b%& Ý [+X$ ¤ `# C b $?(ý+7 K b É ß Ç Ý 8 B... · 9× Ø ^0«) w e s s6Û @ µ þ ï 8 k s ~...
11
WesとMiloの相乗効果で腫瘍微小環境の 免疫細胞集団のプロファイルを作成 イントロダクション ガン免疫療法は劇的な治療効果をもたらしており、腫 瘍の完全な消失もしくは腫瘍縮小が報告され、癌治療 において新しい時代への扉が開かれました。しかしな がら、治療効果のあった患者数は少数であることから、 癌に対する免疫反応をより深く理解することがより重 要になってきました。このため、蓄積されてきた研究 や事実から、腫瘍微小環境 A (tumor microenvironment: TME)の不均一性が、免疫応答の相違の主な原因であ ることが示唆されています 1 。 TMEは、がん細胞や、非がん性および免疫細胞の統合 ネットワークであり、その相互作用により腫瘍の不均 一性、転移の広がり、そして後天的な薬剤耐性が促進 されます。特に、リンパ球、マクロファージ、樹状細 胞などのTMEにおける白血球の浸潤 B は重要な予後予 測因子 C であり、そしてがん免疫療法において期待す る治療効果を妨害する主な障害要因としても認識され ています 2–4 。大まかには、浸潤性のCD8+リンパ球 D は 炎症性TMEの特徴やより良い治療効果と関連がありま す。 その期待する治療効果を維持するために、CD8+ T細 胞動員の陰に潜む細胞決定因子を解明することが必 要です。次に、抑制性免疫細胞とサブセット E が存在 し、これらは一般に、腫瘍特異的な免疫反応を抑制 する働きをします。すなわち今後の複数手段での治 療戦略において魅力的なターゲットでもあります。 TME内に存在する免疫細胞の組成と機能のプロファイ リングによって、免疫療法の治療効果が改善されつつ あり、新しい治療標的や戦略が明らかになるかもしれ ません。しかし、サンプル数の制限や、免疫細胞の特 徴である複雑さと不均一性により、TMEの研究は特に 困難な領域で、より深い分析のための最新のツールが 必要になります。このアプリケーションノートでは、 Wes™とMilo™がどのように連携して、まず、1)サン プルにはどのような種類の免疫細胞が存在するか、そ して、2)何パーセントの細胞が特定の免疫細胞サブタ イプなのかという重要な問いかけに答えるのかを示し ます。このワークフローは、各サンプルを集団レベル およびシングルセルレベルで分析し、結果を得るのに 費やす時間を現在使用している従来のアプローチより も大幅に短縮し、またサンプルを大幅に節約します。 アプリケーションノート
Transcript of b%& Ý [+X$ ¤ `# C b $?(ý+7 K b É ß Ç Ý 8 B... · 9× Ø ^0«) w e s s6Û @ µ þ ï 8 k s ~...

WesとMiloの相乗効果で腫瘍微小環境の
免疫細胞集団のプロファイルを作成
イントロダクション
ガン免疫療法は劇的な治療効果をもたらしており、腫
瘍の完全な消失もしくは腫瘍縮小が報告され、癌治療
において新しい時代への扉が開かれました。しかしな
がら、治療効果のあった患者数は少数であることから、
癌に対する免疫反応をより深く理解することがより重
要になってきました。このため、蓄積されてきた研究
や事実から、腫瘍微小環境A(tumor microenvironment:
TME)の不均一性が、免疫応答の相違の主な原因であ
ることが示唆されています1。
TMEは、がん細胞や、非がん性および免疫細胞の統合
ネットワークであり、その相互作用により腫瘍の不均
一性、転移の広がり、そして後天的な薬剤耐性が促進
されます。特に、リンパ球、マクロファージ、樹状細
胞などのTMEにおける白血球の浸潤Bは重要な予後予
測因子Cであり、そしてがん免疫療法において期待す
る治療効果を妨害する主な障害要因としても認識され
ています2–4。大まかには、浸潤性のCD8+リンパ球Dは
炎症性TMEの特徴やより良い治療効果と関連がありま
す。
その期待する治療効果を維持するために、CD8+ T細
胞動員の陰に潜む細胞決定因子を解明することが必
要です。次に、抑制性免疫細胞とサブセットEが存在
し、これらは一般に、腫瘍特異的な免疫反応を抑制
する働きをします。すなわち今後の複数手段での治
療戦略において魅力的なターゲットでもあります。
TME内に存在する免疫細胞の組成と機能のプロファイ
リングによって、免疫療法の治療効果が改善されつつ
あり、新しい治療標的や戦略が明らかになるかもしれ
ません。しかし、サンプル数の制限や、免疫細胞の特
徴である複雑さと不均一性により、TMEの研究は特に
困難な領域で、より深い分析のための最新のツールが
必要になります。このアプリケーションノートでは、
Wes™とMilo™がどのように連携して、まず、1)サン
プルにはどのような種類の免疫細胞が存在するか、そ
して、2)何パーセントの細胞が特定の免疫細胞サブタ
イプなのかという重要な問いかけに答えるのかを示し
ます。このワークフローは、各サンプルを集団レベル
およびシングルセルレベルで分析し、結果を得るのに
費やす時間を現在使用している従来のアプローチより
も大幅に短縮し、またサンプルを大幅に節約します。
アプリケーションノート

2
WESとMILO : 積極的な連携活用法
Wesで行うSimple Western™アッセイは、キャピラリーを使った完全自動のイムノアッセイで、従来の時間がかかり、
同じ作業を繰り返しおこなうウエスタンブロッティングの多くの課題を解決します。従来法からアッセイを移行するの
は簡単です; 蛋白質のサイズ(分子量)ベースの分離、信頼性の高い標的タンパク質の検出、そして真に定量性のあるデー
タを得て、実験的証拠に基づいた正しい結論を下すことができます。サンプル、抗体、試薬を専用アッセイプレートに
分注し、Wesにキャピラリカートリッジとアッセイプレートをセットしてスタートボタンを押すだけです!わずか3時間
で、標的タンパク質の発現量に関して、最大25サンプルのサイズベースで分離した定量データを取得できます。Wesを
使用すると、サンプルに含まれる免疫細胞の種類を明らかにしたり確認したり、集団内の細胞型特異的マーカーの発現
量を定量化することもできます。発現量の低いタンパク質マーカーもWesにお任せください。ピコグラムレベルの感度
を得るのに必要なサンプル量はわずか3 µLです。Wesは標的タンパク質を確実に検出するだけでなく、貴重なサンプルを
節約し、より少ない労力でより多くのデータを得ることができます。
MiloでSingle-Cell Western™を行うと、サンプルをさらに掘り下げてタンパク質発現の不均一性を測定し、またサンプル
集団内で何パーセントの細胞が、特定の標的タンパク質発現の組み合わせや表現型に特異的かも定量化することができ
ます。Miloは、マルチプレックス(複数同時検出)の要求にも答えます。各ウェルで最大12個の標的タンパク質を検出する
ことができ、TME内の細胞間のばらつきを解析します。わずか10,000細胞の細胞懸濁液を専用のscWestチップにロード
し、キャプチャした細胞を溶解すると、Miloは各シングルセルライセートを約1分間、サイズベースでSDS-PAGE分離し、
各標的タンパク質を明確で定量的なピークとして検出します。重要なことに、WesとMiloは両方ともオープンプラットフ
ォームであるため、ウェスタン検証済みの一次抗体があれば、任意の細胞型マーカーをスクリーニングすることができ
ます。研究者の時間を節約するため、R&D SystemsおよびNovus Biologicalsの提供する抗体に関してはSimple Western
およびSingle-Cell Westernの両方で使用できるかどうかのスクリーニングと検証を行っています。
他のマルチパラメーター法よりもなぜWESとMILOを使うのか? 不均一なTMEの細胞集団の分析には他のマルチパラメーター法でも十分かもしれませんが、WesとMiloのタッグチームは、
細胞タイプの識別と特性情報をアッセイの感度や精度を損なうことなく取得でき、実験台で過ごす時間を大幅に短縮し
ます! さらに、装置のノズルやチューブの詰まり、高度なクリーニングプロトコル、必要なレーザーキャリブレーション
を対処するために、高度な訓練を受けた専門家が機器を操作したり、面倒な日々のメンテナンスを行ったりする必要は
ありません。そしてWesとMiloを使ってサイズベースでタンパク質を分離すると、同じ抗体に結合したバリアント/アイ
ソフォームへのオフターゲット結合に別れを告げることができます—これは例えば、フローサイトメトリーでよく問題
になります。フロー検証済み抗体の探索や、複雑なパネル、アッセイの確立に時間を費やすのをやめましょう。Wesと
Miloを使用すると、Wesで自動化されたシンプルウエスタンイムノアッセイをすることで、シンプルで洗練された細胞集
団ベースの測定ができ、選択した細胞集団をMiloでさらに分析して、ターゲットの局在化に関係なく、選択的かつ定量的
なシングルセルレベルのタンパク質発現情報を得ることができます。FACSで高濃度にソートした後、フローサイトメー
ターに戻すのに十分必要な細胞数がない時は、Miloをタンパク質発現解析に使用しましょう。必要な細胞数はたったの
10,000個です!
ワークフロー概略
8人のドナーから採血した血液をプールし、ヒト末梢血単核細胞F(PBMC)を分離しました。WesおよびMiloで分析する前
に、ヒト末梢血単核細胞を樹状細胞G(DC)、ナチュラルキラー(NK)細胞H、または制御性T(Tregs)細胞Iのいずれかに分化
を誘導しました(図 1)。

3
樹状細胞 (DC) ナチュラルキラー細胞 (NK) cells
Regulatory T (Tregs)細胞
CD14+単核細胞用のMagCellect ナイーブCD4+ T細胞用のMagCellect
CellXVivo™ Human Monocyte- derived DC Differentiation Kit
CellXVivo™ Human NK Cell Expansion Kit CellXVivo™ Human Treg
Cell Differentiation Kit
図1. 細胞タイプ特異的な分化および同定の概要。プールしたPBMCをDC(左)、NK細胞(中央)、Treg(右)に分化誘導した。Wesおよび
Miloで分化した同じ細胞サンプルの分化細胞タイプを示すマーカー発現を分析した。CD14+単球Jを単離後、DCへの分化誘導と増殖
により、PBMCはDCに分化した(左)。NK細胞はPBMCから直接増殖した(中央)。ナイーブ(まだ抗原と出会っていない)CD4+ T細胞を
単離後、Tregへの分化誘導と増殖により、PBMCをTregに分化した(右)。
PBMCs, peripheral blood mononuclear cells; DCs, dendritic cells; NK, natural killer; Tregs, regulatory T cells
樹状細胞への分化誘導
樹状細胞(DC)を得るために、まず、MagCellect™ Human CD14+ Cell Isolation Kit (R&D Systems, MAGH105)の製品添付文書
の推奨プロトコルに従って、CD14陽性ヒト単球を選択 (CD14+ PBMC) しました。次に、CellXVivo™ Human Monocyte-
derived Differentiation Kit (R&D Systems, CDK004)に含まれている培地およびサイトカインを使用して、無血清条件下で
CD14+ PBMCを未成熟および成熟樹状細胞(DC)へ分化誘導しました。 得られた樹状細胞(DC)は、下流のアプリケーションに
使用することができます。
ナチュラルキラー細胞への分化誘導
CellXVivo™ Human NK Cell Expansion Kit (R&D Systems, CDK015)付属のbase mediaとexpander cocktailを使用して、製品
添付文書に記載されている推奨プロトコルに従って、PBMCからNK細胞をexpandしました。このキットは、PBMCから
細胞毒性の高いCD3- CD56+ NK細胞をexpandするのに最適化されており、その後の下流のアプリケーションに使用するこ
とができます。
制御性T細胞への分化誘導
Treg(制御性T細胞)を得るために、まず、MagCellect™ Human Naïve CD4+ T Cell Isolation Kit (R&D Systems, MAGH115)を使
用して、製品添付文書の推奨プロトコルに従って、PBMCからヒトナイーブCD4陽性T細胞をネガティブセレクションで単離
しました。次に、CellXVivo™ Human Treg Cell Differentiation Kit (R&D Systems, CDK006)に含まれている特別に調合された試
薬と成長因子を使用して、FoxP3+ CD25+ Tregへのさらなる分化誘導をしました。得られたTregは下流のアプリケーションに
使用することができます。
PBMCs

4
サンプルの準備とWesとMiloでの測定
Wesの場合、Treg細胞ライセートのFoxP3を検出するために最終濃度0.2mg/mLで、CD25およびCD4を検出するために
0.04mg/mLで調製し、それ以外の各細胞ライセートは0.5mg/mLの最終濃度で調製しました。すべてのサンプルを12-230
kDa Wes Separation Module (ProteinSimple, SM-W003)の推奨プロトコルに従って調製しました。最後に、サンプルを
95°Cで5分間変性、還元してから、Compass for Simple WesternソフトウェアでWesのデフォルトのアッセイ条件と設定
を使用して測定しました。
Miloの場合、各細胞タイプ1 mLをscWestチップに100,000細胞/mLの濃度で播種し、5〜6分間静置しました。DCおよび
NK細胞にはStandard size scWest chip (ProteinSimple, K600)を使用し、TregにはSmall scWest chip (ProteinSimple, K500)
を使用しました。DCおよびNKタイプでは溶解時間を10秒に、Tregでは0秒に設定しました。分析した標的の分子量範囲
に関して、電気泳動実行時間を240 Vで60〜70秒に設定し、240秒間UVでタンパク質を固定しました。
WesおよびMiloで使用した一次抗体および二次抗体の詳細については、このアプリケーションノートの付録およびSimple
WesternおよびSingle-Cell Westernの検証済み抗体データベースを参照してください。
免疫細胞種の同定と特性評価 ナチュラルキラー細胞
ナチュラルキラー(NK)細胞は、癌細胞を標的とするのに抗原特異性に依存しません。 そのため、その細胞傷害性を高め
ることでさまざまな固形腫瘍を消滅させる、格好の自然免疫ベースの治療戦略として知られつつあります5。それでも、
固形腫瘍細胞は、免疫監視K(immune surveillance)から逃れることに長けており、NK細胞機能から逃れた場合、急速に拡
散します。NK細胞の浸潤と腫瘍における機能の両方を監視して、抗腫瘍効果を促進する可能性について有意義な結論を
出す必要があります。図2では、腫瘍におけるNK細胞浸潤と機能をモニタリングすることでWesとMiloの相乗効果を示し
ています。 Wesを使用すると、主要なNK細胞マーカーであるCD56発現量の明確な違いにより、NK細胞のサンプルと
PBMCサンプルを区別することができました(図2A)6。この結果は、PBMCサンプルと比較したNK細胞サンプルでのCD3
発現の欠如によってもさらに裏付けられました(図2B)6。
Miloで同じサンプルをシングルセルレベルでより詳細に解析すると、集団レベルで測定したNK細胞サンプルの
CD56+/CD3-のうち約22%は、CD56+/CD3-NK細胞であったことが明らかになりました(図2C)。興味深いことに、集団
レベルで測定したCD3の発現量は無視できるレベルであるにもかかわらず、そのサンプル内の〜19%のNK細胞サンプル
はCD3+であることがわかりました。これは、細胞集団レベルでは見落す可能性のある少ない細胞サブセットを同定する
のに、シングルセルレベルで測定することは優れた方法であることを示しています。PBMCサンプルでは、細胞の2%が
CD56+細胞として観察され、1.1%がCD56+/CD3+、0.9%がCD56+/CD3-でした。PBMCの0.9%からNK細胞サンプルの
〜22%へのこのCD56+/CD3- NK細胞のシフトは、NK細胞の96%(=(22%-0.9%)/22%)のエンリッチメント(濃縮)を示して
おり、フローサイトメトリーで観察した88%に相当します (データは示していません)。これは、集団レベルで観察した
CD56発現上昇を支持し、WesとMiloが提供する情報の相補性を強調しています。さらに、WesのPBMC集団レベルで測
定した低いCD56の発現と高いCD3の発現細胞は、そのサンプル細胞集団の約19%しか占めていませんでした; PBMCサ
ンプルの〜81%はCD56-およびCD3-であり、他の多くの細胞サブタイプが存在することを示唆しています(図2D)。総細
胞数を計算し、それらの割合を求めるために、Histone H3をローディングコントロールとして使用しました。望んでいた発
現パターンをしている細胞の割合の概要として、図2EにMiloで同定したNK細胞およびPBMCの割合をグラフで表してい
ます。これらのデータは混合集団の細胞サンプルを分析する際に、細胞集団(バルク)とシングルセルの測定技術を組
み合わせることの重要性を証明しており、TMEでのNK細胞の機能を測定する際に、他のアッセイの実験計画の指針とす
ることができます。

5
NK細胞 PBMC
CD56 化学
発光
シグ
ナル
A. B.
160,000 160,000
140,000 140,000
120,000 120,000
100,000 100,000
80,000
60,000
40,000
80,000
60,000
40,000
20,000 20,000
0
12 40 66 116 180 230 MW (kDa)
C.
0
12 40 66 116 180 230 MW (kDa)
CD56+/CD3-
22.3%
CD56+/CD3+
10.6%
CD56-/CD3- 58.5%
CD56-/CD3+ 8.6%
E. 25
20
16.8
22.3
PBMC
NK細胞
15
10 8.6
5
0.9
0 CD56-/CD3+ CD56+/CD3-
細胞の表現型
図2. WesとMiloのNK細胞の同定と特性評価。Wesでは、各サンプルの細胞溶解液0.5 mg / mLをロードすると、NK細胞はPBMCと比較
してより強いCD56シグナルを示した(A)。逆にPBMCではCD3を検出したが、NK細胞サンプルでは検出しなかった(B)。Miloは平均測
定プロファイルを実証し、サンプル内に存在するサブポピュレーションを知る上での手掛かりとなります。Miloでは、サンプル分析に
より、NK細胞サンプルの22%はCD56+/CD3-であったことを示しています(C)。一方PBMCの16.8%はCD56-/CD3+であった。興味深
いことに、分析した10,000細胞/mLのNK細胞サンプルおよびPBMCサンプル内で明らかになった最大の細胞サブセットは、CD56-
/CD3-表現型であった(D)。 NK細胞サンプルとPBMCの典型的な表現型は、分析した集団のパーセントとして示している(E)。
PBMCs, peripheral blood mononuclear cells; NK, natural killer cells
CD3 NK細胞 PBMC
105
104
10³
NK細胞
化学
発光
シグ
ナル
CD
56 ピ
ーク面
積
集団
内での
割合
(%)
CD
56 ピ
ーク面
積
CD3 ピーク面積
D. PBMC
CD56+/CD3- 0.9%
CD56+/CD3+ 1.1%
CD56-/CD3- 81.2%
CD56-/CD3+ 16.8%
104
10³
CD3 ピーク面積
CD
56 ピ
ーク面
積

6
樹状細胞
CD14+未処理細胞
CD83
樹状細胞
CD8+ Tリンパ球は、腫瘍関連抗原に依存して腫瘍細胞を排除します。CD8+ Tリンパ球が持続的に細胞傷害性免疫応答
を誘発するには、DCのようなプロフェッショナル抗原提示細胞による抗原のクロスプレゼンテーションLが必要です7。
それでも、TMEでの腫瘍誘発性免疫抑制のメカニズムは、DC機能とこの抗腫瘍応答を誘発する能力を低下させます。
DC機能の修復または強化を検討する戦略は、CD8+ T細胞ベースのがん免疫療法を改善し、T細胞浸潤の増加との相関
を得るために、CD8+ T細胞の正確な同定と特性評価が不可欠になります7。
異なるDCサブセットには、抗原を処理および提示するさまざまな能力があります。ここでは、最もよく研究されてい
るヒト単球由来のDCに発現しているさまざまなマーカーを検証しました8。Wesで分析したマーカーの相対的発現量に
基づいて、成熟したDC集団とPBMCを明確に区別することができました(図3)。具体的には、単球マーカーCD14は
CD14+未処理細胞で高い発現量を示しましたが、DCでは発現を確認できませんでした(図3、左)9。一方、DC接着分子
CD209(図3、中央)と成熟DCマーカーCD83(図3、右)は、DC細胞で高い発現量を示しましたが、CD14+未処理細胞では
発現を確認できませんでした。特に、CD83は、そのグリコシル化状態により異なる分離プロファイル(図3右)を示す非
常に有望なターゲットとして知られつつあります10。
200,000
200,000
200,000
150,000
150,000
150,000
100,000
100,000
100,000
50,000
50,000
50,000
0
12 40 66 116 180 230 MW (kDa)
0
12 40 66 116 180 230
MW (kDa)
0
12 40 66 116 180 230
MW (kDa)
図3. Wesでの単球由来DCのマーカー発現解析。CD14陽性単球をPBMCから濃縮し、成熟DCに分化誘導した。各細胞のサンプルライ
セート0.5 mg / mLをロードし、CD14の非発現とCD83およびCD209の発現を観察することにより、DCのプロファイルを作成した。エ
レクトロフェログラムは、CD14 +単球細胞サンプルコントロールと比較して、DCでCD209(中央)およびCD83(右)が高く発現している
ことを示している。逆に、予想どおり、CD14は単球サンプルで発現していたが、分析した分化DC集団では発現していなかった(左)。
PBMCs, peripheral blood mononuclear cells; DCs, dendritic cells
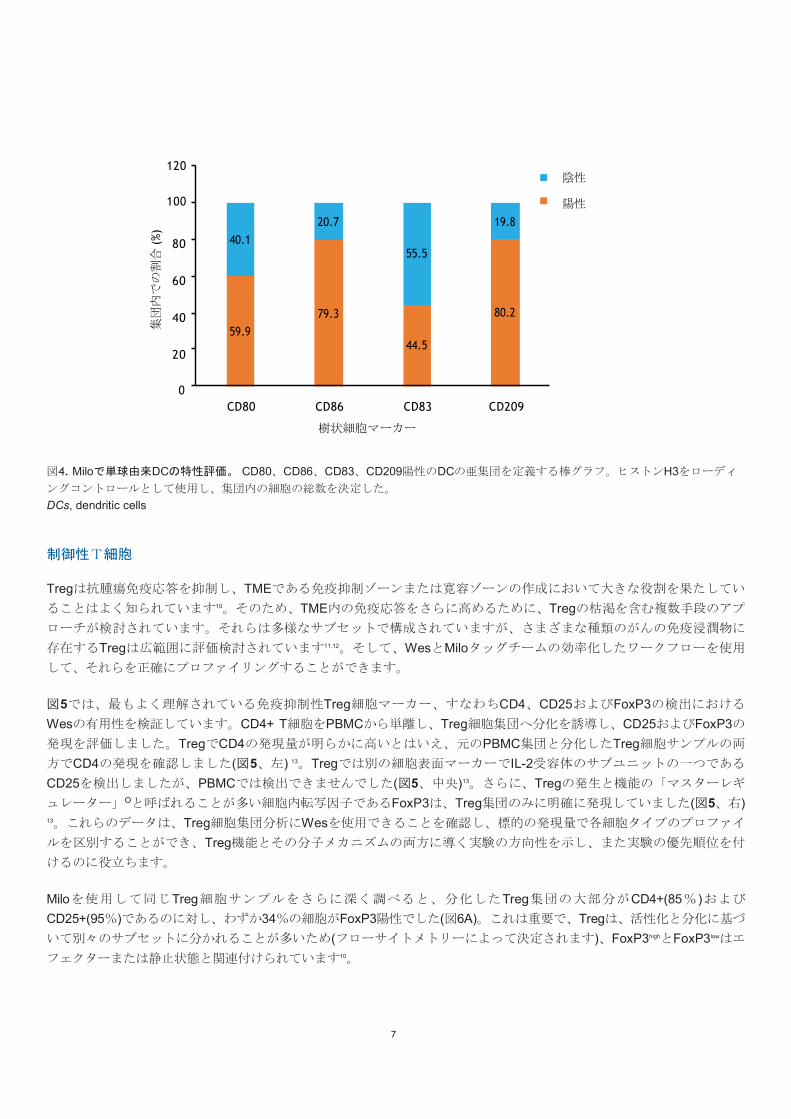
Miloで、同じ単球由来のDCサンプルを使用し免疫表現型をさらに特性評価しました。シングルセル解析により、DCの
60%および79%がそれぞれ共刺激分子MCD80およびCD86を発現していることが明らかになりました。この結果は、この
集団の効率的な抗原提示能力を示しています(図4)。WesでプロファイリングしたDC特異的なCD209およびCD83(図3)は、
各サンプルの約80%および45%の細胞で発現していました。前と同じように、Histone H3をローディングコントロール
として使用して各サンプルの合計細胞数とそれらの割合を計算しました。重要なことに、これらのDCマーカーは従来の
フローサイトメトリー技術を使用しても検出することができますが、Miloではより少ない細胞(〜10,000 : > 100,000)で検
出することができます。これは、TMEから取得したサンプルなど、サンプルに限りがある場合に重要になることがあり
ます。
細胞傷害性T細胞を介した免疫応答を促進する最も強力な抗原提示細胞として、DCがTME内で免疫調節に果たす役割を
理解し、エフェクター機能Nを強化することは、抗腫瘍免疫を高める刺激的な戦略です。WesとMiloを使用すると、腫瘍
内のDCの存在量を決定し、表現型をさらに定義して、存在する細胞集団について正確で機能的な結論を出すことができ
ます。
CD14
樹状細胞
CD14+未処理細胞
樹状細胞
CD14+未処理細胞 CD209
化学発光シグナル
化学発
光シグナル
化学発光
シグナル
7
120
100
陰性
陽性
80
60
40
20
0 CD80 CD86 CD83 CD209
樹状細胞マーカー
図4. Miloで単球由来DCの特性評価。 CD80、CD86、CD83、CD209陽性のDCの亜集団を定義する棒グラフ。ヒストンH3をローディ
ングコントロールとして使用し、集団内の細胞の総数を決定した。
DCs, dendritic cells
制御性T細胞
Tregは抗腫瘍免疫応答を抑制し、TMEである免疫抑制ゾーンまたは寛容ゾーンの作成において大きな役割を果たしてい
ることはよく知られています10。そのため、TME内の免疫応答をさらに高めるために、Tregの枯渇を含む複数手段のアプ
ローチが検討されています。それらは多様なサブセットで構成されていますが、さまざまな種類のがんの免疫浸潤物に
存在するTregは広範囲に評価検討されています11,12。そして、WesとMiloタッグチームの効率化したワークフローを使用
して、それらを正確にプロファイリングすることができます。
図5では、最もよく理解されている免疫抑制性Treg細胞マーカー、すなわちCD4、CD25およびFoxP3の検出における
Wesの有用性を検証しています。CD4+ T細胞をPBMCから単離し、Treg細胞集団へ分化を誘導し、CD25およびFoxP3の
発現を評価しました。TregでCD4の発現量が明らかに高いとはいえ、元のPBMC集団と分化したTreg細胞サンプルの両
方でCD4の発現を確認しました(図5、左) 13。Tregでは別の細胞表面マーカーでIL-2受容体のサブユニットの一つである
CD25を検出しましたが、PBMCでは検出できませんでした(図5、中央)13。さらに、Tregの発生と機能の「マスターレギ
ュレーター」Oと呼ばれることが多い細胞内転写因子であるFoxP3は、Treg集団のみに明確に発現していました(図5、右)
13。これらのデータは、Treg細胞集団分析にWesを使用できることを確認し、標的の発現量で各細胞タイプのプロファイ
ルを区別することができ、Treg機能とその分子メカニズムの両方に導く実験の方向性を示し、また実験の優先順位を付
けるのに役立ちます。
Miloを使用して同じTreg細胞サンプルをさらに深く調べると、分化したTreg集団の大部分がCD4+(85% )および
CD25+(95%)であるのに対し、わずか34%の細胞がFoxP3陽性でした(図6A)。これは重要で、Tregは、活性化と分化に基づ
いて別々のサブセットに分かれることが多いため(フローサイトメトリーによって決定されます)、FoxP3highとFoxP3lowはエ
フェクターまたは静止状態と関連付けられています10。
20.7 19.8
40.1 55.5
79.3 80.2
59.9 44.5
集団内での割合
(%)

8
CD4 CD25 FoxP3
図5. WesでTregマーカー発現解析。CD4+ T細胞をPBMCから濃縮し、Tregに分化誘導し、FoxP3の分析用に各細胞のサンプルライセ
ート0.2mg/mLをロードし、CD25およびCD4の分析用にサンプルライセート0.04mg/mLをロードした。PBMCでCD4の発現を確認した
が、Treg集団の濃縮後の方がその発現量はるかに多かった(左)。PBMCで検出できなかったFoxP3およびCD25マーカーの発現でTreg
を解析評価した(中央、右)。 PBMCs, peripheral blood mononuclear cells
Miloは、検出の難しいFoxP3などの細胞内タンパク質を検出するのに必要なアッセイの柔軟性を提供することにより、フ
ローサイトメトリーのワークフローを簡素化し、細胞の固定や透過処理を必要とせずに表面マーカーと細胞内マーカー
を同時に検出することができます。FoxP3およびCD4発現量をとった散布図(図6B)に示すように、Miloは同じ細胞タイプ
を含むサンプル内の細胞のサブポピュレーションを効果的に発見できます。さらに、Miloの定量測定や、同じ細胞集団
タイプ内の2つの集団を区別する基準を定めることで、Tregに特徴的な不均一性であるFoxP3highとFoxP3lowレベルを確認
することができます(図6C)。Miloがタンパク質の発現レベルの違いで異なる集団を区別する基準(閾値)を定める詳細につ
いては、このアプリケーションノートや利用可能な文献を参照ください14。FoxP3陽性細胞数が少ないため、ここでは分
化したTreg集団は静止状態であると結論付けることができます。今回、Tregの発生源は腫瘍ではなく明らかに健康なド
ナーの末梢血であるため、この結果は合理的かつ適切な判断に基づくように思われます。すなわちこれらのデータは、
TME内での解析にWesとMiloを使用することの合理性を確認しています。

9
A. 120
100
80
60
40
14.9 4.9
85.1
95.1
66.3
陰性
陽性
20 33.7
0 CD4 CD25 FoxP3
Tregマーカー
B. C.
図6. MiloでCD4+ PBMC由来Tregの特性評価。CD4、CD25、およびFoxP33発現に基づきTregサンプル内に存在する細胞亜集団の定量
データを示す棒グラフ(A)。FoxP3とCD4発現の有無に基づき存在するサブポピュレーションの2次元散布図(B)。細胞内FoxP3発現に
基づくTreg集団の分類を示すヒストグラム(C)。
PBMCs, peripheral blood mononuclear cells; Tregs, regulatory T cells
集団内での割合
Cou
nts
Fo
xP3ピ
ーク
面積
11
10
9
8
7
6
5
4
3
2
1
0
FoxP3high
FoxP3low
FoxP3+/CD4- 2.9%
FoxP3+/CD4+ 30.8%
FoxP3-/CD4+ 12.0%
FoxP3-/CD4+ 54.3%
CD4ピーク面積 Log (FoxP3 ピーク面積)

10
まとめ
免疫TME生物学を実際の医療現場で免疫療法に応用するには、さまざまな細胞による複雑さと多様性を解読する
ための高度な技術が必要です。TMEにおけるさまざまな免疫細胞のプロファイリングと特性評価により、転移の
広がりや癌に対する免疫応答を理解し、治療標的の発見および今後の免疫療法戦略への影響を理解することが必要
不可欠です。つまり全ての領域で徹底的な研究が必要不可欠です。このアプリケーションノートでは、R&D
Systemsの細胞分化ソリューションがどのようにWesとMiloのパートナーシップを可能にし、細胞組成や機能を精
細に解析し、前述の目標を達成するのかを実証しました。免疫TMEに取り組む際のアプローチ方法に関係なく、
WesとMiloはきっとあなたを新しい発見に導きます。
参考文献
1. Microenvironmental regulation of therapeutic response in cancer, F Klemm and JA Joyce, Trends Cell Biology, 2015; 25:198–213.
2. HYPE or HOPE: The prognostic value
of infiltrating immune cells in cancer, TA Barnes and E Amir E, British Journal of Cancer, 2017; 117:451–60.
3. Understanding the tumor immune
microenvironment (TIME) for effective therapy, M Binnewies, EW Roberts, K Kersten, V Chan, DF Fearon, M Merad, LM Coussens, DI Gabrilovich, S Ostrand– Rosenberg, CC Hedrick, RH Vonderheide, MJ Pittet, RK Jain, W Zou, TK Howcroft, EC Woodhouse, RA Weinberg and MF Krummel, Nature Medicine, 2018; 24:541–50.
4. Regulatory T cells in the tumor
microenvironment and cancer progression: Role and therapeutic targeting, B Chaudhary B and E Elkord, Vaccines, 2016; 4(3):E28.
5. Overcoming resistance to Natural Killer
cell based immunotherapies for solid tumors, G Nayyar, Y Chu and MS Cairo, Frontiers in Oncology, 2019; 9:51.
6. CD56bright Natural Killer (NK) cells:
An important NK cell subset, A Poli, T Michel, M Thérésine, E Andrès, F Hentges and J Zimmer, Immunology, 2009; 126:458–65.
7. Dendritic cells and CD8 T cell immunity in the tumor microenvironment, F Chunmei and J Aimin, Frontiers in
Immunology, 2018; 9:3059.
8. Antigen cross-presentation by dendritic
cell subsets: one general or all sergeants? S Nierkens, J Tel, E Janssen and GJ Adema, Trends in Immunology, 2013; 34:361–70.
9. Differentiation of human monocytes
into CD14 negative accessory cells: do dendritic cells derive from the monocytic lineage? JH Peters, J Ruppert, RK Gieseler, HM Najar and H Xu, Pathobiology, 1991; 59:122–6.
10. Dendritic cell CD83: A therapeutic
target or innocent bystander? CM Prazma and TF Tedder, Immunology Letters, 2008; 115:1–8.
11. Regulatory T cells in the tumor
microenvironment and cancer progression: Role and therapeutic targeting, B Chaudhary and E Elkord, Vaccines, 2016; 4:E28.
12. Regulatory T cells in cancer
immunotherapy, H Nishikawa and S Sakaguchi, Current Opinion in Immunology, 2014; 27:1–7.
13. Phenotypic alterations, clinical impact and therapeutic potential of regulatory T cells in cancer, B Chaudhary, M Abd Al Samid, BK al-RamadiA and E Elkord, Expert Opinion on Biological Therapy, 2014; 14:931–45.
14. Regulatory T Cells and Foxp3, AY
Rudensky, Immunology Review, 2011; 241:260–68.
15. Profiling human breast epithelial cells
using single cell RNA sequencing identifies cell diversity, QH Nguyen, N Pervolarakis, K Blake, D Ma, RT Davis, N James, AT Phung, E Willey, R Kumar, E Jabart, I Driver, J Rock, A Goga, SA Khan, DA Lawson, Z Werb and K Kessenbrock, Nature Communications, 2018; 9:2028.

付録 表A1. このアプリケーションノートでMiloで使用した抗体。GoatもしくはSheepの一次抗体を使用した場合Milk-Free Antibody Diluentで
希釈したことを除き、一次抗体はAntibody Diluent 2で希釈した。
一次抗体
抗体 メーカー カタログ番号 使用した濃度
Goat Polyclonal Anti-CD4 R&D Systems AF-379-NA 100 µg/mL
Goat Polyclonal Anti-CD25 R&D Systems AF-223-NA 100 µg/mL
Goat Polyclonal Anti-CD56 R&D Systems AF2408 100 µg/mL
Goat Polyclonal Anti-CD80 R&D Systems AF140 100 µg/mL
Goat Polyclonal Anti-CD83 R&D Systems AF2044 100 µg/mL
Goat Polyclonal Anti-CD86 R&D Systems AF-141-NA 100 µg/mL
Sheep Polyclonal Anti-CD209 R&D Systems AF161 100 µg/mL
Goat Polyclonal Anti-FoxP3 R&D Systems BAF3240 100 µg/mL
Rabbit Polyclonal Anti-Histone H3 Novus Biologicals NB500-171 50 µg/mL
Mouse Monoclonal Anti-Histone H3 Novus Biologicals NBP2-36468 50 µg/mL
Rabbit Monoclonal Anti- CD3 ThermoFisher MA5-14524 1:10
用語解説 A腫瘍微小環境: 腫瘍のなかには様々な非腫瘍細胞も多く存在し腫瘍の成長を促しており、これらの細胞や
環境の総称 B浸潤: 本来その組織固有のものでない細胞が,組織中に出現すること C予後予測因子: 治療後の経過を予測し、適切な治療方法を選択するための判断材料 DCD8+ Tリンパ球: リンパ球(T細胞)は、大きくヘルパーT細胞とキラーT細胞に分けられ、キラーT細胞はCD8を発現
しており、ヘルパーT細胞の命令により感染細胞、癌細胞などの異物を認識・破壊するT細胞 Eサブセット: ある全体的なものの一部のこと F末梢血単核細胞: 末梢血から分離した単球(J参照)やリンパ球を含む単核細胞 G樹状細胞: 抗原を自分の細胞表面に掲げ、リンパ球に標的細胞の目印を教える細胞 Hナチュラルキラー細胞: 非抗原特異性で、攻撃の初動部隊としてウイルス感染細胞や、一部のがん細胞を認識して障害す
る免疫細胞 I制御性T細胞: 免疫応答の抑制的制御(免疫寛容)を司るT細胞 J単球: 白血球の一種で樹状細胞等に分化することができる K免疫監視: 侵入してきた細菌などの外敵を退治し、また体内で発生したがん細胞などから身体を守る働きや
仕組みの総称 Lクロスプレゼンテーション: 樹状細胞などが細胞外から取り込んだ外因性抗原を抗原提示分子に結合しCD8+ T細胞に提
示する機能 M共刺激分子: 樹状細胞などに発現誘導される膜タンパク質で、T細胞等の免疫応答へ正と負の調節をする分子 Nエフェクター機能: T細胞が抗原などを認識して活性化し、感染細胞やがん等を排除する機能 Oマスターレギュレーター:多数の下流遺伝子を一括して制御する転写因子
Global bio-techne.com [email protected] TEL +1 612 379 2956 North America TEL 800 343 7475
Europe | Middle East | Africa TEL +44 (0)1235 529449 China [email protected] TEL +86 (21) 52380373 For research use or manufacturing purposes only. Trademarks and registered trademarks are the property of their respective owners.
AN_PWesMilo TME_ASDFY20-9493









![J : ' £ z £ z b J G ^ À H...þ ü £ z £ z b J G ¥ û û û û 1. £ z & K ! J e H K !¹ Q = K¹ . k > Hç* 7 = !¹ Q = K¹ H å ] þ ü ý þ¬ ¤ n µDç ü J ó 2012¬ H ¤](https://static.fdocuments.pl/doc/165x107/5f07ea3f7e708231d41f651d/j-z-z-b-j-g-h-z-z-b-j-g-1-z.jpg)