus spelaeusRosenmüller, 1794 (Rosendhal & Kempe, 2004) col-onized a large part of Europe, migrating to thesouth (central Italy), towards the west (Spain), andtowards the north-west (Great Britain) exhibitingthe most numerous morphological and morphome-trical varieties. Several studies concerning the cavebear group, Ursus spelaeus and its ancestor Ursusdeningeri Von Reichenau, 1906 and U. arctosspecies coeval to U. spelaeus (Rabeder, 1999;Weinstock, 1999; Perego et al., 2001; Rossi & Santi,2001 a, b;; Santi & Rossi, 2001; Santi et al., 2003 andothers) have allowed several hypotheses to beadvanced about the relationships between cave
bears, U. deningeri and coeval species, and aboutpossible phyletic lines indicated by the fossiliferousrecord and by recent mtDNA examination(Hofreiter et al., 2002, 2004). Currently, radiometricdata is only available for a few caves in NorthernItaly: the best known example is the Grotta SopraFontana Marella (Varese Province, Lombardy). Forthe other caves (i.e. the Caverna Generosa, VareseProvince) the data are still incomplete (Bona, 2004).Radiometric data and aminoacid racemization ofthe bear bones from Grotta Sopra Fontana Marellaprovide the following ages: sample FM4 over 26000years BP, FM2, 22310 ±200 years BP and FM121810±200 years BP (Perego et al., 2001). Thewealth of discoveries in various caves in NorthernItaly have shed new light on the distribution of the

vertebrates in this area. In some zones research hasonly just started; one such example is the Valsesia-Valstrona (Piedmont) area (Fig. 1). The aim of thispaper is to summarize previous results regardingthese cave bear populations.
GGeeooggrraapphhiiccaall--ggeeoollooggiiccaall ffrraammee ooff tthhee ssttuuddiieedd aarreeaa
Valstrona is a narrow valley with a V-shaped pro-file in its lower reaches while at its head, near Cimadi Capezzone-Punta del Pizzo (2240 m)-Puntad’Issola (2146 m), it enlarges into a wide cirque. Itwinds for 20 km to Omegna village where itdebouches onto the Orta Lake (Cusio). Near theSambughetto village some caves have formed viakarst processes within the lens of the “MarmoValstrona” formation; this lenticular body is interca-lated between gneisses and micaschists of the “SerieKinzigitico-sillimanitica”. Inside the caves the osteo-logical material, accompanied by yellow loessic clay,collects in the lower parts along the side lanes andcavities. This sediment is frequently covered by hardstalagmitic soil (about 15-20 cm thick), and by greymicaceous and sterile sands interspersed with small-er gravel of more recent age linked to the pluvialwashing away phase. To ensure good preservation
of bones, deposits were only slightly dis-turbed and analysed in situ.
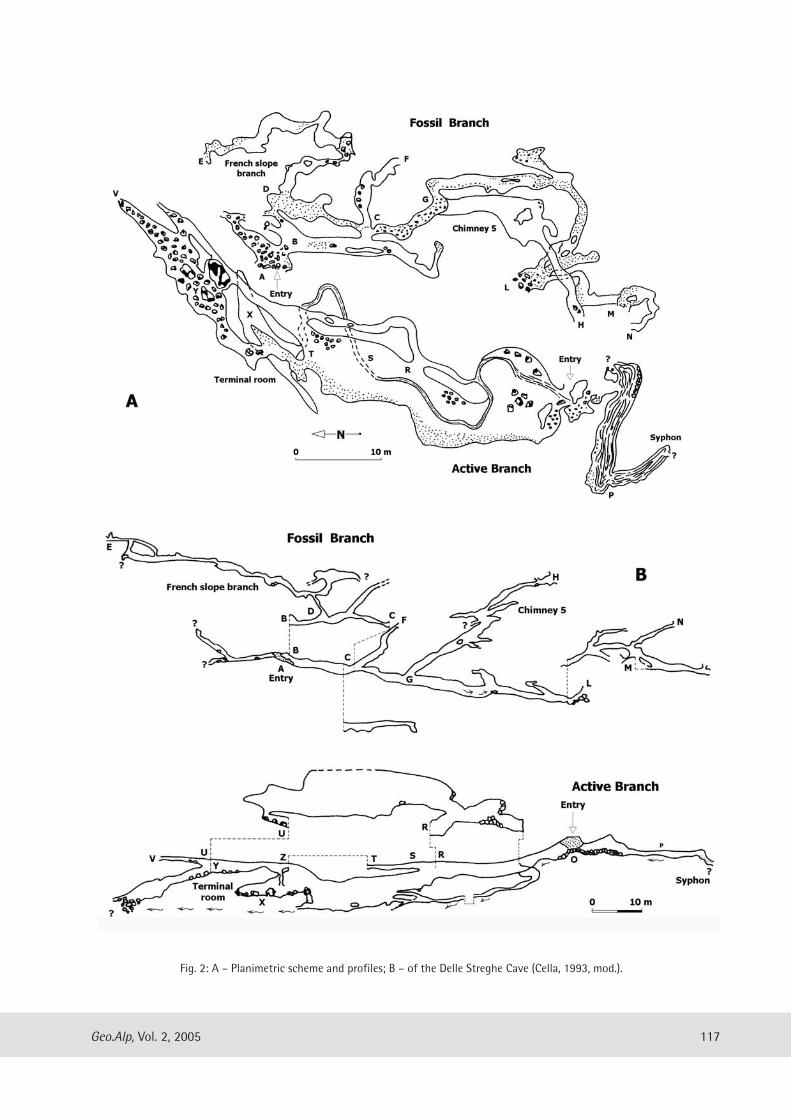
Fossils from Valstrona have beenfound inside the caves known asComplesso dell’Intaglio and Cavernadelle Streghe, near the Cava Sam -bughetto village. The first of these cavesopens out in the upper part of the mar-ble quarry (“Sass Muiè”), it has fiveentrances and a subcircular small gallerycomplex correlated with an older level ofthe water-bearing stratum. The secondcave, called Caverna delle Streghe, is thewidest cave in Verbania Province. It iscomposed of a fossil branch presentlyfoliated by water and by a second activebranch in the marble eroded by the river(Fig. 2). The water source is from theChignolo stream that, after havingcrossed the cave and swelled water fromother tributaries, re-emerges in theStrona River.
The Valsesia fossils are derived from theMt. Fenera (Fig. 3) caves and mainly from the“Ciutarun” and the “Ciota Ciara” cave. The former issituated at 650 m asl, with a large ogival entrance,and it is 55 m long and up to 13 m high. The “CiotaCiara” is located at an altitude of 685 m asl, it is 57m long and the difference in levels internally is upto 18 m. There are two entrances: a southern, nat-ural and a northwestern entrance which wasformed by the collapse of a part of the vault. Thiscave rises upward from SE-NW and ends towardsthe N (Strobino, 1981).
MMaatteerriiaallss aanndd mmeetthhooddss
About one thousand Ursus remains currentlystored in the Museo Civico di Storia Naturale diMilano have been analysed. They have been labelled“MMCCSSNNMM VV”, (abbrevation of MMuseo CCivico di SStoriaNNaturale and VVertebrate), followed by a progressivenumber. A substantial portion of the skeletons ofcubs, juveniles and adult elements is represented(Pls. 1-2). The material is rarely complete, especiallythe skull remains, and in particular in the case ofcubs only skull-caps have been preserved.Preservation is generally good, although some
116 Geo.Alp, Vol. 2, 2005
Fig. 1: Geographic position of the Delle Streghe Cave (Valstrona, Piedmont,Northern Italy).
117Geo.Alp, Vol. 2, 2005
Fig. 2: A – Planimetric scheme and profiles; B – of the Delle Streghe Cave (Cella, 1993, mod.).

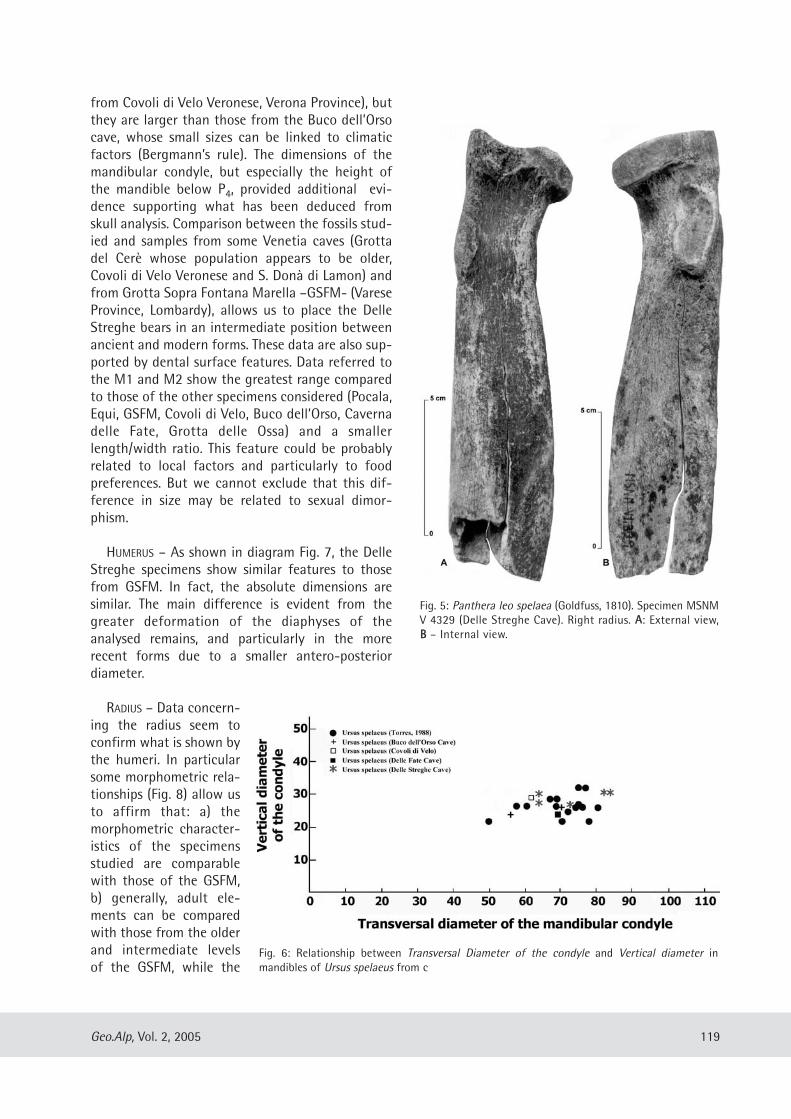
traces of erosion can be found in the proximal anddistal ends of limb bones. In addition, some speci-mens showed traces of pathologies (e.g. periarthri-tis and pesudoarthrosis) and generic malformations,traces of predator activities (Fig. 4). The presence ofpredators is indicated by the catlike remains insidethe Delle Streghe fauna with an incomplete rightradius fragment (MSNM V4329) belonging to
Panthera leo spelaea (Goldfuss, 1810) (Fig. 5). Mostof the fossils belong to Ursus spelaeus Rosenmüller,1794, while others with disputed morphologicalfeatures could be classified as Ursus deningeri VonReichenau 1906. However we have considered theseremains as U. spelaeus on the basis of the broadermorphological relationships within this species.Useful morphometric parameters were deducedfrom Hue (1908), Von den Driesch (1976) and Torres(1988).
MMoorrpphhoommeettrryy
SKULL – These fossils, although incomplete, havesome morphometric features that seem to be typi-cal of cave bears. They are generally similar in sizeto examples of U. spelaeus from caves in Spain andslightly larger than those from Caverna delle Ossa(Zandobbio, Bergamo Province, North of Milan).
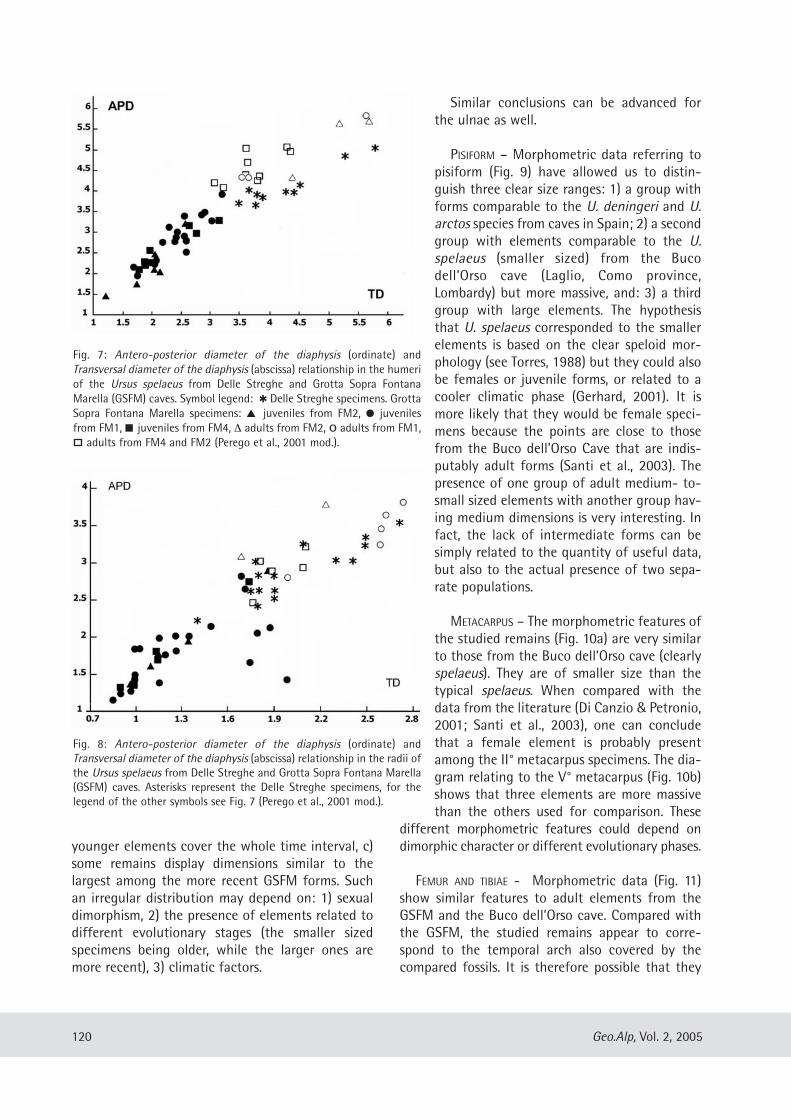
MANDIBLE – The relationship between the trans-versal diameter of the condyle and the verticaldiameter (Fig. 6) confirms what has been inferredregarding skull morphometric analysis. TheSambughetto specimens are similar in size to thetypical spelaeus (in this paper represented by fossils
118 Geo.Alp, Vol. 2, 2005
Fig. 3: Distribution of the main caves in the Fenera Mt. (Valsesia, Piedmont, Northern Italy). Number 1 is the “Ciutarun”, 2 and 3refer to the “Ciota Ciara”. (Strobino, 1981, mod.).
Fig. 4: AA.. Pathological Ursus bone (specimen MSNM V 4362,Delle Streghe Cave). BB.. Predatory activity traces (specimenMSNM V 4097, Delle Streghe Cave).
from Covoli di Velo Veronese, Verona Province), butthey are larger than those from the Buco dell’Orsocave, whose small sizes can be linked to climaticfactors (Bergmann’s rule). The dimensions of themandibular condyle, but especially the height ofthe mandible below P4, provided additional evi-dence supporting what has been deduced fromskull analysis. Comparison between the fossils stud-ied and samples from some Venetia caves (Grottadel Cerè whose population appears to be older,Covoli di Velo Veronese and S. Donà di Lamon) andfrom Grotta Sopra Fontana Marella –GSFM- (VareseProvince, Lombardy), allows us to place the DelleStreghe bears in an intermediate position betweenancient and modern forms. These data are also sup-ported by dental surface features. Data referred tothe M1 and M2 show the greatest range comparedto those of the other specimens considered (Pocala,Equi, GSFM, Covoli di Velo, Buco dell’Orso, Cavernadelle Fate, Grotta delle Ossa) and a smallerlength/width ratio. This feature could be probablyrelated to local factors and particularly to foodpreferences. But we cannot exclude that this dif-ference in size may be related to sexual dimor-phism.
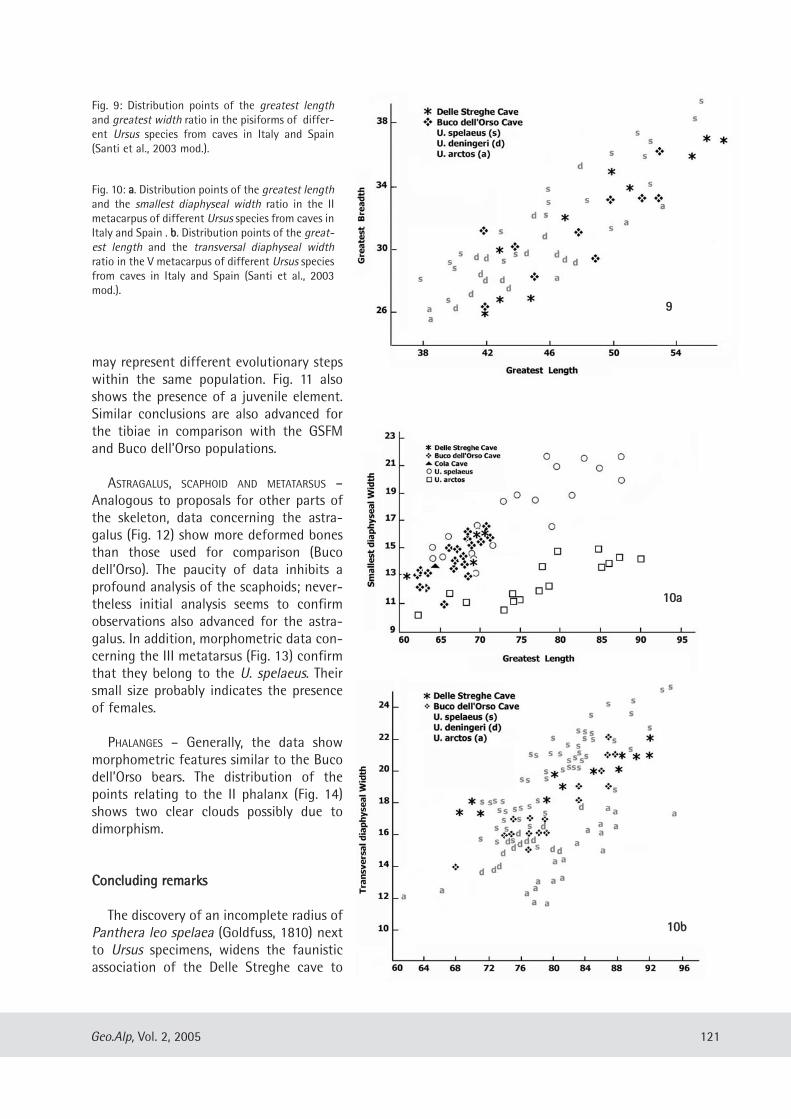
HUMERUS – As shown in diagram Fig. 7, the DelleStreghe specimens show similar features to thosefrom GSFM. In fact, the absolute dimensions aresimilar. The main difference is evident from thegreater deformation of the diaphyses of theanalysed remains, and particularly in the morerecent forms due to a smaller antero-posteriordiameter.
RADIUS – Data concern-ing the radius seem toconfirm what is shown bythe humeri. In particularsome morphometric rela-tionships (Fig. 8) allow usto affirm that: a) themorphometric character-istics of the specimensstudied are comparablewith those of the GSFM,b) generally, adult ele-ments can be comparedwith those from the olderand intermediate levelsof the GSFM, while the
119Geo.Alp, Vol. 2, 2005
Fig. 5: Panthera leo spelaea (Goldfuss, 1810). Specimen MSNMV 4329 (Delle Streghe Cave). Right radius. AA: External view,BB – Internal view.
Fig. 6: Relationship between Transversal Diameter of the condyle and Vertical diameter inmandibles of Ursus spelaeus from c
younger elements cover the whole time interval, c)some remains display dimensions similar to thelargest among the more recent GSFM forms. Suchan irregular distribution may depend on: 1) sexualdimorphism, 2) the presence of elements related todifferent evolutionary stages (the smaller sizedspecimens being older, while the larger ones aremore recent), 3) climatic factors.
Similar conclusions can be advanced forthe ulnae as well.
PISIFORM – Morphometric data referring topisiform (Fig. 9) have allowed us to distin-guish three clear size ranges: 1) a group withforms comparable to the U. deningeri and U.arctos species from caves in Spain; 2) a secondgroup with elements comparable to the U.spelaeus (smaller sized) from the Bucodell’Orso cave (Laglio, Como province,Lombardy) but more massive, and: 3) a thirdgroup with large elements. The hypothesisthat U. spelaeus corresponded to the smallerelements is based on the clear speloid mor-phology (see Torres, 1988) but they could alsobe females or juvenile forms, or related to acooler climatic phase (Gerhard, 2001). It ismore likely that they would be female speci-mens because the points are close to thosefrom the Buco dell’Orso Cave that are indis-putably adult forms (Santi et al., 2003). Thepresence of one group of adult medium- to-small sized elements with another group hav-ing medium dimensions is very interesting. Infact, the lack of intermediate forms can besimply related to the quantity of useful data,but also to the actual presence of two sepa-rate populations.
METACARPUS – The morphometric features ofthe studied remains (Fig. 10a) are very similarto those from the Buco dell’Orso cave (clearlyspelaeus). They are of smaller size than thetypical spelaeus. When compared with thedata from the literature (Di Canzio & Petronio,2001; Santi et al., 2003), one can concludethat a female element is probably presentamong the II° metacarpus specimens. The dia-gram relating to the V° metacarpus (Fig. 10b)shows that three elements are more massivethan the others used for comparison. These
different morphometric features could depend ondimorphic character or different evolutionary phases.
FEMUR AND TIBIAE - Morphometric data (Fig. 11)show similar features to adult elements from theGSFM and the Buco dell’Orso cave. Compared withthe GSFM, the studied remains appear to corre-spond to the temporal arch also covered by thecompared fossils. It is therefore possible that they
120 Geo.Alp, Vol. 2, 2005
Fig. 7: Antero-posterior diameter of the diaphysis (ordinate) andTransversal diameter of the diaphysis (abscissa) relationship in the humeriof the Ursus spelaeus from Delle Streghe and Grotta Sopra FontanaMarella (GSFM) caves. Symbol legend: � Delle Streghe specimens. GrottaSopra Fontana Marella specimens: � juveniles from FM2, � juvenilesfrom FM1, � juveniles from FM4, ∆ adults from FM2, o adults from FM1,
adults from FM4 and FM2 (Perego et al., 2001 mod.).
Fig. 8: Antero-posterior diameter of the diaphysis (ordinate) andTransversal diameter of the diaphysis (abscissa) relationship in the radii ofthe Ursus spelaeus from Delle Streghe and Grotta Sopra Fontana Marella(GSFM) caves. Asterisks represent the Delle Streghe specimens, for thelegend of the other symbols see Fig. 7 (Perego et al., 2001 mod.).
may represent different evolutionary stepswithin the same population. Fig. 11 alsoshows the presence of a juvenile element.Similar conclusions are also advanced forthe tibiae in comparison with the GSFMand Buco dell’Orso populations.
ASTRAGALUS, SCAPHOID AND METATARSUS –Analogous to proposals for other parts ofthe skeleton, data concerning the astra-galus (Fig. 12) show more deformed bonesthan those used for comparison (Bucodell’Orso). The paucity of data inhibits aprofound analysis of the scaphoids; never-theless initial analysis seems to confirmobservations also advanced for the astra-galus. In addition, morphometric data con-cerning the III metatarsus (Fig. 13) confirmthat they belong to the U. spelaeus. Theirsmall size probably indicates the presenceof females.
PHALANGES – Generally, the data showmorphometric features similar to the Bucodell’Orso bears. The distribution of thepoints relating to the II phalanx (Fig. 14)shows two clear clouds possibly due todimorphism.
CCoonncclluuddiinngg rreemmaarrkkss
The discovery of an incomplete radius ofPanthera leo spelaea (Goldfuss, 1810) nextto Ursus specimens, widens the faunisticassociation of the Delle Streghe cave to
121Geo.Alp, Vol. 2, 2005
Fig. 9: Distribution points of the greatest lengthand greatest width ratio in the pisiforms of differ-ent Ursus species from caves in Italy and Spain(Santi et al., 2003 mod.).
Fig. 10: aa. Distribution points of the greatest lengthand the smallest diaphyseal width ratio in the IImetacarpus of different Ursus species from caves inItaly and Spain . bb. Distribution points of the great-est length and the transversal diaphyseal widthratio in the V metacarpus of different Ursus speciesfrom caves in Italy and Spain (Santi et al., 2003mod.).
99
1100aa
1100bb
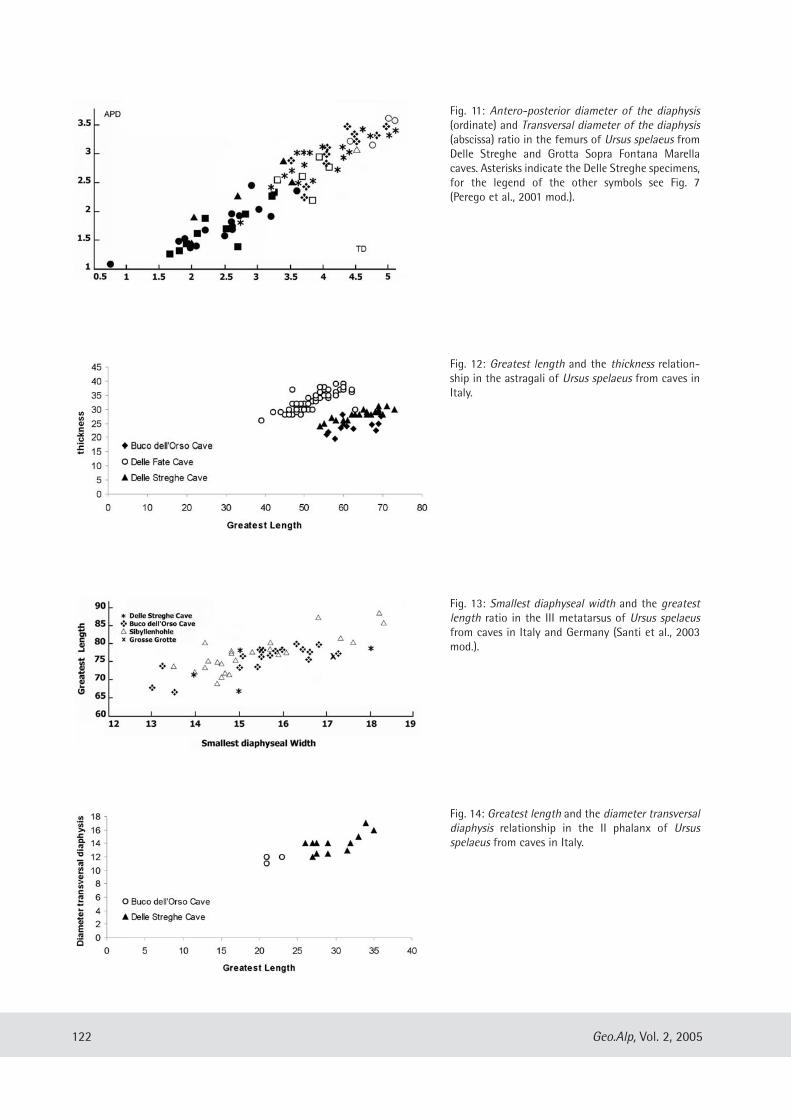
122 Geo.Alp, Vol. 2, 2005
Fig. 11: Antero-posterior diameter of the diaphysis(ordinate) and Transversal diameter of the diaphysis(abscissa) ratio in the femurs of Ursus spelaeus fromDelle Streghe and Grotta Sopra Fontana Marellacaves. Asterisks indicate the Delle Streghe specimens,for the legend of the other symbols see Fig. 7(Perego et al., 2001 mod.).
Fig. 13: Smallest diaphyseal width and the greatestlength ratio in the III metatarsus of Ursus spelaeusfrom caves in Italy and Germany (Santi et al., 2003mod.).
Fig. 14: Greatest length and the diameter transversaldiaphysis relationship in the II phalanx of Ursusspelaeus from caves in Italy.
Fig. 12: Greatest length and the thickness relation-ship in the astragali of Ursus spelaeus from caves inItaly.

other nearby caves (Buco dell’Orso Cave, Delle OssaCave – Zandobbio in Bergamo Province).Pathologies are rare, mainly confined to limbs, andrelated to the senescence of the bears.Morphometric data indicate the presence of at leasttwo populations of cave bears characterized by dif-ferent sizes: the small-size bears are comparable tothe Buco dell’Orso cave bears and those specimensfrom the older levels to the Grotta Sopra FontanaMarella. According to Perego et al. (2001), the dif-ference in size is related to a different evolutionarystep of the bear; small size could correspond tomore ancient forms, namely more primitive ones.The increase in size can be linked to a thermoregu-lation factor following Bergmann’s rule (1847): theincrease in body size yields an advantage in ther-moregulation. Loss of heat in bodies of large size islower, causing a smaller surface-to-volume ratio. Inthis manner large sized populations can colonizecool regions. Moreover, in the case of the studiedbears, an increase in dimensions could also repre-sent a response to a shift towards a cooler climate.In contrast to these authors, Gerhard (2001) andRabeder & Nagel (2001) associate a similar reduc-tion in size to the shift toward cooler conditionsalthough this should be observable in high Alpineregions. The similarity between the Grotta SopraFontana Marella and Delle Streghe Cave fossils leadsus to link this trend to a climatic change, ratherthan to rapid evolution by cave bears.
AAcckknnoowwlleeddggmmeennttss
The authors thank D. Nagel (Vienna) for usefuladvise and critical reading of the manuscript, and G.Papalia (Pavia) for revision of the English. This studywas supported by a FAR grant contribution.
RReeffeerreenncceess
Bergmann, C. (1847): Ueber die Verhaeltnisse derWaemeoekonomie der Thiere zu ihrer Groesse. –Goettinger Studien 3, Pt. 1: 595-708.
Bona, F. (2004): Preliminary analysis on Ursus spelaeusRosenmüller & Heinroth, 1794 populations from“Caverna Generosa” (Lombardy-Italy). – CahiersScientifiques, Hors série 2: 87-98.
Cella, D.G. (1993): Il patrimonio speleologico dellaValstrona. – Labirinti, 13: 2-4.
Di Canzio, E., Petronio, C. (2001): Osservazioni sulla faunaa vertebrati pleistocenici della Grotta Cola ( Abruzzo,Aquila). – Boll. Soc. Paleont. It., 40 (1): 105-114.
Gerhard, W. (2001): The evolution of metapodial bones inthe cave bear group and its biostratigraphical implica-tions. – Cadernos Lab. Xeolòxico de Laxe Coruña, 26 :365-371.
Hofreiter, M., Capelli, C., Krings, M., Waits, L., Conard, N.,Munzel, S., Rabeder, G., Nagel, D., Paunovic, M.,Jambresic, G., Meyer, S., Weiss, G., Pääbo, S. (2002):Ancient DNA analyses reveal high mitochondrial DNAsequence diversity and parallel morphological evolu-tion of late Pleistocene cave bears. – MolecularBiology and Evolution, 19 (8):1244-1250.
Hofreiter, M., Rabeder, G., Jaenicke-Deprés, V., Withalm,G., Nagel, D., Paunovic, M., Jambr?sic, G. & Pääbo, S.(2004): Evidence of reproductive isolation betweencave bear population. – Current Biology, 14: 40-43.
Hue, E. (1907): Musée ostéologique. Étude de la faunequaternaire. Ostéometrie des Mammifères. 2 vol. –Librairie C. Reinwold, Schleicher Frères Editeurs, Paris.
Perego, R., Zanalda, E., Tintori, A. (2001): Ursus spelaeusfrom Grotta sopra Fontana Marella, Campo dei FioriMassif (Varese, Italy): morfometry e paleoecology. –Riv. It. Paleont. Strat., 107 (3): 451-462.
Rabeder, G. (1999): Die Evolution des Höhlenbären gebis -ses. – Mitteilungen der Kommission für Quartär for -schung der Österreichischen Akademie der Wissen -schaften, Band II, 102 pp.
Rabeder, G., Nagel, D. (2001): Phylogenetic problems ofthe Alpine Cave Bears. – Cadernos Lab. Xeológico deLaxe Coruña, 26: 359-364.
Rosendhal, W., Kempe, S. (2004): Johann Christian Rosen -müller und der Höhlenbär (1794-2004). „Lebensbilder“aus 210 Jahren. – Natur und Mensch 2003: 145-159.
Rossi, M., Santi, G. (2001 a): Gli ursidi della Grottadell’Orso (Laglio, Como, Lombardia, Italia Setten -trionale). Analisi morfometrica degli arti: indaginepreliminare. – Atti Soc. it. Sc. Nat. Mus. Civ. St. Nat.Milano, 141/2000 (2): 329-336.
Rossi, M., Santi, G. (2001 b): Archaic and recent Ursusspelaeus forms from Lombardy and Venetia region(North Italy). – Cadernos Lab. Xeológico de LaxeCoruña, 26: 317-323.
Santi, G., Rossi, M. (2001): Bears from the Buco dell’OrsoCave (Laglio-Como, Lombardy-Northern Italy). I:Morphometric study of the cranial and mandibularfossil remains. – Atti Ticinensi di Scienze della Terra,Pavia, 42: 75-100.
Santi, G., Rossi, M., Pomodoro, S. (2003): Cave bearsremains from the Buco dell’Orso cave (Lombardy-
123Geo.Alp, Vol. 2, 2005

Italy). Part III – Morphometric analysis of metapodialbones. – Bull. Inst. Royal Sc. Nat. de Belgique, 73: 195-219.
Strobino, F. (1981): Preistoria in Valsesia. Studi sul MonteFenera. – Società Valsesiana di cultura. pp.89.
Torres Pérez Hidalgo, T. (1988). Osos (Mammalia,Carnivora, Ursidae) del pleistocene Ibérico (U. denin-geri Von Reichenau, U. spelaeus Rosenmüller-Heinroth, U. arctos Linneo). – Boll. Geol. y Min.- lFilogenia, distribution stratigrafica y geografica.Estudio anatomico y metrico del craneo: 3-46. llEstudio anatomico y metrico de la mandibula, hioides,atlas y axis: 220-249. lll Estudio anatomico y metricodel miembro toracico, carpo y metacarpo: 359-412. lVEstudio anatomico y metrico del miembro pelviano,tarso, metatarso y dedos: 516-577. V Dentiction deci-
dual,formula dentaria y denticion superior: 660-714.Vl Denticion inferior: 886-940.
Weinstock, J. (1999): The upper Pleistocene mammalianfauna from the Grosse Grotte near Blauberen(Southwestern Germany). – Stuttgarter Beitr. Naturk.,Serie B: 277, 1-49.
Von den Driesch, A. (1976): A guide to the measurementof animal bones from archaeological sites. – PeabodyMuseum Bullettin, 1: 1-137.
124 Geo.Alp, Vol. 2, 2005
Plate 1: Ursus spelaeus Rosenmüller, 1794. Delle Streghe Cave (Sambughetto Valstrona, Piedmont, North Italy). AA – Skull. SpecimenMSNM V 4486, dorsal view. BB – Skull. Specimen MSNM V 5043, dorsal view. CC – Skull. Specimen MSNM V 5041, dorsal view. DD –Skull-cap of cub. Specimen MSNM V 4614, dorsal view. EE – Skull-cap of cub. Specimen MSNM V 4736, dorsal view. FF – Skull-cap ofcub. Specimen MSNM V 4721, dorsal view. GG - III phalanx. Specimen MSNM V 5028, lateral view. HH – Mandible. Specimen MSNM V5059, internal view. II - Skull. Specimen MSNM V 5043, frontal view. JJ – I phalanx. Specimen MSNM V 4988, dorsal view. KK – Radius.Specimen MSNM V 4331, external view. LL – Scapholunar. Specimen MSNM 4781, lateral view. MM – Astragalus. Specimen MSNM 4874,dorsal view. NN – Femur. Specimen MSNM V 4393, caudal view. OO – Radius. Specimen MSNM V 4304, dorsal view.
Manuscript submitted: November 26, 2004Revised manuscript accepted: June 22, 2005

125Geo.Alp, Vol. 2, 2005
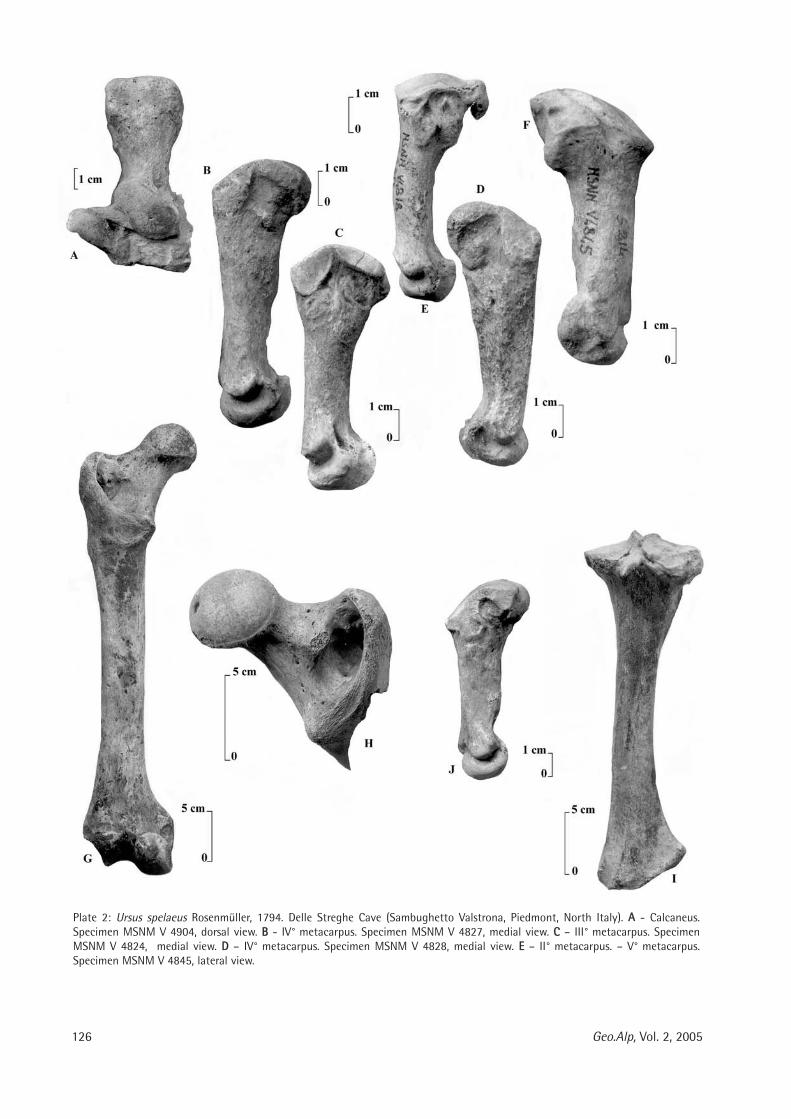
126 Geo.Alp, Vol. 2, 2005
Plate 2: Ursus spelaeus Rosenmüller, 1794. Delle Streghe Cave (Sambughetto Valstrona, Piedmont, North Italy). AA - Calcaneus.Specimen MSNM V 4904, dorsal view. BB - IV° metacarpus. Specimen MSNM V 4827, medial view. CC – III° metacarpus. SpecimenMSNM V 4824, medial view. DD – IV° metacarpus. Specimen MSNM V 4828, medial view. EE – II° metacarpus. – V° metacarpus.Specimen MSNM V 4845, lateral view.


















