
Badanie wrażliwości komórek CHO-K1 w odpowiedzi na ...
17
1 ZESPOŁOWY PROJEKT STUDENCKI 2018/19 pod kierunkiem dr Urszuli Kaźmierczak oraz dr hab. Zygmunta Szeflińskiego Badanie wrażliwości komórek CHO-K1 w odpowiedzi na napromienienie wiązką jonów węgla C-12 Dawid Łyko, Mateusz Wróblewski, Martyna Śpiewak, Marta Senkowska, Joanna Zgardzińska
Transcript of Badanie wrażliwości komórek CHO-K1 w odpowiedzi na ...

1
ZESPOŁOWY PROJEKT STUDENCKI 2018/19
pod kierunkiem dr Urszuli Kaźmierczak
oraz dr hab. Zygmunta Szeflińskiego
Badanie wrażliwości komórek CHO-K1 w odpowiedzi na napromienienie wiązką jonów węgla C-12
Dawid Łyko, Mateusz Wróblewski, Martyna Śpiewak,
Marta Senkowska, Joanna Zgardzińska

2
WPROWADZENIE
Powszechnie widomo, że celem terapii zmian nowotworowych jest zniszczenie patologicznych komórek guza przy jednoczesnym zminimalizowaniu wpływu leczenia na stan komórek prawidłowych. Obecnie wyróżniamy trzy główne metody działania: chirurgiczne usunięcie tkanek nowotworowych, chemioterapię oraz radioterapię. Wykorzystuje się je indywidualnie, jak również łączy ze sobą, decydując się na tak zwaną terapię skojarzoną [1]. Dwie pierwsze ze wspomnianych metod obarczone są dużym ryzykiem naruszenia tkanek zdrowych ze względu na ich mechaniczne uszkodzenie lub działanie środków farmakologicznych na cały organizm. Radioterapia konwencjonalna, wykorzystująca promieniowanie jonizujące do zniszczenia komórek nowotworowych, zdaje się zmniejszać negatywne skutki leczenia.
Rozwój teleradioterapii w ostatnich latach sprzyja ograniczeniu dawki promieniowania dostarczanej do obszarów tkanek zdrowych. Schemat leczenia jest jednak uzależniony od wielu czynników, w głównej mierze od rodzaju i umiejscowienia zmiany nowotworowej ale również od stanu zdrowia pacjenta [1]. W radioterapii konwencjonalnej wykorzystywane są źródła promieniowania emitujące fotony lub elektrony [1;2], a charakterystyka oddziaływania takich wiązek z materią nie pozwala na całkowite wyeliminowanie występowania dawek w tkankach niezmienionych chorobowo. Naturalne więc wydaje się zwrócenie ku innym źródłom promieniowania jonizującego celem dalszej poprawy jakości leczenia. Priorytetem stało się zatem rozwijanie i badanie skuteczności radioterapii hadronowej, gdzie źródłem promieniowania jonizującego są przyspieszone, ciężkie cząstki naładowane, takie jak protony czy jony węgla [1,3–6]. Właściwości fizyczne takich cząstek oraz ich sposób oddziaływania z materią pozwala na uzyskanie bardziej korzystnego rozkładu dawki podczas leczenia oraz większej ochrony zdrowych tkanek niż w przypadku radioterapii konwencjonalnej.
Z przedstawionych powyżej powodów, w ramach Zespołowego Projektu Studenckiego, zajęto się badaniem przeżywalności komórek CHO-K1 napromienionych wiązką jonów C-12.
1. WSTĘP TEORETYCZNY W radioterapii wyleczenie zmiany nowotworowej polega na zabiciu komórek guza, w głównej mierze poprzez uszkodzenie ich materiału DNA. Do utraty zdolności proliferacji (czyli namnażania się) i rozpadu nici DNA mogą prowadzić różne czynniki.
1.1. Skutki oddziaływania promieniowania jonizującego na komórki
Promieniowanie jonizujące może uszkadzać strukturę DNA zarówno w sposób bezpośredni, jak i pośredni. Nie mniej jednak o końcowym losie komórki poddanej wpływowi promieniowania decyduje jej aktywność proliferacyjna oraz zdolności do naprawy uszkodzeń DNA.

3
1.1.1. Zależność od cyklu komórkowego
W cyklu komórkowym, który obejmuje replikację oraz podział komórki, wyróżnia się dwa etapy, czyli interfazę i mitozę. Najdłużej trwająca część cyklu - interfaza składa się z trzech faz:
• faza 𝐺" - to czas, w którym komórka syntezuje różne enzymy potrzebne w kolejnej fazie cyklu;
• faza 𝑆 - w tym czasie w komórce następuje replikacja, czyli podwojenie ilości DNA, czego wynikiem jest powstanie chromatyd siostrzanych;
• faza 𝐺$ - ta faza charakteryzuje się zwiększoną syntezą białek potrzebnych później w procesie mitozy;
Najistotniejszym etapem cyklu komórkowego jest mitoza, inaczej faza M. To w tej fazie następuje podział jądra komórkowego oraz cytoplazmy. Schemat całego cyklu przedstawiono na Rysunku 1.
Rysunek 1. Schemat cyklu komórkowego.[13]
Podczas napromieniania duże znaczenie odgrywa to, w jakiej fazie cyklu aktualnie jest komórka. W trakcie każdej z faz komórka wykazuje inną promieniowrażliwość. Komórki są najbardziej wrażliwe w późnej fazie 𝐺$ oraz M, zaś najmniej podczas fazy 𝑆. Nie do końca wiadomo jakie są przyczyny różnej wrażliwości, lecz najprawdopodobniej jest to związane ze zmianami organizacji chromatyny.
1.1.2. Rodzaje uszkodzeń komórek i ich naprawa Promieniowanie jonizujące w głównej mierze oddziałuje na jądro komórkowe i zawarte w nim DNA. W wyniku bezpośredniej lub pośredniej (przez wolne rodniki) depozycji energii promieniowanie powoduje uszkodzenia jednej nici DNA, pęknięcia obu nici a także uszkodzenia zasad. Losy napromienianej komórki zależą od jej zdolności do naprawy uszkodzeń DNA. Komórka uruchamia różne procesy naprawcze w zależności od rodzaju uszkodzeń. Rozróżniamy dwa główne uszkodzenia nici DNA poddanej wpływowi promieniowania jonizującego:
• pęknięcia jednoniciowe - są jednoznaczne z uszkodzeniem tylko jednej z dwóch nici tworzących helisę, druga nić może wtedy stanowić matrycę w trakcie naprawy;
• pęknięcia podwójnonciowe -to co najmniej dwa pęknięcia jodnoniciowe na obu niciach występujące w odległości mniejszej niż 10 par zasad.
Dzięki różnym mechanizmom uszkodzenia mogą być bezbłędnie naprawiane, co pozwala komórce na przeżycie. Naprawa pojedynczych pęknięć lub uszkodzonych zasad z reguły przebiega szybciej i sprawniej niż w przypadku podwójnoniciowych pęknięć.

4
Kiedy jednak mechanizmy naprawy nie działają dobrze może dojść do mutacji, czyli trwałych zmian materiału genetycznego. Mutacją są na przykład aberracje chromosomowe, czyli trwałe zmiany struktury chromosomów wywołane głównie przez podwójnoniciowe pęknięcia. Charakteryzują się zmianami w liczbie lub strukturze chromosomów, a najczęstszymi zmianami są translokacje (przemieszczenia fragmentu chromosomu w tym samym lub do innego chromosomu) oraz dicentryki (połączenie dwóch chromosomów). Podczas podziałów komórki z uszkodzonych chromosomów mogą powstawać również mikrojądra. Różne przyadki zachowania komórki po napromienieniu przedstawiono na Rysunku 2.
Rysunek 2. Zachowanie komórki po ekspozycji na promieniowanie.[13]
1.1.3. Wpływ LET promieniowania na RBE LET - liniowy przekaz energii. To współczynnik służący do określania ilości zdeponowanej energii promieniowania jonizującego na jednostkę drogi w absorbencie. Każdy rodzaj promieniowania charakteryzuje się innym współczynnikiem LET. Ciężkie cząstki mają wysoki LET ze względu na występowanie dużej ilości oddziaływań z cząsteczkami materii przez którą przechodzą, z kolei fotony mają niski współczynnik, ponieważ oddziałują z mniejszą ilością cząsteczek na jednostkę drogi. RBE - względna skuteczność biologiczna. Współczynnik ten służy do określania skuteczności niszczenia nowotworów danego rodzaju promieniowania względem promieniowania X.
Promieniowanie o niskim 𝐿𝐸𝑇 powoduje w znakomitej większości jednoniciowe pęknięcia nici DNA, które są niewystarczające do uśmiercenia komórek, ponieważ istnieją wyspecjalizowane mechanizmy naprawcze zdolne do odbudowy pojedynczej nici [1,5]. Z kolei wiązki jonów węgla, które charakteryzują się wysokim LET, „bombardując” komórkę powodują jej bezpośrednie

5
uszkodzenia, doprowadzając do dwuniciowych pęknięć DNA w jednym akcie jonizacji, co w sposób decydujący przekłada się na wartość RBE [1,4,5,9].
1.1.4. Oddziaływanie ciężkich jonów z materią Charakterystyka ciężkich cząstek naładowanych wykorzystywanych w radioterapii hadronowej jest nieco inna niż charakterystyka wykorzystywanych w radioterapii konwencjonalnej elektronów i fotonów. Zasadniczą przewagą cząstek naładowanych w kontekście radioterapii jest ich korzystniejszy rozkład dawki względem głębokości [1,3], gdzie przez dawkę rozumiemy ilość energii zaabsorbowanej przez jednostkę masy materii : 𝐷 = *
+. Porównanie rozkładów dawki względem
głębokości dla fotonów oraz ciężkich cząstek, jak również ich znaczenie w radioterapii przedstawia Rysunek 3.
Rysunek 3. Rozkłady dawki dla jonów węgla oraz fotonów o różnej energii w funkcji głębokości w
ośrodku wodnym (po lewej) [3] oraz ich znaczenie w radioterapii (po prawej).[13]
Jak można zauważyć na Rysunku 3, rozkład dawki w funkcji głębokości, inaczej przebiega inaczej dla fotonów, a inaczej dla ciężkich cząstek. Depozycja energii przez fotony dość szybko rośnie do momentu osiągnięcia wartości maksymalnej, po czym następuje powolny spadek. W przypadku ciężkich jonów obserwuje się początkowo powolny wzrost dawki do chwili w której następuje depozycja znacznej większości energii (wysoki pik) co w rezultacie powoduje spadek dawki do zera. Punkt maksymalnej depozycji dawki dla fotonów jak i dla ciężkich cząstek naładowanych zależy od ich energii, co oznacza, że im większa energia tym dalej zostanie zdeponowana największa dawka.

6
2. MATERIAŁY I METODY
2.1 Linia komórkowa CHO-K1
Linia komórkowa CHO-K1 wyizolowana została w wyniku biopsji jajnika dorosłego chomika chińskiego. Komórki te posiadają 21 chromosomów [12,13], a ich cykl komórkowy trwa 11,00 ± 3,42 h [13,14]. Hodowane mogą być w monowarstwie oraz wykazują zdolność adhezji (łączenia się ze sobą powierzchniowych warstw ciał fizycznych lub faz), dzięki czemu często wykorzystywane są do badań laboratoryjnych oraz medycznych [13].
2.2 Układ eksperymentalny
Do przeprowadzenia pomiarów w celu określenia promieniowrażliwości linii komórkowej CHO-K1 wykorzystano układ do badań radiobiologicznych znajdujący się w Środowiskowym Laboratorium Ciężkich Jonów Uniwersytetu Warszawskiego (ŚLCJ UW), w którym wiązkę jonów węgla dostarcza cyklotron U-200P. Układ ten podaje szeroką wiązkę równolegle względem podłoża, w stałym, określonym obszarze, dlatego też badany układ umieszczony jest w specjalnym uchwycie przesuwnego robota pomiarowego wykonującego ruchy w kierunku pionowym względem wiązki [13].
Wiązka jonów o energii pierwotnej 48 MeV wprowadzana była jonowodem do komory rozpraszającej o promieniu 8 cm, wewnątrz której zainstalowano złotą tarczę rozpraszającą o grubości 13 mg/cm2. Po rozproszeniu na tarczy wiązka pokonywała drogę 233 cm wzdłuż poziomego jonowodu i przechodziła przez okno wyjściowe wykonane z havaru o grubości 2,3 mg/cm2 [13]. Schemat układu pomiarowego przedstawiono na Rysunku 4 oraz 5.
Rysunek 4. Schemat układu pomiarowego używanego do badań radiobiologicznyh w ŚLCJ UW.[13]

7
Rysunek 4. Układ pomiarowy w przekroju poprzecznym.[13]
Dodatkowo, w układzie znajdują się dwa detektory krzemowe. Jeden z nich umieszczony w komorze, pod kątem 20° względem osi wiązki oraz w odległości 11,8 cm od środka folii rozpraszającej, monitorujący wiązkę bezpośrednio podczas napromieniania próbki. Drugi detektor instalowany jest podczas kalibracji, pod kątem 0° w pozycji próbki. Sygnały z obu detektorów służą do skoordynowania pracy układu dostarczającego wiązkę i robota przesuwającego próbkę tak by cała próbka została równomiernie napromieniona dawką w całym swoim obszarze. Liczba jonów, rejestrowana pod kątem 20° w komorze rozproszeń jest proporcjonalna do liczby jonów zarejestrowanych przez detektor kalibracyjny, co pozwala na określenie jaka dawka została zaabsorbowana w danym obszarze próbki i odpowiednio do ustalonych wytycznych, zatrzymanie napromieniania i przesunięcie próbki względem układu o zadaną odległość. Specjalne oprogramowanie do sterowania robotem podczas napromieniania napisała dr Joanna Czub (Uniwersytet Jana Kochanowskiego w Kielcach) [15].
2.3 Szalki do napromieniania komórek CHO-K1
Ze względu na specyficzną budowę układu źródło-próbka, w którym źródło emituje cząstki równolegle względem podłoża i prostopadle względem próbki oraz mając na uwadze energię uzyskiwanej wiązki, do badań wykorzystano szlaki własnej konstrukcji. Szklane lub plastikowe szalki Petriego, zwyczajowo stosowane w tego typu eksperymentach nie mogły zostać użyte, bowiem nie zapewniały dostatecznej szczelności wymaganej podczas ekspozycji w pionowej orientacji, uwarunkowanej budową układu. Ponadto przy uzyskiwanej energii wiązek, cząstki deponowałaby by część lub jej całość w materiale szalki, a sam badany materiał nie zostałby napromieniowany lub dawka przez niego pochłonięta odbiegałaby od zaplanowanej.
„Do konstrukcji szalek wykorzystane zostały plastikowe krążki o wysokości 10 mm, powstałe z pocięcia polipropylenowej rury. Ich średnica wewnętrzna wynosiła 40 mm, zaś zewnętrzna 63 mm. Dzięki temu powstałe z nich szalki miały 12,57 ml pojemności. Dno szalki stanowiła poliestrowa folia mylar o grubości 6 µm, przyklejona do wyszlifowanego i czystego krążka za pomocą taśmy dwustronnej. Ponieważ pierwotnie taśma pokrywała nie tylko grubą ściankę krążka, ale również dno powstającej szalki, niepotrzebne jej fragmenty były usuwane. Taśma, stanowiąca w danej chwili dno szalki, była odcinana skalpelem, przesuwanym wzdłuż wewnętrznej ścianki krążka. Jej nadmiar,

8
znajdujący się na zewnątrz krążka był usuwany dopiero po przyklejeniu folii mylar. Chwilowe pozostawienie fragmentów taśmy klejącej na zewnątrz krążka pomagało bowiem w dokładnym, gładkim przyklejeniu cienkiej folii. Gruba ścianka krążka, w całości pokryta taśmą dwustronną, gwarantowała szczelne przyklejenie foliowego dna. W celu usunięcia ewentualnych zanieczyszczeń, po dokładnym odcięciu zbędnych fragmentów folii, szalka była przemywana 80% etanolem. Do szczelnego zamykania naczynek zastosowano paralm (specjalnie spreparowana parana), dociskany za pomocą plastikowych obręczy. Zestaw takich obręczy przygotowano z rury polipropylenowej o średnicy wewnętrznej mniejszej niż średnica zewnętrzna szalki i wynoszącej 61 mm oraz zewnętrznej 70 mm. Rurę pocięto na krążki o wysokości 7 mm, a następnie każdy z nich dodatkowo wydrążono, powiększając średnicę wewnętrzną do 64 mm na głębokości 5 mm. W ten sposób krążki posiadały dwie średnice wewnętrzne: 64 mm przez 5 mm wysokości oraz 61 mm na głębokość 2 mm. Tym samym uzyskano krawędź wewnętrzną o wymiarach 1,5 x 2 mm. Tak powstała obręcz, ze średnicą wewnętrzną niewiele większą od średnicy zewnętrznej szalki i krawędzią pozwalającą na blokadę głębokości docisku obręczy. W połączeniu z paralmem zapewniała ona bardzo dobre zamknięcie szalki, nawet w pionowym ustawieniu. Wszystkie elementy mogące mieć bezpośredni kontakt z komórkami lub ich pożywką musiały być sterylne. Dlatego też przygotowane szalki oraz wycięte z paralmu kółka były pakowane i przewożone do IChTJ, gdzie poddawano je procesowi sterylizacji radiacyjnej wiązką wysokoenergetycznych elektronów wytwarzaną w akceleratorze Elektronika 10/10 [16]. Szalki do napromieniania oraz krążki z paralmu były przygotowywane każdorazowo na potrzeby przeprowadzanych eksperymentów.”[13]
2.3.1 Przygotowanie próbek biologicznych do napromienienia
W każdej szalce wysiano 1 mln komórek CHO-K1, umieszczonych w umieszczane w
pożywce 5A McCoy (Gibco, USA) zawierającej 10% płodowej surowicy bydlęcej (ang. Fetal Bovine Serum, FBS), 1% penicyliny i streptomycyny [13,17]. Po wysianiu, szalki uszczelniano paralmem i plastikową obręczą, a tak przygotowaną próbkę przekładano do plastikowego naczynka o średnicy 10cm, chroniącego dno szalki przed uszkodzeniem w trakcie przechowywania i transportu. Przed umieszczeniem próbek w inkubatorze, usuwano zabezpieczający palarm i obręcz. Inkubacja odbywała się w ściśle określonych warunkach, przy zachowaniu temperatury 37 ℃ i stężenia CO2 na poziomie 5%. W tym czasie komórki przytwierdzały się do dna szalki w monowarstwie[13].
Rysunek 5. Szalka z badaną próbką, w pełni przygotowaną do eksperymentu [13].
Bezpośrednio przed napromienianiem szalki były całkowicie wypełniane pożywką oraz ponownie zabezpieczane w sposób identyczny jak po wysianiu. Podczas procedury zabezpieczającej dbano by

9
nie powstawały pęcherzyki powietrza zamknięte wewnątrz pożywki. Sytuacja taka mogłaby bowiem zaburzyć jednorodny rozkład dawki w napromienianym obszarze.
2.3.2 Napromienianie
W hali eksperymentalnej tuż przed napromienianiem, wyjmowano szalkę z plastikowego naczynka ochronnego i starannie mocowano w uchwycie tak by dno szalki zwrócone było równolegle do wylotu jonowodu. Dodatkową stabilizacje zapewniała cienka gumka poprowadzona po przekątnej między szalką a uchwytem. Napromienianie komórek rozpoczynało się w momencie wprowadzenia wiązki jonów do układu. Dawka promieniowania i ruch szalki względem wiązki został uprzednio zaplanowany w oprogramowaniu, o którym mowa w podrozdziale 2.2. Wiązka jonów by ulec absorbcji w badanym materiale biologicznym, musiała pokonać warstwę powietrza oraz dna szalki. W zależności od zaplanowanej dawki, czas napromieniania ulegał zmianom, jednak nie przekraczał 15 minut, ponieważ tak długotrwały proces mógłby spowodować odklejanie się kolonii od dna szalki i w efekcie uzyskane wyniki byłyby niemiarodajne [13]. Po napromienieniu szalkę demontowano z uchwytu, przy czym zachowywano szczególną ostrożność by nie doprowadzić do skażenia materiału biologicznego. Następnie umieszczano ją wewnątrz plastikowej szalki zabezpieczającej i opisywano.
Każda z przygotowanych próbek napromieniana była w jednakowych warunkach, a następnie transportowana do laboratorium, w którym odpowiednio przygotowywano je do przeprowadzenia testu klonogennego.
2.4 Test Klonogenny
Test klonogenny, inaczej przeżywalności, wykonywany jest na komórkach hodowanych in vitro. Pozwala, dla danego rodzaju komórek, określić ich zdolności do proliferacji, a co za tym idzie do tworzenia kolonii. Aby przeprowadzić taki test należy wysiać dostateczną liczbę napromienionych komórek na szalkę, następnie są one inkubowane w odpowiednich warunkach, a po tym czasie dokonywana jest analiza, która polega na zliczeniu utworzonych kolonii. Za kolonię uznawane jest zbiorowiska co najmniej 50 komórek. Test przeżywalności jest powszechnie stosowany, ponieważ jest dokładny nawet gdy ustosowane są niskie dawki promieniowanie jonizującego (0,05 Gy).
Do oceny zdolności proliferacyjnych komórek wykorzystuje się parametr 𝑃𝐸 czyli wydajność klonowania zdefiniowaną jako :
𝑃𝐸 = ./0123567893ł;0<=6.6>//./01237;8/3>;0<=6?óAB=
,
Przeżywalność określa się parametrem 𝑆𝐹 (ang. survival fraction):
𝑆𝐹 =wydajnośćklonowanianapromienianejpróbkiwydajnośćklonowaniakomórekkontrolnych
,
gdzie komórki kontrolne są komórkami nienapromieniowanymi.

10
2.5 Metody pomiaru
2.5.1. Analiza obrazów w programie ImageJ
W pracy analizowano serię obrazów szalek z komórkami, w tym celu korzystano z ogólnodostępnego pakietu BioVoxxel w programie ImageJ.
Analiza przebiegała w kolejnych krokach:
• dopasowanie kontrastu obrazu i binaryzacja • rozmycie obrazu przy pomocy filtra "Gaussian Blur" • sprawdzenie "Treshold Check"(narzędzie z pakietu BioVoxxel), na podstawie którego
następnie • wybierano odpowiedni treshold przy pomocy narzędzia ”Autotreshold” • z pakietu "BioVoxxel"wybiearno ”Extended Particle Analyzer”, w którym ustawiano wiele
parametrów (największe znaczenie miała wielkość zliczanych kolonii dostosowywana w oknie „Area”)
Na Rysunku 6. Przedstawiono poszczególne etapy analizy liczby kolonii komórkowych. Na Rysunku 7 widać które kolonie zostały policzone, w przypadku dużej liczby komórek nienumerowanych zmieniano wartości minimalnej powierzchni w oknie „Area”, lub doliczano pozostałe komórki manualnie.
Rysunek 6. Początkowe kroki analizy obrazów (binaryzacja).

11
Rysunek 7. Obraz analizowany pakietem BioVoxxel.
W celu określenia przeżywalności komórkowych wyznaczono wydajność klonowania komórek jako stosunek liczby zliczonych kolonii do liczby wysianych pierwotnie komórek ze wzoru (3). Wydajność klonowania komórek kontrolnych uśredniono, by otrzymać wynik dla całej próby. W kolejnym kroku wyznaczono przeżywalność komórek jako iloraz wydajności klonowania komórek napromienianych, oznaczonych jako e, oraz średniej wydajności klonowania komórek kontrolnych k ze wzoru (4). Wartości te uśredniono dla każdej z dawek i otrzymano średnią przeżywalność komórkową po napromienieniu daną dawką. Wartości te przedstawiono na wykresie zależności przeżywalności SF od dawki D (Rysunek 8).

12
Rysunek 8. Wykres zależność frakcji przeżywalności SF od dawki D.
Dane doświadczalne dla poszczególnych dawek: 0,1 Gy, 1 Gy oraz 4 Gy przedstawiono w Tabelach 1-3. Analizowane szalki podpisane są w następujący sposób: e1a, e1b, e1c, gdzie e1 to numer napromienionej szalki , zaś a, b, c to oznaczenie powtórzenia technicznego wysiewania komórek. Oznacza to, że napromieniono np. 1 milion komórek, które znajdowały się w szalce E1. Ponieważ jest to zbyt duża liczba komórek, żeby możliwe było przeprowadzenie tygodniowej inkubacji materiału biologicznego w danej szalce, po napromienieniu policzono komórki w szalce i określoną ich liczbę (np. 1000) wysiano do nowej szalki - np. ela. Nie chcąc tracić pozostałych, napromienionych komórek, wykonano jeszcze tzw. powtórzenia techniczne - z tej samej napromienionej szalki wysiano komórki do kolejnych pustych szalek, tworząc w ten sposób np. elb i elc.
Do szalek kontrolnych k wysiewano po 300 komórek nienapromienionych, natomiast do szalek e w zależności od dawki, którą zostały napromienione komórki – wysiewano odpowiednio 600 komórek napromienionych dawką 0,1Gy, 1000 komórek napromienionych dawką 1 Gy oraz 6000 komórek napromienionych dawką 4Gy.
13
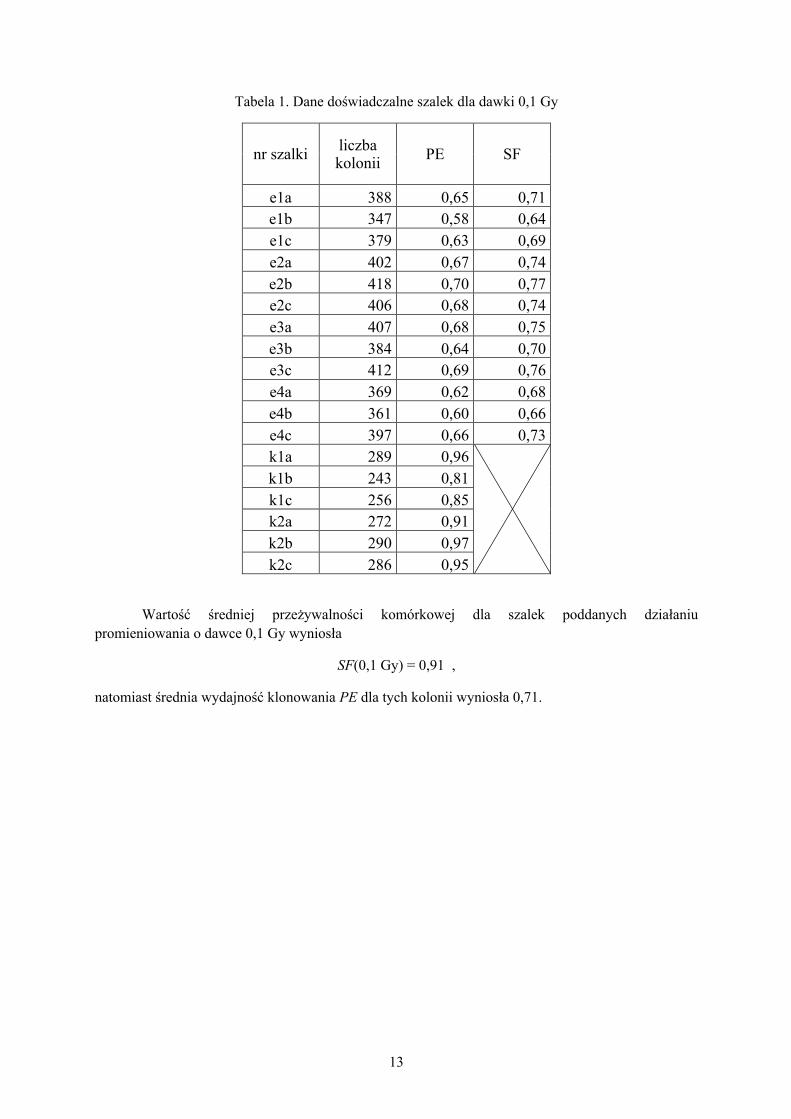
Tabela 1. Dane doświadczalne szalek dla dawki 0,1 Gy
nr szalki liczba kolonii PE SF
e1a 388 0,65 0,71 e1b 347 0,58 0,64 e1c 379 0,63 0,69 e2a 402 0,67 0,74 e2b 418 0,70 0,77 e2c 406 0,68 0,74 e3a 407 0,68 0,75 e3b 384 0,64 0,70 e3c 412 0,69 0,76 e4a 369 0,62 0,68 e4b 361 0,60 0,66 e4c 397 0,66 0,73 k1a 289 0,96
k1b 243 0,81 k1c 256 0,85 k2a 272 0,91 k2b 290 0,97 k2c 286 0,95
Wartość średniej przeżywalności komórkowej dla szalek poddanych działaniu promieniowania o dawce 0,1 Gy wyniosła
SF(0,1 Gy) = 0,91 ,
natomiast średnia wydajność klonowania PE dla tych kolonii wyniosła 0,71.

14
Tabela 2. Dane doświadczalne szalek dla dawki 1 Gy
nr szalki liczba kolonii PE SF
e1a 408 0,41 0,51 e1b 428 0,43 0,54 e1c 323 0,32 0,40 e2a 415 0,42 0,52 e2b 405 0,41 0,51 e2c 353 0,35 0,44 e3a 422 0,42 0,53 e3b 414 0,41 0,52 e3c 387 0,39 0,48 e4a 319 0,32 0,40 e4b 302 0,30 0,38 e4c 403 0,40 0,50 k1a 284 0,95
k1b 268 0,89 k1c 249 0,83 k2a 294 0,98 k2b 277 0,92 k2c 285 0,95
Wartość średniej przeżywalności komórkowej dla szalek poddanych działaniu promieniowania o dawce 1 Gy wyniosła
SF(1 Gy) = 0,92 ,
natomiast średnia wydajność klonowania PE dla tych kolonii wyniosła 0,48.

15
Tabela 3. Dane doświadczalne szalek dla dawki 4 Gy
nr szalki liczba kolonii PE SF
e1a 571 0,10 0,12 e1b 512 0,09 0,11 e1c 514 0,09 0,11 e2a 557 0,09 0,12 e2b 397 0,07 0,08 e2c 427 0,07 0,09 e3a 516 0,09 0,11 e3b 445 0,07 0,09 e3c 510 0,09 0,11 e5a 501 0,08 0,10 e5b 521 0,09 0,11 e5c 506 0,08 0,11 e6a 417 0,07 0,09 e6b 493 0,08 0,10 e6c 491 0,08 0,10 k1a 271 0,90
k1b 253 0,84 k1c 289 0,96 k3a 274 0,91 k3c 268 0,89
Wartość średniej przeżywalności komórkowej dla szalek poddanych działaniu promieniowania o dawce 0,1 Gy wyniosła
SF(0,1 Gy) = 0,90 ,
natomiast średnia wydajność klonowania PE dla tych kolonii wyniosła 0,10.
3.2 Dyskusja i wnioski
Wyznaczone doświadczalnie wartości testu klonogennego dla szalek poddanych działaniu promieniowania o dawkach 0,1 Gy, 1 Gy i 4 Gy wyniosły kolejno: 𝑆𝐹(0,1 Gy) = 0,71 , 𝑆𝐹(1 Gy) = 0,48 , 𝑆𝐹(4 Gy) = 0,09, co było zgodne z oczekiwaniami, że wraz ze wzrostem dawki promieniowania, przeżywalność napromienianych komórek będzie spadać.
Wiedząc, że wartość 𝑃𝐸 dla większości linii komórkowych waha się w zakresie 80-90% [18], co jest zgodne z uzyskanymi doświadczalnie wartościami, można stwierdzić, że eksperyment przebiegł poprawnie, a przedstawione metody są skuteczne.
Spadek 𝑆𝐹 wraz ze wzrostem dawki promieniowania charakterystyczny jest również dla wiązek konwencjonalnych, jednak charakter uzyskiwanej krzywej jest zgoła odmienny i wskazuję na wyższą skuteczność biologiczną wiązek hadronowych. Mianowicie uzyskany na podstawie danych rysunek 8 ukazuje, że kształt krzywej przeżywalności jest zbliżony do linii prostej,

16
zatem współczynnik przeżycia komórek jest wykładniczą funkcją dawki. Jest on zbliżony do radiobiologicznego modelu jednotarczowego, którego teoria zakłada, że już pojedyncze trafienie w tarcze komórki powoduję utratę klonogenności lub śmierć.
IV.BIBLIOGRAFIA
[1] Malicki J, Ślosarek K, Via Medica. Planowanie leczenia i dozymetria w radioterapii. T. 2 T. 2. Gdańsk: Via Medica; 2018.
[2] Kukołowicz PF, Polska Akademia Nauk, Komitet Fizyki Medycznej R i DO, Polskie Towarzystwo Fizyki Medycznej. Charakterystyka wiązek terapeutycznych stosowanych w radioterapii. Wrocław: Komitet Fizyki Medycznej, Radiobiologii i Diagnostyki Obrazowej PAN : Polskie Towarzystwo Fizyki Medycznej; 2017.
[3] Kraft, G. (2005). Terapia nowotworów przy użyciu ciężkich jonów. Verein zur Förderung der Tumortherapie mit Schweren Ionen eV, Darmstadt.
[4] Ebner DK, Kamada T. The Emerging Role of Carbon-Ion Radiotherapy. Front Oncol. 2016 Jun;6. DOI: 10.3389/fonc.2016.00140
[5] Ohno T. Particle radiotherapy with carbon ion beams. EPMA J. 2013 Dec;4(1). DOI: 10.1186/1878-5085-4-9
[6] Durante M, Loeffler JS. Charged particles in radiation oncology. Nat Rev Clin Oncol. 2010 Jan;7(1):37–43.
[7] Glowa C, Peschke P, Brons S, Neels OC, Kopka K, Debus J, et al. Carbon ion radiotherapy: impact of tumor differentiation on local control in experimental prostate carcinomas. Radiat Oncol. 2017 Nov;12(1):174.
[8] Raju MR, Carpenter SG. A heavy particle comparative study. Part IV: acute and late reactions. Br J Radiol. 1978 Sep;51(609):720–7.
[9] Hamada N, Imaoka T, Masunaga S, Ogata T, Okayasu R, Takahashi A, et al. Recent Advances in the Biology of Heavy-Ion Cancer Therapy. J Radiat Res (Tokyo). 2010;51(4):365–83.
[10] Ando K, Kase Y. Biological characteristics of carbon-ion therapy. Int J Radiat Biol. 2009 Jan;85(9):715–28.
[11] Cui X, Oonishi K, Tsujii H, Yasuda T, Matsumoto Y, Furusawa Y, et al. Effects of Carbon Ion Beam on Putative Colon Cancer Stem Cells and Its Comparison with X-rays. Cancer Res. 2011 May;71(10):3676–87.
[12] Xu X, Nagarajan H, Lewis NE, Pan S, Cai Z, Liu X, et al. The genomic sequence of the Chinese hamster ovary (CHO)-K1 cell line. Nat Biotechnol. 2011 Aug;29(8):735–41.
[13] Kaźmierczak Urszula. Dawka lokalna i jej rola w biologicznej odpowiedzi linii komórkowych ssaków in vitro, Rozprawa doktorska,Wydział Fizyki Instytut Fizyki Doświadczalnej Zakład Fizyki Jądrowej, Warszawa. 2015

17
[14] Vasila Packeer Mohamed. Chinese hamster ovary (CHO-K1) cells expressed native insulin-like growth factor-1 (IGF-1) gene towards efficient mammalian cell culture host system. Afr J Biotechnol. 2011 Dec;10(81). DOI: 10.5897/AJB11.2743
[15] Czub J. Biologiczne oddziaływanie ciężkich jonów o niskiej energii,Rozprawa doktorska, Uniwersytet Humanistyczno-Przyrodniczy Jana Kochanowskiego w Kielcach, 2011. 2011
[16] Strona internetowa Instytutu Chemii i Techniki Jądrowej [dostęp: 14.08.2019], [Internet]Available from: http://www.ichtj.waw.pl/drupal/?q=node/233.
[17] Kaźmierczak U, Banaś D, Braziewicz J, Buraczewska I, Czub J, Jaskóła M, et al. Investigation of the bystander effect in CHO-K1 cells. Rep Pract Oncol Radiother. 2014 May;19:S37–41.
[18] Munshi A, Hobbs M, Meyn RE. Clonogenic Cell Survival Assay. Chemosensitivity. New Jersey: Humana Press; 2005; pp 021–8.


















