
Inhibition of oral morphogenesis during conjugation of Tetrahymena thermophila and its resumption...
11
Europ.]. Protisto!' 29, 359 -369 (1993 ) November 12, 1993 EUr0\>C3n Journal of PROTISTOLOGY Inhibition of Oral Morphogenesis During Conjugation of Tetrahymena thermophila and its Resumption After Cell Separation Mauryla Kiersnowska and Andrzej Kaczanowski Zoological Institute, University of Warsaw, Warsaw, Poland Gerard de Haller Department of Zoologyand Animal Biology, Un iversity of Geneva, Geneva, Switzerland SUMMARY During the early stages of the conjugation of Tetrahymena thermophila, the oral apparatus undergoes partial regression: the deep fiber bundle is resorbed and oral membranelles are partially deciliared, However, further progress of the oral replacement is arrested until cell separation. Only then proliferation of basal bodies for a new oral primordium is unblocked and concomitantly the old oral str uctures are resorbed. Th is step is followed by the assembly of new oral membranelles and resorption of the old ones. Thus, the cell separation itself may be the source of an important signal for the cortical morphogenesis. To test this hypothesis we have used micronuclearly defective "*" cells which separate after conjugation about 2.5 h earlier than normal cells and do not undergo postzygotic nuclear divisions. In the " *" crosses (the crosses of the "*" X wt cells) the oral primordium appeared before the cell separation (at the stage of hemikarya) following partial regression of the old oral structures, but only in those pairs in which the separa tion was already advanced in the separation. Consequently onset of conjugational morphogenesis in the "*" pairs was correlated with the time of the cell seI?aration, in spite of the differences in the course of nuclear events in the two kinds of pairs. Introduction There are two morphogenetical pathways in the ciliate Tetrahymena thermoph ila: i) divisional morphogenesis [7, 8, 14], ii) in situ reorganization of the oral apparatus, called an oral replacement, which occurs under stationary culture conditions [6, 13, reviewed in 7, 8] and after the process of conjugation [4]. During divisional morphogenesis, ciliary basal bodies in the midregion of the ciliary meridian * 1 proliferate into the intermeridional space forming an anarchic field of basal bodies. During oral replacement the top of ciliary row * 1 (instead of its middle segment) and the undulating membrane participate in the basal body proliferation for the anarchic field; subsequently the basal bodies assemble © 1993 by Gustav Fischer Verlag, Stuttgart into the oral membranelles. During oral replacement the old oral membranelles are fully resorbed concomitantly with the in situ formation of new ones [reviewed in 7 and 8J. Dur ing the somatic cell cycle the oral morphogenesis is synchronised with the nuclear divisions, thu s micro nuclear elongation corresponding to metaphase-anaphase transi- tion appears during stage 5 of the oral morphogenesis. On the other hand, under stationary and starvation conditions nond ividing cells undergo many cycles of the oral replace- ment without cell divisions [6, 13]. During Tetrahymena conjugation the nuclei undergo 5 nuclear divisions: meiosis I, meiosis II, 3rd prezygotic division and 2 postzygotic divisions after crossfertilization (Fig. 1A). It was not known whether conjugation morpho- genesis is synchronised with these nuclear divisions. 0932-4739/93/0029-0359$3.50/0
Transcript of Inhibition of oral morphogenesis during conjugation of Tetrahymena thermophila and its resumption...

Europ.]. Protisto!' 29, 359-369 (1993)November 12, 1993
EUr0\>C3n Journal of
PROTISTOLOGY
Inhibition of Oral Morphogenesis During Conjugationof Tetrahymena thermophila and its ResumptionAfter Cell Separation
Mauryla Kiersnowska and Andrzej KaczanowskiZoological Institute, University of Warsaw, Warsaw, Poland
Gerard de HallerDepartment of Zoologyand AnimalBiology, University of Geneva, Geneva,Switzerland
SUMMARY
During the early stages of the conjugation of Tetrahymena thermophila, the oral appara tusundergoes part ial regression: the deep fiber bundle is resorbed and oral membranelles arepartially deciliared, However, further progress of the oral replacement is arrested until cellseparation. Only then proliferation of basal bodies for a new oral primordium is unblocked andconcomitantly the old oral structures are resorbed. Th is step is followed by the assembly of neworal membranelles and resorption of the old ones. Thu s, the cell separation itself may be thesource of an import ant signal for the cortical morphogenesis. To test this hypothesis we haveused micronu clearly defective "*" cells which separate after conjugation about 2.5 h earlierthan normal cells and do not undergo postzygotic nuclear divisions. In the "*" crosses (thecrosses of the "*" X wt cells) the oral primordium appeared before the cell separation (at thestage of hemikarya) following partial regression of the old oral structures, but only in those pairsin which the separa tion was already advanced in the separation. Consequently onset ofconjugational morph ogenesis in the "*" pairs was correlated with the time of the cellseI?aration, in spite of the differences in the course of nuclear events in the two kinds ofpairs.
Introduction
There are two morphogenetical path ways in the ciliateTetrahymena thermoph ila: i) divisional morphogenesis [7,8, 14], ii) in situ reorganization of the oral apparatus,called an oral replacement, which occurs under stationaryculture conditions [6, 13, reviewed in 7, 8] and after theprocess of conjugation [4].
During divisional morphogenesis, ciliary basal bodies inthe midregion of the ciliary meridian * 1 proliferate intothe intermeridional space forming an anarchic field ofbasal bodies. During oral replacement the top of ciliaryrow * 1 (instead of its middle segment) and the undulatingmembrane participate in the basal body proliferation forthe anarchic field; subsequently the basal bodies assemble
© 1993 by Gustav Fischer Verlag, Stuttgart
into the oral membranelles. During oral replacement theold oral membranelles are fully resorbed concomitantlywith the in situ formation of new ones [reviewed in 7 and8J. During the somatic cell cycle the oral morphogenesis issynchronised with the nuclear divisions, thu s micronuclearelongation corre sponding to metaphase-anaphase transition appears during stage 5 of the oral morphogenesis. Onthe other hand , unde r stationary and starvation conditionsnond ividing cells undergo many cycles of the oral replacement without cell divisions [6, 13].
During Tetrahymena conjugation the nuclei undergo 5nuclear divisions: meiosis I, meiosis II, 3rd prezygoticdivision and 2 postzygotic divisions after crossfertilization(Fig. 1A). It was not known whether conjugation morphogenesis is synchronised with these nuclear divisions.
0932-4739/93/0029-0359$3.50/0

360 . M. Kiersnowska, A. Kaczanowski and G. de Haller
It was observed previously that the oral apparatus ispartl y reduced early in conjugation , but the oral morphogenesis is completed only after separation of the conjugant s [4]. Therefore our first aim was to establish whetherthe old oral apparatus becomes "fro zen" early in conjugation for the remaining time of pairing, or whether it stillundergoes morphogenetical events before cell separation.
The pair separation is a source of important signals forthe anlagen DNA replication and for the completion of theresorption of the old macronucleus [15]. But the separation could also be a source of signals for the commencement of the oral morphogenesis. For this reason wefollowed the events occurring around the time of cellseparation, and attempted to corre late these events withthe resumption of oral morphogenesis. We used a micronuclearly defective strain, called the "*" strain, becauseduring its conjugation with a tester strain, the pairssepara te about 2.5 h earlier than contro l pairs under thesame conditions (Fig. 1B) [10, 16].
Resumption of food uptake in the exconjugant cellsoccurs only after the completion of the oral morphogenesis. In this case resumption of feeding activity in theabortive "*"exconjugant s retaining old macronuclei wasexpected to occur earlier, thus suggesting that the cellsepa ration was a source of the morphogenetic signal actingon the posttranslational level.
Material and Methods
The following strains of T. tbermopbila have been used in thepresent study: CU 399 strain mt VI kindly provided by Dr.P. Bruns, WU 131, WU 113 mt IV, WU 131 mt IV, WU 132mt VI, and WU 132 obtained in our laboratory, and twomicronuclearly defective strains A* III and A* V kindly pro-
vided by Dr. Frankel (see [23] for the origin and characteristics ofthese two "*" strains; [1, 5 and 16] for cytogenetics of the "*"strain conjugation, crossed to a diploid tester).
Inducing ofconjugation. Cells of complementary mating typeswere prestarved in a 10 mM TRIS HCl buffer of pH 7.4 for about20-24 h and then mixed for conjugation at 30°C, according toBruns and Brussard [2] and Martindale et al. [17].
The feeding activity of conjugants and exconjugants wasassayed by the cont inuous exposure of cells to an Indian inksuspension beginning after 10 h of conjugation of normal cells,and after 6 h of the "*" cells conjugation. The Indian ink had noadverse effect on the cells, judging by a comparison withuntreated controls.
Cytological methods. Giemsa staining was performed according to Gaertig et al. [11], protargol staining according to Ng andNelsen [19].
Immunofluorescence staining was performed according toCohen er al. [3]. Monoclonal anrityrosine tubuline (clone TublA2) was obtained from Sigma. Nuclei were costained with DAP]fluorochrome.For scanning electron microscopy (SEM) the cells were preparedin the following way: first they were deciliated by Thompson'smethod modified by Gaert ig et al. [11]. 5 min after deciliation thecells were fixed in 2% (w/v) osmium tetroxide, dehydrated in aseries of alcohol and prepared according to the procedure ofRuffolo [21]. The scanning microscope JEOL-JSM 6400 wasapplied.
For transmission electron microscopy (TIM) the cells werefixed for 2 h with 2% glutar aldehyde at room tempera ture and7.2 pH (cacodylate buffer), rinsed 3 times with the buffer andpostfixed with 1% osmium tetroxide. Finally the cells werewashed, dehydrated and embedded in the Spurr's medium. Gridswere stained with 2% (w/v) uranyl acetate followed by leadacetate. The stages of macronuclear development I and II and thestagesof oral development (1-6) will be referred to as described in[17] and [8], respectively.
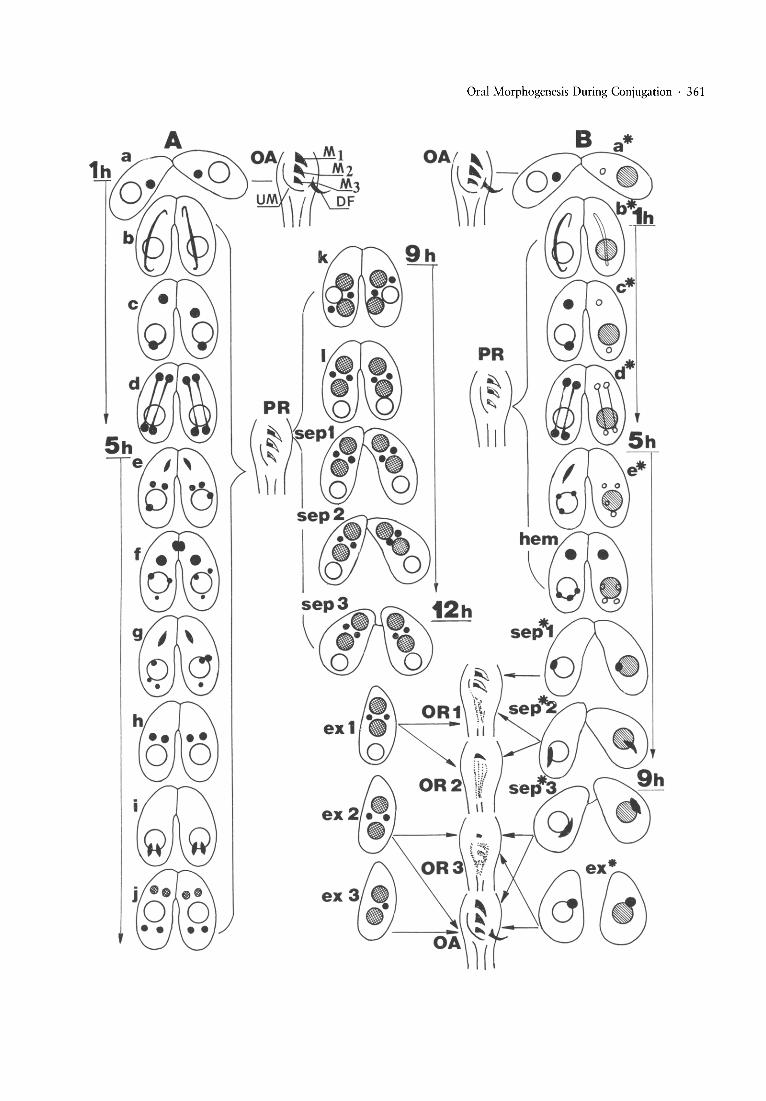
Fig. 1. Relative timing of nuclear events, cell separation and oral morph ogenesis during conjugation of Tetrahymena thermophila. ~
Approximate times in hours (1 h,5 h and so on) from the mixing of the cellsare given only for some conjugation stages; Panel A: normalconjugation (tester x tester) (cross-hatched circles represent macronuclear anlagen, open circles old macronuclear); Panel B: abortivecon jugation: tester X "*" strain cross (small filled circles represent the micronuclei of wt parent and hemikarya, open and smallercircles the micronuclei of the "*" parent , open and hatched large circles the retained macronuclei of wt and "*" parent s, respectively).Oral structures are shown between both panels; Arrows and brackets indicate the correspondence of the exconjugant stage and oralreplacement stage in the normal sequence and of the cell separation stage and oral replacement in the " * " conjugation.Abbreviations for panel A: nuclear events during early conjugation (a = early pairs, b = meiotic prophase, c = produ cts of the meiosis](interkinesis), d = meiosis II, e = 3rd prezygotic division, f = pronuclei, g = 1st postzygotic division, h = products of the 1stposrzygotic division, i = 2d postzygotic division, j = macronuclear development I, k = rearran gement of nuclei, I = macronucl eardevelopment II), sep = separation of pairs (sep 1 = beginning of separation, sep 2 = advanced separation, sep 3 = end of separation),ex = exconjugants (ex-l with 2 micronu clei and the relic of the old macronucl eus, ex-Z with resorbed old macronucleus, ex-3 withresorbed old macronucleus and 1 of the 2 micronuclei. Approximate time of appearance of some stages from the mixing of cells isindicated .Abbreviations for panel B: stages a*, b*, c* , d*, and e* correspond to the respective stages a-e of the normal sequence, hem =hemikarya , sep* = separation of the "*" pairs: sep*-l = beginning, sep* -2 = advanced and sep-Jx = end of the separation ;hemikaryon during these stages is shifted posteriorly and elongated, the old macronucleus is retained, ex* = "*" exconjugants.Abbreviations for oral apparatus: OA = complete oral apparatus at the beginning of pairing and at the end of postconjugationalmorphogenesis (M I-M 3 = adoral membranelles, UM = undulating membrane, OF = deep fiber bundle), PR = partial regression oforal structures (black sectors represent ciliated basal bodies, white nonciliated ), brackets in both panels indicate the conjugation stageswith "frozen" partially regressed oral apparatus, OR-l, OR-2, OR-3 = stages 1, 2 and 4- 5 of the postconjugation oralreplacement.
Oral Morphogenesis During Conjugation· 361
362 . M. Kiersnowska, A. Kaczanowski and G. de Haller
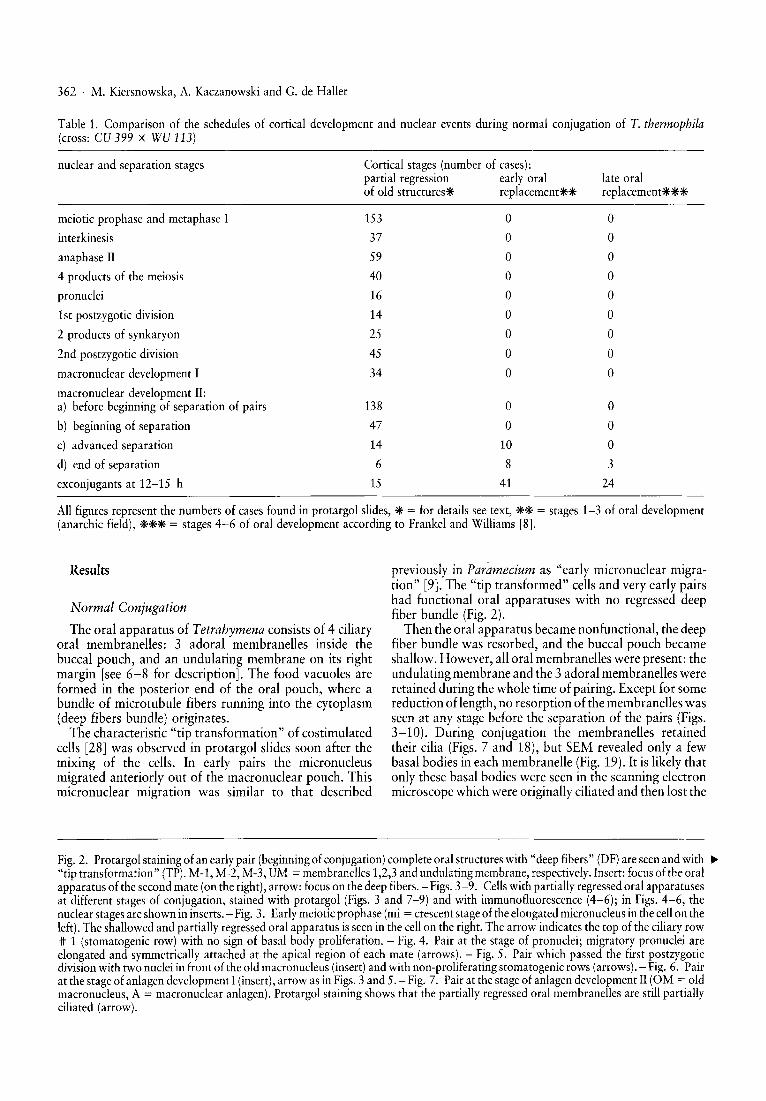
Table 1. Comparison of the schedules of cortical development and nuclear events during normal conjugation of T. thermophila(cross: CU 399 X WU 113)
nuclear and separation stages Cortical stages (number of cases):partial regression early oralof old structurese replacementeeee
late oralreplacement***
meiotic prophase and metaphase I
interkinesis
anaphase II
4 products of the meiosis
pronuclei
1st postzygotic division
2 products of synkaryon
2nd postzygotic division
macronuclear development I
macronuclear development II:a) before beginning of separation of pairs
b) beginning of separation
c) advanced separation
d) end of separation
exconjugants at 12-15 h
153
37
59
40
16
14
25
45
34
138
47
14
6
15
ooooooooo
oo
10
8
41
ooooooooo
ooo3
24
All figures represent the numbers of cases found in protargol slides, * = for details see text, ** = stages 1-3 of oral development(anarchic field), *** = stages 4-6 of oral development according to Frankel and Williams [8].
Results
Normal Conjugation
The oral apparatus of Tetrahymena consists of 4 ciliaryoral membranelles: 3 adoral membranelles inside thebuccal pouch, and an undulating membrane on its rightmargin [see 6-8 for description]. The food vacuoles areformed in the posterior end of the oral pouch, where abundle of microtubule fibers running into the cytoplasm(deep fibers bundle) originates.
The characteristic "tip transformation" of costimulatedcells [28J was observed in protargol slides soon after themixing of the cells. In early pairs the micronucleusmigrated anteriorly out of the macronuclear pouch. Thismicronuclear migration was similar to that described
previously in Paramecium as "early micronuclear migration" [9J. The "tip transformed" cells and very early pairshad functional oral apparatuses with no regressed deepfiber bundle (Fig. 2).
Then the oral apparatus became nonfunctional, the deepfiber bundle was resorbed, and the buccal pouch becameshallow. However, all oral membranelles were present: theundulating membrane and the 3 adoral membranelles wereretained during the whole time of pairing. Except for somereduction of length, no resorption of the membranelles wasseen at any stage before the separation of the pairs (Figs.3-10). During conjugation the membranelles retainedtheir cilia (Figs. 7 and 18), but SEM revealed only a fewbasal bodies in each membranelle (Fig. 19). It is likely thatonly these basal bodies were seen in the scanning electronmicroscope which were originally ciliated and then lost the
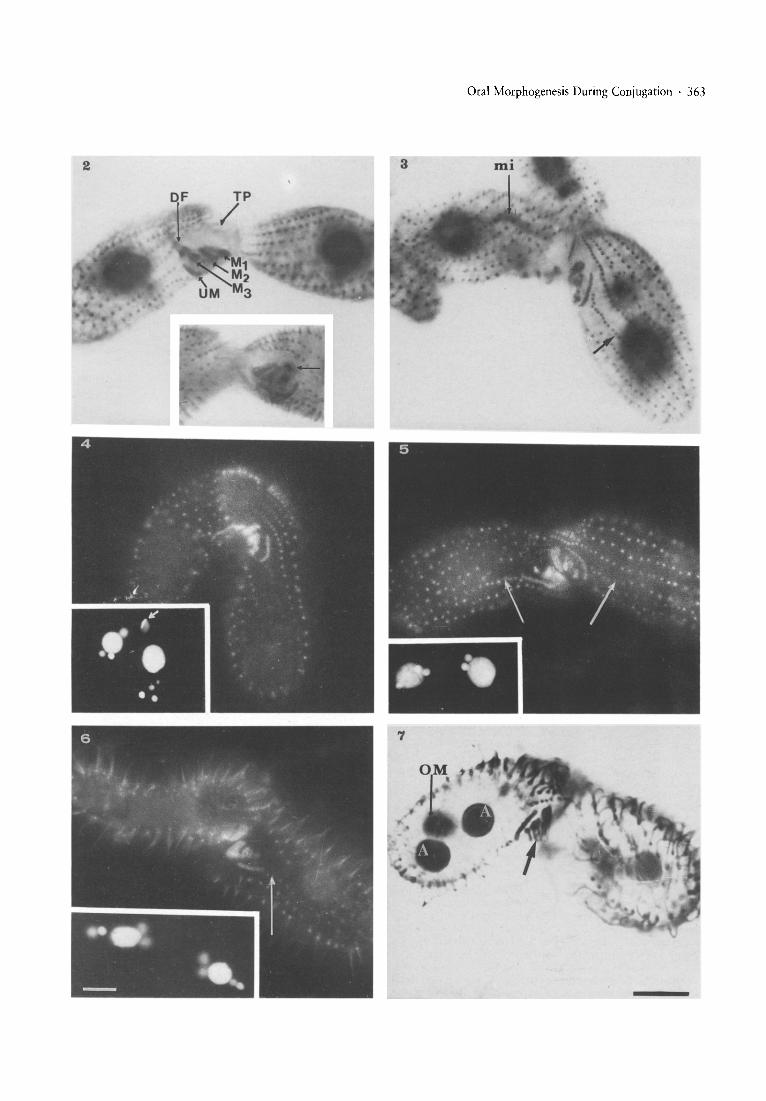
Fig. 2. Protargol staining of an early pair (beginning of conjugation) complete oral structures with "deep fibers" (DF) are seen and with ~
"tip transformation" (TP). M-l, M-2, M-3, UM = membranelles 1,2,3 and undulating membrane, respectively. Insert: focus of the oralapparatus of the second mate (on the right), arrow: focus on the deep fibers. - Figs. 3-9. Cells with partially regressed oral apparatusesat different stages of conjugation, stained with protargol (Figs. 3 and 7-9) and with immunofluorescence (4-6); in Figs. 4-6, thenuclear stages are shown in inserts. - Fig. 3. Early meiotic prophase (mi = crescent stage of the elongated micronucleus in the cellon theleft). The shallowed and partially regressed oral apparatus is seen in the cell on the right. The arrow indicates the top of the ciliary row'If 1 (stomatogenic row) with no sign of basal body proliferation. - Fig. 4. Pair at the stage of pronuclei; migratory pronuclei areelongated and symmetrically attached at the apical region of each mate (arrows). - Fig. 5. Pair which passed the first postzygoticdivision with two nuclei in front of the old macronucleus (insert) and with non-proliferating stomatogenic rows (arrows). - Fig. 6. Pairat the stage of anlagen development I (insert), arrow as in Figs. 3 and 5. - Fig. 7. Pair at the stage of anlagen development II (OM = oldmacronucleus, A = macro nuclear anlagen). Protargol staining shows that the partially regressed oral membranelles are still partiallyciliated (arrow).
Oral Morphogenesis During Conjugation . 363
TP
/r2
"1
364 . M. Kiersnowska, A. Kaczanowski and G. de Haller
9
jO
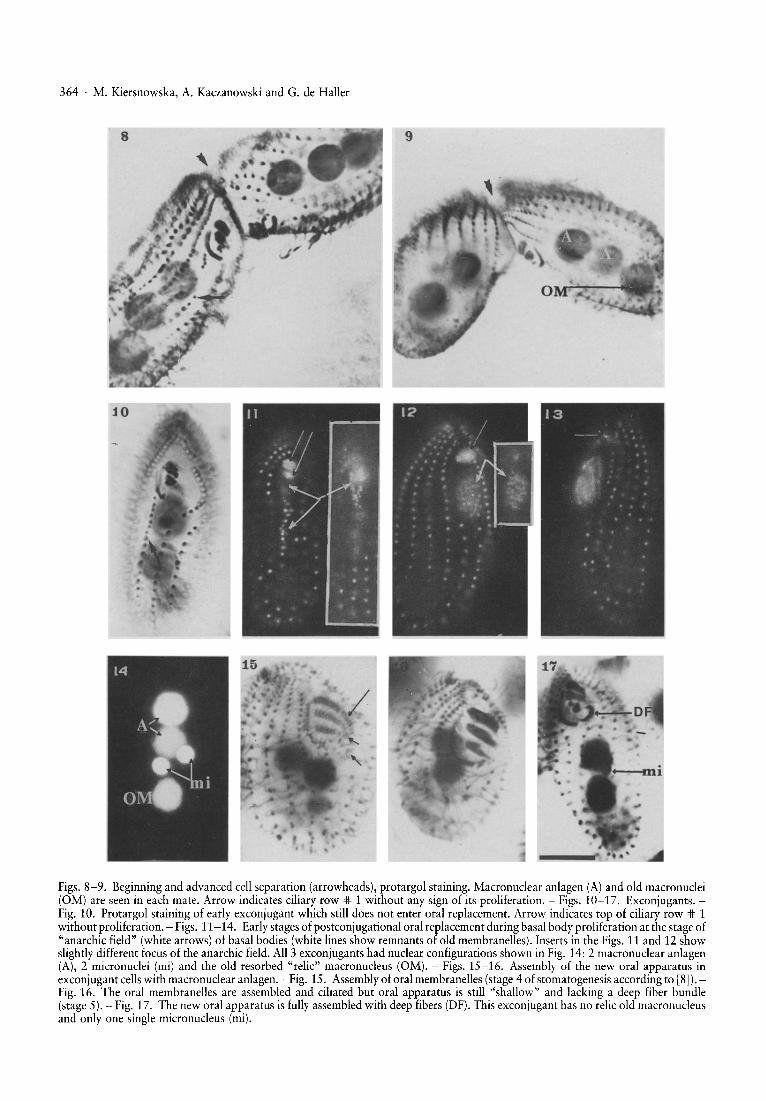
Figs. 8-9. Beginning and advanced cell separation (arrowheads), protargol staining. Macronuclear anlagen (A) and old macronuclei(OM) are seen in each mate. Arrow indicates ciliary row '* 1 without any sign of its proliferation. - Figs. 10-17. Exconjugants.Fig. 10. Protargol staining of early exconjugant which still does not enter oral replacement. Arrow indicates top of ciliary row '* 1without proliferation. - Figs. 11-14. Early stages of postconjugational oral replacement during basal body proliferation at the stage of"anarchic field" (white arrows) of basal bodies (white lines show remnants of old membranelles). Inserts in the Figs. 11 and 12 showslightly different focus of the anarchic field. All 3 exconjugants had nuclear configurations shown in Fig. 14: 2 macronuclear anlagen(A), 2 micronuclei (mi) and the old resorbed "relic" macronucleus (OM). - Figs. 15-16. Assembly of the new oral apparatus inexconjugant cellswith macronuclear anlagen. - Fig. 15. Assembly of oral mernbranelles (stage 4 of stomatogenesis according to [8]).Fig. 16. The oral membranelles are assembled and ciliated but oral apparatus is still "shallow" and lacking a deep fiber bundle(stage 5). - Fig. 17. The new oral apparatus is fully assembled with deep fibers (DF). This exconjugant has no relic old macronucleusand only one single micronucleus (mi).
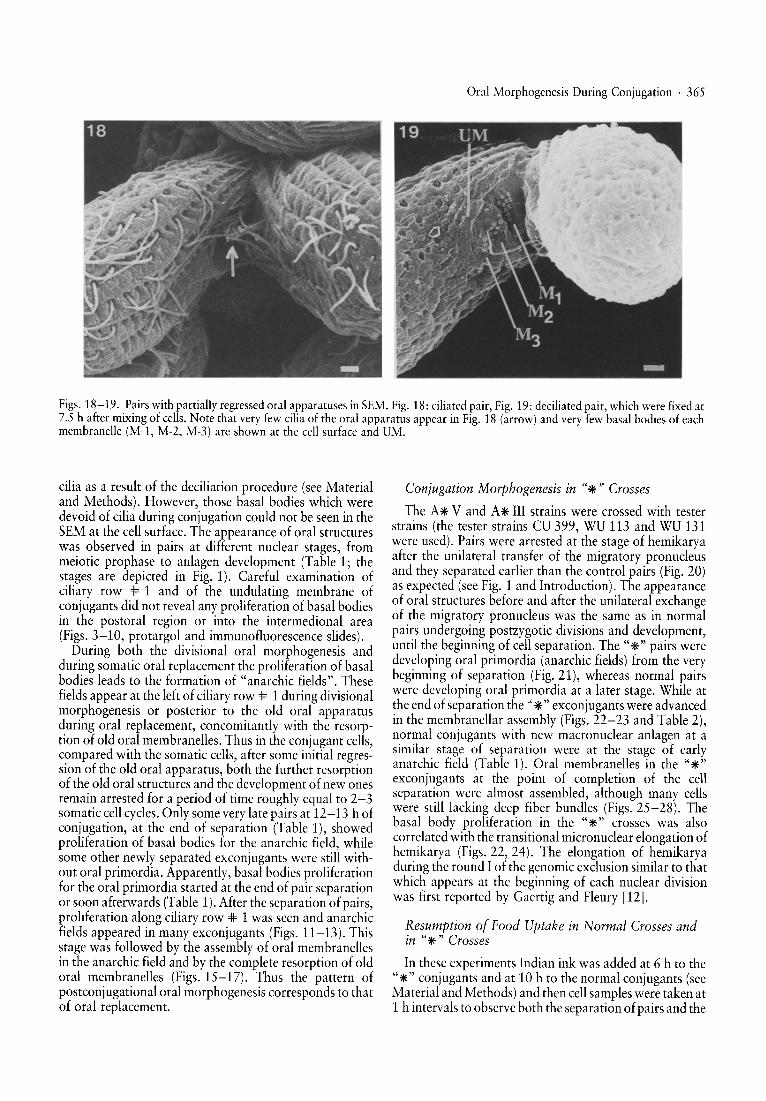
Oral Morphogenesis During Conjugation . 365
Figsh1~-19 ..P~irs withpartiallyregressed oral ~l?paratuses inSEM. Fig. 18: ciliated pair, Fig. 19: deciliated pair, whichwerefixed at
7.5 ba ter mlxmg of cells. Note that veryfewciliaof the oral apparatus appear in Fig. 18 (arrow) and veryfewbasal bodies of eachmem randle (Mvl, M-2, M-3) are shown at the cell surface and UM.
cilia as a result of the deciliation procedure (see Materialand Methods). However, those basal bodies which weredevoid of cilia during conjugation could not be seen in theSEM at the cell surface. The appearance of oral structureswas observed in pairs at different nuclear stages frommeiotic prophase to anlagen development (Tablel ; thestages are depicted in Fig. 1). Careful examinatio~ ofcili~ry row .# 1 and of the undulating membrane ofconjugants did not reveal any proliferation of basal bodiesin the postoral region or into the intermedional area(Figs. ~-10, protargol and immunofluorescence slides).~unng bo~h the divisional oral morphogenesis and
dun.ng somatic oral replacement the proliferation of basalbodies leads to the formation of "anarchic fields". Thesefields appear at the left of ciliary row * 1 during divisionalmo~phogenesis or posterior to the old oral apparatus~unng oral replacement, concomitantly with the resorption of old o~al membran~lles. Thus in the conjugant cells,c?mpared WIth the somatic cells, after some initial regresSIOn of the old oral apparatus, both the further resorptionof th~ old oral structures and the development of new onesremau?- arrested for a period of time roughly equal to 2-3s0m.atic ~ell cycles. Only some very late pairs at 12-13 h ofconJ~gatI?n, at the end of separation (Table 1), showedproliferation of basal bodies for the anarchic field whilesome other newly separated exconjugants were still without oral primordia. Apparently, basal bodies proliferationfor the oral primordia started at the end of pair separationor s~on a~terwards (~~ble 1). After the separation of pairs,proliferation along ciliary row * 1 was seen and anarchicfields appeared in many exconjugants (Figs. 11-13). Thisstage was followed by the assembly of oral membranellesin the anarchic field and by the complete resorption of oldoral me.mbr~nelles (Figs. 15-17). Thus the pattern ofpostconjugational oral morphogenesis corresponds to thatof oral replacement.
Conjugation Morphogenesis in "*" Crosses
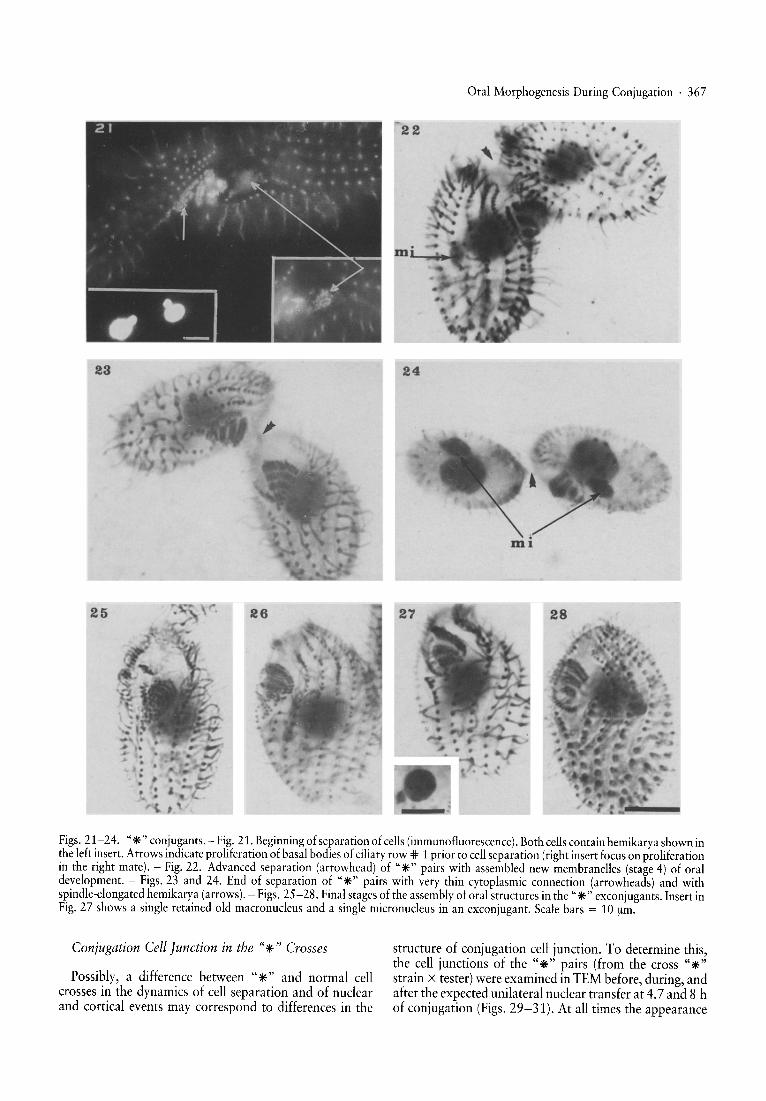
The A* V and A* III strains were crossed with testerstrains (the tester strains CD 399, WU 113 and WU 131were used). ~airs were arrested at the stage of hemikaryaafter the unilateral transfer of the migratory pronucleusand they separated earlier than the control pairs (Fig. 20)as expected (see Fig. 1 and Introduction). The appearanceof oral st~uctures before and after the unilateral exchangeof the migratory pronucleus was the same as in normalpai~s underg?in~ postzygotic divisions and development,until th~ begmnmg. of cel.l separation. The "*" pairs weredev~lo~mg oral pnm?rdia \anarchic fields) from the verybeginning of .separatIon. (FIg..21), whereas normal pairswere developmg oral primordia at a later stage. While at~he end of separation the" *" exconjugants were advancedin the membranellar assembly (Figs. 22-23 and Table 2)n.or~al conjugants with new macronuclear anlagen at ~SImIlar. st~ge of separation were at the stage of earlyanarchic field (Table 1). Oral membranelles in the "*"exconjugants at the point of completion of the cellseparat~on we:e almost ~ssembled, although many cellswere still lacking deep fiber bundles (Figs. 25-28). Thebasal body proliferation in the "*" crosses was alsocorr~lated wit~ the transitional micronuclear elongation ofhe~Ikarya (FIgS. 22, 24). The elongation of hemikaryadunng the round I of the genomic exclusion similar to thatwhich appears at the beginning of each nuclear divisionwas first reported by Gaertig and Fleury [12].
Resumption of Food Uptake in Normal Crosses andin "*" Crosses
In these experiments Indian ink was added at 6 h to the"*" conjugants and at 10 h to the normal conjugants (seeMa~erial and Methods) and then cell samples were taken at1 h intervals to observe both the separation of pairs and the
366 . M. Kiersnowska, A. Kaczanowski and G. de Haller
Table 2. Comparison of the schedules of cortical development and nuclear events during round I of the genomic exclusion(cross: A* V x CU 399)
nuclear and separation stages cortical stages (number of cases):partial regression early oralof old structures replacement
late oralreplacement
meiotic prophase and metaphase
interkinesis
anaphase II
4 meiotic products
3rd prezygotic division
pronuclei
hemikarya before separation of pairs
beginning of separation
advanced separation
end of separation
130
36
23
53
26
66
37
44oo
ooooooo
10
23
o
oooooooo
15
11
All explanations as in Table 1.
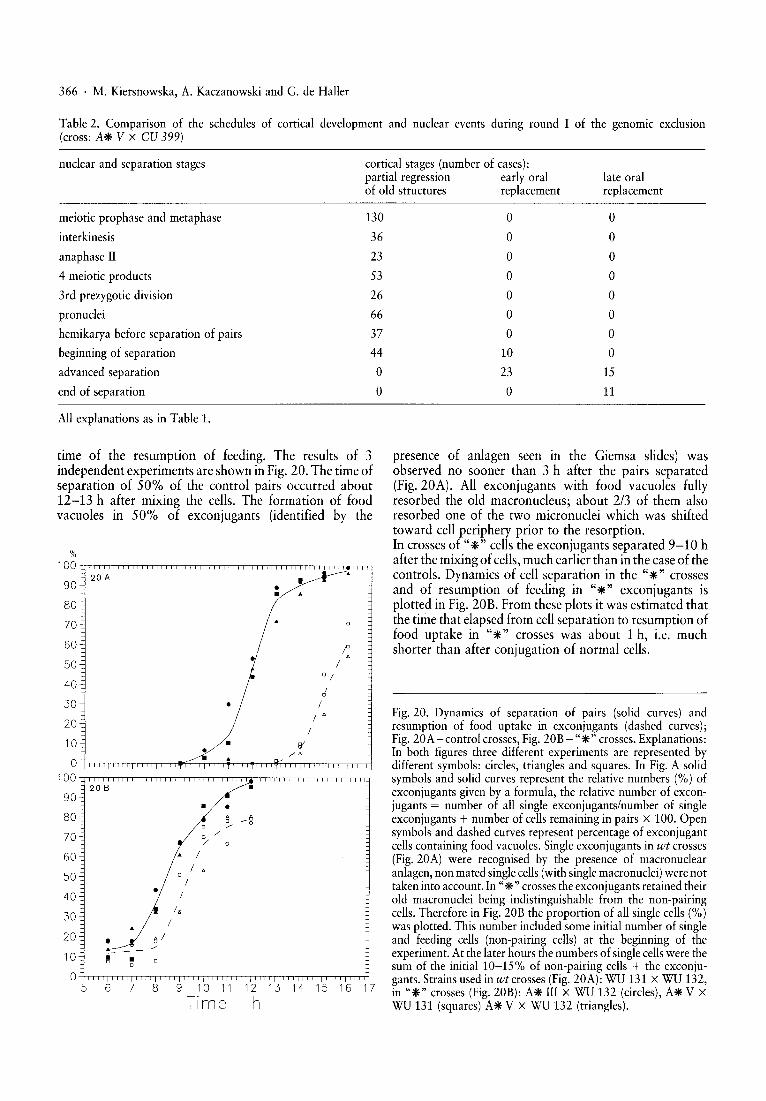
presence of anlagen seen in the Giemsa slides) wasobserved no sooner than 3 h after the pairs separated(Fig.20A). All exconjugants with food vacuoles fullyresorbed the old macronucleus; about 2/3 of them alsoresorbed one of the two micronuclei which was shiftedtoward cell periphery prior to the resorption.In crosses of "*" cells the exconjugants separated 9-10 hafter the mixing of cells,much earlier than in the case of thecontrols. Dynamics of cell separation in the "*" crossesand of resumption of feeding in "*" exconjugants isplotted in Fig. 20B. From these plots it was estimated thatthe time that elapsed from cell separation to resumption offood uptake in "*" crosses was about 1 h, i.e. muchshorter than after conjugation of normal cells.
Fig. 20. Dynamics of separation of pairs (solid curves) andresumption of food uptake in exconjugants (dashed curves);Fig. 20A-control crosses, Fig. 20B - "*" crosses. Explanations:In both figures three different experiments are represented bydifferent symbols: circles, triangles and squares. In Fig. A solidsymbols and solid curves represent the relative numbers (%) ofexconjugants given by a formula, the relative number of exconjugants = number of all single exconjugants/number of singleexconjugants + number of cells remaining in pairs x 100. Opensymbols and dashed curves represent percentage of exconjugantcells containing food vacuoles. Single exconjugants in wt crosses(Fig. 20A) were recognised by the presence of macronuclearanlagen, non mated singlecells (with single macronuclei) were nottaken into account. In "*" crosses the exconjugants retained theirold macronuclei being indistinguishable from the non-pairingcells. Therefore in Fig. 20B the proportion of all single cells (%)was plotted. This number included some initial number of singleand feeding cells (non-pairing cells) at the beginning of theexperiment. At the later hours the numbers of singlecellswere thesum of the initial 10-15% of non-pairing cells + the exconjugants. Strains used in wt crosses (Fig. 20A): WU 131 X WU 132,in "*" crosses (Fig. 20B): A* III X WU 132 (circles), A* V XWU 131 (squares) A* V X WU 132 (triangles).
%100
20 A •90 ••80
70
60
50 •
40,f
30 • /
20/ 0
/10 • W• /0
0100
20 B •90
• •80 ~ --e70 0
-:/ 0
60 I
/ 0
50 /• /40
30I.
I
20 • e I/'
10 ~ •00
5 6 7 8 9 10 11 12 13 14 15 16 17
Time h
time of the resumption of feeding. The results of 3independent experiments are shown in Fig. 20. The time ofseparation of 50% of the control pairs occurred about12-13 h after mixing the cells. The formation of foodvacuoles in 50% of exconjugants (identified by the
23
Oral Morphogenesis During Conjugation . 367
24
25 28
Figs. 21-24. "*"conjugants. - Fig. 21. Beginning of separat ion of cells (immunofluorescence). Borh cells contain hemikarya shown inthe left insert . Arrows indicate prol iferat ion of basal bodies of ciliary row * 1 prior to cell separation (right insert focus on proliferationin the right mare). - Fig. 22. Advanced sepa ration (arrow head) of "*"pairs with assembled new membranelles (stage 4) of oraldevelopment. - Figs. 23 and 24. End of separarion of "*" pairs with very thin cytopl asmic connection (arrowheads) and withspindle-elongated hemikarya (arrows). - Figs. 25-28. Final stages of the assembly of oral structures in the "*" exconjugants . Insert inFig. 27 shows a single retained old macronucleus and a single micronucleus in an exconjugant. Scale bars = 10 urn.
Conjugation Cell Junction in the "*" Crosses
Possibly, a difference between "*" and normal cellcrosses in the dynamics of cell separation and of nuclearand cortical events may corre spond to differences in the
structure of conjugation cell junct ion. To determine this,the cell junctions of the "*" pairs (from the cross "*"strain X tester) were examined in TEM before, during, andafter the expected unilate ral nuclear transfer at 4.7 and 8 hof conjugation (Figs. 29-31). At all times the appearance

368 . M. Kiersnowska, A. Kaczanowski and G. de Haller
Figs. 29-31. Conjugation cell junction in "*" pairs (A* V X tester) at4, 7 and 8 h, respectively. In all 3 figures the cell junction with itscharacteristic pores is shown (arrows). In Fig. 29, there is also visible a tip of the micronucleus (mi) of the non-star mate which iselongated (crescent stage of the meiotic prophase); Fig. 30 shows a micronucleus (mi) which attaches to the conjugation cell junction(presumably it is the migratory pronucleus). Scale bar for Figs. 29-31 = 1 urn.
of the junction in the "*" crosses was similar to that whichwas observed in the wt cross both in this study and asreported previously [26,27].
Discussion
The process of pairing at 30°C takes about 10 h [10, 15,17 and this paper], in contrast to the 3-4 h of cell cycle inthe growing culture of Tetrahymena. Thus the time ofconjugation was roughly 2.5-3 times as long as thesomatic cell cycle. During the cell cycle, the divisionalmorphogenesis is synchronised with nuclear divisions.Stationary and starved nondividing cellsof T. thermophilaalso continually undergo an oral morphogenesis called theoral replacement in absence of nuclear divisions. Surprisingly, although numerous nuclear divisions were takingplace during as long time of pairing as 10 h, the oralapparatus remained "frozen" at the stage of partialregression, in agreement with the previous report of Coleand Frankel [4]. During this stage some but not all basalbodies of oral membranelles are ciliated and can be seen onthe cell surface in SEM. Tsunemoto et al. [22] in their SEMstudy have also observed very few ciliary basal bodies inthe partly regressed oral structures during conjugation ofTetrahymena, but we believe that they did not see thoseunciliated basal bodies which were underneath the cellsurface.
Our results suggest that the separation of the conjugation pairs is the most important signal for the unblockingof basal bodies proliferation for postconjugational oralreplacement. The proliferation of ciliary basal bodies for
the oral primordium could have begun shortly beforeseparation but in most cases it appeared to begin only afterthe final separation.
The oral morphogenesis was also arrested in the "*"cells during prezygotic divisions and at the stage ofhemikarya after unilateral nuclear transfer. These pairsseparated much earlier than the control pairs as observedpreviously [10, 16]. The first proliferation of ciliary basalbodies for the oral primordia appeared in the "*" crossesduring the stage of "beginning separation" (Fig. 1) and bythe end of separation they were well advanced in the oralmorphogenesis. The resumption of the oral morphogenesistook place before the cell separation in the "*" crosses andafter cell separation in control (wt) crosses, but in bothcases it was correlated with the separation. Thus, in spite ofthe different sequence the cell separation in the"*" crossescould still be an important signal for the resumption of oralmorphogenesis. Cole and Frankel [4] have observedblocked separation in cortical mutant pairs jan x jan aswell as blocked resorption of old structures even after 20 hof conjugation. Kaczanowski [15] observed that theresorption of old macronucleus and cell separation werealso blocked in mra mutant cells (mra for macronuclearresorption arrest). These conjugants were blocked inpostconjugational oral morphogenesis in the same way asjan X jan conjugants (Kiersnowska and Kaczanowski,unpublished results). Apparently, cellunion in conjugation"freezes" progress of the oral morphogenesis in Tetrahymena.
The time elapsing from cell separation to feeding activitywas much shorter in "*" pairs (about 1 h) than in normalcontrols (about 3 h). This time difference is correlated with

the earlier onset of oral morphogenesis in the "*" pair sand can be due to any yet unknown property of theconjugation cell junction. Ho wever, we did not find anydifference in the structure of the junct ion between the totand"*"pairs in the TEM and this structure was similar tothat observed previously by Wolfe [26, 27]. However, it isalso possible that the late morphogenetic events (assemblyof basal bodies into new membranelles, formation of oralpouch and deep fiber bundle ) may require new transcriptswhich are produced with some time lag during normalconjugation. In the late stages of normal conjugation(about 3 h before cell separation), the old macronucleus issilenced when it becomes highly condensed, forming onebig phagosome, enwrapped in additional membranes [24].Subsequently, the expression of known new macronucleargenes begins at a certain time after pair separation [18,20],although some transcriptional activity of the new macronucleus was observed autoradiographically at an earliertime [25]. In contrast the "*" pair s do not resorb the oldmacronucleus, and one may expect that this nucleus istranscriptionally active throughout the conjugation,speeding up the process of postconjugational oral morphogenesis.
Acknowledgements
Thi s work was supported by a gram of Th e Polish Ministry ofEducation G-MEN-75 to A.K. and by gra nts from DonationGeorges and Antoine Claraz to G.B. Thanks are due to Dr. JaninaKaczanowska for the inspiring discussion s, and to Marie-PauleBarrillat and Barbara Swiderski for help with electron microscop y.
References
1 Allen S. L. (1967) : Cytogenetics of genomic exclusion inTetrahymena. Genetics, 55, 797-822.
2 Bruns P. J. and Brussard T . B. (1974): Pair formation inTetrahymena pyriformis, an inducible developmental system.J. Exp, Zool., 188,337-344.
3 Cohen J., Adoutte A., Grandchamp S., Houdebine L. M. andBeisson]. (1982) : Immunocytochemical study of microtubular st ructures throughout the cell cycle of Paramecium. Biol.Cell , 44, 35- 44 .
4 Cole E. and Frankel J. (199 1): Conjugal blocks in Tetrahymena pattern mutants and their cytoplasmic rescue. 11. janusA. Dev. Biol. , 148,420-428.
5 Doerder F. P. and Shab atu ra S. K. (1980): Genomic exclu sionin Tetrahymena thermophila cytoge netic fluorometric stu dy.Dev. Genet. , 1, 205-21 8.
6 Frankel]. (196 9): Participation of th e undulatin g membranein th e formation of the oral replacement primordium ofTetrahymena pyriform is. J. Protozool. , 16,26-35.
7 Frankel J. (1989) : Pattern formation . Ciliate studies andmodels. Oxford University Press, Oxford, New York .
8 Frankel]. and Williams N. E. (1973) : Cortical development inTetrahymena. In : Elliott A. M. (ed.) : Th e biology of Tetrahymena , pp. 375- 409. Dowden, Hutchison and Ross Inc.,Strudsburg, Pa.
9 Fujishima M. and Hiwat ashi K. (1977) : An earl y step ininitiation of fertilization in Paramecium: early micronuclearmigration . ]. Exp. Zool., 201 , 127-134.
Oral Morphogenesis During Conjugation . 369
10 Gaert ig J. and Kaczanowski A. ( 1986): Corr elation betweenth e shortened period of cell pairing during genomic exclu sionand the block in po stransfer nucl ear development in Tetrahymena tbermopbila. Develop. Growth and Differ. , 29,553-562.
11 Gae rt igJ., Kiersno wska M. and Iftode F. (1988): Induction ofhybrid stra ins of Tetrahymena by electro fusion. J. Cell Sci.,89, 253 -261.
12 Gae rtig J. and Fleur y A. (1992 ): Spatiotemporal reorganization of intracyroplasmati c microtubules is associated withnucl ear selection and differenti ation during development alpro cess in the ciliate Tetrahymena thermo phila. Protoplasma,167, 74- 87 .
13 Kaczanowski A. (197 6) : An ana lysis of mp gene affectedmorphogenesis in Tetrahymena pyriformis, syngen 1 (species1).]. Exp. Zool., 196, 215-230.
14 Kaczanowski A. (1978): Gradient s of proliferation of ciliarybasal bodies and determination of the position of oralprimordium in Tetrahymena. ]. Exp . Zool., 204, 417-430.
15 Kaczanowski A. (1992) : Mutation affecting cell separationand macronuclear resorption dur ing conjugation in Tetrahymena ther mo phila. Early expressio n of zygo tic phenotype.Dev. Genetics, 121, 37-45 .
16 Kaczanowski A., Piorrowska 1. and Cleffmann G. (1989):Arrest of micronuclear DN A replication during genomicexclusion produces haploid strai ns. Genetics, 121, 37-45.
17 Martindale D. W. , Allis C. D. and Bruns P. J. (1982) :Co njugation in Tetrahymena thermophila: a temporal analysis of cytological stages. Exp. Cell Res., 140, 227-236.
18 Mayo K. A. and Orias E. (1986): Developmental regul ation ofgene expression in Tetrahymena. Dev. Biol., 11 6,302-3 13.
19 Ng S. F. and Nelsen E. M. (1977): The protargol stainingtechn ique. An improved version for Tetrahym ena pyriformis.Tran s. Amer. Soc., 96, 36 9-376 .
20 Orias E. (1986): Ciliate conjuga tion. In: Gall]. G. (ed.): Themolecular biology of ciliated prot ozoa, pp. 45-H4. AcademicPress, New York.
21 Ruffolo].]. Jr. (1974 ): Critical point drying of protozoancells and other biological specimens for scanning electronmicroscopy. Trans Am. Micros. Soc., 93, 124-131.
22 Tsunemoto M., Numata 0., Sugai T. and Watanabe Y.(198 8): Analysis of oral replacem ent by scanning electronmicroscop y and immunofluorescence microscopy in Tetrahy mena tbermopbila during conju gation. Zuo!' Sci. (Tokyo), 5,119-131.
23 Weindruch R. H. and Doerder F. P. (1975): Age dependentmicronuclear deterioration in Tetrahymena pyriformis, syngen 1. Mech. Ageing and Dev., 4, 263- 279 .
24 Weiske-Benner A. and Eckert W. A. (1987): Differentiation ofnucl ear structure during th e sexua l cycle in Tetrahymenathermo phila. Degeneration and autolysis of macro- andmicronuclei degeneration . Differentiation, 34, 1-12.
25 Wenk ert D. and Allis C. D. (1984) : Timing of the appearanceof macronuclear-specific histone variant hv 1 and geneexpression of developing new macronuclei of Tetr ahym enathermophila. J. Ce ll Biol., 98, 21 07- 211 7.
26 Wolfe J. (1982): Th e conjugat ion junct ion of Tetrahymena:its structu re and development. J. Morphol. , 172, 159-178.
27 Wolfe J. (1985): Cytoskeleral reorganization and plasmafusion in conjugating Tetrahymena. ]. Cell Sci., 73, 65 - 85.
28 Wolfe ]. and Grimes G. W. (1979): Tip transformation inTetrahymena: morphogenetic response to interactionsbetw een mating types. ]. Protozo ol., 26, 82-89.
Key words: Tetrahymena thermophila - Conjugation - Morphogenesis
Mauryla Kiersnowska, Dept. of Cyto logy, Zoological Institute, University of Warsaw, Warsaw 00-927, Poland

















![MACROALGA Ulva intestinalis (L.) OCCURRENCE IN …for the Wielkopolska area, two sites of Ulva intestinalis occurrence were de-scribed [Messyasz – in press]. Different oral reports](https://static.fdocuments.pl/doc/165x107/5f21b62502ad3e52be4b1bfa/macroalga-ulva-intestinalis-l-occurrence-in-for-the-wielkopolska-area-two-sites.jpg)
