Metody analizy decyzji Wykład 7 – sekwencyjne problemy decyzyjne

Metody analizy genomu
1. Mapowanie restrykcyjne.
2. Sondy do rozpoznawania DNA
3. FISH
4. Odczytanie sekwencji DNA
5. Interpretacja sekwencji DNA genomu
6. Transkryptom
7. Proteom

Były to pierwsze mapy
„molekularne”. genomu
Przykładem mapowania
restrykcyjnego jest strawienie
odcinka DNA dwoma enzymami
osobno i razem.
Poznajemy położenie i sekwencję
miejsc restrykcyjnych, sekwencji
pomiędzy nimi nie.
1. Mapy restrykcyjne

Mapowanie restrykcyjne daje
najlepsze wyniki dla stosunkowo
krótkich odcinków DNA, poniżej 50
kB, bo większe odcinki się zlewają
Gdy pozwolimy na pulsowanie pola
elektrycznego, nawet tak długie
odcinki jak 2Mb (małe
chromosomy, fragmenty dużych,
duże plazmidy, etc.) mogą być
rozdzielone.
1. Mapy restrykcyjne

2. Sondy do rozpoznawania DNA - identyfikowanie genu
na odcinku DNA
Możemy mieć informację o interesującym nas genie, a nie znamy jego
miejsca na chromosomie. Posiadana informacja może posłużyć do
skonstruwanaia sondy (ang. probe), która odszuka ten gen.
Sondy takie mogą być:
- otrzymane na bazie badanego mRNA, czyli są wyznakowanym cDNA
- wyznakowanymi fragmenty homologicznych genów znanych z innych
organizmów
- sztucznie zsyntetyzowane na podstawie sekwencji aminokwasów -
białka: -aa – aa – aa – aa – Tyr – Met – His – aa – aa – aa –
kodony TAC ATG CAC
lub inne TAT CAT

znacznik DNA
i próbki elektroforeza
bibuła
blot – przeniesienie
na filtr
sondy –
odszukują
sekwencje
komplementarne
błona
fotograficzna
Technika bibułowa - blotting
2. Sondy do
rozpoznawania
DNA
Zaczęło się od
omawianej wcześniej
hybrydyzacji metodą
Southerna.

3. FISH (fluorescent in situ
hibridization) - sonda wprost
na chromosom
Komórki z chromosomami w
stadium metafazy denaturujemy
chemicznie i prostujemy na
płytce.
Dodajemy sondę wyznakowaną
fluorescencyjnie.
Mapować można pod
mikroskopem !!

3. FISH (fluorescent in situ
hibridization) - sonda wprost na
chromosom
Na tym przykładzie widzimy
chromosom metafazowy (para
chromosomów homologicznych,
każdy ma dwie chromatydy).
Dodano tu na raz 18 różnych sond.

5. Odczytanie sekwencji DNA – metoda Sangera (tradycyjna)
Reakcja sekwencjonowania DNA polega na przerywanej syntezie DNA.
Wśród normalnych nukleotydów znajduje się domieszka nukleotydów
zmienionych tak, że uniemożliwiają one dalsze wydłużanie łańcucha. Te
zmienione nukleotydy maja 4 markery fluorescencyjne: A, G, T, C.
Tu przykład reakcji
kończącej się na A, inne
kończą się na G, T, C. .

Mieszanina reakcji
kończących się na
A, G, T, C
(z jednej matrycy DNA,
długiej do 700 bp)
jest rozdzielana w jednej
kapilarze sekwenatora
wyposażonego w odczyt
fluorescencji

5. Odczytanie sekwencji DNA
Dawniej rozdzielano i
odczytywano na ogromnych
żelach poliakrylamidowych
Jeden nukleotyd daje
widoczną róznice w tempie
migracji

5. Odczytanie sekwencji CAŁEGO GENOMU
Pewnym sposobem jest poznane wcześniej fizyczne porządkowanie kolekcji
klonów i sekwencjonowanie jej elementów. Stworzenie takiej biblioteki jest
jednak pracochłonne.
Trzeba robić sondy z poszczególnych fragmentów i nimi szukać następnych,
tak by się zazębiały.

Przykladem jest STS (sequence tagged site) mapping
Potrzebnymi unikatowymi znacznikami (tutaj A-I) mogą być sondy zrobione
z cDNA genów mających jedna kopię w genomie.
Możemy uporządkować naszą bibliotekę, czyli, zbudować kontig klonów.

5. Odczytanie sekwencji CAŁEGO GENOMU
Metodą szybszą jest losowe wielokrotne losowe sekwencjonowanie
fragmentów i numeryczne układanie ich w komputerze
- tzw, metoda „shutgun”
- nie wymaga fizycznego klonowania
- obecnie zdecydowanie dominująca
Widoczne pokrycie jest słabe, jak uzyskać setki fragmentów na
danym odcinku?

5. Nowe metody odczytywania sekwencji DNA – analizator Solexa
Na takich płytach
rozprowadza się
fragmenty DNA.
Losowy fragment
(kilkadziesiąt
nukleotydów) ma
wokół wolny promień
około kilku mikronów.
Pozwala to umieścić
miliony różnych
fragmentów na cm
kwadratowym

5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa
Pofragmentowane
DNA (nawet cały
genom na raz –
oczywiście w wielu
kopiach) jest
ligowane ze
standardowymi
starterami
polimeryzacji

Złączone fragmenty
DNA i wolne startery
są umieszczane na
płycie.
Losowy fragment
(kilkadziesiąt
nukleotydów) ma
wokół wolny promień
około kilku mikronów.
Pozwala to umieścić
miliony różnych
fragmentów na cm
kwadratowym.
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Mostkowanie – fragment
z dwoma starterami
odnajduje starter
komplementarny
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Replikacja do
fragmentu
dwuniciowego
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Rozdzielenie do
dwóch pojedynczych
nici
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa
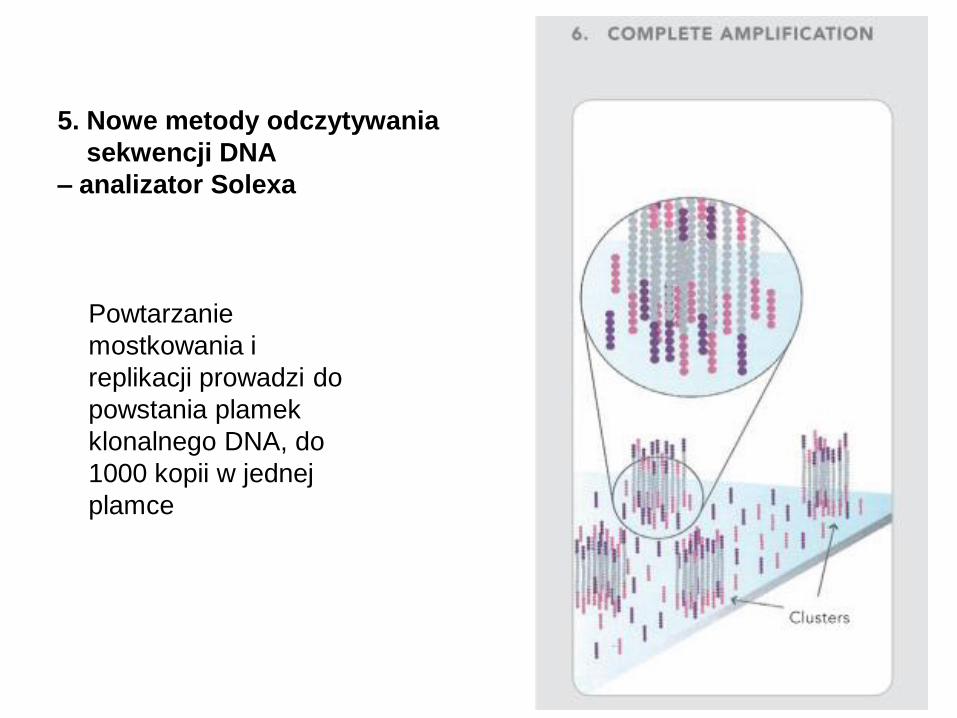
Powtarzanie
mostkowania i
replikacji prowadzi do
powstania plamek
klonalnego DNA, do
1000 kopii w jednej
plamce
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Dodawanie
pierwszego
nukleotydu
warunkowo
terminującego i
podświetlenie w celu
identyfikacji
przyłączonego
nukleotydu
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Identyfikacja
pierwszego
nulkeotydu
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Odblokowanie i
„odbarwienie” pierwszego
nukleotydu i dołączenie
drugiego świecącego i
blokującego nukleotydu
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Identyfikacja drugiego
nukleotydu
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Wielokrotne
powtarzanie
identyfikacji
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

Złożenie sekwencji w
kontig
5. Nowe metody odczytywania
sekwencji DNA
– analizator Solexa

ORF (open reading frame) to każda
sekwencja mająca na początku kodon
START (ATG), dalej inne kodony
aminokwasów, a kończąca się kodonem
STOP (TAG, TGA, lub TAA).
W sekwencji 4522
zasad są dwa
prawdziwe geny
kodujące białka
(ciemne linie) i
wiele fałszywych
(jasne).
Prawdziwe geny
są odpowiednio
długie i nie
zachodzą na
siebie (z pewnymi
wyjątkami).
6. Interpretacja sekwencji DNA –
czyli gdzie są geny?

6. Interpretacja sekwencji DNA
Poszukiwanie ORF jest utrudnione przez obecność intronów, szczególnie u
wyższych eukariontów.
W rozpoznaniu genów pomaga porównanie sekwencji DNA bliskich sobie
gatunków.

5. Finałem jest podanie sekwencji i ADNOTACJI genów
Celem mapowania fizycznego jest uzyskanie informacji, najlepiej z
dokładnością co do jednej zasady, gdzie w genomie umiejscowiony
jest interesujący nas element (gen, promotor, mutacja punktowa,
profag, etc.) - poniżej przykład mapy chromosomu drożdży.

6) Transkryptom
Transkryptom to ogół mRNA
obecnych w danym momencie w
komórce. Nowoczesna technologia
pozwala na badanie ekspresji
wszystkich genów równocześnie!
Sondy dla pojedynczych genów
układa się w mikropanelach. Trzeba
znać sekwencję badanych genów.
Taki mikropanel jest następnie
eksponowany na ekstrakt mRNA
uzyskany z genów ulegających
ekspresji, a powstawanie hybryd
wykrywa się laserem.

6) Transkryptom
Celem badań ekspresji wszystkich genów jest charakterystyka zmian jakim
ulega metabolizm komórki w różnych tkankach, warunkach
środowiskowych, stanach chorobowych, etc.

6) Transkryptom
Jeżeli badany jest jakieś zjawisko
trwające w czasie, to można pobierać
w zabiegu eksperymentalnym próbki
mRNA co pewien okres i uzyskać w
ten sposób obraz podniesienia
(zielono) lub obniżenia (czerwono)
ekspresji poszczególnych genów w
badanym czasie.

7. Proteom
Proteom to ogół białek obecnych w komórce. Białka zawarte w komórce
można rozdzielić w dwukierunkowej elektroforezie.
Obrót
Po wycięciu i oczyszczeniu
z żelu białko poddawane
jest spektrometrii masowej
dla zidentyfikowania
fragmentów (po strawieniu
na odcinki 5-70 aminokw.) a
potem ułożeniu ich w kontig
sekwencji białka.

7. Proteom
Chcielibyśmy znać nie tylko listę białek, ale też ich oddziaływania ze
sobą. W systemie dwuhybrydowym, dwa dowolne białka (np. ludzkie)
które podejrzewany o oddziaływanie łączymy z dwoma białkami drożdży.

7. Proteom
Wykorzystujemy białka drożdży, które muszą się zbliżyć do siebie i
dołączyć do odpowiednich miejsc przed genem reporterowym (np.
barwnik) by uruchomić transkrypcję. To się udaje, jeśli sztucznie
doczepione białka ludzkie oddziaływują ze sobą.

7. Proteom
Efektem tych
badań są sieci
oddziaływań
białek,
pokazujące
zwłaszcza te
białka, które
maja najwięcej
partnerów.
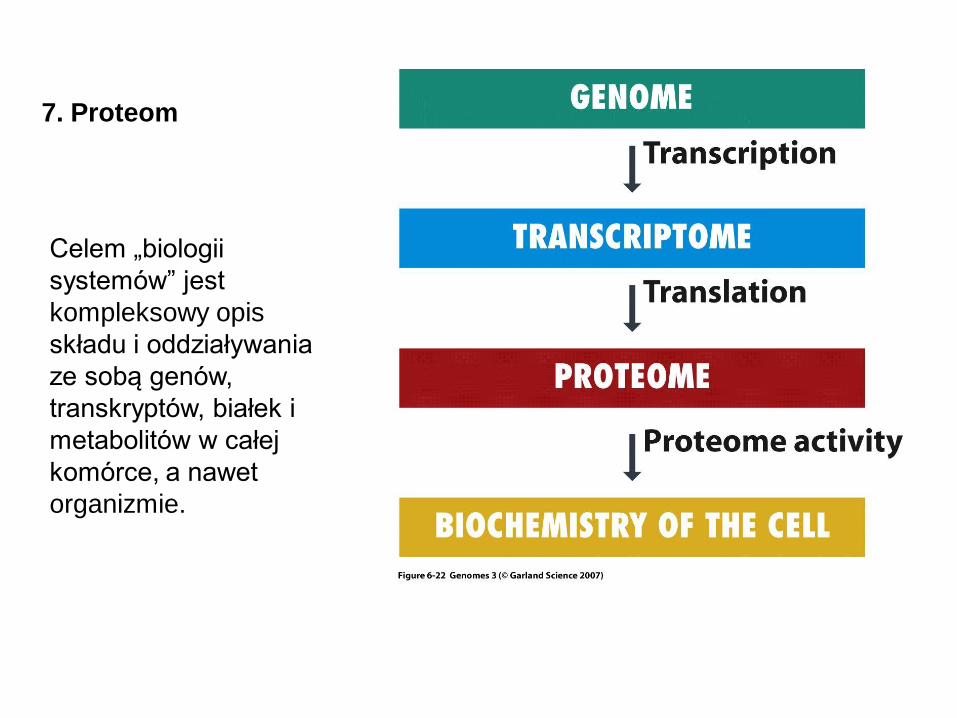
7. Proteom
Celem „biologii
systemów” jest
kompleksowy opis
składu i oddziaływania
ze sobą genów,
transkryptów, białek i
metabolitów w całej
komórce, a nawet
organizmie.